Kladogramma - Cladogram

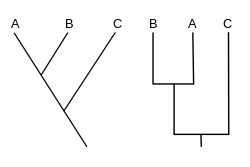
A kladogramma (dan.) Yunoncha kladolar "filial" va grammatika "belgi") - bu ishlatilgan diagramma kladistika organizmlar o'rtasidagi munosabatlarni ko'rsatish. Ammo kladogramma emas evolyutsion daraxt chunki bu ajdodlarning avlodlar bilan qanday aloqasi borligini va ularning qanchalik o'zgarganligini ko'rsatmaydi, shuning uchun ko'plab evolyutsion daraxtlar bir xil kladogramma bilan mos kelishi mumkin.[1][2][3][4][5] Kladogrammada a bilan tugaydigan har xil yo'nalishlarga bo'linadigan chiziqlar ishlatiladi qoplama, a bilan birga bo'lgan organizmlar guruhi so'nggi umumiy ajdod. Kladogrammalarning ko'p shakllari bor, ammo ularning barchasi boshqa chiziqlardan ajralib turadigan chiziqlarga ega. Chiziqlar ularni tarvaqaylab ketgan joyiga qarab kuzatilishi mumkin. Ushbu tarvaqaylab qo'yilgan nuqtalar gipotetik ajdodni anglatadi (haqiqiy shaxs emas), uning ustidagi terminal taksonlari o'rtasida umumiy xususiyatlarni namoyish qilish mumkin.[4][6] Keyinchalik, bu taxminiy ajdod ajdodlar haqida turli xil xususiyatlarning rivojlanishi, moslashuvi va boshqa evolyutsion rivoyatlari to'g'risida maslahatlar berishi mumkin. Garchi an'anaviy ravishda bunday kladogrammalar asosan morfologik belgilar asosida tuzilgan bo'lsa ham, DNK va RNK ma'lumotlar ketma-ketligi va hisoblash filogenetikasi hozirda kladogrammalar avlodida o'z-o'zidan yoki morfologiya bilan birgalikda juda keng qo'llaniladi.
Kladogramma yaratish
Ushbu bo'lim uchun qo'shimcha iqtiboslar kerak tekshirish. (2016 yil aprel) (Ushbu shablon xabarini qanday va qachon olib tashlashni bilib oling) |
Molekulyar va morfologik ma'lumotlar
Kladogramma yaratish uchun ishlatiladigan xususiyatlarni taxminan morfologik (sinapsid bosh suyagi, issiq qonli, notoxord, bir hujayrali va boshqalar) yoki molekulyar (DNK, RNK yoki boshqa genetik ma'lumotlar).[7] DNK sekvensiyasi paydo bo'lishidan oldin, kladistik tahlilda asosan morfologik ma'lumotlar ishlatilgan. Xulq-atvor ma'lumotlari (hayvonlar uchun) ham ishlatilishi mumkin.[8]
Sifatida DNKning ketma-ketligi arzonlashdi va osonlashdi, molekulyar sistematikasi filogenetik gipotezalarni xulosa qilishning tobora ommalashgan usuliga aylandi.[9] Parsimonlik mezonidan foydalanish - bu molekulyar ma'lumotlardan filogeniya xulosa chiqarishning bir necha usullaridan biridir. Kabi yondashuvlar maksimal ehtimollik, ketma-ketlik evolyutsiyasining aniq modellarini o'z ichiga olgan, ketma-ketlik ma'lumotlarini baholashning Hennigiya bo'lmagan usullari. Filogeniyalarni tiklashning yana bir kuchli usuli - bu genomikdan foydalanish retrotranspozon markerlari, muammoga kamroq moyil deb hisoblangan orqaga qaytish ketma-ketlik ma'lumotlarini azoblaydi. Ular, odatda, homoplaziya bilan kasallanish darajasi past deb taxmin qilishadi, chunki ilgari ularning integratsiyasi genom butunlay tasodifiy edi; Biroq, bu hech bo'lmaganda ba'zida bunday bo'lmasligi mumkin.

