Intron - Intron - Wikipedia
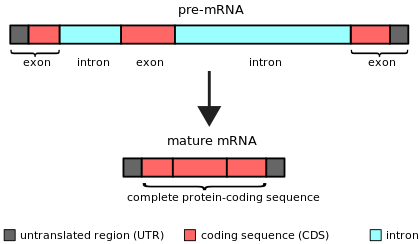
An intron (uchun intragenik mintaqa) har qanday nukleotidlar ketma-ketligi ichida a gen tomonidan olib tashlangan RNK qo'shilishi davomida kamolot oxirgi RNK mahsulotining.[1][2] Boshqacha qilib aytganda, intronlar - bu RNK transkriptining kodlamaydigan hududlari yoki uni kodlovchi DNK, bu oldin biriktirish yo'li bilan yo'q qilinadi. tarjima.[3][4] So'z intron atamasidan kelib chiqqan intragenik mintaqa, ya'ni gen ichidagi mintaqa.[5] Atama intron gen ichidagi DNK ketma-ketligini va RNKdagi tegishli ketma-ketlikni anglatadi stenogrammalar.[6] RNK qo'shilishidan so'ng yakuniy etuk RNKda birlashtirilgan qatorlar exons.[7]
Intronlar ko'pchilik organizmlar va ko'plab viruslarning genlarida uchraydi va genlarning keng doiralarida, shu jumladan hosil qiluvchi genlarda joylashgan bo'lishi mumkin. oqsillar, ribosomal RNK (rRNK) va transfer RNK (tRNA). Intron o'z ichiga olgan genlardan oqsillar hosil bo'lganda, RNK qo'shilishi RNKni qayta ishlash yo'lining bir qismi sifatida sodir bo'ladi. transkripsiya va tarjimadan oldin.[7]
Kashfiyot va etimologiya
Intronlar birinchi marta proteinlarni kodlovchi genlarida topilgan adenovirus,[8][9] va keyinchalik transfer RNK va ribosomal RNK genlarini kodlovchi genlarda aniqlandi. Hozirgi vaqtda intronlar barcha biologik shohliklar tarkibidagi barcha organizmlar va viruslarda genlar tarkibida bo'lganligi ma'lum.
Genlar bo'lganligi bo'lingan yoki uzilgan intronlar tomonidan mustaqil ravishda 1977 yilda kashf etilgan Fillip Allen Sharp va Richard J. Roberts, buning uchun ular birgalikda bo'lishdi Fiziologiya yoki tibbiyot bo'yicha Nobel mukofoti 1993 yilda.[10] Atama intron Amerika tomonidan kiritilgan biokimyogar Valter Gilbert:[5]
". Tushunchasi tsistron [ya'ni, gen] ..., etuk xabarchidan yo'qoladigan mintaqalarni o'z ichiga olgan transkripsiya birligi bilan almashtirilishi kerak - bu bizni intronlar (intragenik mintaqalar uchun) deb atashni tavsiya etamiz - ifoda etiladigan mintaqalar bilan almashtirib turamiz - ekzonlar. "(Gilbert 1978)
Atama intron shuningdek, murojaat qiladi intrakistron, ya'ni a ichida paydo bo'lgan qo'shimcha DNK bo'lagi tsistron.[11]
Ba'zida intronlar chaqiriladi oraliq ketma-ketliklar,[12] "oraliq ketma-ketlik" atamasi so'nggi gen mahsulotida mavjud bo'lmagan ichki nuklein kislota ketma-ketliklarining har qanday oilasiga tegishli bo'lishi mumkin, shu jumladan tamsayılar, tarjima qilinmagan ketma-ketliklar (UTR ) va nukleotidlar tomonidan olib tashlangan RNK tahriri, intronlardan tashqari.
Tarqatish
Turli xil genomlar ichidagi intronlarning chastotasi biologik organizmlar spektrida keng farq qilishi kuzatiladi. Masalan, intronlar jag 'umurtqali hayvonlarning (masalan, odamlar va sichqonlar) yadro genomida juda keng tarqalgan bo'lib, bu erda oqsillarni kodlovchi genlar deyarli har doim bir nechta intronlarni o'z ichiga oladi, ba'zi eukaryotik mikroorganizmlarning yadro genlarida esa intronlar kam uchraydi,[13] masalan novvoy / pivo achitqi (Saccharomyces cerevisiae). Aksincha, mitoxondriyal genomlar umurtqali hayvonlar umuman intronlardan mahrum, ökaryotik mikroorganizmlar esa ko'plab intronlarni o'z ichiga olishi mumkin.[14]
Ayniqsa, haddan tashqari holat Drosophila dhc7 ≥3,6 megabaza (Mb) intronini o'z ichiga olgan gen, bu transkripsiya qilish uchun uch kun davom etadi.[15][16] Boshqa tomondan, yaqinda o'tkazilgan bir tadqiqot shuni ko'rsatadiki, eng qisqa vaqt ichida eukaryotik intron uzunligi odamga tegishli bo'lgan 30 tayanch jufti (bp). MST1L gen.[17]
Tasnifi
Barcha intron o'z ichiga olgan RNK molekulalarining birikishi yuqorida ta'riflanganidek, yuzaki o'xshashdir. Shu bilan birga, intronlarning turlarini DNK ketma-ketligini tahlil qilish va RNK qo'shilish reaktsiyalarining genetik va biokimyoviy tahlillari bilan intron tuzilishini tekshirish orqali aniqlandi.
