Oxirgi plastinka potentsiali - End-plate potential

Plitaning so'nggi potentsiali (RaI) depolarizatsiyaga olib keladigan kuchlanishdir skelet mushaklari sabab bo'lgan tolalar neyrotransmitterlar postsinaptik membranaga bog'lanib asab-mushak birikmasi. Ular "so'nggi plitalar" deb nomlanadi, chunki mushak tolalarining postsinaptik terminallari katta, likopchaga o'xshash ko'rinishga ega. Qachon harakat potentsiali ga etadi akson terminali a vosita neyroni, nörotransmitterlarni tashiydigan pufakchalar (asosan atsetilxolin ) bor exocytosed va tarkibi asab-mushak birikmasiga chiqariladi. Ushbu nörotransmitterlar postsinaptik membranadagi retseptorlari bilan bog'lanib, uning depolarizatsiyasiga olib keladi. Harakat potentsiali bo'lmagan taqdirda, atsetilxolin pufakchalari o'z-o'zidan nerv-mushak birikmasiga oqib chiqadi va postsinaptik membranada juda kichik depolarizatsiyani keltirib chiqaradi. Ushbu kichik javob (~ 0.4mV)[1] miniatyuradagi so'nggi plastinka potentsiali (MEPP) deb nomlanadi va bitta atsetilxolin o'z ichiga olgan pufakcha tomonidan hosil bo'ladi. Bu mushak ichiga tushishi mumkin bo'lgan eng kichik depolarizatsiyani anglatadi.
Nerv-mushak birikmasi

Nerv-mushak birikmasi - bu an o'rtasida hosil bo'lgan sinaps alfa motorli neyron (a-MN) va skelet mushak tolasi. Mushak qisqarishi uchun harakat potentsiali birinchi navbatda vosita neyronining akson terminaliga yetguncha asab orqali tarqaladi. Keyinchalik, vosita neyroni mushak tolalarini asab-mushak birikmasining postsinaptik membranasida harakat potentsialini keltirib chiqarishi bilan qisqarishgacha innervatsiya qiladi.
Asetilkolin
Oxirgi plastinka potentsiali deyarli butunlay skelet mushaklaridagi atsetilxolin nörotransmitteri tomonidan ishlab chiqariladi. Asetilkolin organizmdagi ikkinchi muhim qo'zg'atuvchi nörotransmitterdir glutamat. Bu boshqaradi somatosensor tizim bu tegish, ko'rish va eshitish hislarini o'z ichiga oladi. Bu 1914 yilda aniqlangan birinchi neyrotransmitter edi Genri Deyl. Asetilxolin neyronning sitoplazmasida sintezlanadi xolin va atsetil-KoA. Xolin asiltransferaza - bu atsetilxolinni sintez qiladigan ferment va ko'pincha atsetilxolin ishlab chiqarish bilan bog'liq tadqiqotlarda marker sifatida ishlatiladi. Asetilkolinni ishlatadigan neyronlarga xolinergik neyronlar deyiladi va ular mushaklarning qisqarishi, xotira va o'rganishda juda muhimdir.[2]
Ion kanallari
Membranalarning polarizatsiyasi tomonidan boshqariladi natriy, kaliy, kaltsiy va xlorid ion kanallari. Nerv-mushak birikmasi va so'nggi plastinka potentsialida ikki xil ion kanallari mavjud: kuchlanishli ionli kanal va ligandli ionli kanal. Voltajli yopiq ionli kanallar membranali voltajning o'zgarishiga ta'sir qiladi, bu esa kuchlanishli ionli kanalning ochilishiga olib keladi va ma'lum ionlarning o'tishiga imkon beradi. Ligand eshikli ion kanallari nörotransmitterlar kabi ba'zi molekulalarga javob beradi. A ning majburiyligi ligand ion kanali oqsilidagi retseptorga konformatsion o'zgarishni keltirib chiqaradi, bu esa ma'lum ionlarning o'tishiga imkon beradi.