Plesiomorfiyalar va sinapomorfiyalar
Tadqiqotchilar qaysi belgilar davlatlari "ajdodlar" ekanligiga qaror qilishlari kerak (plesiomorfiyalar ) va ular olingan (sinapomorfiyalar ), chunki faqat sinapomorfik belgilar holatlari guruhlanishni isbotlaydi.[10] Ushbu aniqlash odatda bir yoki bir nechtasining xarakter holatlari bilan taqqoslash yo'li bilan amalga oshiriladi guruhlar. Guruh a'zolari va guruhning ayrim a'zolari o'rtasida birgalikda yashaydigan davlatlar simplesiomorfiyalardir; guruhning faqat bir qismida mavjud bo'lgan holatlar sinapomorfiyalardir. Shuni esda tutingki, bitta terminalga xos bo'lgan belgilar holatlari (autapomorphies) guruhlarga oid dalillarni keltirmaydi. Chiqish guruhini tanlash kladistik tahlilning hal qiluvchi bosqichidir, chunki turli xil guruhlar chuqur topologiyalarga ega daraxtlarni hosil qilishi mumkin.
Gomoplaziyalar
A gomoplaziya biron sababga ko'ra ikki yoki undan ortiq taksonlar bilan bo'lishadigan belgi holatidir boshqa umumiy ajdodlardan ko'ra.[11] Gomoplaziyaning ikkita asosiy turi bu konvergentsiya (kamida ikkita alohida nasl-nasabdagi "bir xil" belgining evolyutsiyasi) va reversiya (ajdodlar xarakteriga qaytish). Gomoplastik xarakterga ega bo'lgan belgilar, masalan, Arktika sutemizuvchilarining turli nasl-nasabidagi oq mo'yna, filogenetik tahlilga belgi sifatida kiritilmasligi kerak, chunki ular o'zaro munosabatlarni tushunishimizga hech qanday hissa qo'shmaydi. Shu bilan birga, gomoplaziya ko'pincha belgining o'zini tekshirishda aniq ko'rinmaydi (masalan, DNK ketma-ketligida bo'lgani kabi) va keyinchalik uning eng parsimon kladogrammada nomuvofiqligi (simsiz tarqalishi) bilan aniqlanadi. Gomoplastik belgilar hali ham filogenetik signalni o'z ichiga olishi mumkinligini unutmang.[12]
Konvergent evolyutsiyasi tufayli homoplaziyaning taniqli namunasi "qanotlarning mavjudligi" xarakteri bo'lishi mumkin. Qushlarning qanotlari bo'lsa ham, ko'rshapalaklar, va hasharotlar bir xil funktsiyani bajaradi, ularning har biri mustaqil ravishda rivojlanib borgan, chunki ular buni ko'rishlari mumkin anatomiya. Agar qush, ko'rshapalak va qanotli hasharotlar "qanotlarning borligi" belgisi uchun to'plangan bo'lsa, ma'lumotlar bazasiga homoplaziya kiritilishi mumkin va bu tahlilni chalkashtirib yuborishi mumkin, bu esa munosabatlarning yolg'on faraziga olib kelishi mumkin. Albatta, birinchi navbatda gomoplazani tanib olishning yagona sababi shundaki, uning gomoplastik taqsimlanishini ochib beradigan munosabatlar modelini nazarda tutadigan boshqa belgilar mavjud.