Kamida to'rtta intronlarning aniq sinflari aniqlandi:[1]
- Yadro oqsilini kodlovchi genlarning intronlari tomonidan olib tashlangan splitseozomalar (splitseozoma intronlari)
- Oqsillar (tRNK intronlari) bilan chiqarib tashlanadigan yadro va arxaeal RNK genlaridagi intronlar
- O'z-o'zini qo'shish I guruh intronlari tomonidan olib tashlangan RNK katalizi
- O'z-o'zini qo'shish II guruh intronlari ular RNK kataliz yordamida olib tashlanadi
III guruh intronlari beshinchi oila bo'lish taklif qilingan, ammo ularning birikishida vositachilik qiladigan biokimyoviy apparatlar haqida kam narsa ma'lum. Ular II guruh intronlari, va ehtimol splitseozomal intronlar bilan bog'liq ko'rinadi.[18]
Splitseozoma intronlari
Yadrodan oldingi mRNA intronlari (splitseozomal intronlar) intronlar va ekzonlar orasidagi chegaralarda joylashgan o'ziga xos intron sekanslari bilan tavsiflanadi.[19] Ushbu ketma-ketliklar splitseozomal RNK molekulalari tomonidan qo'shilish reaktsiyalari boshlanganda tan olinadi.[20] Bundan tashqari, ular tarkibida tarmoqlanish nuqtasi, intronning 3 'uchi yaqinidagi ma'lum bir nukleotidlar ketma-ketligi, ular qo'shilish jarayonida intronning 5' uchi bilan kovalent ravishda bog'lanib, tarvaqaylab hosil bo'ladi (lariya) intron. Ushbu uchta saqlanib qolgan elementlardan tashqari, mRNA yadrosi old intron sekanslari juda o'zgaruvchan. Yadrodan oldingi mRNA intronlari ko'pincha atrofdagi ekzonlarnikidan ancha uzunroq.
tRNA intronlari
Olib tashlash uchun oqsillarga bog'liq bo'lgan uzatish RNK intronlari, biriktirilmagan tRNA prekursorlarining antikodon tsikli ichida ma'lum bir joyda sodir bo'ladi va tRNK biriktiruvchi endonukleaza orqali olib tashlanadi. Keyin ekzonlar ikkinchi protein - tRNK biriktiruvchi ligaza bilan birlashtiriladi.[21] E'tibor bering, o'z-o'zidan birikadigan intronlar ba'zan tRNK genlarida ham uchraydi.[22]
I va II guruh intronlari
I va II guruh intronlari oqsillarni kodlovchi genlarda uchraydi (xabarchi RNK ), transfer RNK va ribosomal RNK juda keng tirik organizmlarda.,[23][24] RNKga transkripsiyadan so'ng, I va II guruh intronlari, shuningdek, o'ziga xos, murakkab bo'lib katlanishga imkon beradigan keng ichki o'zaro ta'sirlarni amalga oshiradi. uch o'lchovli arxitektura. Ushbu murakkab me'morchiliklar ba'zi I va II guruh intronlarining bo'lishiga imkon beradi o'z-o'zini qo'shish, ya'ni intron o'z ichiga olgan RNK molekulasi intronni aniq olib tashlash va ekzonlarni to'g'ri tartibda bir-biriga bog'lab qo'yish uchun o'zining kovalent tuzilishini o'zgartirishi mumkin. Ba'zi hollarda, o'zaro bog'laydigan oqsillar biriktirishda qatnashadilar va shu tarzda harakat qiladilar, ular intronga o'z-o'zini biriktirish faoliyati uchun zarur bo'lgan uch o'lchovli tuzilishga katlanishga yordam beradi. I va II guruh intronlari turli xil ichki saqlangan ketma-ketliklar va bukilgan tuzilmalar to'plamlari bilan ajralib turadi va II guruh intronlarini o'z ichiga olgan RNK molekulalarining birlashishi tarvaqaylab intronlar hosil qiladi (splitseozomal RNKlar singari), I guruh intronlarida esa bo'lmagan qo'shimchani boshlash uchun kodlangan guanozin nukleotidi (odatda GTP), uni eksizlangan intronning 5'-uchiga qo'shadi.
