Genom - Genome
| Qismi bir qator kuni |
| Genetika |
|---|
 |
| Asosiy komponentlar |
| Tarix va mavzular |
| Tadqiqot |
| Shaxsiylashtirilgan tibbiyot |
| Shaxsiylashtirilgan tibbiyot |

Dalalarida molekulyar biologiya va genetika, a genom organizmning genetik materialidir. U quyidagilardan iborat DNK (yoki RNK yilda RNK viruslari ). Genom ikkalasini ham o'z ichiga oladi genlar (the kodlash mintaqalari ) va kodlamaydigan DNK,[1] shu qatorda; shu bilan birga mitoxondrial DNK[2] va xloroplast DNK. Genomni o'rganish deyiladi genomika.
Terminning kelib chiqishi
Atama genom tomonidan 1920 yilda yaratilgan Xans Vinkler,[3] professor botanika da Gamburg universiteti, Germaniya. Oksford lug'atida bu nom so'zlarning aralashmasi ekanligi taxmin qilinadi gen va xromosoma.[4] Biroq, qarang omika batafsilroq muhokama qilish uchun. Bir nechta tegishli - bir kabi so'zlar allaqachon mavjud edi biom va ildizpoyali so'z birikmasini shakllantirish genom muntazam ravishda mos keladi.[5]
Tartiblash va xaritalash
Genom ketma-ketligi - bu to'liq ro'yxat nukleotidlar (DNK genomlari uchun A, C, G va T) xromosomalar shaxs yoki tur. Bir tur ichida nukleotidlarning aksariyati jismoniy shaxslar o'rtasida bir xil, ammo genetik xilma-xillikni tushunish uchun bir nechta shaxslarni tartiblashtirish zarur.

1976 yilda, Valter Feyers da Gent universiteti (Belgiya) birinchi bo'lib virusli RNK-genomning to'liq nukleotidlar ketma-ketligini o'rnatdi (Bakteriyofag MS2 ). Keyingi yil, Fred Sanger birinchi DNK-genom ketma-ketligini yakunladi: B-X174 bosqichi, 5386 tayanch juftligidan.[6] Hayotning uchta sohasi orasida birinchi to'liq genomlar ketma-ketligi 90-yillarning o'rtalarida qisqa vaqt ichida paydo bo'ldi: birinchi navbatda bakterial genom ketma-ketligi Gemofilus grippi, da bir jamoa tomonidan yakunlandi Genomik tadqiqotlar instituti 1995 yilda. Bir necha oy o'tgach, birinchi eukaryotik genom tugallandi, undagi xamirturushning 16 xromosomalari ketma-ketligi Saccharomyces cerevisiae 1980-yillarning o'rtalarida boshlangan Evropa boshchiligidagi sa'y-harakatlar natijasida nashr etilgan. An uchun birinchi genom ketma-ketligi arxeon, Metanococcus jannaschii, 1996 yilda, yana Genomik tadqiqotlar instituti tomonidan yakunlandi.
Yangi texnologiyalarning rivojlanishi genomlar ketma-ketligini keskin arzonlashtirdi va osonlashtirdi va to'liq genom sekanslari soni tez o'sib bormoqda. The AQSh milliy sog'liqni saqlash institutlari genomik ma'lumotlarning bir nechta to'liq ma'lumotlar bazalaridan birini saqlaydi.[7] Minglab yakunlangan genomlarni tartiblashtirish loyihalari qatoriga quyidagilar kiradi guruch, a sichqoncha, o'simlik Arabidopsis talianasi, puffer baliq va bakteriyalar E. coli. 2013 yil dekabrda olimlar birinchi navbatda butunni ketma-ketlik qildilar genom a Neandertal, yo'q bo'lib ketgan turlari odamlar. Genom olingan oyoq suyagi dan topilgan 130000 yoshli neandertalning Sibir g'ori.[8][9]
Kabi yangi ketma-ketlik texnologiyalari massiv parallel ketma-ketlik kashshof sifatida diagnostika vositasi sifatida shaxsiy genomlarni ketma-ketlashtirish istiqbollarini ochdi Manteia Bashoratli tibbiyot. Ushbu maqsad sari katta qadam 2007 yilda yakunlandi to'liq genom ning Jeyms D. Uotson, DNK tuzilishini birgalikda kashf etganlardan biri.[10]
Genom ketma-ketligi genomdagi har bir DNK asosining tartibini sanab o'tgan bo'lsa, genom xaritasi diqqatga sazovor joylarni aniqlaydi. Genom xaritasi genomlar ketma-ketligidan kamroq ma'lumotga ega va genom atrofida harakat qilishda yordam beradi. The Inson genomining loyihasi uchun tashkil etilgan xarita va ga ketma-ketlik The inson genomi. Loyihadagi asosiy qadam bu batafsil genomik xaritani chiqarish edi Jan Vaysenbax va uning jamoasi Genoskop Parijda.[11][12]
Yo'naltiruvchi genom ketma-ketliklar va xaritalar yangilanishda davom etmoqda, xatolarni olib tashlaydi va yuqori allelik murakkablikdagi hududlarni aniqlaydi.[13] Genomik xaritalash narxining pasayishiga yo'l qo'yildi nasabga oid uni xizmat sifatida taqdim etadigan saytlar,[14] genomini topshirishi mumkin bo'lgan darajada olomon manbasi kabi ilmiy harakatlar DNK da Nyu-York Genom markazi,[15] ikkalasining ham misoli o'lchov iqtisodiyoti va of fuqarolik fani.[16]
Virusli genomlar
Virusli genomlar yoki RNK, ham DNKdan iborat bo'lishi mumkin. Ning genomlari RNK viruslari ham bo'lishi mumkin bitta zanjirli RNK yoki ikki zanjirli RNK, va bir yoki bir nechta alohida RNK molekulalarini o'z ichiga olishi mumkin (segmentlar: monopartit yoki ko'p partiyali genom). DNK viruslari bitta yoki ikki qatorli genomlarga ega bo'lishi mumkin. Ko'pgina DNK viruslari genomlari DNKning bitta, chiziqli molekulasidan iborat, ammo ba'zilari aylana shaklidagi DNK molekulasidan iborat.[17]Shuningdek, bitta zanjirli RNK deb nomlangan virusli RNK mavjud: mRNK sintezi uchun andoza vazifasini bajaradi[18] va bitta zanjirli RNK: DNK sintezi uchun shablon bo'lib xizmat qiladi.
Virusli konvert[19] virusli genomlar mezbon hujayraga kirish uchun foydalanadigan membrananing tashqi qatlami. Virusli DNK va RNKning ayrim sinflari virusli konvertdan iborat, ba'zilari esa yo'q.
| Sinf / Oila | Konvertmi? |
|---|---|
| Ikki qatorli DNK | |
| Adenovirus | Yo'q |
| Papillomavirus | Yo'q |
| Polyomavirus | Yo'q |
| Herpesvirus | Ha |
| Poxvirus | Ha |
| Bir qatorli DNK | |
| Parvovirus | Yo'q |
| Ikki qatorli RNK | |
| Reovirus | Yo'q |
| Bir qatorli RNK | |
| Picornavirus | Yo'q |
| Koronavirus | Ha |
| Flavivirus | Ha |
| Togavirus | Ha |
| Bir qatorli RNK: mRNK sintezi uchun andoza vazifasini bajaradi | |
| Filovirus | Ha |
| Orthomyxovirus | Ha |
| Paramyxovirus | Ha |
| Rabdovirus | Ha |
| Bir qatorli RNK: DNK sintezi uchun shablon sifatida xizmat qiladi | |
| Retrovirus | Ha |
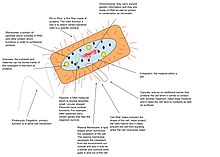
Prokaryotik genomlar
Prokaryotlar va eukariotlar DNK genomiga ega. Arxeya va ko'pgina bakteriyalarda bitta dumaloq xromosoma,[20] ammo, ba'zi bakteriyalar turlari chiziqli yoki ko'p xromosomalarga ega.[21][22] Agar DNK bakteriyalar hujayralari bo'linishidan tezroq takrorlansa, bitta hujayrada xromosomaning bir nechta nusxalari bo'lishi mumkin, agar hujayralar DNKning ko'payishidan tezroq bo'linadigan bo'lsa, bo'linish sodir bo'lishidan oldin xromosomaning ko'p marta ko'payishi boshlanadi, qiz hujayralari to'liq genomlarni va allaqachon qisman takrorlangan xromosomalarni meros qilib olishga imkon beradi. Ko'pgina prokaryotlarning genomida takrorlanadigan DNK juda kam.[23] Biroq, ba'zilari simbiyotik bakteriyalar (masalan, Serratiya simbiotikasi ) genomlari kamaygan va psevdogenlarning yuqori qismi: ularning DNKlarining atigi ~ 40% oqsillarni kodlaydi.[24][25]
Ba'zi bakteriyalar yordamchi genetik materialga, shuningdek, genomning bir qismiga ega plazmidlar. Buning uchun so'z genom ning sinonimi sifatida ishlatilmasligi kerak xromosoma.
