ESCRT - ESCRT - Wikipedia
The endosomal tashish uchun zarur bo'lgan saralash komplekslari (ESCRT) texnika tashkil topgan sitosolik ESCRT-0, ESCRT-I, ESCRT-II va ESCRT-III deb nomlanuvchi oqsil komplekslari. Bir qator aksessuar oqsillari bilan birgalikda ushbu ESCRT komplekslari noyob rejimini ta'minlaydi membrana qayta qurish natijasida membranalar egilib / tomurcuklanıyor sitoplazma.[1][2] Ushbu ESCRT komponentlari ajratilgan va bir qator organizmlarda o'rganilgan, shu jumladan xamirturush va odamlar.[3]
ESCRT apparati bir qator uyali jarayonlarda, shu jumladan muhim rol o'ynaydi multivesular tanasi (MVB) biogenez, uyali abscission va virusli tomurcuklanma. Multivesular tanadagi (MVB) biogenez bu jarayon hamma joyda belgilangan oqsillar kiradi organoidlar hosil bo'lishi orqali endosomalar deb ataladi pufakchalar. Ushbu jarayon hujayralar noto'g'ri katlanmış va shikastlangan oqsillarni yo'q qilish uchun juda muhimdir.[4] ESCRT uskunalarisiz bu oqsillar ko'payishi va olib kelishi mumkin neyrodejenerativ kasallik. Masalan, ESCRT-III tarkibiy qismlarining anormalliklari kabi nevrologik kasalliklarga olib kelishi mumkin irsiy spastik paraplegiya (HSP).[5] Uyali abscission, ikkita qiz hujayralarni birlashtirgan membranani ajratish jarayoni ham ESCRT apparati vositasida amalga oshiriladi. ESCRT komplekslarisiz, qiz hujayralar ikki baravar ko'proq o'z ichiga olgan g'ayritabiiy hujayralarni ajrata olmadi DNK hosil bo'ladi. Ushbu hujayralar muqobil ravishda ma'lum bo'lgan jarayon orqali yo'q qilinadi apoptoz. Va nihoyat, ESCRT apparati yo'qligida virusli kurtaklanish yoki ma'lum turdagi viruslarning hujayralardan chiqishi jarayoni sodir bo'lmasligi mumkin. Bu muqarrar ravishda viruslarning hujayradan hujayraga tarqalishini oldini oladi.
ESCRT komplekslari va qo'shimcha oqsillar

ESCRT komplekslari va qo'shimcha oqsillarning har biri o'ziga xos biokimyoviy funktsiyalarni ta'minlaydigan noyob tuzilmalarga ega. ESCRT texnikasining har bir oqsil komponenti uchun xamma xamirturush uchun ham bir qator sinonimlar mavjud metazoanlar. Ushbu oqsillarning barchasi haqida qisqacha jadval quyida keltirilgan.