Kladogramma emas
Kladogramma taksonlarni faqat sinapomorfiyalar asosida guruhlaydigan tahlilning diagramma natijasidir. Ma'lumotlarga biroz boshqacha munosabatda bo'ladigan va natijada kladogrammalarga o'xshash, ammo kladogrammalar bo'lmagan filogenetik daraxtlarni yaratadigan boshqa ko'plab filogenetik algoritmlar mavjud. Masalan, UPGMA va Neighbour-Joining kabi fenetik algoritmlar umumiy o'xshashlik bo'yicha guruhlanadi va ikkala sinapomorfiya va simplesiomorfiyani guruhlashning dalili sifatida qabul qiladi, natijada olingan diagrammalar kladogrammalar emas, fenogrammalardir, Xuddi shunday modelga asoslangan usullarning natijalari (Maksimum Ikkala dallanish tartibini va "novdalar uzunligini" hisobga oladigan "ehtimollik yoki Bayescha yondashuvlar" ikkala sinapomorfiya va avtapomorfiyani ham guruhlash uchun yoki qarshi ko'rsatma sifatida hisoblaydi, bunday tahlil natijalaridan olingan diagrammalar ham kladogrammalar emas.[13]
Kladogramma tanlovi
Bir nechtasi bor algoritmlar "eng yaxshi" kladogrammani aniqlash uchun mavjud.[14] Ko'pgina algoritmlarda a metrik nomzod kladogrammasi ma'lumotlarga qanchalik mos kelishini o'lchash. Ko'pgina kladogramma algoritmlari ning matematik usullaridan foydalaniladi optimallashtirish va minimallashtirish.
Umuman olganda, kladogramma yaratish algoritmlari kompyuter dasturlari sifatida amalga oshirilishi kerak, ammo ba'zi bir algoritmlar ma'lumotlar to'plamlari kam bo'lganida qo'l bilan bajarilishi mumkin (masalan, bir nechta tur va bir nechta xususiyatlar).
Ba'zi algoritmlar faqat xarakteristik ma'lumotlar molekulyar (DNK, RNK) bo'lganda foydali bo'ladi; boshqa algoritmlar faqat xarakterli ma'lumotlar morfologik bo'lganda foydalidir. Xarakterli ma'lumotlar molekulyar va morfologik ma'lumotlarni o'z ichiga olganda boshqa algoritmlardan foydalanish mumkin.
Kladogramma yoki boshqa turdagi filogenetik daraxtlarning algoritmlariga quyidagilar kiradi eng kichik kvadratchalar, qo'shni qo'shilish, parsimonlik, maksimal ehtimollik va Bayes xulosasi.
Biologlar ba'zan bu atamani qo'llashadi parsimonlik kladogramma yaratish algoritmining o'ziga xos turi uchun va ba'zan barcha filogenetik algoritmlar uchun soyabon atamasi sifatida.[15]
Optimallashtirish vazifalarini bajaradigan algoritmlar (masalan, kladogrammalar yaratish) kirish ma'lumotlari (turlar ro'yxati va ularning xususiyatlari) taqdim etilgan tartibda sezgir bo'lishi mumkin. Ma'lumotlarni har xil tartibda kiritish bir xil algoritmda har xil "eng yaxshi" kladogrammalar hosil bo'lishiga olib kelishi mumkin. Bunday vaziyatlarda foydalanuvchi ma'lumotlarni har xil tartibda kiritishi va natijalarni taqqoslashi kerak.
Bitta ma'lumotlar to'plamida turli xil algoritmlardan foydalanish ba'zida har xil "eng yaxshi" kladogrammalarni keltirib chiqarishi mumkin, chunki har bir algoritm "eng yaxshi" ning o'ziga xos ta'rifiga ega bo'lishi mumkin.
Mumkin bo'lgan kladogrammalarning astronomik soni tufayli algoritmlar bu yechim eng yaxshi echim ekanligiga kafolat bera olmaydi. Agar dastur kerakli global minimal darajaga emas, balki mahalliy minimal darajaga o'tsa, untimal bo'lmagan kladogramma tanlanadi.[16] Ushbu muammoni hal qilishda yordam berish uchun ko'plab kladogramma algoritmlari simulyatsiya qilingan tavlanish tanlangan kladogrammaning optimal bo'lishi ehtimolini oshirishga yondashish.[17]
The bazal holat - bu ildiz otgan filogenetik daraxt yoki kladogramma asosining (yoki ildizining) yo'nalishi. Bazal qoplama - bu kattaroq qoplama ichida tarvaqaylab ketish uchun eng erta qoplama (berilgan taksonomik darajaga [a]).