Eukaryotik genomlar
Eukaryotik genomlar bir yoki bir nechta chiziqli DNK xromosomalaridan iborat. Xromosomalar soni juda katta farq qiladi Jek sakrash chumolilar va an jinssiz nemotod,[26] ularning har birida faqat bitta juftlik bor, a fern turlari bu 720 juftlikdir.[27] Oddiy hujayrada 22 nusxadan ikkitadan nusxa bor autosomalar, har bir ota-onadan bittadan, ikkitadan jinsiy xromosomalar, uni diploid holga keltiradi. Gametalar, masalan, tuxumdonlar, sperma, spora va gulchanglar gaploid bo'lib, ular har bir xromosomaning faqat bitta nusxasini olib yurishadi.
Yadroda xromosomalardan tashqari, kabi organoidlar xloroplastlar va mitoxondriya o'zlarining DNKlariga ega. Ba'zan mitoxondriyaning o'ziga xos genomi bor, deyishadimitoxondriyal genom ". Xloroplast tarkibidagi DNK" deb nomlanishi mumkinplastome ". Ular paydo bo'lgan bakteriyalar singari, mitoxondriya va xloroplastlar ham aylana xromosomasiga ega.
Prokaryotlardan farqli o'laroq, eukaryotlar oqsillarni kodlovchi genlarning ekzon-intronli tashkil etilishiga va o'zgaruvchan miqdordagi takrorlanadigan DNKga ega. Sutemizuvchilar va o'simliklarda genomning ko'p qismi takrorlanadigan DNKdan iborat.[28]
Kodlash ketma-ketliklari
Oqsillarni ishlab chiqarish bo'yicha ko'rsatmalarga ega bo'lgan DNK sekanslari kodlash ketma-ketligi deb nomlanadi. Kodlash ketma-ketliklari bilan band bo'lgan genomning nisbati juda katta farq qiladi. Kattaroq genomda ko'proq genlar bo'lishi shart emas va takrorlanmaydigan DNK ulushi murakkab eukaryotlarda genom kattaligi ortishi bilan kamayadi.[28]

Kodlashning ketma-ketligi
Kodlashning ketma-ketligi kiradi intronlar, kodlamaydigan RNKlar, tartibga soluvchi mintaqalar va takrorlanadigan DNK uchun ketma-ketliklar. Kodlashsiz ketma-ketliklar inson genomining 98 foizini tashkil qiladi. Genomda takrorlanadigan DNKning ikkita toifasi mavjud: tandem takrorlanadi va intervalgacha takrorlash.[29]
Tandem takrorlaydi
Boshdan quyruqgacha takrorlanadigan qisqa, kodlamaydigan ketma-ketliklar deyiladi tandem takrorlanadi. Mikrosatellitlar 2-5 taglik takroriy takrorlashdan iborat bo'lsa, minisatellit takrorlashlar 30-35 bp. Tandem takrorlanishi inson genomining taxminan 4% va mevali chivin genomining 9% ni tashkil qiladi.[30] Tandemni takrorlash funktsional bo'lishi mumkin. Masalan, telomerlar sutemizuvchilarda takrorlanadigan TTAGGG tandemidan tashkil topgan va ular xromosoma uchlarini himoya qilishda muhim rol o'ynaydi.
Boshqa hollarda, ekzonlar yoki intronlarda tandemning takrorlanish sonining kengayishi sabab bo'lishi mumkin kasallik.[31] Masalan, odamning ovlanadigan geni odatda CAG nukleotidlarining 6-29 tandem takrorlanishini o'z ichiga oladi (poliglutamin traktini kodlash). 36 dan ortiq takroriy natijalar uchun kengayish natijaga olib keladi Xantington kasalligi, neyrodejenerativ kasallik. Yigirmata odamning buzilishi turli xil genlardagi o'xshash tandem takroriy kengayishlaridan kelib chiqishi ma'lum. Kengaytirilgan poligulatamin traktiga ega oqsillarning neyronlarning o'limiga sabab bo'lish mexanizmi to'liq tushunilmagan. Ehtimollardan biri shundaki, oqsillar to'g'ri katlanamaz va degradatsiyani oldini oladi, aksincha muhim transkripsiya omillarini ajratadigan va shu bilan gen ekspressionini o'zgartiradigan agregatlarda to'planadi.[31]
Tandemning takrorlanishiga odatda replikatsiya paytida siljish, tengsiz o'tish va gen konversiyasi sabab bo'ladi.[32]
Transposable elementlar
Transposable elementlar (TE) - bu genomda joylashishini o'zgartirishga qodir bo'lgan aniqlangan tuzilishga ega DNKning ketma-ketliklari.[30][23][33] TElar I-darajali TE-larga bo'linadi, ular nusxa ko'chirish va joylashtirish mexanizmi bilan takrorlanadi yoki genomdan chiqarilib, yangi joyga joylashtirilishi mumkin bo'lgan II TE-lar.