Biologik funktsiyalar va evolyutsiya
Intronlar oqsil mahsulotlarini kodlamasa ham, ular gen ekspressionini boshqarishda ajralmas hisoblanadi. Ba'zi intronlarning o'zi ishlab chiqarish uchun biriktirilgandan keyin keyingi ishlov berish orqali funktsional RNKlarni kodlashadi kodlashsiz RNK molekulalar.[25] Shu bilan bir qatorda qo'shilish bitta gendan bir nechta oqsillarni hosil qilish uchun keng qo'llaniladi. Bundan tashqari, ba'zi intronlar genlarni ekspression tartibga solish funktsiyalarining keng doiralarida muhim rol o'ynaydi Bema'ni vositachilik bilan parchalanish[26] va mRNA eksporti.[27]
Intronlarning biologik kelib chiqishi qorong'u. Eukaryotik yadroning oqsillarni kodlovchi genlaridagi intronlarning dastlabki kashfiyotidan so'ng, hozirgi organizmlardagi intronlar umumiy qadimgi ajdoddan meros bo'lib o'tganmi (intronlar - dastlabki gipoteza deb nomlangan) yoki ular paydo bo'lganligi to'g'risida muhim bahslar bo'lib o'tdi. yaqinda evolyutsion jarayonda genlar (intron-kech gipotezasi deb nomlangan). Yana bir nazariya shundan iboratki splitseozoma va genlarning intron-ekzon tuzilishi - bu qoldiq RNK dunyosi (intronlar-birinchi gipoteza).[28] Ushbu farazlarning qaysi biri eng to'g'ri ekanligi haqida hali ham munozaralar davom etmoqda. Ayni paytda mashhur kelishuvga ko'ra, intronlar eukaryot nasabida paydo bo'lgan xudbin unsurlar.[29]
Organizmlarning keng doirasidan olingan genomik DNK ketma-ketliklarini dastlabki tadqiqotlar shuni ko'rsatadiki, turli xil organizmlarda gomologik genlarning intron-ekzon tuzilishi juda xilma-xil bo'lishi mumkin.[30] Hammasi yaqinda olib borilgan tadqiqotlar ökaryotik genomlar endi intronlarning uzunligi va zichligi (intronlar / gen) turdosh turlar orasida sezilarli darajada o'zgarib turishini ko'rsatdi. Masalan, inson genomida o'rtacha 8,4 intron / gen (genomda 139,418) bo'lsa, bir hujayrali qo'ziqorin Ensefalitozoon kunikuli faqat 0,0075 intron / genni o'z ichiga oladi (genomda 15 ta intron).[31] Eukaryotlar umumiy ajdoddan kelib chiqqanligi sababli (umumiy nasl ), evolyutsiya davrida intronlarning katta yutug'i yoki yo'qotilishi bo'lgan bo'lishi kerak.[32][33] Ushbu jarayon tanlanishga bo'ysunadi, deb hisoblashadi, chunki ularning soni kichikroq bo'lganligi sababli yirik turlarda intron daromad olish tendentsiyasi va kichikroq (ayniqsa bir hujayrali) turlarda aksincha.[34] Biologik omillar, shuningdek, genomdagi qaysi genlar intronlarni yo'qotishi yoki to'planishiga ta'sir qiladi.[35][36][37]
Shu bilan bir qatorda qo'shilish Intron eksizyonidan so'ng gen ichidagi ekzonlar bitta gendan tarjima qilingan oqsillar ketma-ketligining katta o'zgaruvchanligini keltirib chiqaradi, bu esa bitta gen va bitta prekursor mRNA transkriptidan ko'plab bog'liq oqsillarni hosil bo'lishiga imkon beradi. Muqobil RNK qo'shilishini boshqarish hujayra ichidagi va hujayradan tashqari signallarning keng doirasiga javob beradigan signal beruvchi molekulalarning murakkab tarmog'i tomonidan amalga oshiriladi.
Intronlar samarali biriktirish uchun muhim bo'lgan bir nechta qisqa ketma-ketliklarni o'z ichiga oladi, masalan intronning har ikki uchida joylashgan aktseptor va donor joylari, shuningdek, to'g'ri biriktirish uchun zarur bo'lgan filial nuqtasi joyi. splitseozoma. Ba'zi intronlar, ular tarkibidagi gen ekspresiyasini kuchaytirishi ma'lum bo'lgan jarayon bilan ma'lum intron vositachilik bilan takomillashtirish (IME).
Viloyatlarning faol ravishda transkripsiyasi DNK tez-tez shakllantiradi R-ko'chadan himoyasiz DNKning shikastlanishi. Yuqori darajada ifoda etilgan xamirturush genlarida intronlar R-tsikl hosil bo'lishini va DNK zararlanishini oldini oladi.[38] Xamirturush va odamlarda o'tkazilgan genom tahlillari shuni ko'rsatdiki, intron o'z ichiga olgan genlar shu kabi ekspressionning intronless genlariga nisbatan R-loop darajasini pasaytirgan va DNK zararini kamaytirgan.[38] R-tsiklga moyil gen ichiga intron kiritilishi ham R-loop shakllanishini bostirishi mumkin rekombinatsiya. Bonnet va boshq. (2017)[38] intronlarning genetik barqarorlikni saqlashdagi funktsiyasi ularning ma'lum joylarda, xususan yuqori darajada ekspresiya qilingan genlarda evolyutsiyasini ta'minlashi mumkin.