Presinaptik membrana
Odatda vosita neyronining dam oluvchi membrana potentsiali -70mV dan -50 darajagacha ushlab turiladi, uning tashqarisida natriy yuqori va ichkarida kaliy yuqori bo'ladi. Harakat potentsiali asab bo'ylab tarqalib, vosita neyronining akson terminaliga etib borganida, membranadagi kuchlanishning o'zgarishi kaltsiy kuchlanishining kaltsiy ionlarining kirib kelishiga imkon beradigan eshik kanallarining ochilishiga olib keladi. Ushbu kaltsiy ionlari presinaptik membranaga biriktirilgan atsetilxolin pufakchalari orqali atsetilxolinni chiqarishga olib keladi. ekzotsitoz sinaptik yoriqqa.[3]
Postsinaptik membrana
EPP asosan atsetilxolinning postsinaptik membranadagi retseptorlari bilan bog'lanishidan kelib chiqadi. Asetilkolin retseptorlari ikki xil: nikotinik va muskarinik. Nikotin retseptorlari tez uzatish uchun ligandli eshikli kanallar. Nerv-mushak birikmasidagi barcha atsetilxolin retseptorlari nikotinikdir. Muskarinik retseptorlari bor G oqsillari bilan bog'langan retseptorlari ikkinchi xabarchi ishlatadigan. Ushbu retseptorlar sekin va shuning uchun miniatyura so'nggi plastinka potentsialini (MEPP) o'lchashga qodir emas. Ular joylashgan parasempatik asab tizimi vagus asabida va oshqozon-ichak trakti. Xomilalik rivojlanish davrida atsetilxolin retseptorlari postsinaptik membranada to'planib, o'sib chiqayotgan embriondagi asab terminalining butun yuzasi signal berishdan oldin ham qoplanadi. To'rt xil genning to'rt xil oqsilidan tashkil topgan beshta bo'linma nikotin atsetilxolin retseptorlarini o'z ichiga oladi, shuning uchun ularni qadoqlash va yig'ish turli xil omillarga ega bo'lgan juda murakkab jarayon. Enzim mushaklariga xos kinaz (MuSK) rivojlanayotgan postsinaptik mushak hujayrasida signalizatsiya jarayonlarini boshlaydi. U postsinaptik asetilkolin retseptorlari klasterlarini barqarorlashtiradi, sinaptik genlarning mushak tolalari yadrolari bilan transkripsiyasini osonlashtiradi va akson o'sish konusining differentsiatsiyasini keltirib, differentsial nerv terminalini hosil qiladi.[4] Substrat laminin myotubalar yuzalarida atsetilxolin retseptorlari klasterlarining rivojlanib borishini keltirib chiqaradi.[5]
Boshlash
Sinaptik pufakchalar
Barcha neyrotransmitterlar ekzotsitoz orqali sinaptik yorilishga chiqadi sinaptik pufakchalar. Ikki xil nörotransmitter pufakchalari mavjud: katta zich yadro pufakchalari va mayda tiniq pufakchalar. Katta zich yadro pufakchalarida neyropeptidlar va neyronning hujayra tanasida hosil bo'lgan va keyinchalik akson terminaliga tez aksonal transport orqali uzatiladigan yirik neyrotransmitterlar mavjud. Kichik tiniq yadro pufakchalari, presinaptik terminallarda mahalliy sintez qilingan kichik molekula nörotransmitterlarini tashiydi. Tugatilgan nörotransmitter pufakchalar presinaptik membrana bilan bog'langan. Harakat potentsiali motor neyron aksonida tarqalib, akson terminaliga kelganda akson terminalining depolarizatsiyasini keltirib chiqaradi va kaltsiy kanallarini ochadi. Bu nörotransmitterlarning vazikula ekzositozi orqali chiqarilishiga olib keladi.
Ekzotsitozdan keyin pufakchalar sinaptik pufak tsikli deb ataladigan jarayon davomida qayta ishlanadi. Olingan vazikulyar membranalar bir nechta hujayra ichidagi bo'linmalar orqali o'tib, u erda yangi sinaptik pufakchalar hosil qilish uchun o'zgartiriladi. Keyinchalik ular neyrotransmitterlarni tashish va chiqarish uchun yana kerak bo'lguncha zaxira hovuzida saqlanadi.