TElarning harakati eukaryotlarda genom evolyutsiyasining harakatlantiruvchi kuchidir, chunki ularning kiritilishi gen funktsiyalarini buzishi mumkin, TElar orasidagi gomologik rekombinatsiya takrorlanishlarni keltirib chiqarishi mumkin, TE esa ekzonlar va tartibga soluvchi ketma-ketlikni yangi joylarga aralashtirib yuborishi mumkin.[34]
Retrotranspozonlar
Retrotranspozonlar[35] asosan eukaryotlarda uchraydi, ammo prokaryotlarda uchramaydi va retrotranspozonlar ko'plab eukaryotlar genomlarining katta qismini tashkil qiladi. Retrotranspozon - bu an orqali o'tadigan transposable element RNK oraliq. Retrotranspozonlar[36] tarkib topgan DNK, ammo transpozitsiya uchun RNKga transkripsiya qilinadi, so'ngra RNK transkripti teskari transkriptaz deb ataladigan ma'lum bir ferment yordamida DNK hosil bo'lishiga ko'chiriladi. O'z genida teskari transkriptazni olib boradigan retrotranspozonlar o'z transpozitsiyasini qo'zg'atishi mumkin, ammo teskari transkriptaz bo'lmagan genlar boshqa retrotranspozon tomonidan sintez qilingan teskari transkriptazdan foydalanishi kerak. Retrotranspozonlar RNKga ko'chirilishi mumkin, keyin ular boshqa joyda genomga ko'paytiriladi.[37] Retrotranspozonlarni ikkiga bo'lish mumkin uzoq terminal takrorlanadi (LTRlar) va uzoq bo'lmagan terminal takrorlashlari (LTR bo'lmaganlar).[34]
Uzoq terminal takrorlashlari (LTR) qadimiy retrovirus infektsiyalaridan kelib chiqadi, shuning uchun ular retrovirus oqsillari bilan bog'liq oqsillarni, shu jumladan gag (virusning strukturaviy oqsillari), pol (teskari transkriptaz va integralaza), pro (proteaz) va ba'zi hollarda env (konvert) genlarini kodlaydi.[33] Ushbu genlarning yonida 5 'va 3' uchlarida ham uzun takrorlanishlar mavjud. Ma'lum qilinishicha, LTRlar o'simliklarning ko'pgina genomlaridagi eng katta fraktsiyadan iborat bo'lib, genom hajmining katta o'zgarishini keltirib chiqarishi mumkin.[38]
Uzoq bo'lmagan terminal takrorlashlari (LTR bo'lmaganlar) sifatida tasniflanadi uzun yadro elementlari (Chiziqlar), intervalgacha bo'lgan qisqa yadro elementlari (SINEs) va Penelopaga o'xshash elementlar (PLEs). Yilda Dictyostelium discoideum, DIRS-ga o'xshash boshqa elementlar LTR bo'lmaganlarga tegishli. LTR bo'lmaganlar eukaryotik genomlarda keng tarqalgan.[39]
Uzoq interpersent elementlar (LINEs) teskari transkriptaz va endonukleaza uchun genlarni kodlaydi va ularni avtonom transposable elementlarga aylantiradi. Odam genomida taxminan 500,000 LINE mavjud, bu genomning 17% ni tashkil qiladi.[40]
Qisqa interfaol elementlar (SINEs) odatda 500 tagacha juftlikdan kam va avtonom emas, shuning uchun ular transpozitsiya uchun LINElar tomonidan kodlangan oqsillarga ishonadilar.[41] The Alu elementi primatlarda uchraydigan eng keng tarqalgan SINE. Bu taxminan 350 ta asosiy juft bo'lib, taxminan 150000 nusxada inson genomining taxminan 11% ni egallaydi.[34]
DNK transpozonlari
DNK transpozonlari teskari terminal takrorlashlari o'rtasida transpozaza fermentini kodlash. Ekspresatsiya qilinganida, transpozaza transpozonning yon tomonidagi terminalning teskari takrorlanishini tan oladi va uning eksizyoni va yangi maydonda qayta joylashishini katalizlaydi.[30] Ushbu kesish va yopishtirish mexanizmi odatda transpozonlarni asl joyiga yaqin joyda (100kb ichida) qayta joylashtiradi.[34] DNK transpozonlari bakteriyalarda uchraydi va inson genomining 3% va yumaloq qurt genomining 12% ni tashkil qiladi. C. elegans.[34]
Genom hajmi
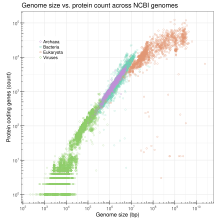
Genom hajmi - bu gaploid genomning bitta nusxasidagi DNK asoslari juftlarining umumiy soni. Genom kattaligi turlar bo'yicha juda xilma-xil. Odamlarda yadro genomiga DNKning taxminan 3,2 milliard nukleotidi kiradi, ularning har biri boshqa xromosomada joylashgan 24 ta chiziqli molekulaga, eng uzuni 50 000 000 nukleotidga va eng uzuni 260 000 000 nukleotidga bo'linadi.[42] Ikkalasida ham morfologik murakkablik va genom kattaligi o'rtasida aniq va izchil bog'liqlik mavjud emas prokaryotlar yoki pastroq eukaryotlar.[28][43] Genom kattaligi asosan takrorlanadigan DNK elementlarining kengayishi va qisqarishiga bog'liq.
Genomlar juda murakkab bo'lganligi sababli, bitta tadqiqot strategiyasi - genomdagi genlar sonini minimal darajaga kamaytirish va shu organizmni omon qolish uchun. Bir hujayrali organizmlar uchun minimal genomlar va ko'p hujayrali organizmlar uchun minimal genomlar bo'yicha eksperimental ishlar olib borilmoqda (qarang. Rivojlanish biologiyasi ). Ish ikkalasi ham jonli ravishda va silikonda.[44][45]
Bu erda ba'zi bir muhim yoki vakillik genomlarining jadvali mavjud. Qarang #Shuningdek qarang ketma-ket genomlar ro'yxati uchun.
| Organizm turi | Organizm | Genom hajmi (tayanch juftliklari ) | Taxminan. yo'q. genlar | Eslatma | |
|---|---|---|---|---|---|
| Virus | Cho'chqa sirkovirusi 1 turi | 1,759 | 1.8kb | Avtonom tarzda takrorlanadigan eng kichik viruslar ökaryotik hujayralar.[46] | |
| Virus | Bakteriyofag MS2 | 3,569 | 3.5kb | Birinchi ketma-ket RNK-genom[47] | |
| Virus | SV40 | 5,224 | 5.2kb | [48] | |
| Virus | B-X174 bosqichi | 5,386 | 5.4kb | Birinchi navbatda DNK-genom[49] | |
| Virus | OIV | 9,749 | 9,7 kg | [50] | |
| Virus | Faj f | 48,502 | 48,5 kg | Ko'pincha rekombinant DNKni klonlash uchun vektor sifatida ishlatiladi. | |
| Virus | Megavirus | 1,259,197 | 1.3Mb | 2013 yilgacha ma'lum bo'lgan eng katta virusli genom.[54] | |
| Virus | Pandoravirus salinusi | 2,470,000 | 2.47Mb | Eng katta ma'lum bo'lgan virusli genom.[55] | |
| Eukaryotik organelle | Inson mitoxondriya | 16,569 | 16,6 kg | [56] | |
| Bakteriya | Nasuia deltocephalinicola (NAS-ALF shtamm) | 112,091 | 112kb | 137 | Virusli bo'lmagan eng kichik genom. Symbiont barg barglari.[57] |
| Bakteriya | Carsonella ruddii | 159,662 | 160kb | An endosimbiont ning psilid hasharotlar | |
| Bakteriya | Buchnera aphidicola | 600,000 | 600kb | Ning endosimbionti shira[58] | |
| Bakteriya | Wigglesworthia glossinidia | 700,000 | 700Kb | Ning ichaklaridagi simbiont tsetse fly | |
| Bakteriya – siyanobakteriya | Proxlorokokk spp. (1,7 Mb) | 1,700,000 | 1.7Mb | 1,884 | Siyanobakteriyalarning eng kichik genomi. Yerdagi asosiy fotosintezatorlardan biri.[59][60] |
| Bakteriya | Gemofilus grippi | 1,830,000 | 1.8Mb | Tirik organizmning birinchi genomi ketma-ketligi, 1995 yil iyul[61] | |
| Bakteriya | Escherichia coli | 4,600,000 | 4.6Mb | 4,288 | [62] |
| Bakteriya - siyanobakteriya | Nostoc punctiforme | 9,000,000 | 9Mb | 7,432 | 7432 ochiq o'qish ramkalari[63] |
| Bakteriya | Solibacter usitatus (Ellin 6076 shtamm) | 9,970,000 | 10Mb | [64] | |