Ochlikka moslashish
Intronlarning fizik mavjudligi, oziq moddalarni sezuvchi yo'llarning ribosomal oqsil genlarining intron kuchaygan repressiyasi orqali ochlikka qarshi uyali qarshilikka yordam beradi.[39]
Mobil genetik elementlar sifatida
Intronlar yo'qolishi yoki evolyutsion vaqt davomida qo'lga kiritilishi mumkin, bu ko'plab taqqoslash tadqiqotlari ko'rsatgan ortologik genlar. Keyingi tahlillar natijasida intron yo'qotilishi va yutuq hodisalarining minglab misollari aniqlandi va eukaryotlarning paydo bo'lishi yoki eukaryotik evolyutsiyaning dastlabki bosqichlari intron invaziyasini o'z ichiga olganligi taklif qilindi.[40] Intron yo'qotilishining ikkita aniq mexanizmi, teskari transkriptaz vositasida intron yo'qotilishi (RTMIL) va genomik o'chirishlar aniqlandi va ularning paydo bo'lishi ma'lum.[41] Intron daromadning aniq mexanizmlari, ammo tushunarsiz va bahsli bo'lib qolmoqda. Hozirgacha kamida etti intron yutish mexanizmi haqida xabar berilgan: Intron transpozitsiyasi, transposon qo'shilishi, tandemning genomik ko'payishi, intronning o'tkazilishi, ikki qatorli tanaffusni tiklash paytida intron daromad (DSBR), II guruh intronini kiritish va intronizatsiya. Nazariyada, yaqinda erishilgan intronlarning kelib chiqishini xostlar tomonidan uyg'unlashgan mutatsiyalar yo'qligi sababli aniqlash osonroq bo'lishi kerak, ammo hatto yaqinda olingan intronlar ham yuqorida aytib o'tilgan mexanizmlardan kelib chiqmagan. Shunday qilib, ushbu topilmalar intron yutishning tavsiya etilgan mexanizmlari ko'plab yangi intronlarning mexanistik kelib chiqishini tavsiflay olmaydimi yoki yo'qmi degan savolni tug'diradi, chunki ular intron yutishning aniq mexanizmlari emas yoki yangi, ammo hali kashf qilinmagan yangi jarayonlar mavjud bo'lsa intronlar.[42]
Intron transpozitsiyasida, eng ko'p uchraydigan intron daromad mexanizmi, birlashtirilgan intron ilgari intronisiz holatda o'z mRNKiga yoki boshqa mRNKga qo'shilishni qaytaradi deb o'ylashadi. Ushbu intron o'z ichiga olgan mRNK teskari transkripsiyadan o'tkaziladi va natijada intron o'z ichiga olgan cDNA o'z genomik lokusi bilan to'liq yoki qisman rekombinatsiya orqali intron daromadiga olib kelishi mumkin. Transpozon qo'shimchalari intron yaratilishiga ham olib kelishi mumkin. Bunday qo'shilish transpozonni AGGT qatoriga kiritganda kodlash ketma-ketligini buzmasdan transpozonni intronizatsiya qilishi mumkin, natijada transpozonning har ikki tomonida bu ketma-ketlik takrorlanadi. Ushbu elementlar nima uchun tasodifan yoki transpozon tomonidan biron bir imtiyozli harakatlar tufayli birlashtirilishi hali tushunilmagan. Tandem genomik takrorlanishida, ikkalasi ham AGGTga o'xshash bo'lgan konsensus donor va akseptor qo'shilish joylari o'rtasidagi o'xshashlik tufayli, AGGT ketma-ketligini saqlaydigan ekzonik segmentning tandem genomik takrorlanishi ikki potentsial qo'shilish joyini hosil qiladi. Splitseozoma tomonidan tan olinganida, asl va takrorlangan AGGT orasidagi ketma-ketlik qo'shiladi, natijada genning kodlash ketma-ketligini o'zgartirmasdan intron hosil bo'ladi. Yaqinda tadqiqotchilar Dafniyada erishilgan intronlarning 43% atrofida qisqa muddatli to'g'ridan-to'g'ri takrorlanishlarni aniqlaganda, gomologik bo'lmagan qo'shilish orqali ikki qatorli tanaffusni tiklash intron yutuq manbai sifatida aniqlandi.[42] Ushbu raqamlar statistik jihatdan ahamiyatliligi uchun boshqa organizmlarda takrorlanishlar bilan himoyalangan intronlar soni bilan taqqoslanishi kerak. II guruh intronini kiritish uchun II guruh intronini yadro geniga retrohom qilish so'nggi splitseozomal intron yutuqlarni keltirib chiqarish uchun taklif qilingan.