Zaxira hovuzidan farqli o'laroq, sinaptik pufakchalarning osongina bo'shatilishi mumkin bo'lgan havzasi ishga tushirishga tayyor. Tezda bo'shatiladigan basseyndan pufakchalarning tükenmesi uzoq muddatli yuqori chastotali stimulyatsiya paytida yuzaga keladi va uyg'otilgan EPP hajmi kamayadi. Ushbu asab-mushak tushkunligi stimulyatsiya paytida kamroq nörotransmitter ajralib chiqishi bilan bog'liq. Zaiflashmaslik uchun, 30 Hz dan past bo'lgan stimulyatsiya chastotalarida sodir bo'lishi mumkin bo'lgan to'ldirish va tükenme o'rtasida muvozanat bo'lishi kerak.[6]
Vezikula nörotransmitterlarini ekzotsitoz orqali chiqarganda, u barcha tarkibni sinaptik yoriqqa bo'shatadi. Vesikulalardan neyrotransmitterning chiqarilishi kvant deb aytiladi, chunki faqat butun sonli pufakchalar chiqarilishi mumkin. 1970 yilda, Bernard Kats dan London universiteti g'olib bo'ldi Nobel mukofoti fiziologiya yoki tibbiyot uchun asab-mushak birikmasidagi shovqinni tahlil qilish asosida atsetilxolin pufakchalarining miqdoriy miqdorini statistik jihatdan aniqlash uchun. Mexanik statistika bo'yicha kitobdan foydalanish[tushuntirish kerak ], u bir vaqtning o'zida sodir bo'layotgan alohida voqealar hajmini taxmin qilishga qodir edi.
Asetilkolinning sinaptik pufakchalari diametri 30 nm bo'lgan aniq yadroli sinaptik pufakchalardir. Har bir atsetilxolin pufakchasida taxminan 5000 atsetilxolin molekulasi mavjud. Vesikulalar atsetilxolinning butun miqdorini chiqaradi va bu amplituda 1mV dan kam bo'lgan va chegaraga etishish uchun etarli bo'lmagan miniatyuradagi so'nggi plastinka potentsiallarini (MEPP) paydo bo'lishiga olib keladi.[7]
Miniatyuradagi so'nggi plastinka potentsiali (MEPP)
Miniatyuradagi so'nggi plastinka potentsiallari - bu bitta vazikulaning sinaptik yoriqqa chiqishi natijasida kelib chiqadigan postsinaptik terminalning kichik (~ 0,4mV) depolarizatsiyasi. Atsetilkolinni o'z ichiga olgan neyrotransmitter pufakchalar o'z-o'zidan nerv terminali bilan to'qnashadi va aksondan signal olmasa ham asetilkolinni nerv-mushak birikmasiga chiqaradi. Ushbu kichik depolarizatsiya chegaraga erishish uchun etarli emas va shuning uchun postsinaptik membranada harakat potentsiali paydo bo'lmaydi.[8]MEPPlar bilan tajriba o'tkazishda, odatda o'z-o'zidan paydo bo'ladigan ta'sir potentsiallari paydo bo'lishi aniqlandi, ular oddiy tirnoqli mushaklarda so'nggi plastinka pog'onalari deb ataladi, bu esa hech qanday stimulga ega emas. Ushbu so'nggi plastinka tirnoqlari tufayli mushak tolalari shikastlanishi yoki tirnash xususiyati natijasida paydo bo'lgan deb ishonishgan elektrodlar. So'nggi tajribalar shuni ko'rsatdiki, bu so'nggi plastinka tirnoqlari aslida sabab bo'lgan mushak millari va ikkita aniq naqshga ega: kichik va katta. Kichik uchli plastinka pog'onalari signalning tarqalishisiz salbiy boshlanishiga ega va katta plastinka pog'onalari motor birligining potentsialiga (MUP) o'xshaydi. Mushaklar shpindellari - bu mushaklarning cho'zilishini yoki cho'zilishini o'lchaydigan sezgir retseptorlari bo'lib, tegishli javob uchun ma'lumotni orqa miya yoki miyaga etkazadi.[9]
To'siq salohiyati ("Hammasi yoki yo'q")
Harakat potentsiali ko'plab atsetilxolin pufakchalari chiqarilishiga olib kelganda, atsetilxolin nerv-mushak birikmasi bo'ylab tarqaladi va mushak tolasidagi ligandli nikotinik retseptorlari (selektiv bo'lmagan kation kanallari) bilan bog'lanadi. Bu natriy va kaliy ionlari oqimining ko'payishiga imkon beradi, bu esa sarkolemmaning (mushak hujayralari membranasi) depolarizatsiyasini keltirib chiqaradi. Atsetilkolinning individual sinaptik pufakchadan chiqishi bilan bog'liq bo'lgan kichik depolarizatsiya miniatyuradagi so'nggi plastinka potentsiali (MEPP) deb nomlanadi va uning kuchi + 0,4mV ga teng. MEPPlar qo'shimcha bo'lib, oxir-oqibat plastinka potentsialini (EPP) -100mV dan -60mV pol potentsialigacha oshiradi, bu erda postsinaptik membranadagi kuchlanishli ion kanallari ochilib, natriy ionlarining to'satdan oqimiga imkon beradi. sinaps va depolarizatsiyadagi keskin pog'ona. Ushbu depolarizatsiya kuchlanishining ko'tarilishi postsinaptik membranada tarqaladigan harakat potentsialini keltirib chiqaradi, bu esa mushaklarning qisqarishiga olib keladi. Shuni ta'kidlash kerakki, EPPlar harakat potentsiali emas, balki ular harakat potentsialini qo'zg'atadi, normal mushak qisqarishida taxminan 100-200 atsetilxolin pufakchalari ajralib chiqadi, bu depolarizatsiyaga olib keladi, bu kattaligi MEPPdan 100 baravar katta. Bu esa membrana potentsialini -100mV dan -60mV gacha bo'lgan chegarada + 40mV (100 x 0,4mV = 40mV) depolarizatsiyaga olib keladi.[7]
Harakatning potentsial bosqichlari
Membrana potentsiali pol darajasiga etganidan so'ng, harakat potentsiali paydo bo'ladi va membrana qutblanishida keskin pog'onani keltirib chiqaradi. Harakat potentsialining beshta bosqichi mavjud: chegara, depolarizatsiya, tepalik, repolarizatsiya va giperpolarizatsiya.