| Ameboid | Polychaos dubiumi ("Amoeba" dubiya) | 670,000,000,000 | 670Gb | Eng katta ma'lum bo'lgan genom.[65] (Bahsli)[66] | |
| O'simlik | Genlisea tuberosa | 61,000,000 | 61Mb | Eng kichigi yozilgan gullarni o'simlik genom, 2014 yil.[67] | |
| O'simlik | Arabidopsis talianasi | 135,000,000[68] | 135 Mb | 27,655[69] | Birinchi o'simlik genomining ketma-ketligi, 2000 yil dekabr.[70] |
| O'simlik | Populus trichocarpa | 480,000,000 | 480Mb | 73,013 | Birinchi daraxt genomining ketma-ketligi, 2006 yil sentyabr[71] |
| O'simlik | Fritillaria assyriaca | 130,000,000,000 | 130Gb | ||
| O'simlik | Parij yaponikasi (Yapon tilida tug'ilgan, rangpar barg) | 150,000,000,000 | 150Gb | Ma'lum bo'lgan eng katta o'simlik genomi[72] | |
| O'simlik – mox | Physcomitrella patenlari | 480,000,000 | 480Mb | A ning birinchi genomi bryofit ketma-ketligi, 2008 yil yanvar.[73] | |
| Qo'ziqorin – xamirturush | Saccharomyces cerevisiae | 12,100,000 | 12,1 Mb | 6,294 | Birinchi ökaryotik genom ketma-ketligi, 1996 yil[74] |
| Qo'ziqorin | Aspergillus nidulans | 30,000,000 | 30Mb | 9,541 | [75] |
| Nematod | Pratylenchus coffeae | 20,000,000 | 20Mb | [76] Ma'lum bo'lgan eng kichik hayvon genomi[77] | |
| Nematod | Caenorhabditis elegans | 100,300,000 | 100Mb | 19,000 | Birinchi ko'p hujayrali hayvon genomi ketma-ketligi, 1998 yil dekabr[78] |
| Hasharot | Drosophila melanogaster (mevali chivin) | 175,000,000 | 175Mb | 13,600 | Kuchlanish asosida o'lchamlarning o'zgarishi (175-180Mb; standart) y w shtamm 175Mb)[79] |
| Hasharot | Apis mellifera (Asalari) | 236,000,000 | 236Mb | 10,157 | [80] |
| Hasharot | Bombyx mori (ipak kuya) | 432,000,000 | 432Mb | 14,623 | 14,623 ta taxmin qilingan genlar[81] |
| Hasharot | Solenopsis invicta (olov chumoli) | 480,000,000 | 480Mb | 16,569 | [82] |
| Sutemizuvchi | Muskul mushak | 2,700,000,000 | 2.7 Gb | 20,210 | [83] |
| Sutemizuvchi | Pan paniskus | 3,286,640,000 | 3.3Gb | 20,000 | Bonobo - taxminiy genom hajmi 3.29 milliard bp[84] |
| Sutemizuvchi | Homo sapiens | 3,000,000,000 | 3Gb | 20,000 | Homo sapiens 2001 yilda 3.2 Gbp ga baholangan genom hajmi[85][86] Inson genomini dastlabki tartiblash va tahlil qilish[87] |
| Qush | Gallus gallus | 1,043,000,000 | 1,0 Gb | 20,000 | [88] |
| Baliq | Tetraodon nigroviridis (puffer baliq turi) | 385,000,000 | 390Mb | 340 Mb deb taxmin qilingan eng kichik umurtqali genom[89][90] - 385 Mb.[91] | |
| Baliq | Protopterus aethiopicus (marmar o'pka baliqlari) | 130,000,000,000 | 130Gb | Ma'lum bo'lgan eng katta umurtqali genom | |
Genomik o'zgarishlar
Organizmning barcha hujayralari bitta hujayradan kelib chiqadi, shuning uchun ularning genomlari bir xil bo'lishi kutilmoqda; ammo, ba'zi hollarda farqlar paydo bo'ladi. Hujayraning bo'linishi paytida DNKni nusxalash jarayoni ham, atrofdagi mutagenlarga ta'sir qilish ham somatik hujayralarda mutatsiyalarga olib kelishi mumkin. Ba'zi hollarda, bunday mutatsiyalar saratonga olib keladi, chunki ular hujayralarni tezroq bo'linishiga va atrofdagi to'qimalarga kirib borishiga olib keladi.[92] Inson immunitet tizimidagi ba'zi limfotsitlarda, V (D) J rekombinatsiyasi har xil hujayra noyob antikor yoki T hujayra retseptorlarini ishlab chiqaradigan har xil genomik ketma-ketlikni hosil qiladi.
Davomida mayoz, diploid hujayralar ikki marta bo'linib, gaploid jinsiy hujayralarni hosil qiladi. Ushbu jarayon davomida rekombinatsiya natijasida gomologik xromosomalardan genetik material o'zgaradi, shuning uchun har bir jinsiy hujayralar o'ziga xos genomga ega bo'ladi.
Genom bo'yicha qayta dasturlash
Sichqoncha bo'yicha genom bo'yicha qayta dasturlash ibtidoiy jinsiy hujayralar o'z ichiga oladi epigenetik izlarni o'chirishga olib keladi totipotensiya. Qayta dasturlash faol tomonidan osonlashtiriladi DNK demetilatsiyasi, DNKni o'z ichiga olgan jarayon asosiy eksizyonni ta'mirlash yo'l.[93] Ushbu yo'l o'chirishda ishlatiladi CpG metilatsiyasi (5mC) ibtidoiy jinsiy hujayralarda. 5mC o'chirish uning konvertatsiya qilish orqali sodir bo'ladi 5-gidroksimetilsitozin (5hmC) yuqori darajadagi o'n o'n bir dioksigenaza fermentlari tomonidan boshqariladi TET1 va TET2.[94]
Genom evolyutsiyasi
Genomlar organizmning yig'indisidan ko'proq genlar va bo'lishi mumkin bo'lgan xususiyatlarga ega o'lchangan va ma'lum bir genlar va ularning mahsulotlarining tafsilotlariga murojaat qilmasdan o'rganilgan. Tadqiqotchilar kabi xususiyatlarni taqqoslashadi karyotip (xromosoma raqami), genom hajmi, genlar tartibi, kodondan foydalanish tarafkashligi va GK-tarkib bugungi kunda mavjud bo'lgan turli xil genomlarni qanday mexanizmlar yaratishi mumkinligini aniqlash uchun (so'nggi sharhlar uchun qarang: Braun 2002; Sakkone va Pesol 2003; Benfey va Protopapas 2004; Gibson va Muse 2004; Riz 2004; Gregori 2005).
Dublikatlar genomni shakllantirishda katta rol o'ynaydi. Duplikatsiya kengaytmasidan boshlab bo'lishi mumkin qisqa tandem takrorlanadi, genlar klasterining takrorlanishiga va butun xromosomalarning takrorlanishiga yoki hatto butun genomlar. Bunday takrorlanishlar, ehtimol, genetik yangilikni yaratish uchun asosdir.
Genlarni gorizontal ravishda uzatish Ikki organizm genomining kichik qismlari o'rtasida juda uzoq o'xshashlik borligini tushuntirish uchun chaqiriladi. Genlarning gorizontal ravishda o'tkazilishi ko'pchilik orasida keng tarqalgan ko'rinadi mikroblar. Shuningdek, eukaryotik hujayralar ularning ba'zi bir genetik materiallarini uzatishni boshdan kechirgan ko'rinadi xloroplast va mitoxondrial ularning yadro xromosomalariga genomlar. So'nggi empirik ma'lumotlar, genetik yangilik va tabiiy genomni tahrirlash uchun asosiy harakatlantiruvchi rolni namoyish etadigan viruslar va sub-virusli RNK-tarmoqlarning muhim rolini ko'rsatadi.
Badiiy adabiyotda
Ilmiy fantastika asarlari genomlar ketma-ketligi mavjudligidan xavotirlarni tasvirlaydi.
Maykl Krixtonning 1990 yilgi romani Yura parki va keyingi film olis orolda klonlangan dinozavrlar parkini yaratadigan milliarder haqida hikoya qiling, natijasi halokatli. Genetika mutaxassisi qadimgi chivinlar qonidan dinozavr DNKini ajratib oladi va bo'shliqlarni zamonaviy turlardan DNK bilan to'ldirib, bir nechta dinozavr turlarini yaratadi. Xaos nazariyachisidan dinozavrlar bilan ekotizimni yaratish xavfsizligi to'g'risida ekspert xulosasini berishni so'rashadi va u bir necha bor loyihaning natijalarini oldindan aytib bo'lmaydigan va oxir-oqibat nazoratsiz bo'lishini ogohlantiradi. Genomik ma'lumotlardan foydalanish xavfi to'g'risida ushbu ogohlantirishlar kitobning asosiy mavzusidir.