Parol yoki psevdogen intronga ega bo'lganda intron yutuqqa olib keladi va keyinchalik bu intronni rekombinatsiya orqali singlisi paralogidagi intron bo'lmagan joyga o'tkazadi. Intronizatsiya - bu mutatsiyalar ilgari ekzonik ketma-ketlikdan yangi intronlarni yaratish jarayoni. Shunday qilib, intron yutishning boshqa taklif qilingan mexanizmlaridan farqli o'laroq, ushbu mexanizm yangi intron yaratish uchun DNK qo'shilishi yoki hosil bo'lishini talab qilmaydi.[42]
So'nggi intron daromadning to'g'ridan-to'g'ri dalillarga ega bo'lmagan yagona faraz qilingan mexanizmi - bu in Vivo jonli ravishda namoyish etilganda, gen ekspresiyasini bekor qiladigan II guruh intron qo'shilishidir.[43] Shuning uchun II guruh intronlari splitseozomal intronlarning taxmin qilingan ajdodlari bo'lib, ular o'ziga xos retroelementlar vazifasini bajaradi va endi intronning ortishi uchun javobgar emas.[44][45] Tandemning genomik takrorlanishi - bu in vivo jonli eksperimental dalillarni qo'llab-quvvatlovchi yagona taklif qilingan mexanizm: qisqa intragenik tandemning takrorlanishi yangi intronni oqsillarni kodlovchi genga kiritib, tegishli peptidlar ketma-ketligini o'zgarishsiz qoldirishi mumkin.[46] Ushbu mexanizm, shuningdek, tandemning genomik takrorlanishi intron daromad olishning keng tarqalgan mexanizmi ekanligi haqidagi g'oyani qo'llab-quvvatlovchi keng miqdordagi bilvosita dalillarga ega. Vivo jonli ravishda taklif qilingan boshqa mexanizmlarni, xususan DSBR, intron uzatish va intronizatsiya paytida intron yutuqlarni sinab ko'rish mumkin, ammo bu mexanizmlar ularni intron yutishning haqiqiy mexanizmlari sifatida mustahkamlash uchun in vivo jonli ravishda namoyish etilishi kerak. Keyinchalik genomik tahlillar, ayniqsa populyatsiya darajasida amalga oshirilganda, har bir mexanizmning nisbiy hissasini aniqlab berishi mumkin, ehtimol turli xil turlar orasida intron yutuqlarining turli darajalariga oydinlik kiritishi mumkin bo'lgan turlarga xos tomonlarni aniqlash mumkin.[42]
Shuningdek qarang
Tuzilishi:
Birlashtirish:
Funktsiya
Boshqalar:
Adabiyotlar
- ^ a b Alberts, Bryus (2008). Hujayraning molekulyar biologiyasi. Nyu-York: Garland fani. ISBN 978-0-8153-4105-5.
- ^ Strayer, Lyubert; Berg, Jeremi Mark; Timoczko, Jon L. (2007). Biokimyo. San-Fransisko: W.H. Freeman. ISBN 978-0-7167-6766-4.
- ^ Ghosh, Shampa; Sinha, Jitendra Kumar (2017), "Intron", Vonkda, Jennifer; Shackelford, Todd (tahr.), Hayvonlarni bilish va xulq-atvori ensiklopediyasi, Springer International Publishing, 1-5 betlar, doi:10.1007/978-3-319-47829-6_70-1, ISBN 978-3-319-47829-6
- ^ Tahrirlovchilar, B. D. (2017 yil 6-avgust). "Intron". Biologiya lug'ati. Olingan 1 dekabr 2019.CS1 maint: qo'shimcha matn: mualliflar ro'yxati (havola)
- ^ a b Gilbert, Uolter (1978). "Nima uchun genlar bo'laklarda". Tabiat. 271 (5645): 501. Bibcode:1978 yil Noyabr.271..501G. doi:10.1038 / 271501a0. PMID 622185. S2CID 4216649.
- ^ Kinniburg, Alan; mertz, j; Ross, J. (1978 yil iyul). "Sichqoncha g-globinli xabarchi RNK ning kashfiyotchisi oraliq RNKning ikkita ketma-ketligini o'z ichiga oladi". Hujayra. 14 (3): 681–693. doi:10.1016/0092-8674(78)90251-9. PMID 688388. S2CID 21897383.
- ^ a b Lewin, Benjamin (1987). Genlar (3-nashr). Nyu-York: Vili. 159–179, 386-betlar. ISBN 0-471-83278-2. OCLC 14069165.