Statistika
Mos kelmaslik uzunligi farqi testi (yoki bo'limning bir xilligi testi)
Mos kelmaslik uzunligi farqi testi (ILD) - bu turli xil ma'lumotlar to'plamlarining (masalan, morfologik va molekulyar, plastid va yadroviy genlarning) kombinatsiyasi uzunroq daraxtga qanday hissa qo'shishini o'lchashdir. Dastlab har bir qismning umumiy daraxt uzunligini hisoblash va ularni yig'ish orqali o'lchanadi. Keyin nusxalar asl bo'limlardan tashkil topgan tasodifiy yig'ilgan qismlarni yasash orqali amalga oshiriladi. Uzunliklar yig'iladi. Agar 99 ta takrorlash uzunroq uzunlikdagi daraxt uzunligiga ega bo'lsa, 100 ta takrorlash uchun 0,01 p qiymati olinadi.
Gomoplazani o'lchash
Ba'zi chora-tadbirlar ma'lumotlar to'plamidagi homoplaziya miqdorini daraxtga qarab o'lchashga harakat qiladi,[18] ammo ushbu choralar qaysi xususiyatni aniqlashga qaratilganligi aniq aniq emas[19]
Mustahkamlik ko'rsatkichi
Tutarlılık indeksi (CI) daraxtning bir qator ma'lumotlarga mosligini o'lchaydi - bu daraxt tomonidan nazarda tutilgan minimal gomoplaziya o'lchovidir.[20] Ma'lumotlar to'plamidagi minimal o'zgarishlarni hisoblash va uni kladogramma uchun zarur bo'lgan o'zgarishlarning haqiqiy soniga bo'lish yo'li bilan hisoblanadi.[20] Muvofiqlik indeksini individual belgi uchun ham hisoblash mumkin men, v bilan belgilanadimen.
Gomoplazi miqdorini aks ettirish bilan bir qatorda metrik ma'lumotlar bazasidagi taksonlar sonini ham aks ettiradi,[21] (kamroq darajada) ma'lumotlar to'plamidagi belgilar soni,[22] har bir belgining filogenetik ma'lumotni ko'tarish darajasi,[23] va qo'shimcha belgilarning kodlanishi, uni maqsadga muvofiq bo'lmagan holatga keltiradigan moda.[24]
vmen 1 dan 1 gacha bo'lgan oraliqni egallaydi / [n.taxa/ 2] teng holat taqsimotiga ega ikkilik belgilarda; uning minimal qiymati holatlar bir tekis tarqalmaganida katta bo'ladi.[23][18] Umuman olganda, bilan ikkilik yoki ikkilik bo'lmagan belgi uchun , vmen dan 1 gacha bo'lgan oraliqni egallaydi .[23]


Saqlash indeksi
Saqlash ko'rsatkichi (RI) "ba'zi bir ilovalar uchun" CI-ni takomillashtirish sifatida taklif qilingan[25] Ushbu o'lchov homoplaziya miqdorini o'lchashga qaratilgan, ammo sinapomorfiyalar daraxtni qanchalik yaxshi tushuntirganligini o'lchaydi. U (daraxtdagi o'zgarishlarning maksimal sonini chiqarib tashlagan holda daraxtdagi maksimal o'zgarishlarning sonini) va (daraxtdagi maksimal o'zgarishlarning sonini va ma'lumotlar to'plamidagi minimal o'zgarishlarni) hisobga olgan holda hisoblanadi.
Qayta tiklangan qat'iylik ko'rsatkichi (RC) CI ni RIga ko'paytirish yo'li bilan olinadi; aslida bu CI diapazonini uzaytiradi, shunda uning minimal nazariy jihatdan erishiladigan qiymati 0 ga ko'tariladi, maksimal darajasi esa 1 ga teng bo'ladi.[18][25] Gomoplazi indeksi (HI) oddiygina 1 - CI.