Xamirturushlarda quyidagi komplekslar / aksessuar oqsillari quyidagicha mavjud:
ESCRT-0
ESCRT-0 kompleksi hamma joyda mavjud bo'lgan oqsillarni va / yoki biriktirish va klasterlash orqali ko'p qirrali jismlarni hosil qilishda muhim rol o'ynaydi. retseptorlari hujayra yuzasida Keyinchalik kompleks a bilan bog'lanish uchun javobgardir lipid endosomaga ushbu belgilangan oqsillarni jalb qiladigan endosomal membranada.[6] Bir marta to'g'ri mahalliylashtirilgan, keyinchalik bu oqsillar pufakchalar orqali endosomaga kirib, multivikulyar jismlarni hosil qiladi va oxiriga etkaziladi lizosoma qaerda ular tanazzulga uchragan. Ushbu jarayon juda zarur, chunki u orqali o'tgan zararlangan oqsillarni parchalanishi uchun asosiy yo'ldir Golgi.[4] ESCRT-0 kompleksining tarkibiy qismlari quyidagicha mavjud:
Kompleks 1: 1 heterodimer Vps27 (vakuolyar oqsillarni saralash oqsil 27) va Hse1.[1][5] Vps27 va Hse1 xiralashmoq orqali antiparallel o'ralgan lasan GAT (GGA va Tom1 oqsillari nomi bilan atalgan) domenlari.[1] Vps27 va Hse1 ikkalasida ham an mavjud amino-terminal VHS domeni (shunday nomlangan, chunki u tarkibida mavjud Vps27, HRS va STAM oqsillari).[7] Ushbu VHS domenlari hujayraning parchalanishini maqsad qilgan oqsillar bilan ubiqitinni bog'laydi. Ubiquitin, shuningdek, Hse1-dagi yoki Vps27-da joylashgan ikki tomonlama domen kabi o'zaro ta'sir qiluvchi motiflar bilan bog'lanishi mumkin. A FYVE domeni (dastlab u aniqlangan to'rtta oqsil nomi bilan atalgan: Fab1p, YOTB, Vac1 va EEA1) VPS va Vb27 ning o'zaro ta'sir qiluvchi motiflari domenlari o'rtasida joylashgan.[5][8] Fosfatidilinozitol 3-fosfat, keng tarqalgan endosomal lipid, ushbu FYVE domeniga bog'lanib, endosomaga ESCRT-0 ni jalb qilishga olib keladi.[5]
ESCRT-I
ESCRT-I kompleksining roli hamma joyda mavjud bo'lgan oqsillarni klasterlash va ESCRT-0 va ESCRT-II komplekslari o'rtasida ko'prik vazifasini bajarib, ko'p qirrali jismlarni yaratishda yordam berishdan iborat.[9] Shuningdek, u membrananing tanib olinishi va membrananing abscissiyasi paytida qayta tiklanishida rol o'ynaydi, chunki uning ikkala tomonida halqalar hosil bo'ladi o'rtamiyona bo'linadigan hujayralar. ESCRT-I shuningdek hujayralarni ajratishidan oldin siqilish zonasini tashkil etadigan ESCRT-IIIni jalb qilish uchun javobgardir.[10] Bundan tashqari, ESCRT-I o'ziga xos virusli oqsillar bilan o'zaro ta'sirlashib, virusni kurtaklashda rol o'ynaydi va qo'shimcha ravishda ESCRT mexanizmlarini virusni chiqarilish joyiga jalb qilishga olib keladi.[11] ESCRT-I texnikasi tafsilotlari quyida tavsiflangan.
ESCRT-I kompleksi a heterotetramer (1: 1: 1: 1) ning Vps23, Vps28, Vps37 va Mvb12.[3] O'rnatilgan heterotetramer Vps23, Vps37 va Mvb12 dan tashkil topgan novda shaklidagi sopi bo'lib ko'rinadi. spirallar Vps23, Vps28 va Vps37.[3][5] Vps23 tarkibiga bitta ubikuitin E2 variant domeni kiradi, u ubikuitin, ESCRT-0 kompleksi va PTAP (prolin, txreonin, alanin, prolin) virusli motiv Gag oqsillari.[3][5] Ushbu ubiquitin E2 variant domenidan so'ng prolinga boy motif (GPPX)3Y) mavjud bo'lib, ESCRT-I ni membranani abscissiyasi paytida o'rta tanaga yo'naltiradi.[5] Mvb12 ubiqitinni ham uning yordamida bog'lashi mumkin karboksi-terminal. Vps28 GLUE domeni bilan bog'lanish orqali ESCRT-I va ESCRT-II ning o'zaro ta'siri uchun javobgardir (GRAM-Like Ubiqitin bilan bog'langan EVps36 ning AP45) karboksi-terminali orqali to'rt spiral to'plami domen.[1]
