Apoptosoma - Apoptosome
The apoptosoma katta to'rtinchi davrdir oqsil jarayonida shakllangan tuzilish apoptoz. Uning shakllanishi ozod etilishi bilan boshlanadi sitoxrom v dan mitoxondriya ichki (ichki) yoki tashqi (tashqi) hujayralarni o'lim stimuliga javoban. Stimullar o'zgarishi mumkin DNKning shikastlanishi va virusli infektsiya a-ning degradatsiyasiga olib keladigan kabi rivojlanish belgilariga turpole's quyruq.
Sutemizuvchilar hujayralarida bir marta sitoxrom v ajralib chiqadi, u sitosolik oqsil bilan bog'lanadi Apaf-1 apoptosoma hosil bo'lishini engillashtirish uchun. Dastlabki biokimyoviy tadqiqotlar apoptosoma hosil bo'lishi uchun sitoxrom c va apaf-1 nisbatlarini ikkitadan bittaga taklif qiladi. Ammo yaqinda o'tkazilgan tizimli tadqiqotlar sitokrom c va apaf-1 nisbati birma-bir ekanligini ko'rsatadi. Shuningdek, nukleotid ekanligi isbotlangan dATP uchinchi komponent apaf-1 bilan bog'langanligi sababli, uning aniq roli hali ham muhokama qilinmoqda. Sutemizuvchilar apoptosomasi hech qachon kristallanmagan edi, ammo odamning APAF-1 / sitoxrom-c apoptosomasi pastki qismida tasvirlangan (2 nm ) kriogen bilan aniqlik uzatish elektron mikroskopi 2002 yilda,[2] 7 barobar simmetriyaga ega geptamerik g'ildirakka o'xshash zarrachani ochib berish. Yaqinda odam apoptosomasining o'rtacha o'lchamlari (9,5 igngstrom) tuzilishi ham hal qilindi kriyo-elektron mikroskopi, bu barcha APAF-1 domenlari (CARD, NBARC va WD40) va sitokrom c pozitsiyalarida aniq xulosa chiqarishga imkon beradi. Hozirgi vaqtda monomerik, faol bo'lmagan Apaf-1 kichik birligining (PDB 3SFZ) kristalli tuzilishi mavjud.[1][3]
Yaratilgandan so'ng, apoptosoma faol bo'lmaganlarni jalb qilishi va faollashtirishi mumkin pro-kaspaz-9. Faollashtirilgandan so'ng, ushbu tashabbuskor kaspaza effektor kaspazlarini faollashtirishi va apoptozga olib keladigan hodisalar kaskadini boshlashi mumkin.
Tarix
Apoptosoma atamasi birinchi bo'lib 1998 yilda Yoshihide Tsujimotoning "Rol Bcl-2 oilaviy oqsillari yilda apoptoz: apoptosomalar yoki mitoxondriya ?".[4] Biroq, Apoptosoma bu vaqtgacha a uchlamchi kompleks. Ushbu kompleks jalb qilingan kaspaz-9 va Bcl-XL har biri o'ziga xos xususiyatga ega Apaf-1 domen. Keyinchalik ushbu kompleksning shakllanishi tartibga soluvchi rol o'ynaydi deb ishonilgan sutemizuvchi hujayralar o'limi.[5] O'sha yilning dekabr oyida "Journal of Biological Chemistry" jurnalida Apas-1 prokaspaza-9 ni faollashtirish orqali apoptozning regulyatori ekanligi haqida yana bir maqola e'lon qilindi.[6]
Apoptosoma mezonlari 1999 yilda ishlab chiqilgan. Birinchidan, u katta kompleks bo'lishi kerak (1,3 million Daltondan katta). Ikkinchidan, uning shakllanishi talab qiladi gidroliz ning yuqori energiya aloqasi ATP yoki dATP. Va nihoyat, u procaspase-9ni funktsional shaklida faollashtirishi kerak. Ushbu kompleksning shakllanishi qaytarilmaslik nuqtasidir va apoptoz paydo bo'ladi. Barqaror APAF-1 va sitoxrom mutimerik kompleks Ushbu tavsifga mos keladi va endi apoptosoma deb nomlanadi.[7]
Apoptosoma a deb o'ylangan mutimerik kompleks ikki sababga ko'ra. Birinchidan, bir nechta procaspase-9 molekulalarini bo'linish uchun bir-biriga yaqinlashtirish. Ikkinchidan, apoptoz eshigini ko'tarish, shuning uchun nonspesifik oqish sitoxrom v apoptozga olib kelmaydi.[7]