- ^ Chow LT, Gelinas RE, Broker TR, Roberts RJ (sentyabr 1977). "Adenovirus 2 messenjeri RNKning 5 'uchidagi ajoyib ketma-ketlik tartibi". Hujayra. 12 (1): 1–8. doi:10.1016/0092-8674(77)90180-5. PMID 902310. S2CID 2099968.
- ^ Berget SM, Mur S, Sharp PA (1977 yil avgust). "Adenovirus 2 ning kechki mRNA ning 5 'uchidagi qo'shilgan segmentlari". Proc. Natl. Akad. Ilmiy ish. AQSH. 74 (8): 3171–5. doi:10.1073 / pnas.74.8.3171. PMC 431482. PMID 269380.
- ^ https://www.nobelprize.org/nobel_prizes/medicine/laureates/1993/press.html
- ^ Tonegava, S .; Maksam, A. M .; Tizard, R .; Bernard, O .; Gilbert, V. (1978 yil 1 mart). "Immunoglobulin yorug'lik zanjirining o'zgaruvchan mintaqasi uchun sichqoncha urug'i-chizig'i genining ketma-ketligi". Milliy fanlar akademiyasi materiallari. 75 (3): 1485–1489. Bibcode:1978PNAS ... 75.1485T. doi:10.1073 / pnas.75.3.1485. ISSN 0027-8424. PMC 411497. PMID 418414.
- ^ Tilg'man, S. M .; Tiemeier, D. C .; Seidman, J. G.; Peterlin, B. M.; Sallivan M.; Mayzel, J. V .; Leder, P. (1978 yil 1-fevral). "Sichqoncha beta-globin genining tuzilish qismida aniqlangan DNKning oraliq ketma-ketligi". Milliy fanlar akademiyasi materiallari. 75 (2): 725–729. Bibcode:1978PNAS ... 75..725T. doi:10.1073 / pnas.75.2.725. ISSN 0027-8424. PMC 411329. PMID 273235.
- ^ Stajich JE, Dietrich FS, Roy SW (2007). "Qo'ziqorin genomlarini qiyosiy genomik tahlili intronga boy ajdodlarni ochib beradi". Genom Biol. 8 (10): R223. doi:10.1186 / gb-2007-8-10-r223. PMC 2246297. PMID 17949488.
- ^ Taanman, Jan-Villem (1999). "Mitokondriyal genom: tuzilishi, transkripsiyasi, tarjimasi va replikatsiyasi". Biochimica et Biofhysica Acta (BBA) - Bioenergetika. 1410 (2): 103–123. doi:10.1016 / s0005-2728 (98) 00161-3. PMID 10076021 - Elsevier Science Direct orqali.
- ^ Tollervey, Devid; Caceres, Xaver F (2000 yil noyabr). "RNKni qayta ishlash marshlari". Hujayra. 103 (5): 703–709. doi:10.1016 / S0092-8674 (00) 00174-4. PMID 11114327.
- ^ Reugels, AM; Kurek, R; Lammermann, U; Bünemann, H (2000 yil fevral). "Heteroxromatik Y xromosomasidagi dyhein geni DhDhc7 (Y) tarkibidagi mega-intronlar Drosophila hydei ning birlamchi spermatotsitlarida ulkan iplar halqalarini keltirib chiqaradi". Genetika. 154 (2): 759–69. PMC 1460963. PMID 10655227. Olingan 12 dekabr 2014.
- ^ Piovesan, Allison; Karakausi, Mariya; Ricci, Marko; Strippoli, Pierluigi; Vitale, Lorenza; Pelleri, Mariya Chiara (2015 yil 1-dekabr). "GeneBase, NCBI Gen ma'lumotlar bazasini tahlil qilish uchun qulay vosita GeneBase orqali minimal eukaryotik intronlarni aniqlash". DNK tadqiqotlari. 22 (6): 495–503. doi:10.1093 / dnares / dsv028. PMC 4675715. PMID 26581719.
- ^ Copertino DW, Hallick RB (1993 yil dekabr). "II va III guruh intrintronlarining intronlari: yadrodan oldingi mRNA intronlari bilan potentsial munosabatlar". Biokimyo tendentsiyalari. Ilmiy ish. 18 (12): 467–71. doi:10.1016 / 0968-0004 (93) 90008-b. PMID 8108859.
- ^ Padgett RA, Grabovski PJ, Konarska MM, Seiler S, Sharp PA (1986). "Xabarchilarning RNK prekursorlarini birlashtirish". Annu. Rev. Biochem. 55: 1119–50. doi:10.1146 / annurev.bi.55.070186.005351. PMID 2943217.
- ^ Guthrie C, Patterson B (1988). "Spliceosomal snRNAs". Annu. Rev. Genet. 22: 387–419. doi:10.1146 / annurev.ge.22.120188.002131. PMID 2977088.