ESCRT-II
ESCRT-II kompleksi birinchi navbatda multivikulyar jismlarning biogenezi va ubikuitin etiketli oqsillarni endosomaga etkazib berish jarayonida ishlaydi. Ubiquitin bilan belgilangan oqsillar ESCRT-0 dan ESCRT-I ga, so'ngra ESCRT-II ga o'tkaziladi. ESCRT-II ESCRT-III bilan bog'lanadi, u vesikula yopiq bo'lgan yukni chimchilaydi.[5] ESCRT-II ning o'ziga xos jihatlari quyidagilardan iborat:
ESCRT-II - bu heterotetramer (2: 1: 1) ikkitadan iborat Vps25 subbirlik, bittasi Vps22 va bittasi Vps36 subbirlik.[3] Vps25 molekulalarida bog'langan PPXY motiflari mavjud qanotli spiral (WH) Vps22 va Vps36 asoslari Vps22 va Vps36 asos bo'lib, Vps25 molekulalari qurol sifatida Y shaklidagi kompleks hosil qiladi.[3][5] Vps25 molekulalarida ESCRT-II ning ESCRT-III bilan o'zaro ta'siri uchun mas'ul bo'lgan WH motiflari ham mavjud. Vps36 tarkibida ESCRT-I ning fosfatidilinozitol 3-fosfat va Vps28 ni bog'laydigan GLUE domeni mavjud.[3][5] Ikki sink barmog'i domenlar xamirturush VPS36 ning GLUE domeniga ulanadi. Ushbu sink barmoqlari domenlaridan biri Vps28 ning karboksi-terminal domenini, ikkinchisi esa ubikuitin bilan bog'laydi.[5]
ESCRT-III
ESCRT-III kompleksi, ehtimol ESCRT mexanizmlarining eng muhimidir, chunki u ESCRT vositachiligidagi barcha jarayonlarda rol o'ynaydi.[12] Membrananing abscissiyasi va virusli kurtaklari paytida ESCRT-III membranani yorilishidan oldin membrana toraygan joy atrofida o'ralgan uzun iplarni hosil qiladi.[10][13] Absitsiyaning ushbu vositachiligi markaziyspindlin murakkab.[14] Ushbu filamentli tuzilmalar multivikulyar tanani shakllantirish jarayonida ham mavjud va yuk oqsillarini hujayra sitozoliga kirib ketishining oldini olish uchun kurtak ochuvchi pufakchani tiqib qo'yadigan halqaga o'xshash panjara vazifasini bajaradi.[10] ESCRT-III mavjud va quyidagicha ishlaydi:
ESCRT-III kompleksi boshqa barcha ESCRT mexanizmlaridan faqat vaqtinchalik mavjudligi bilan ajralib turadi va tarkibida muhim va keraksiz qismlar mavjud.[1][10] Muhim bo'linmalar tegishli tartibda yig'ilishi kerak (Vps20, Snf7, Vps24, keyin Vps2) mashinaning ishlashi uchun.[5] Muhim bo'lmagan bo'linmalarga Vps60, Did2 va Ist1 kiradi.[10] Vps20 ESCRT-IIIni Snf7 polimer birikmasining yadrosi sifatida harakat qilib yig'ishni boshlaydi. Keyinchalik Vps24 kompleksni yopish va Vps2 ni jalb qilish uchun Snf7 bilan bog'lanadi.[1][3] Keyin Vps2 Vps4 ni kompleksga olib keladi.[15] Har bir kichik birlikning barcha "erkin" sitosolik shakllari yopiq hisoblanadi. Ya'ni, har bir bo'linmaning karboksi-terminali qismi o'z-o'zidan avtoinhibitatsion tarzda buklanib, monomerik subbirliklar.[1][3] ESCRT-III subbirliklarining ko'pchiligining karboksi-terminali muhim va ahamiyatsiz bo'lib, MIMlarni o'z ichiga oladi (MIT (mikrotubula o'zaro ta'sir va transport sohasi) menteskari mmotiflar.[16] Ushbu motiflar Vps4 va the bog'lash uchun javobgardir AAA-ATPase spastin.[3]