1997 yilgi film Gattaka bolalarning genomlari ota-onalari xususiyatlarining eng ideal kombinatsiyasini o'z ichiga oladigan futuristik jamiyatda o'rnatiladi va yurak xastaligi xavfi va umr ko'rish davomiyligi kabi ko'rsatkichlar har bir inson uchun ularning genomiga qarab hujjatlashtiriladi. "In-Valids" deb nomlanuvchi evgenika dasturidan tashqarida homilador bo'lgan odamlar kamsitilishlarga duch kelmoqdalar va yomon kasblarga jo'natildilar. Filmning bosh qahramoni - taxmin qilingan genetik ehtimollarni bartaraf etish va kosmik navigator sifatida ishlash orzusiga erishish uchun ishlaydigan In-Valid. Film kelajakda genomik ma'lumot xurujni kuchaytirishi va genetik jihatdan ishlab chiqilgan bolalarni sotib olishga qodir bo'lmagan va qila olmaydiganlar o'rtasidagi haddan tashqari sinfiy farqlarni keltirib chiqaradi.[95]
Shuningdek qarang
- Bakteriyalarning genom hajmi
- Hayvonlarning genetik resurslarini kriyokonservatsiya qilish
- Genom brauzeri
- Genom kompilyatori
- Genom topologiyasi
- Genom bo'yicha assotsiatsiyani o'rganish
- Hayvonlarning ketma-ket genomlari ro'yxati
- Arxeologik genomlar ketma-ketligi
- Tizimli bakterial genomlar ro'yxati
- Tartibga qo'yilgan ökaryotik genomlar ro'yxati
- Qatorli zamburug'lar genomlari ro'yxati
- Tartiblangan o'simlik genomlari ro'yxati
- Tartiblangan plastomalar ro'yxati
- Tartiblangan protist genomlari ro'yxati
- Metagenomika
- Mikrobiom
- Molekulyar epidemiologiya
- Molekulyar patologik epidemiologiya
- Molekulyar patologiya
- Nuklein kislota ketma-ketligi
- Pan-genom
- Aniq tibbiyot
- Regulyator geni
- Keyingi
- Butun genomlar ketma-ketligi
Adabiyotlar
- ^ Brosius, J (2009), "Parchalangan gen", Nyu-York Fanlar akademiyasining yilnomalari, 1178 (1): 186–93, Bibcode:2009NYASA1178..186B, doi:10.1111 / j.1749-6632.2009.05004.x, PMID 19845638, S2CID 8279434
- ^ Ridli M (2006). Genom: 23 bobda turning tarjimai holi (PDF). Nyu-York: Harper ko'p yillik. ISBN 978-0-06-019497-0. Arxivlandi asl nusxasi (PDF) 2018 yil 24 oktyabrda. Olingan 11 may 2016.
- ^ Vinkler HL (1920). Verbreitung und Ursache der Parthenogenesis im Pflanzen- und Tierreiche. Jena: Verlag Fischer.
- ^ "Genomning ta'rifi Oksford lug'atida". Olingan 25 mart 2014.
- ^ Ledbergberg J, Makkrey AT (2001). "'Ome Sweet 'Omics - so'zlarning nasabiy xazinasi " (PDF). Olim. 15 (7). Arxivlandi asl nusxasi (PDF) 2006 yil 29 sentyabrda.
- ^ "Genlar haqida hamma narsa". www.beowulf.org.uk.
- ^ "Genom uyi". 8 dekabr 2010 yil. Olingan 27 yanvar 2011.
- ^ Zimmer C (2013 yil 18-dekabr). "Oyoq barmoqlarining qoldiqlari to'liq neandertal genomini beradi". The New York Times. Olingan 18 dekabr 2013.
- ^ Prüfer K, Racimo F, Patterson N, Jey F, Sankararaman S, Soyer S va boshq. (2014 yil yanvar). "Oltoy tog'laridan kelgan neandertalning to'liq genom ketma-ketligi". Tabiat. 505 (7481): 43–49. Bibcode:2014 yil Natur.505 ... 43P. doi:10.1038 / tabiat12886. PMC 4031459. PMID 24352235.
- ^ Wade N (2007 yil 31-may). "DNK kashshofi genomi dehifrlangan". The New York Times. Olingan 2 aprel 2010.
- ^ "Genom nima?". Genomenewsnetwork.org. 2003 yil 15-yanvar. Olingan 27 yanvar 2011.
- ^ NCBI_user_services (2004 yil 29 mart). "Ma'lumotlar xaritasini xaritalash". Arxivlandi asl nusxasi 2010 yil 19-iyulda. Olingan 27 yanvar 2011.
- ^ Genom ma'lumotnomasi konsortsiumi. "Genomni yig'ish". Olingan 23 avgust 2016.
- ^ Kaplan, Sara (2016 yil 17-aprel). "Sizning 20000 geningiz juda ko'p turli xil xususiyatlarni qanday aniqlaydi? Ular ko'p vazifali". Washington Post. Olingan 27 avgust 2016.
- ^ Hayden, Erika (2015) ni tekshiring. "Olimlar" DNA.LAND "ga millionlab odamlarni jalb qilishlariga umid qilishadi'". Tabiat. doi:10.1038 / tabiat.2015.18514.
- ^ Zimmer, Karl. "Genomlar o'yini, 13-qism: Javoblar va savollar". STAT. Olingan 27 avgust 2016.
- ^ Gelderblom, Xans R. (1996). Tibbiy mikrobiologiya (4-nashr). Galveston, TX: Texas universiteti tibbiyot filiali Galveston.
- ^ Urry, Lisa A. (2016). Kempbell biologiyasi: ettinchi nashr. Nyu-York: Xoboken: Pirson oliy ma'lumoti. 403-404 betlar. ISBN 0134093410.
- ^ Urry, Lisa A (2016). Kempbell biologiyasi: ettinchi nashr. Nyu-York: Xoboken: Pirson oliy ma'lumoti. 403-404 betlar. ISBN 0134093410.
- ^ Samson RY, Bell SD (2014). "Arxeoal xromosoma biologiyasi". Molekulyar mikrobiologiya va biotexnologiya jurnali. 24 (5–6): 420–27. doi:10.1159/000368854. PMC 5175462. PMID 25732343.
- ^ Chakonas G, Chen CW (2005). "Chiziqli bakterial xromosomalarning ko'payishi: endi aylana bo'ylab aylanish kerak emas". Bakterial xromosoma: 525–540. doi:10.1128 / 9781555817640.ch29. ISBN 9781555812324.
- ^ "Bakterial xromosomalar". Mikrobial genetika. 2002.
- ^ a b Koonin EV, Wolf YI (2010 yil iyul). "Genom va molekulyar-fenom evolyutsiyasidagi cheklovlar va plastiklik". Tabiat sharhlari. Genetika. 11 (7): 487–98. doi:10.1038 / nrg2810. PMC 3273317. PMID 20548290.
- ^ McCutcheon JP, Moran NA (2011 yil noyabr). "Simbiotik bakteriyalarning haddan tashqari genom kamayishi". Tabiat sharhlari. Mikrobiologiya. 10 (1): 13–26. doi:10.1038 / nrmicro2670. PMID 22064560. S2CID 7175976.
- ^ Land M, Hauser L, Jun SR, Nookaew I, Leuze MR, Ahn TH, Karpinets T, Lund O, Kora G, Wassenaar T, Poudel S, Ussery DW (mart 2015). "Bakterial genomning 20 yillik sekvensiyasi haqidagi tushunchalar". Funktsional va integral genomika. 15 (2): 141–61. doi:10.1007 / s10142-015-0433-4. PMC 4361730. PMID 25722247.
- ^ "Olimlar 18 million yillik nasabga ega bo'lgan aseksual mayda chuvalchangni ketma-ketligi". ScienceDaily. Olingan 7-noyabr 2017.
- ^ Xandelval S (1990 yil mart). "Ophioglossum L. turkumidagi xromosoma evolyutsiyasi". Linnean Jamiyatining Botanika jurnali. 102 (3): 205–17. doi:10.1111 / j.1095-8339.1990.tb01876.x.
- ^ a b v Lewin B (2004). Genlar VIII (8-nashr). Yuqori Saddle River, NJ: Pearson / Prentice Hall. ISBN 978-0-13-143981-8.
- ^ Stojanovich N, tahrir. (2007). Hisoblash genomikasi: hozirgi usullar. Wymondham: Horizon Bioscience. ISBN 978-1-904933-30-4.
- ^ a b v Padeken J, Zeller P, Gasser SM (aprel 2015). "Genomni tashkil qilish va barqarorlikda DNKni takrorlang". Genetika va rivojlanish sohasidagi dolzarb fikrlar. 31: 12–19. doi:10.1016 / j.gde.2015.03.009. PMID 25917896.
- ^ a b Usdin K (2008 yil iyul). "Oddiy tandem takrorlanishining biologik ta'siri: takroriy kengayish kasalliklaridan saboq". Genom tadqiqotlari. 18 (7): 1011–19. doi:10.1101 / gr.070409.107. PMC 3960014. PMID 18593815.