- ^ Greer CL, Peebles CL, Gegenheimer P, Abelson J (fevral 1983). "TRNK qo'shilishida xamirturush RNK ligazasining ta'sir mexanizmi". Hujayra. 32 (2): 537–46. doi:10.1016/0092-8674(83)90473-7. PMID 6297798. S2CID 44978152.
- ^ Reinhold-Hurek B, Shub DA (may 1992). "Keng tarqalgan divergent bakteriyalarning tRNK genlaridagi o'z-o'zini biriktiruvchi intronlar". Tabiat. 357 (6374): 173–6. Bibcode:1992 yil Natura. 357..173R. doi:10.1038 / 357173a0. PMID 1579169. S2CID 4370160.
- ^ Cech TR (1990). "I guruh intronlarining o'z-o'zini biriktirishi". Annu. Rev. Biochem. 59: 543–68. doi:10.1146 / annurev.bi.59.070190.002551. PMID 2197983.
- ^ Mishel F, Ferat JL (1995). "II guruh intronlarining tuzilishi va faoliyati". Annu. Rev. Biochem. 64: 435–61. doi:10.1146 / annurev.bi.64.070195.002251. PMID 7574489.
- ^ Rearick D, Prakash A, McSweeny A, Shepard SS, Fedorova L, Fedorov A (2011 yil mart). "NcRNA ning intronlar bilan tanqidiy aloqasi". Nuklein kislotalari rez. 39 (6): 2357–66. doi:10.1093 / nar / gkq1080. PMC 3064772. PMID 21071396.
- ^ Bicknell AA, Cenik C, Chua HN, Roth FP, Mur MJ (dekabr 2012). "UTRlarda intronlar: nega ularni e'tiborsiz qoldirishimiz kerak". BioEssays. 34 (12): 1025–34. doi:10.1002 / bies.201200073. PMID 23108796. S2CID 5808466.
- ^ Cenik, mumkin; Chua, Xon Nian; Chjan, Xuy; Tarnavskiy, Stefan P.; Akef, Abdalla; Derti, Adnan; Tasan, Murat; Mur, Melissa J.; Palazzo, Aleksandr F.; Rot, Frederik P. (2011). Snayder, Maykl (tahrir). "Genom tahlillari sekretor va mitoxondriyal genlar uchun 5′UTR intronlari va yadroviy mRNA eksporti o'rtasidagi o'zaro bog'liqlikni ochib beradi". PLOS Genetika. 7 (4): e1001366. doi:10.1371 / journal.pgen.1001366. ISSN 1553-7404. PMC 3077370. PMID 21533221.
- ^ Penny D, Hoeppner MP, Poole AM, Jeffares DC (Noyabr 2009). "Intronlar-birinchi nazariyaga umumiy nuqtai". Molekulyar evolyutsiya jurnali. 69 (5): 527–40. Bibcode:2009JMolE..69..527P. doi:10.1007 / s00239-009-9279-5. PMID 19777149. S2CID 22386774.
- ^ Kavalier-Smit, T (1985). "Xudbin DNK va intronlarning kelib chiqishi". Tabiat. 315 (6017): 283–4. Bibcode:1985 yil natur.315..283C. doi:10.1038 / 315283b0. PMID 2987701. S2CID 4367253.
- ^ Rodriges-Trelles F, Tarrio R, Ayala FJ (2006). "Splitseozoma intronlarining kelib chiqishi va rivojlanishi". Annu. Rev. Genet. 40: 47–76. doi:10.1146 / annurev.genet.40.110405.090625. PMID 17094737.
- ^ Mourier T, Jeffares DC (may 2003). "Eukaryotik intron yo'qotilishi". Ilm-fan. 300 (5624): 1393. doi:10.1126 / science.1080559. PMID 12775832. S2CID 7235937.
- ^ Roy SW, Gilbert V (mart 2006). "Splitseozoma intronlarining rivojlanishi: naqshlar, jumboqlar va taraqqiyot". Genetika haqidagi sharhlar. 7 (3): 211–21. doi:10.1038 / nrg1807. PMID 16485020. S2CID 33672491.
- ^ de Souza SJ (2003 yil iyul). "Intron evolyutsiyasining sintetik nazariyasining paydo bo'lishi". Genetika. 118 (2–3): 117–21. doi:10.1023 / A: 1024193323397. PMID 12868602. S2CID 7539892.
- ^ Lynch M (aprel 2002). "Intron evolyutsiya populyatsion-genetik jarayon sifatida". Milliy fanlar akademiyasi materiallari. 99 (9): 6118–23. Bibcode:2002 yil PNAS ... 99.6118L. doi:10.1073 / pnas.092595699. PMC 122912. PMID 11983904.
- ^ Jeffares DC, Mourier T, Penny D (2006 yil yanvar). "Intron yutuq va yo'qotish biologiyasi". Genetika tendentsiyalari. 22 (1): 16–22. doi:10.1016 / j.tig.2005.10.006. PMID 16290250.