Vps4-Vta1
Vps4-Vta1 oqsillari ma'lum bir jarayon tugagandan so'ng boshqa ESCRT tarkibiy qismlarini (odatda ESCRT-III) membranalardan tozalash uchun talab qilinadi. Vps4 ESCRT-III kompleksini ajratib oladimi yoki kompleksni qayta tuzadimi yoki yo'qmi, shuning uchun bir vaqtning o'zida bitta tarkibiy qism to'kilishi haqida ba'zi munozaralar mavjud.[11] Vta1 Vps4-ning faollashtiruvchisi bo'lib, uning yig'ilishiga yordam beradi va AAA-ATPase faolligini oshiradi.[12][17] Ushbu oqsillarning ishlash tartibi quyidagicha:
Vps4 subbirliklarida ikkita funktsional domen mavjud, aminok terminal MIT domeni va markaziy AAA-ATPase domeni.[3] MIT domeni Vps4 ning VPS2 ning MIM domeni bilan o'zaro ta'siri uchun javobgardir.[1] AAA-ATPase domeni ESCRT-III kompleksini quvvatni demontaj qilish uchun ATPni gidrolizlaydi.[10] ESCRT-III ning ushbu "yalang'ochlanishi" barcha tegishli subbirliklarni keyingi foydalanish uchun qayta ishlashga imkon beradi.[10][11] Vta1 bitta VSL domenini o'z ichiga olgan dimerik oqsildir (u oqsillarda bo'lgani uchun shunday nomlangan Vps4, SBP1 va LIP5), bu Vps4 bilan bog'lanishni ta'minlaydi va ESCRT-III kichik birligi Vps60 bilan bog'lanish uchun MIT domeni. Vta1 muhim bo'lmagan bo'lsa-da, Vps4 halqasini yig'ishda yordam beradi, Vsp4 ning ATPase faolligini tezlashtiradi va ESCRT-III demontajini rag'batlantiradi.[5]
Bro1
Bro1-ning asosiy vazifasi - yollash deubikuitinazalar ESCRT-III kompleksiga.[18] Bu multisikulyar jismlar paydo bo'lishidan oldin lizozomada degradatsiyaga qaratilgan oqsillardan ubikuitin teglarini olib tashlashga olib keladi. Shuningdek, Bro1 ESCRT-IIIni barqarorlashtirishga yordam beradi, ammo ubikuitin yorliqlari yuk oqsillaridan ajralib chiqadi.[18]
Bro1, ESCRT-III ning Snf7 bilan bog'langan Bro1 amino-terminal domenini o'z ichiga oladi.[19] Ushbu birikma Bro1ni membranani abscissiya joyiga olib keladi. Bro1 shuningdek bog'laydi katalitik Doa4 domeni, ubikuitin gidrolaza (deubikuitinaza), uni abssitsiya joyiga olib keladi. Doa4 lizozomaga yo'naltirilgan yuk oqsillaridan ubikuitinni olib tashlaydi.[19]
Multivesular korpus biogenezi va yuklarni tashish
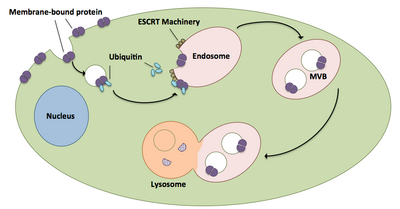
Ko'p qirrali jismlar hamma joyda mavjud bo'lgan oqsillar va retseptorlarni lizosomaga etkazishda katta rol o'ynaydi.[20] ESCRT komplekslari hamma joyda joylashgan yuklarni to'g'ridan-to'g'ri hujayraning endosomal bo'linmasiga kirib, ko'p qirrali jismlarni hosil qiladigan uyali pufakchalarga etkazadi.[20] Ushbu multivikulyar jismlar oxir-oqibat yukning parchalanishiga olib keladigan lizosoma bilan birlashadi.[15] Jarayonni, shu jumladan tegishli texnikani yanada chuqurroq tavsifi quyidagicha mavjud:
- ESCRT-0 komponentlari Vps27 va Hse1 har biri hamma joyda yuk bilan bog'lanadi.[1][20]
- Vps27 endosomal lipid bo'lgan fosfatidilinozitol 3-fosfat bilan bog'lanib, keyinchalik u butun kompleksni endosomaga qo'shib qo'yadi.[1][20]