- ^ Li YC, Korol AB, Fahima T, Beiles A, Nevo E (dekabr 2002). "Mikrosatellitlar: genomik tarqalish, taxminiy funktsiyalar va mutatsion mexanizmlar: sharh". Molekulyar ekologiya. 11 (12): 2453–65. doi:10.1046 / j.1365-294X.2002.01643.x. PMID 12453231. S2CID 23606208.
- ^ a b Wessler SR (2006 yil noyabr). "Transposable elementlar va eukaryotik genomlarning rivojlanishi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 103 (47): 17600–01. Bibcode:2006 yil PNAS..10317600W. doi:10.1073 / pnas.0607612103. PMC 1693792. PMID 17101965.
- ^ a b v d e Kazazian HH (2004 yil mart). "Mobil elementlar: genom evolyutsiyasi drayverlari". Ilm-fan. 303 (5664): 1626–32. Bibcode:2004 yil ... 303.1626K. doi:10.1126 / science.1089670. PMID 15016989. S2CID 1956932.
- ^ "Transposon | genetika". Britannica entsiklopediyasi. Olingan 5 dekabr 2020.
- ^ Sanders, Mark Frederik (2019). Genetik tahlil: integral yondashuv uchinchi nashr. Nyu-York: Pearson, doimo o'rganadi va o'zlashtiradi. p. 425. ISBN 9780134605173.
- ^ Deininger PL, Moran QK, Batzer MA, Kazazian HH (2003 yil dekabr). "Ko'chma elementlar va sutemizuvchilar genomining evolyutsiyasi". Genetika va rivojlanish sohasidagi dolzarb fikrlar. 13 (6): 651–58. doi:10.1016 / j.gde.2003.10.013. PMID 14638329.
- ^ Kidwell MG, Lisch DR (mart 2000). "Transposable elementlar va mezbon genom evolyutsiyasi". Ekologiya va evolyutsiya tendentsiyalari. 15 (3): 95–99. doi:10.1016 / S0169-5347 (99) 01817-0. PMID 10675923.
- ^ Richard GF, Kerrest A, Dujon B (dekabr 2008). "Eukaryotlarda takrorlanadigan DNKning qiyosiy genomikasi va molekulyar dinamikasi". Mikrobiologiya va molekulyar biologiya sharhlari. 72 (4): 686–727. doi:10.1128 / MMBR.00011-08. PMC 2593564. PMID 19052325.
- ^ Cordaux R, Batzer MA (oktyabr 2009). "Retrotranspozonlarning inson genomi evolyutsiyasiga ta'siri". Tabiat sharhlari. Genetika. 10 (10): 691–703. doi:10.1038 / nrg2640. PMC 2884099. PMID 19763152.
- ^ Xan JS, Boeke JD (2005 yil avgust). "LINE-1 retrotranspozonlari: sutemizuvchilar genlari ekspressioni miqdori va sifatining modulyatorlari?". BioEssays. 27 (8): 775–84. doi:10.1002 / bies.20257. PMID 16015595. S2CID 26424042.
- ^ "Inson genomi". Olingan 19 avgust 2016.
- ^ Gregori TR, Nikol JA, Tamm H, Kullman B, Kullman K, Leitch IJ, Murray BG, Kapraun DF, Greilhuber J, Bennet MD (yanvar 2007). "Eukaryotik genom hajmi ma'lumotlar bazalari". Nuklein kislotalarni tadqiq qilish. 35 (Ma'lumotlar bazasi muammosi): D332-38. doi:10.1093 / nar / gkl828. PMC 1669731. PMID 17090588.
- ^ Glass JI, Assad-Garcia N, Alperovich N, Yooseph S, Lyuis MR, Maruf M, Hutchison CA, Smith HO, Venter JC (yanvar 2006). "Minimal bakteriyaning muhim genlari". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 103 (2): 425–30. Bibcode:2006 yil PNAS..103..425G. doi:10.1073 / pnas.0510013103. PMC 1324956. PMID 16407165.
- ^ Forster AC, Cherkov GM (2006). "Minimal hujayraning sintezi tomon". Molekulyar tizimlar biologiyasi. 2 (1): 45. doi:10.1038 / msb4100090. PMC 1681520. PMID 16924266.
- ^ Mankertz P (2008). "Cho'chqa sirkoviruslarining molekulyar biologiyasi". Hayvonlarning viruslari: Molekulyar biologiya. Caister Academic Press. ISBN 978-1-904455-22-6.
- ^ Fiers W, Contreras R, Duerinck F, Haegeman G, Izerentant D, Merregaert J, Min Jou V, Molemans F, Raeymaekers A, Van den Berghe A, Volckaert G, Ysebaert M (aprel 1976). "MS2 RNK bakteriyofagining to'liq nukleotidlar ketma-ketligi: replikaza genining birlamchi va ikkilamchi tuzilishi". Tabiat. 260 (5551): 500–07. Bibcode:1976 yil natur.260..500F. doi:10.1038 / 260500a0. PMID 1264203. S2CID 4289674.
- ^ Fiers W, Contreras R, Haegemann G, Rogiers R, Van de Voorde A, Van Heuverswyn H, Van Herreweghe J, Volckaert G, Ysebaert M (may 1978). "SV40 DNKning to'liq nukleotidlar ketma-ketligi". Tabiat. 273 (5658): 113–20. Bibcode:1978 yil Noyabr.273..113F. doi:10.1038 / 273113a0. PMID 205802. S2CID 1634424.
- ^ Sanger F, Air GM, Barrell BG, Brown NL, Coulson AR, Fiddes CA, Hutchison CA, Slocombe PM, Smith M (Fevral 1977). "Phi X174 DNK bakteriyofagining nukleotidlar ketma-ketligi". Tabiat. 265 (5596): 687–95. Bibcode:1977 yil natur.265..687S. doi:10.1038 / 265687a0. PMID 870828. S2CID 4206886.
- ^ "Virusologiya - inson immunitet tanqisligi virusi va yordami, tuzilishi: OIV genomi va oqsillari". Pathmicro.med.sc.edu. 2010 yil 1-iyul. Olingan 27 yanvar 2011.
- ^ Thomason L, Court DL, Bubunenko M, Costantino N, Wilson H, Datta S, Oppenheim A (aprel 2007). "Rekombinatsiya: gomologik rekombinatsiyadan foydalangan holda bakteriyalarda genetik muhandislik". Molekulyar biologiyaning amaldagi protokollari. 1-bob: 1.16-birlik. doi:10.1002 / 0471142727.mb0116s78. ISBN 978-0-471-14272-0. PMID 18265390. S2CID 490362.
- ^ Sud DL, Oppenheim AB, Adhya SL (yanvar 2007). "Lambda bakteriofagining genetik tarmoqlariga yangi ko'rinish". Bakteriologiya jurnali. 189 (2): 298–304. doi:10.1128 / JB.01215-06. PMC 1797383. PMID 17085553.
- ^ Sanger F, Coulson AR, Hong GF, Hill DF, Petersen GB (dekabr 1982). "Lambda DNK bakteriofagining nukleotidlar ketma-ketligi". Molekulyar biologiya jurnali. 162 (4): 729–73. doi:10.1016/0022-2836(82)90546-0. PMID 6221115.
- ^ Legendre M, Arslan D, Abergel C, Claverie JM (yanvar 2012). "Megavirus genomikasi va hayotning tushunarsiz to'rtinchi domeni". Kommunikativ va integral biologiya. 5 (1): 102–06. doi:10.4161 / cib.18624. PMC 3291303. PMID 22482024.
- ^ Filipp N, Legendre M, Doutre G, Couté Y, Puaro O, Leskot M, Arslan D, Seltzer V, Bertaux L, Bruley C, Garin J, Claverie JM, Abergel C (2013 yil iyul). "Pandoraviruslar: genomlari 2,5 Mb gacha bo'lgan amabo viruslari, parazitik ökaryotlar" (PDF). Ilm-fan. 341 (6143): 281–86. Bibcode:2013 yil ... 341..281P. doi:10.1126 / science.1239181. PMID 23869018. S2CID 16877147.
- ^ Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJ, Staden R, Young IG (aprel 1981). "Inson mitoxondriyal genomining ketma-ketligi va tashkil etilishi". Tabiat. 290 (5806): 457–65. Bibcode:1981 yil Noyabr.290..457A. doi:10.1038 / 290457a0. PMID 7219534. S2CID 4355527.