Apoptosoma procaspase-9 faollashtiruvchisi sifatida o'rnatilgandan so'ng, ushbu yo'l ichidagi mutatsiyalar muhim tadqiqot maydoniga aylandi. Ba'zi misollarga inson kiradi leykemiya hujayralar, tuxumdon saratoni va virusli infektsiyalar.[8][9][10] Ushbu yo'lning dolzarb tadqiqot yo'nalishlari batafsilroq muhokama qilinadi. Hujayra o'limi uchun ham maxfiy yo'llar mavjud, ular APAF-1 va shuning uchun apoptosomadan mustaqil. Ushbu marshrutlar, shuningdek, kaspaz-3 va 9-dan mustaqil bo'lib, apoptozning yashirin yo'llari sekinroq, ammo keyingi tadqiqotlar davomida foydali bo'lishi mumkin.[11]

(Yuan va boshq. 2010, apoptosoma-procaspase-9 CARD kompleksining tuzilishi[1]
Tuzilishi
Apoptosoma multimolekulyar hisoblanadi holoenzim adapter oqsili atrofida yig'ilgan murakkab Apaf1 (apoptotik proteaz faollashtiruvchi omil 1) ustiga mitoxondriya - vositachilik apoptoz apoptosomaning T shakllanishi ba'zi bir stress signallari bilan rag'batlantirilishi kerak ATP / dATP va sitoxrom v ichida sitozol.[12]Stress qo'zg'atuvchisi sitoxrom c ning sitoplazma ichiga chiqarilishini keltirib chiqarishi mumkin, so'ngra C-terminali bir nechta WD-40 takrorlanishini o'z ichiga olgan mintaqada Apaf-1.[2] The oligomerizatsiya Apaf-1 ning sinxronlashtirilgan yollash bilan birga bo'lishi ko'rinadi procaspase-9 uchun KARTA motifi Apaf-1da N-terminali.[2] Apoptosoma aktivatsiyani ishga tushiradi kaspalar ichida ichki yo'l ning apoptoz.[12]
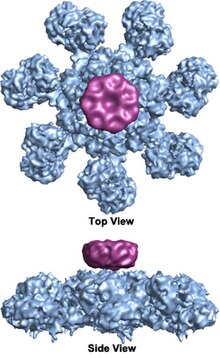
G'ildirak shaklida geptamerik apoptosomaning ettita simmetriya tuzilishiga ega bo'lgan kompleks birinchi marta 27 piksellar sonida aniqlandi elektron kriyomikroskopiyasi texnikasi va hisoblangan massasi taxminan 1 MDa (Acehan va boshq. 2002).[2] Ushbu g'ildirakka o'xshash zarrachaning ettita spiksi va markaziy uyasi bor. Gapiruvchining distal mintaqasi aniq Y shakliga ega.[12] Hub domeni Y domeniga egilgan qo'l bilan bog'langan. Har bir Y domeni ikkita lobni o'z ichiga oladi (katta va kichik), ular orasida sitoxrom v bog'lanish joylari mavjud.[12] Apoptosoma tuzilishining rezolyutsiyasi nisbatan past bo'lganligi sababli, apoptosomani yig'ish uchun ikkita munozarali model taklif qilindi. Bitta model NOD domenlari markaziy markazni va CARD domenlari NOD mintaqasining yuqori qismida erkinroq uzuk hosil qilishini taklif qiladi.[2] Boshqa bir model Apaf-1 ni N-terminalli CARD va nukleotidlarni bog'lash mintaqasi apoptosomaning markaziy markazini tashkil etadigan darajada kengaytirilgan tarzda tashkil qilishni taklif qiladi, 13 WD-40 takrorlanadi ikkita lobni tashkil qiladi.[12] Katta lob ettita takrorlanish va kichik lob oltita takrorlash bilan hosil bo'ladi.[12] Har bir kaspaz-9 molekulasi markaziy markazda CARD domenini bog'lab, gumbaz shaklidagi tuzilmani hosil qiladi.[12] Ushbu tortishuv yaqinda inson apoptosomasi-procaspase-9 CARD kompleksining yuqori aniqlikdagi tuzilishi bilan hal qilindi.[1] Ushbu tuzilma faqat NOD mintaqalari apoptosomaning markaziy markazini tashkil etishini aniq ko'rsatdi (rasmlarga qarang), CARD esa apoptosoma platformasi bilan moslashuvchan bog'lanib, asosiy holat apoptosomasida tartibsiz bo'lib qoladi.[1] Apoptosoma procaspase-9 bilan bog'langandan so'ng, Apaf-1 CARDs va procaspase-9 CARDs platformada o'tirgan diskka o'xshash egiluvchan strukturani hosil qiladi.[1] Soni WD-40 takrorlanadi shuningdek, 13 o'rniga 15 ekanligi isbotlangan,[1] va u 7 ta pichoqli beta-pervaneldan va 8 ta pichoqli beta-pervanadan iborat.[1]