- Vps27 ESCRT-I ning Vps23 subunitini bog'lab, ESCRT-I ni endosomaga keltiradi. ESCRT-I hamma joyda mavjud bo'lgan oqsillarni ham bog'lashi mumkin.[1][20]
- Vps36 ESCRT-I kichik birligi Vps28 bilan bog'lanadi, natijada ESCRT-II kompleksi ishga olinadi.[1]
- ESCRT-II ning Vps25 kichik birligi ESCRT-III kompleksining Vps20 bilan bog'lanadi va faollashtiradi.[1][15][20]
- Vps20, keyinchalik Vps24 tomonidan yopilgan Snf7 iplarining hosil bo'lishiga yadro beradi.[15]
- Vps24 Vps2-ni ishga oladi, bu Vps4-ni kompleksga olib keladi.[15]
- Vps4 ikkitadan qilingan teshik hosil qiladi geksamerik Vta1 bog'laydigan uzuklar.[1] Ushbu Vps4-Vta1 kompleksi ESCRT-III ning demontajini keltirib chiqaradi va multivikulyar tanani shakllantirishni tugatadi.[2]
Membranani yo'q qilish

Membranani yo'q qilish davomida sitokinez - bu ikkita qiz hujayralarni birlashtirgan membrananing yorilishi hujayraning bo'linishi. U bir qatorda saqlanib qolganligi sababli Arxeya, membranani abscissiyasi ESCRT texnikasi uchun eng qadimgi rol hisoblanadi.[5] Jarayon sentrosomal oqsil CEP55 MKLP1, a bilan birgalikda bo'linadigan hujayralar o'rta tanasiga jalb qilinadi mitotik kinesin - mikrotubulalar bilan birikadigan oqsil kabi.[5][22] So'ngra CEP55, ESCRT-I ning Vps23 subunitini va o'rta tanasining har ikki tomonida halqalarni hosil qiluvchi ALIX aksessuar oqsilini jalb qiladi.[5][10][11] ESCRT-I va ALIX o'zlarining Snf7 kichik birligi orqali ESCRT-IIIni yollashadi.[5] VSC20, Snf7, Vps24, Vps2 va Did2 ESCRT-III subbirliklari Vps23 tomonidan hosil qilingan halqalarga tutash spiral shaklidagi fibrilga aylanadi.[1][11][18] Ushbu spiralga o'xshash strukturaning hosil bo'lishi membranani deformatsiyalaydi va AAA-ATPaz spastinini Mid2 da hosil bo'lgan mikrotubulalarni ajratish uchun Did2 va Ist1 olib keladi.[11][18] Vps4 keyin kataliz qiladi ESCRT-III kompleksini demontaj qilish natijasida yangi ajratilgan ikkita qiz hujayralari paydo bo'ldi.[18] Metazoanlarda bu jarayon ko'proq o'rganilganligi sababli metazoan oqsillari yordamida membrana abscissiyasi jarayoni tasvirlangan.
Virusli tomurcuklanma

Virusli zarralarning chiqarilishi, shuningdek ma'lum virusli tomurcuklanma, bu bepul bo'lgan jarayon virionlar hujayralar ichidan ESCRT mexanizmini olib qochish orqali chiqariladi.[1][13] Retroviruslar, kabi OIV-1 va inson T-limfotrop virusi, shuningdek, bir qator o'ralgan viruslar shu jumladan Ebola virusi, xujayraning hujayrasidan chiqish uchun ESCRT texnikasini talab qiling.[1] Jarayon, virusli Gag oqsillari, o'zaro ta'sir o'tkazadigan retrovirus paltosining asosiy tarkibiy oqsillari tomonidan boshlanadi TSG101 ESCRT-I kompleksi va ALIX aksessuar oqsili.[11][12] ESCRT-III kichik birliklari (faqat CHMP4 va CHMP2 muhim ahamiyatga ega [9]) sitokinoz paytida membrananing abscissiyasi uchun ta'riflanganga o'xshash tarzda kurtakning bo'ynini toraytirish va kesish uchun virusli tomurcuklanma joyiga jalb qilinadi.[1][5][11] Keyin Vps4 ESCRT-III komponentlarini sitosolga qayta ishlaydi va virus hujayradan ajralib chiqadi.[5] Bu erda tasvirlangan mexanizm metazoan oqsillaridan foydalanadi, chunki virusli tomurcuk metazoanlarda kengroq o'rganilgan.