- ^ Jeffares DC, Penkett CJ, Bähler J (avgust 2008). "Tez tartibga solinadigan genlar intron kambag'aldir". Genetika tendentsiyalari. 24 (8): 375–8. doi:10.1016 / j.tig.2008.05.006. PMID 18586348.
- ^ Castillo-Devis CI, Mehedov SL, Xartl DL, Koonin EV, Kondrashov FA (avgust 2002). "Yuqori darajada ifodalangan genlarda qisqa intronlar uchun tanlov". Tabiat genetikasi. 31 (4): 415–8. doi:10.1038 / ng940. PMID 12134150. S2CID 9057609.
- ^ a b v Bonnet A, Grosso AR, Elkaoutari A, Coleno E, Presle A, Sridhara SC, Janbon G, Géli V, de Almeyda SF, Palancade B (2017). "Intronlar eukaryotik genomlarni transkripsiyaga bog'liq genetik beqarorlikdan himoya qiladi". Mol. Hujayra. 67 (4): 608-621.e6. doi:10.1016 / j.molcel.2017.07.002. PMID 28757210.
- ^ Parenteau, Julie; Maignon, Laurine; Bertumie, Melodi; Katala, Matyo; Gagnon, Vanessa; Abou Elela, Sherif (2019 yil 16-yanvar). "Intronlar - hujayralarning ochlikka javob berish vositachilari". Tabiat. 565 (7741): 612–617. Bibcode:2019Natur.565..612P. doi:10.1038 / s41586-018-0859-7. ISSN 1476-4687. PMID 30651641. S2CID 58014466.
- ^ Rogozin, I. B.; Karmel, L .; Tsuros, M.; Koonin, E. V. (2012). "Splitseozoma intronlarining kelib chiqishi va rivojlanishi". Biologiya to'g'ridan-to'g'ri. 7: 11. doi:10.1186/1745-6150-7-11. PMC 3488318. PMID 22507701.
- ^ Derr, L. K .; Strathern, J. N. (1993). "Genlarni konversiyalashda teskari transkriptlarning roli". Tabiat. 361 (6408): 170–173. Bibcode:1993 yil 36-iyun. doi:10.1038 / 361170a0. PMID 8380627. S2CID 4364102.
- ^ a b v d Yenerall, P .; Chjou, L. (2012). "Intron daromad mexanizmlarini aniqlash: taraqqiyot va tendentsiyalar". Biologiya to'g'ridan-to'g'ri. 7: 29. doi:10.1186/1745-6150-7-29. PMC 3443670. PMID 22963364.
- ^ Chalamcharla, V. R .; Curcio, M. J .; Belfort, M. (2010). "II guruh intronining yadroviy ifodasi splitseozoma intron ajdodiga mos keladi". Genlar va rivojlanish. 24 (8): 827–836. doi:10.1101 / gad.1905010. PMC 2854396. PMID 20351053.
- ^ Cech, T. R. (1986). "O'z-o'zini biriktiruvchi RNKning umumiyligi: yadro mRNK qo'shilishi bilan bog'liqligi". Hujayra. 44 (2): 207–210. doi:10.1016/0092-8674(86)90751-8. PMID 2417724. S2CID 11652546.
- ^ Dikson, L .; Xuang, H. -R .; Liu, L .; Matsuura M.; Lambovits, A. M.; Perlman, P. S. (2001). "Xamirturush guruhi II intronining retrotranspozitsiyasi to'g'ridan-to'g'ri ektopik DNK joylariga teskari qo'shilish orqali sodir bo'ladi". Milliy fanlar akademiyasi materiallari. 98 (23): 13207–13212. Bibcode:2001 yil PNAS ... 9813207D. doi:10.1073 / pnas.231494498. PMC 60849. PMID 11687644.
- ^ Xellsten, U .; Aspden, J. L .; Rio, D.C .; Roxsar, D. S. (2011). "Segmental genomik takrorlanish funktsional intron hosil qiladi". Tabiat aloqalari. 2: 454–. Bibcode:2011 yil NatCo ... 2..454H. doi:10.1038 / ncomms1461. PMC 3265369. PMID 21878908.
Tashqi havolalar
- NCBI tomonidan belgilangan exon / intron ketma-ketliklarini qidirish mexanizmi
- Bryus Alberts, Aleksandr Jonson, Julian Lyuis, Martin Raff, Kit Roberts va Piter Uolter Hujayraning molekulyar biologiyasi, 2007, ISBN 978-0-8153-4105-5. To'rtinchi nashrni NCBI Bookshelf orqali onlayn ravishda olish mumkin: havola
- Jeremi M Berg, Jon L Timoczko va Lyubert Strayer, Biokimyo 5-nashr, 2002 yil, V H Freeman. Onlaynda NCBI Bookshelf orqali mavjud: havola
- O'simliklar genomik ketma-ketliklari uchun intron topuvchi vosita
- Exon-intron grafik ishlab chiqaruvchisi