- ^ Bennet GM, Moran NA (2013 yil 5-avgust). "Kichkina, kichikroq, eng kichigi: Phloem bilan oziqlanadigan hasharotda qadimgi ikkilangan simbiozlarning kelib chiqishi va rivojlanishi". Genom biologiyasi va evolyutsiyasi. 5 (9): 1675–88. doi:10.1093 / gbe / evt118. PMC 3787670. PMID 23918810.
- ^ Shigenobu S, Vatanabe H, Hattori M, Sakaki Y, Ishikava H (sentyabr 2000). "Aphidlarning endocellular bakterial simbiontining genom ketma-ketligi Buchnera sp. APS". Tabiat. 407 (6800): 81–86. Bibcode:2000. Nat.407 ... 81S. doi:10.1038/35024074. PMID 10993077.
- ^ Rocap G, Larimer FW, Lamerdin J, Malfatti S, Chain P, Ahlgren NA va boshq. (2003 yil avgust). "Ikki Proklorokokk ekotipidagi genom divergensiyasi okean niche farqini aks ettiradi". Tabiat. 424 (6952): 1042–47. Bibcode:2003 yil natur.424.1042R. doi:10.1038 / nature01947. PMID 12917642. S2CID 4344597.
- ^ Dufresne A, Salanoubat M, Partenskiy F, Artiguenave F, Axmann IM, Barbe V va boshq. (2003 yil avgust). "Prochlorococcus marinus SS120 siyanobakteriyasining genom ketma-ketligi, deyarli minimal oksifototrofik genom". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 100 (17): 10020–25. Bibcode:2003 PNAS..10010020D. doi:10.1073 / pnas.1733211100. PMC 187748. PMID 12917486.
- ^ Fleischmann RD, Adams MD, White O, Kleyton RA, Kirkness EF, Kerlavage AR, Bult CJ, Tomb JF, Dougherty BA, Merrick JM (1995 yil iyul). "Haemophilus influenzae Rd ning genogenom tasodifiy ketma-ketligi va yig'ilishi". Ilm-fan. 269 (5223): 496–512. Bibcode:1995 yilgi ... 269..496F. doi:10.1126 / science.7542800. PMID 7542800. S2CID 10423613.
- ^ Blattner FR, Plunkett G, Bloch CA, Perna NT, Burland V, Riley M va boshq. (1997 yil sentyabr). "Escherichia coli K-12 ning to'liq genom ketma-ketligi". Ilm-fan. 277 (5331): 1453–62. doi:10.1126 / science.277.5331.1453. PMID 9278503.
- ^ Meeks JC, Elhai J, Thiel T, Potts M, Larimer F, Lamerdin J, Predki P, Atlas R (2001). "Nostoc punctiforme, ko'p hujayrali, simbiyotik siyanobakteriya genomiga umumiy nuqtai". Fotosintez tadqiqotlari. 70 (1): 85–106. doi:10.1023 / A: 1013840025518. PMID 16228364. S2CID 8752382.
- ^ Challacombe JF, Eichorst SA, Hauser L, Land M, Xie G, Kuske CR (15 sentyabr 2011). Shtinke D (tahrir). "Candidatus Solibacter usitatus Ellin6076 yirik genomida qadimgi genlarni olish va takrorlashning biologik oqibatlari". PLOS ONE. 6 (9): e24882. Bibcode:2011PLoSO ... 624882C. doi:10.1371 / journal.pone.0024882. PMC 3174227. PMID 21949776.
- ^ Parfrey LW, Lahr DJ, Katz LA (2008 yil aprel). "Eukaryotik genomlarning dinamik tabiati". Molekulyar biologiya va evolyutsiya. 25 (4): 787–94. doi:10.1093 / molbev / msn032. PMC 2933061. PMID 18258610.
- ^ ScienceShot: Hech qachon eng katta Genom Arxivlandi 2010 yil 11 oktyabr Orqaga qaytish mashinasi, sharhlar: "Juda katta genomga ega ekanligi xabar qilingan Amoeba dubia va boshqa protozoa uchun o'lchov 1960 yillarda qo'pol biokimyoviy yondoshuv yordamida amalga oshirildi, bu hozirgi vaqtda genom hajmini aniq aniqlash uchun ishonchsiz usul hisoblanadi."
- ^ Fleischmann A, Maykl TP, Rivadavia F, Sousa A, Vang V, Temsch EM, Greilxuber J, Myuller KF, Heubl G (dekabr 2014). "Genlisea (Lentibulariaceae) yirtqich o'simliklarning genom kattaligi va xromosoma sonining evolyutsiyasi, angiospermlarda minimal genom hajmini yangi baholash bilan". Botanika yilnomalari. 114 (8): 1651–63. doi:10.1093 / aob / mcu189. PMC 4649684. PMID 25274549.
- ^ "Genom assambleyasi". Arabidopsis Axborot Resursi (TAIR).
- ^ "Tafsilotlar - Arabidopsis thaliana - Ensembl Genomes 40". o'simliklar.ensembl.org.
- ^ Greilhuber J, Borsch T, Myuller K, Worberg A, Porembski S, Barthlott V (noyabr 2006). "Lentibulariaceae-da topilgan eng kichik angiosperm genomlari, bakteriyalar kattaligi xromosomalari". O'simliklar biologiyasi. 8 (6): 770–77. doi:10.1055 / s-2006-924101. PMID 17203433.
- ^ Tuskan GA, Difazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U va boshq. (2006 yil sentyabr). "Populus trichocarpa (Torr. Va Gray)" qora paxta daraxti genomi " (PDF). Ilm-fan. 313 (5793): 1596–604. Bibcode:2006 yil ... 313.1596T. doi:10.1126 / science.1128691. PMID 16973872. S2CID 7717980.
- ^ Pellicer J, Fay MF, Leitch IJ (2010 yil 15 sentyabr). "Barchasining eng katta eukaryotik genomi?". Linnean Jamiyatining Botanika jurnali. 164 (1): 10–15. doi:10.1111 / j.1095-8339.2010.01072.x.
- ^ Lang D, Zimmer AD, Rensing SA, Reski R (oktyabr 2008). "O'simliklar bioxilma-xilligini o'rganish: Physcomitrella genomi va boshqalar". O'simlikshunoslik tendentsiyalari. 13 (10): 542–49. doi:10.1016 / j.tplants.2008.07.002. PMID 18762443.
- ^ "Saccharomyces Genom ma'lumotlar bazasi". Yeastgenome.org. Olingan 27 yanvar 2011.
- ^ Galagan JE, Calvo SE, Cuomo C, Ma LJ, Wortman JR, Batzoglou S va boshq. (2005 yil dekabr). "Aspergillus nidulans ketma-ketligi va A. fumigatus va A. oryzae bilan qiyosiy tahlil". Tabiat. 438 (7071): 1105–15. Bibcode:2005 yil. Tabiat. 438.1105G. doi:10.1038 / nature04341. PMID 16372000.
- ^ Leroy S, Bouamer S, Morand S, Fargette M (2007). "O'simlik-parazit nematodalarning genom kattaligi". Nematologiya. 9 (3): 449–50. doi:10.1163/156854107781352089.
- ^ Gregori TR (2005). "Hayvonlar genomlari hajmi bo'yicha ma'lumotlar bazasi". Gregori, T.R. (2016). Hayvonlar genomlari hajmi bo'yicha ma'lumotlar bazasi.
- ^ The C. elegans Tartiblash konsortsiumi (1998 yil dekabr). "C. elegans nematodasining genom ketma-ketligi: biologiyani o'rganish platformasi". Ilm-fan. 282 (5396): 2012–18. Bibcode:1998 yil ... 282.2012.. doi:10.1126 / science.282.5396.2012. PMID 9851916. S2CID 16873716.
- ^ Ellis LL, Huang Vt, Quinn AM, Ahuja A, Alfrejd B, Gomes FE, Hjelmen Idoralar, Mur KL, Mackay TF, Johnston JS, Tarone AM (iyul 2014). "D. melanogasterdagi intrapopulyatsiya genomining o'zgarishi hayot tarixining o'zgarishi va plastisiyasini aks ettiradi". PLOS Genetika. 10 (7): e1004522. doi:10.1371 / journal.pgen.1004522. PMC 4109859. PMID 25057905.