Adabiyotlar
- ^ a b v d e f g h men j k l m n o p q r s Shmidt O, Teis D (2012 yil fevral). "ESCRT apparati". Curr. Biol. 22 (4): R116-20. doi:10.1016 / j.cub.2012.01.028. PMC 3314914. PMID 22361144.
- ^ a b Babst M (avgust 2011). "MVB pufakchasining shakllanishi: ESCRTga bog'liq, ESCRTga bog'liq emas va orasidagi hamma narsa". Curr. Opin. Hujayra biol. 23 (4): 452–7. doi:10.1016 / j.ceb.2011.04.008. PMC 3148405. PMID 21570275.
- ^ a b v d e f g h men j k l Hurli JH, Xanson PI (avgust 2010). "ESCRT apparati tomonidan membrananing kurtaklanishi va sinishi: barchasi bo'ynida". Nat. Rev. Mol. Hujayra biol. 11 (8): 556–66. doi:10.1038 / nrm2937. PMC 2922035. PMID 20588296.
- ^ a b Piper RC, Katzmann DJ (2007). "Multivikulyar jismlarning biogenezi va funktsiyasi". Annu. Rev. Cell Dev. Biol. 23: 519–47. doi:10.1146 / annurev.cellbio.23.090506.123319. PMC 2911632. PMID 17506697.
- ^ a b v d e f g h men j k l m n o p q r s t Hurli JH (2010 yil dekabr). "ESCRT komplekslari". Krit. Rev. Biochem. Mol. Biol. 45 (6): 463–87. doi:10.3109/10409238.2010.502516. PMC 2988974. PMID 20653365.
- ^ Wollert T, Xarli JH (2010 yil aprel). "ESCRT komplekslari bo'yicha multivikulyar tanadagi biogenezning molekulyar mexanizmi". Tabiat. 464 (7290): 864–9. Bibcode:2010 yil natur.464..864W. doi:10.1038 / nature08849. PMC 2851844. PMID 20305637.
- ^ Ren X, Xarli JH (2010 yil mart). "ESCRT-0 ning VHS domenlari poliubiqitinatsiyalangan yuklarni yuqori avidlilikda bog'lashda hamkorlik qiladi". EMBO J. 29 (6): 1045–54. doi:10.1038 / emboj.2010.6. PMC 2845278. PMID 20150893.
- ^ Banerji S, Basu S, Sarkar S (2010). "Qiyosiy genomika eukaryotik nasl-nasablar bo'yicha FYVE va PX domen oqsillarining selektiv tarqalishi va domen tashkilotini ochib beradi. BMC Genomics. 11: 83. doi:10.1186/1471-2164-11-83. PMC 2837644. PMID 20122178.
- ^ a b Morita E, Sandrin V, Makkulaf J, Katsuyama A, Baci Xamilton I, Sundquist WI (mart 2011). "OIV-1 kurtaklari uchun ESCRT-III oqsil talablari". Hujayra xosti mikrobidir. 9 (3): 235–42. doi:10.1016 / j.chom.2011.02.004. PMC 3070458. PMID 21396898.
- ^ a b v d e f g h Adell MA, Teis D (2011 yil oktyabr). "ESCRT-III membrana sintez kompleksini yig'ish va demontaj qilish". FEBS Lett. 585 (20): 3191–6. doi:10.1016 / j.febslet.2011.09.001. PMC 3192940. PMID 21924267.
- ^ a b v d e f g h Myuller M, Adell MA, Teis D (avgust 2012). "Membrananing abscissioni: dinamik ESCRT-larda birinchi qarash". Curr. Biol. 22 (15): R603-5. doi:10.1016 / j.cub.2012.06.063. PMC 3414845. PMID 22877781.