Eshik - bu MEPPlarning yig'indisi ma'lum potentsialga yetganda va kuchlanishli ionli kanallarning ochilishini keltirib chiqaradi. Natriy ionlarining tez oqishi membrana potentsialini ijobiy zaryadga olib keladi. Kaliy ionlari kanallari natriy ionlari kanallariga qaraganda sekinroq ishlaydi va shuning uchun membrana potentsiali eng yuqori darajaga ko'tarila boshlagach, kaliy ionlari kanallari ochilib, natriy oqimiga qarshi kaliy chiqishiga olib keladi. Eng yuqori nuqtada kaliy oqimi natriy oqimiga teng keladi va membrana qutblanishni o'zgartirmaydi.
Repolarizatsiya paytida natriy kanallari faolsizlana boshlaydi, natijada kaliy ionlarining aniq oqimi hosil bo'ladi. Bu membrana potentsialining -100mV tinchlanadigan membrana potentsialiga tushishiga olib keladi. Giperpolarizatsiya sekin ta'sir etuvchi kaliy kanallarini o'chirish uchun ko'proq vaqt talab etilishi sababli yuzaga keladi, shuning uchun membrana dam olish potentsialini haddan tashqari oshirib yuboradi. U asta-sekin dam olish potentsialiga qaytadi va boshqa harakat potentsiali paydo bo'lishiga tayyor.
Giperpolarizatsiya bosqichidan oldin harakat potentsiali davomida membrana har qanday stimulga javob bermaydi. Boshqa harakat potentsialini keltirib chiqara olmaslik bu mutlaq refrakter davr deb nomlanadi. Giperpolarizatsiya davrida membrana yana stimulyatsiyalarga javob beradi, ammo harakat potentsialini kuchaytirish uchun bu juda yuqori kirishni talab qiladi. Ushbu bosqich nisbiy refrakter davr sifatida tanilgan.