- ^ Asal ari genomini ketma-ketlashtirish konsortsiumi (2006 yil oktyabr). "Apis mellifera asal asalining genomidan olingan ijtimoiy hasharotlar to'g'risida tushunchalar". Tabiat. 443 (7114): 931–49. Bibcode:2006 yil natur.443..931T. doi:10.1038 / tabiat05260. PMC 2048586. PMID 17073008.
- ^ Xalqaro ipak qurti genomi (2008 yil dekabr). "Lepidopteran modeli hasharotining genomi, ipak qurti Bombyx mori". Hasharotlar biokimyosi va molekulyar biologiya. 38 (12): 1036–45. doi:10.1016 / j.ibmb.2008.11.004. PMID 19121390.
- ^ Wurm Y, Vang J, Riba-Grognuz O, Corona M, Nygaard S, Hunt BG va boshq. (2011 yil aprel). "Solenopsis invicta olovli chumolining genomi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 108 (14): 5679–84. Bibcode:2011PNAS..108.5679W. doi:10.1073 / pnas.1009690108. PMC 3078418. PMID 21282665.
- ^ Cherkov DM, Goodstadt L, Hillier LW, Zody MC, Goldstein S, She X va boshq. (2009 yil may). Roberts RJ (tahrir). "Sichqoncha tugallangan genom majmuasi tomonidan aniqlangan naslga xos biologiya". PLOS biologiyasi. 7 (5): e1000112. doi:10.1371 / journal.pbio.1000112. PMC 2680341. PMID 19468303.
- ^ "Pan paniskus (pigmentli shimpanze)". nih.gov. Olingan 30 iyun 2016.
- ^ Erik Lander; va boshq. (2001 yil 15 fevral). "Inson genomini dastlabki ketma-ketligi va tahlili". Tabiat. 409 (6822): 860–921. doi:10.1038/35057062. PMID 11237011. Jadval 8.
- ^ "Funktsional va qiyosiy Genomika ma'lumotlari". Ornl.gov. Arxivlandi asl nusxasi 2008 yil 20 sentyabrda.
- ^ Venter JC, Adams MD, Myers EW, Li PW, Mural RJ, Satton GG va boshq. (2001 yil fevral). "Inson genomining ketma-ketligi". Ilm-fan. 291 (5507): 1304–51. Bibcode:2001 yil ... 291.1304V. doi:10.1126 / science.1058040. PMID 11181995.
- ^ Xalqaro tovuq genomini tartiblash konsortsiumi (2004 yil dekabr). "Tovuq genomining ketma-ketligi va qiyosiy tahlili umurtqali hayvonlar evolyutsiyasining o'ziga xos istiqbollarini beradi". Tabiat. 432 (7018): 695–716. Bibcode:2004 yil natur.432..695C. doi:10.1038 / nature03154. ISSN 0028-0836. PMID 15592404.
- ^ Roest Crollius H, Jaillon O, Dasilva C, Ozouf-Costaz C, Fizames C, Fischer C, Bouneau L, Billault A, Quetier F, Saurin W, Bernot A, Weissenbach J (iyul 2000). "Tetraodon nigroviridis chuchuk suvli pushti baliqning ixcham genomini tavsiflash va takroriy tahlil qilish". Genom tadqiqotlari. 10 (7): 939–49. doi:10.1101 / gr.10.7.939. PMC 310905. PMID 10899143.
- ^ Jaillon O, Aury JM, Brunet F, Petit JL, Stange-Thomann N, Mauceli E va boshq. (2004 yil oktyabr). "Tetraodon nigroviridis teleost baliqlarida genomning ko'payishi erta umurtqali proto-karyotipni ochib beradi". Tabiat. 431 (7011): 946–57. Bibcode:2004 yil natur.431..946J. doi:10.1038 / tabiat03025. PMID 15496914.
- ^ "Tetraodon loyihasi haqida ma'lumot". Arxivlandi asl nusxasi 2012 yil 26 sentyabrda. Olingan 17 oktyabr 2012.
- ^ Martincorena I, Kempbell PJ (sentyabr 2015). "Saraton va normal hujayralardagi somatik mutatsiya". Ilm-fan. 349 (6255): 1483–89. Bibcode:2015 yil ... 349.1483 million. doi:10.1126 / science.aab4082. PMID 26404825. S2CID 13945473.
- ^ Xajkova P, Jeffri SJ, Li S, Miller N, Jekson SP, Surani MA (iyul 2010). "Sichqoncha urug'i chizig'ida genom bo'yicha qayta dasturlash eksizyonni tiklash yo'lini talab qiladi". Ilm-fan. 329 (5987): 78–82. Bibcode:2010Sci ... 329 ... 78H. doi:10.1126 / science.1187945. PMC 3863715. PMID 20595612.
- ^ Hackett JA, Sengupta R, Zylicz JJ, Murakami K, Lee C, Down TA, Surani MA (yanvar 2013). "Germline DNK demetilatsiyasi dinamikasi va 5-gidroksimetilsitozin orqali izlarni yo'q qilish". Ilm-fan. 339 (6118): 448–52. Bibcode:2013 yil ... 339..448H. doi:10.1126 / science.1229277. PMC 3847602. PMID 23223451.
- ^ "Gattaka (film)". Rotten Pomidor.
Qo'shimcha o'qish
- Benfey P, Protopapas AD (2004). Genomika asoslari. Prentice Hall.
- Jigarrang TA (2002). Genomlar 2. Oksford: Bios Scientific Publishers. ISBN 978-1-85996-029-5.
- Gibson G, Muse SV (2004). Genom fanining asosiy yo'nalishi (Ikkinchi nashr). Sanderlend, Mass: Sinayer dots. ISBN 978-0-87893-234-4.
- Gregori TR (2005). Genomning rivojlanishi. Elsevier. ISBN 978-0-12-301463-4.
- Reece RJ (2004). Genlar va genomlar tahlili. Chichester: John Wiley & Sons. ISBN 978-0-470-84379-6.
- Sakkone C, Pesole G (2003). Qiyosiy genomika bo'yicha qo'llanma. Chichester: John Wiley & Sons. ISBN 978-0-471-39128-9.
- Verner E (2003 yil dekabr). "Ko'p hujayrali silika tizimlarida biologiya va minimal genomlar". Bugungi kunda giyohvand moddalarni kashf etish. 8 (24): 1121–27. doi:10.1016 / S1359-6446 (03) 02918-0. PMID 14678738.
Tashqi havolalar
- UCSC Genome brauzeri - 80 dan ortiq organizmlar uchun genom va izohlarni ko'rish.
- genomecenter.howard.edu
- DNK molekulasini yarating
- Ba'zi taqqoslanadigan genom o'lchamlari
- DNKning interaktiv: DNK fanining tarixi
- Boshidan DNK
- Inson genomi loyihasi haqida hamma narsa - Genome.gov-dan
- Hayvonlarning genom hajmi ma'lumotlar bazasi
- O'simliklar genomlari hajmi ma'lumotlar bazasi
- OLTIN: OnLine ma'lumotlar bazasini yaratadi
- Genom yangiliklar tarmog'i
- NCBI Entrez Genome Project ma'lumotlar bazasi
- NCBI Genom Primer
- Generkartalar - inson genlarining yaxlit ma'lumotlar bazasi
- BBC News - yakuniy genomning "bobi" nashr etildi
- IMG (Integrated Microbial Genomes system) - DOE-JGI tomonidan genomni tahlil qilish uchun
- GeKnome Technologies Keyingi avlod ketma-ketligini tahlil qilish - keyingi avlod uchun ketma-ketlik ma'lumotlarini tahlil qilish Illumina va 454 GeKnome Technologies xizmati.
| Asosiy komponentlar | |
|---|---|
| Maydonlar | |
| Arxeogenetika ning | |
| Tegishli mavzular | |
| Ro'yxatlar | |
| |
| Genomika | |
|---|---|
| Bioinformatika | |
| Strukturaviy biologiya | |
| Tadqiqot vositalari | |
| Tashkilotlar |
|
| |