- ^ a b v McDonald B, Martin-Serrano J (iyul 2009). "Iplar biriktirilmagan: virusli tomurcuklanma va sitokinezdagi ESCRT apparati". J. Cell Sci. 122 (Pt 13): 2167-77. doi:10.1242 / jcs.028308. PMC 2723143. PMID 19535732.
- ^ a b Jouvenet N, Jadina M, Bieniasz PD, Simon SM (2011 yil aprel). "Retrovirusli yig'ilish paytida ESCRT oqsillarini jalb qilish dinamikasi". Nat. Hujayra biol. 13 (4): 394–401. doi:10.1038 / ncb2207. PMC 3245320. PMID 21394083.
- ^ Glotzer, Maykl. "Sitokinesis: Membrana langari sifatida Centralspindlin oy nurlari", Hozirgi biologiya, 2013 yil 18-fevral
- ^ a b v d e Teis D, Saksena S, Judson BL, Emr SD (mart 2010). "ESCRT-II yuklarni saralash va multivesular korpus pufakchalarini shakllantirish uchun ESCRT-III iplarini yig'ishni muvofiqlashtiradi". EMBO J. 29 (5): 871–83. doi:10.1038 / emboj.2009.408. PMC 2837172. PMID 20134403.
- ^ Scott A, Gaspar J, Stuchell-Brereton MD, Alam SL, Skalicky JJ, Sundquist WI (sentyabr 2005). "Inson VPS4A ning MIT domenining tuzilishi va ESCRT-III oqsillari bilan o'zaro ta'siri". Proc. Natl. Akad. Ilmiy ish. AQSH. 102 (39): 13813–8. Bibcode:2005 yil PNAS..10213813S. doi:10.1073 / pnas.0502165102. PMC 1236530. PMID 16174732.
- ^ Azmi I, Devies B, Dimaano C, Peyn J, Ekkert D, Babst M, Katsmann DJ (fevral 2006). "AAA-ATPase Vps4 tomonidan ESCRTlarni qayta ishlash Vta1 da saqlangan VSL mintaqasi tomonidan tartibga solinadi". J. Hujayra Biol. 172 (5): 705–17. doi:10.1083 / jcb.200508166. PMC 2063703. PMID 16505166.
- ^ a b v d e Babst M, Devies BA, Katzmann DJ (oktyabr 2011). "MVB saralash va sitokinez paytida Vps4 ning regulyatsiyasi". Yo'l harakati. 12 (10): 1298–305. doi:10.1111 / j.1600-0854.2011.01230.x. PMC 3171586. PMID 21658171.
- ^ a b Vemmer M, Azmi I, G'arbiy M, Devis B, Katsmann D, Odorizzi G (yanvar 2011). "Bro1 Snf7 bilan bog'lanishi xamirturushdagi ESCRT-III membrana sinitsion faolligini tartibga soladi". J. Hujayra Biol. 192 (2): 295–306. doi:10.1083 / jcb.201007018. PMC 3172170. PMID 21263029.
- ^ a b v d e f Hurli JH, Emr SD (2006). "ESCRT komplekslari: membrana savdosi tarmog'ining tuzilishi va mexanizmi". Annu Rev Biofhys Biomol tuzilishi. 35: 277–98. doi:10.1146 / annurev.biophys.35.040405.102126. PMC 1648078. PMID 16689637.
- ^ Karmena M (2012 yil iyul). "Abscission tekshiruv punkti nazorati: Aurora B bilan o'rtada qolib ketgan". Biol-ni oching. 2 (7): 120095. doi:10.1098 / rsob.120095. PMC 3411112. PMID 22870391.
- ^ Zhu C, Bossy-Wetzel E, Jiang V (iyul 2005). "INCENP tomonidan shpindel midzone / midbody-ga MKLP1ni jalb qilish o'rta hujayralarni hosil qilish va inson hujayralarida sitokinezni yakunlash uchun juda muhimdir". Biokimyo. J. 389 (Pt 2): 373-81. doi:10.1042 / BJ20050097. PMC 1175114. PMID 15796717.