Nerv-mushak birikmasida harakat potentsiali tugagach, ishlatilgan atsetilxolin ferment tomonidan sinaptik yoriqdan tozalanadi. atsetilxolinesteraza. Fermentlarning neyrotransmitterlarni sinaptik yoriqdan tozalay olmasliklari natijasida bir qator kasalliklar va muammolar paydo bo'lishi mumkin, bu esa potentsial tarqalishining davom etishiga olib keladi.[10]
Klinik qo'llanmalar
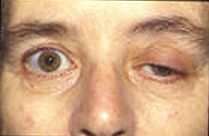
Hozirgi tadqiqotlar so'nggi plastinka potentsiallari va ularning mushaklarning faoliyatiga ta'siri haqida ko'proq ma'lumot olishga harakat qilmoqda. Ko'pgina hozirgi kasalliklar buzilgan so'nggi plastinka potentsial faolligini o'z ichiga oladi. Altsgeymer kasalligida, beta amiloid atsetilxolin retseptorlariga yopishadi va atsetilxolinning bog'lanishini inhibe qiladi. Bu signalning kamroq tarqalishiga va eshikka etib bormaydigan kichik RaIlarga olib keladi. Asetilkolin bilan miya jarayonlarini tahlil qilib, shifokorlar beta amiloidning qancha ekanligini o'lchashlari va uning Altsgeymerga ta'sirini baholashlari mumkin.[11]Myasthenia gravis bu otoimmun kasallik, bu erda organizm asab-mushak birikmasidagi postsinaptik membranadagi atsetilxolin retseptorlariga qarshi antitellar ishlab chiqaradi. Mushaklarning charchoqlanishi va zaifligi, foydalanish paytida yomonlashadi va dam olish bilan yaxshilanadi, bu kasallikning o'ziga xos xususiyati. Bog'lanish uchun mavjud bo'lgan atsetilxolin retseptorlari miqdori cheklanganligi sababli simptomatik davolash asetilkolinni asab-mushak birikmasidagi parchalanishini kamaytirish uchun atsetilxolinesteraza inhibitori yordamida amalga oshiriladi, shuning uchun blokirovka qilinmagan retseptorlarning oz miqdori uchun atsetilxolin etarli bo'ladi. oxirgi plastinka atsetilxolin esteraza (AChE) etishmovchiligi natijasida yuzaga keladigan anormallik miyastenik gravis uchun patofizyologik mexanizm bo'lishi mumkin. AChE etishmovchiligi bo'lgan bemorga o'tkazilgan tadqiqotda shifokorlar uning boshqa mushaklarda titrash bilan proksimal va trunkal mushaklarning kuchli zaifligini rivojlantirganligini ta'kidladilar. Atsetilkolin retseptorlari chayqalishi va blokirovka tezligining kombinatsiyasi miyasteniya gravis holatlarida kuzatilganga o'xshash pasaygan plastinka potentsialini keltirib chiqarishi aniqlandi.[12]Dvigatel birligining potentsialini (MUP) tadqiq qilish, patologik kasalliklarning miogen yoki neyrogen kelib chiqishiga qarab o'zgarishini doimiyligini o'lchash yo'li bilan baholashda mumkin bo'lgan klinik qo'llanmalarga olib keldi. Dvigatel birligining potentsiallari - bu motor birliklari tomonidan ishlab chiqarilgan, amplituda, davomiylik, faza va tepalik bilan tavsiflanishi mumkin bo'lgan elektr signallari va tartibsizlik koeffitsienti (IQ) eng yuqori raqamlar va amplitudalar asosida hisoblanadi.[13]Lambert-Eaton miyastenik sindromi bu presinaptik kaltsiy kanallari otoimmun buzilishiga olib keladigan buzilishdir, bu esa kamroq neyrotransmitter pufakchalarning ekzotsitlanishiga olib keladi. Bu kamroq pufakchalar chiqarilishi sababli kichik RaIlarni keltirib chiqaradi. Ko'pincha kichikroq RaIlar eshigiga etib bormaydilar, bu esa bemorlarda mushaklarning kuchsizligi va charchoqni keltirib chiqaradi. Ko'pgina hayvonlar o'zlarini himoya qilish va o'ljani o'ldirish uchun neyrotoksinlardan foydalanadilar. Tetrodotoksin kabi ba'zi bir zaharli baliqlarda uchraydigan zahar puferfish va triggerfish bu natriy ion kanallarini to'sib qo'yadi va postsinaptik membranada ta'sir potentsialini oldini oladi. Tetraetilammoniy hasharotlar tarkibida kaliy kanallarini bloklaydi. Ilonlarda uchraydigan alfa nörotoksin atsetilxolin retseptorlari bilan bog'lanib, atsetilxolinning bog'lanishiga yo'l qo'ymaydi. Alfa-latrotoksin ichida topilgan qora beva o'rgimchaklar akson terminalida katta miqdordagi kaltsiy oqimini keltirib chiqaradi va neyrotransmitterning ortiqcha tarqalishiga olib keladi.Botulinum toksini bakteriyalar tomonidan ishlab chiqarilgan Clostridium botulinum eng kuchli toksik oqsil hisoblanadi. Nörotransmitter pufakchalarini joylashtirishni inhibe qilish orqali asab-mushak birikmasida atsetilxolin ajralishini oldini oladi.
Shuningdek qarang
- Asetilkolin
- Harakat salohiyati
- Alfa-latrotoksin
- Altsgeymer kasalligi
- Botulinum toksini
- Motor neyron
- Muskarinik retseptorlari
- Myasthenia gravis
- Nerv-mushak birikmasi
- Neyrotransmitter
- Nikotin retseptorlari
- Skelet mushaklari
- Sinaptik vazikula
- Tetraetilammoniy
- Tetrodotoksin
Adabiyotlar
- ^ Boron, V.; Boulpaep, E. (2012). Tibbiy fiziologiya. Filadelfiya, Pensilvaniya: Sonders, Elsevier inc. p. 224. ISBN 978-0-8089-2449-4.