Gomoplazaning ortiqcha nisbati
Bu daraxtda kuzatilgan gomoplaziya miqdorini nazariy jihatdan mavjud bo'lishi mumkin bo'lgan gomoplazaning maksimal miqdoriga nisbatan o'lchaydi - 1 - (kuzatilgan gomoplazaning ortiqcha) / (maksimal gomoplazaning ortiqcha).[22] 1 qiymati gomoplazaning yo'qligini bildiradi; 0 to'liq tasodifiy ma'lumotlar to'plamida mavjud bo'lgan miqdordagi gomoplazani aks ettiradi va salbiy qiymatlar hanuzgacha ko'proq gomoplazani bildiradi (va faqat uydirma misollarda yuzaga keladi).[22] HER hozirgi vaqtda mavjud bo'lgan eng yaxshi homoplaziya o'lchovi sifatida taqdim etilgan.[18][26]
Shuningdek qarang
Adabiyotlar
- ^ Mayr, Ernst (2009). "Kladistik tahlilmi yoki kladistik tasnifmi?". Zoologik sistematika va evolyutsion tadqiqotlar jurnali. 12: 94–128. doi:10.1111 / j.1439-0469.1974.tb00160.x.
- ^ Foote, Mayk (1996 yil bahor). "Ajdodlarning fotoalbomlarda saqlanish ehtimoli to'g'risida". Paleobiologiya. 22 (2): 141–51. doi:10.1017 / S0094837300016146. JSTOR 2401114.
- ^ Dayrat, Benoit (2005 yil yoz). "Ajdodlar-avlodlar o'rtasidagi munosabatlar va hayot daraxtini tiklash". Paleobiologiya. 31 (3): 347–53. doi:10.1666 / 0094-8373 (2005) 031 [0347: aratro] 2.0.co; 2. JSTOR 4096939.
- ^ a b Posada, Devid; Crandall, Keyt A. (2001). "Turlar ichidagi gen nasablari: Tarmoqlarga payvand qilinadigan daraxtlar". Ekologiya va evolyutsiya tendentsiyalari. 16: 37–45. doi:10.1016 / S0169-5347 (00) 02026-7.
- ^ Podani, Xanos (2013). "Daraxtlarni o'ylash, vaqt va topologiya: evolyutsion / filogenetik sistematikada daraxt diagrammalarini izohlash bo'yicha sharhlar" (PDF). Kladistika. 29 (3): 315–327. doi:10.1111 / j.1096-0031.2012.00423.x.
- ^ Schuh, Randall T. (2000). Biologik sistematika: asoslari va qo'llanilishi. ISBN 978-0-8014-3675-8.[sahifa kerak ]
- ^ DeSalle, Rob (2002). Molekulyar sistematika va evolyutsiyasi usullari. Birxauzer. ISBN 978-3-7643-6257-7.[sahifa kerak ]
- ^ Wenzel, John W. (1992). "Xulq-atvor homologiyasi va filogeniyasi". Annu. Vahiy Ekol. Syst. 23: 361–381. doi:10.1146 / annurev.es.23.110192.002045.
- ^ Xillis, Devid (1996). Molekulyar sistematika. Sinur. ISBN 978-0-87893-282-5.[sahifa kerak ]
- ^ Xenig, Villi (1966). Filogenetik sistematikasi. Illinoys universiteti matbuoti.
- ^ G'arbiy-Eberxard, Meri Jeyn (2003). Rivojlanishning plastikligi va evolyutsiyasi. Oksford universiteti. Matbuot. pp.353 –376. ISBN 978-0-19-512235-0.
- ^ Kalersjo, Mari; Albert, Viktor A.; Farris, Jeyms S. (1999). "Gomoplazi filogenetik tuzilishini oshiradi". Kladistika. 15: 91–93. doi:10.1111 / j.1096-0031.1999.tb00400.x.
- ^ Brower, Endryu V.Z. (2016). "Kladogramma nima va nima yo'q?". Kladistika. 32 (5): 573–576. doi:10.1111 / kla.12144.
- ^ Kitching, Yan (1998). Kladistika: Parsimonlarni tahlil qilish nazariyasi va amaliyoti. Oksford universiteti matbuoti. ISBN 978-0-19-850138-1.[sahifa kerak ]
- ^ Styuart, Karo-Bet (1993). "Parsimonlikning kuchlari va tuzoqlari". Tabiat. 361 (6413): 603–7. Bibcode:1993 yil 361..603S. doi:10.1038 / 361603a0. PMID 8437621.