- ^ Kimura Y; Oda Y; Deguchi T; Higashida H. (1992). "Sichqoncha xolin-asetiltransferaza CDNA bilan transfektsiya qilingan neyroblastoma X glioma gibrid NG108-15 hujayralarida atsetilxolin sekretsiyasining kuchayishi". FEBS xatlari. 314 (3): 409–412. doi:10.1016 / 0014-5793 (92) 81516-O. PMID 1468577. S2CID 4956377.
- ^ Lin S, Landmann L, Ruegg MA, Brenner HR (2008). "Sutemizuvchilarning asab-mushak birikmasining shakllanishida nervga qarshi mushaklardan kelib chiqadigan omillarning roli" (PDF). Neuroscience jurnali. 28 (13): 3333–3340. doi:10.1523 / JNEUROSCI.5590-07.2008. PMID 18367600. S2CID 18659773.
- ^ Cole RN, Reddel SW, Gervasio OL, Phillips WD (2008). "Anti-MuSK bilan kasallangan antikorlar sichqonchani nerv-mushak birikmasini buzadi". Nevrologiya yilnomalari. 63 (6): 782–789. doi:10.1002 / ana.21371. PMID 18384168. S2CID 205340971.
- ^ Teressa G, Prives J (2008). "Mushak hujayralarida postsinaptik membrana yig'ilishini hujayra madaniyati asosida tahlil qilish". Onlayn biologik protseduralar. 10 (1): 58–65. doi:10.1251 / bpo143. PMC 2683546. PMID 19461953.
- ^ Van Lunteren E, Moyer M (2005). "Sichqoncha diafragmasida so'nggi plastinka potentsialini tiklashning ikki fazali reatiyasini modulyatsiya qilish". Mushak va asab. 31 (3): 321–330. doi:10.1002 / mus.20245. PMID 15654692. S2CID 31071429.
- ^ a b Takeda T, Sakata A, Matsuoka T (1999). "Umurtqali hayvonlarning nerv-mushak birikmasida miniatyuradagi so'nggi plastinka potentsialining paydo bo'lishidagi fraktal o'lchamlar". Neyro-psixofarmakologiya va biologik psixiatriyadagi taraqqiyot. 23 (6): 1157–1169. doi:10.1016 / S0278-5846 (99) 00050-0. PMID 10621955. S2CID 30988488.
- ^ Sellin LC, Molgo J, Thornquist K, Hansson B, Thesleff S (1996). "Mumkin kelib chiqishi to'g'risida ulkan yoki sekin ko'tarilgan asab-mushak birikmasidagi miniatyura so'nggi plastinka potentsiali ". Pflygers Archiv: Evropa fiziologiyasi jurnali. 431 (3): 325–334. doi:10.1007 / BF02207269. PMID 8584425. S2CID 8748384.
- ^ Partanen J (1999). "Inson elektromiyogrammasidagi so'nggi plastinka pog'onalari. Fusimotorlar nazariyasini qayta ko'rib chiqish". Fiziologiya jurnali-Parij. 93 (1–2): 155–166. doi:10.1016 / S0928-4257 (99) 80146-6. S2CID 4961877.
- ^ Purves D, Augustine G va boshq. "Asab hujayralarining elektr signallari". Nevrologiya. Sinauer Associates, Inc: Sanderlend, Massachusets, 2008. 25-39.
- ^ Prives J, Stoni Brukdagi Nyu-York shtat universiteti farmakologiya professori. Suhbatdosh Per Uotson. 2008-11-18.
- ^ Kohara N, Lin TS, Fukudome T, Kimura J, Sakamoto T va boshq. (2002). "Tug'ma plastinka atsetilxolinesteraza etishmovchiligi bo'lgan bemorda zaiflik patofiziologiyasi". Mushak va asab. 25 (4): 585–592. doi:10.1002 / mus.10073. PMID 11932977. S2CID 45891411.
- ^ Zalewska E, Hausmanova-Petrushevich I, Stahlberg E (2004). "Noto'g'ri motor birligining potentsiallari bo'yicha modellashtirish ishlari". Klinik neyrofiziologiya. 115 (3): 543–556. doi:10.1016 / j.clinph.2003.10.031. PMID 15036049. S2CID 43828995.