- ^ Foley, Piter (1993). Klasistik: sistematikaning amaliy kursi. Oksford universiteti. Matbuot. p.66. ISBN 978-0-19-857766-9.
- ^ Nikson, Kevin C. (1999). "Parsimoniyalik Ratchet, tezkor parsimonlik tahlili uchun yangi usul". Kladistika. 15 (4): 407–414. doi:10.1111 / j.1096-0031.1999.tb00277.x.
- ^ a b v d ko'rib chiqildi Archi, Jeyms V. (1996). "Gomoplaziya choralari". Sandersonda Maykl J.; Xufford, Larri (tahrir). Gomoplaziya. pp.153 –188. doi:10.1016 / B978-012618030-5 / 50008-3. ISBN 9780126180305.
- ^ Chang, Jozef T.; Kim, Junhyong (1996). "Gomoplazani o'lchash: stoxastik ko'rinish". Gomoplaziya. 189–203 betlar. doi:10.1016 / b978-012618030-5 / 50009-5. ISBN 9780126180305.
- ^ a b Klyuge, A. G.; Farris, J. S. (1969). "Miqdoriy filetika va anurans evolyutsiyasi". Tizimli zoologiya. 18 (1): 1–32. doi:10.2307/2412407. JSTOR 2412407.
- ^ Archi, J. V.; Felsenshteyn, J. (1993). "Tasodifiy evolyutsion ma'lumotlar uchun tasodifiy va minimal uzunlikdagi daraxtlar bo'yicha evolyutsion qadamlar soni". Aholining nazariy biologiyasi. 43: 52–79. doi:10.1006 / tpbi.1993.1003.
- ^ a b v Archie, J. W. (1989). "GOMOPLAZIYA KO'RSATIShI KO'RSATIShLARI: FILOGENETIK TIZIMDA GOMOPLAZIYA DARAJALARINI OLISHNING YANGI KO'RSATIShLARI VA MUVOFIQLIK KO'RSATIShI TANGISI". Tizimli zoologiya. 38 (3): 253–269. doi:10.2307/2992286. JSTOR 2992286.
- ^ a b v Xoyal Kutill, Jennifer F.; Breddi, Simon J.; Donoghue, Filip C. J. (2010). "Ko'p bosqichli belgilarda mumkin bo'lgan maksimal qadamlar formulasi: matritsa parametrlarining evolyutsion yaqinlashuv ko'rsatkichlariga ta'sirini ajratish". Kladistika. 26: 98–102. doi:10.1111 / j.1096-0031.2009.00270.x.
- ^ Sanderson, M. J .; Donoghue, M. J. (1989). "Gomoplaziya darajalarining o'zgarishi naqshlari". Evolyutsiya. 43 (8): 1781–1795. doi:10.2307/2409392. JSTOR 2409392.
- ^ a b Farris, J. S. (1989). "Saqlash indeksi va qayta tiklangan barqarorlik indeksi". Kladistika. 5 (4): 417–419. doi:10.1111 / j.1096-0031.1989.tb00573.x.
- ^ Hoyal Cuthill, Jennifer (2015). "Belgilar holati maydoni homoplaziyaning paydo bo'lishi va aniqlanishiga ta'sir qiladi: tartibsiz filogenetik belgilar uchun mos kelmaslik ehtimolini modellashtirish". Nazariy biologiya jurnali. 366: 24–32. doi:10.1016 / j.jtbi.2014.10.033. PMID 25451518.
Tashqi havolalar
 Bilan bog'liq ommaviy axborot vositalari Kladogrammalar Vikimedia Commons-da
Bilan bog'liq ommaviy axborot vositalari Kladogrammalar Vikimedia Commons-da
| Tegishli maydonlar | ||
|---|---|---|
| Asosiy tushunchalar | ||
| Xulosa qilish usullari | ||
| Dolzarb mavzular | ||
| Guruh xususiyatlari | ||
| Guruh turlari | ||
| Nomenklatura | ||
| ||