Vang va uning hamkasblaridan olingan dalillar shuni ko'rsatadiki, majmua ichidagi procaspase-9 va Apaf-1 stexiometrik nisbati taxminan 1: 1 ni tashkil qiladi.[7] Bu miqdoriy mass-spektrometriya tahlili bilan yana bir bor isbotlandi.[13] Sitafrom c ning Apaf-1 ga qadar bo'lgan stexiometriyasi 1: 1 ekanligi isbotlangan.[1] Oligomerizatsiyadan so'ng sitokrom c ning apoptosomaga barqaror qo'shilishi zarurmi yoki yo'qligi haqida ba'zi munozaralar mavjud, ammo so'nggi tuzilmaviy ma'lumotlar odamning oligomerik apoptosomasini stabillashtiradi degan fikrni ma'qullaydi.[1] Shu bilan birga, qurtlar va mevali pashshalar singari sutemizuvchilardan tashqari turlarda apoptosomani yig'ish uchun sitoxrom c talab qilinishi mumkin emas.[14] Bundan tashqari, bir nechta boshqa molekulalar, xususan, kaspaza-3, apoptosoma bilan birgalikda tozalanganligi haqida xabar berilgan[7] va kaspaza-3 apoptosoma-procaspase-9 kompleksini bog'lashi mumkinligi isbotlangan.[13]
Apaf-1 apoptosomaning asosini tashkil qiladi. Uning uchta alohida mintaqasi bor: N-terminal kaspaza-yollash domeni (CARD, qoldiqlari 1-90), markaziy nukleotid bilan bog'lovchi va oligomerizatsiya hududi (NB-ARC / NOD, 128-586) va C-terminalli WD40 mintaqasi ( 613–1248) taxminan 140 KDa oqsilni tashkil qiladi.[2]
- CARD domeni Apaf-1 procaspase-9 bilan o'zaro aloqada bo'lib, apoptosoma ichida ishga qabul qilish bilan bog'liq.[2]
- NB-ARC / NOD mintaqasi C. elegans Ced-4 oqsiliga o'xshash ketma-ket o'xshashlikni namoyish etadi.[2]
- Apaf1-ning WD40 mintaqasidagi C-terminalida 15 ta mavjud WD-40 takrorlanadi pervanel shaklidagi ikkita domenga tuzilgan.[1] WD-40 takrorlanishi - bu Trp-Asp bilan tugaydigan va odatda oqsil-oqsilning o'zaro ta'sirida ishtirok etadigan 40 ta aminokislotaning ketma-ketligi.[2]
Konservalangan Walker qutilari A (p-loop 155-161) va B (239-243) o'z ichiga olgan a / b domenlari (NBD) bilan bog'langan qisqa bog'lovchi va nukleotid N-terminal CARD domeniga amal qiladi.[2] Walker qutilari A / B dATP / ATP va Mg2 + ulanishi uchun juda muhimdir.[1][2] NBD-dan keyin kichik spiral domen (HD1), ikkinchi bog'lovchi va konservalangan qanotli spiral domeni (WHD) joylashgan.[2] NOD mintaqasi NBD, HD1 va WHD ni o'z ichiga oladi va ATPase domenini yaratadi AAA + ATPase oilasi.[1][2] NOD va WD-40 takrorlanishlari orasidagi bog'lanishda super spiral domen (HD2) mavjud.[1] WD40 takrorlashlari sakkiz va ettitadan iborat bo'lib, ularni bog'lovchi bilan bog'laydi.[1]

Boshqa organizmlardagi apoptosomalar
Yuqoridagi tavsiflar inson apoptosomasi uchun. Boshqa organizmlarning apoptosomali murakkab tuzilmalari ko'p o'xshashliklarga ega, ammo rasmda ko'rsatilgandek, subbirliklarning kattaligi va soni juda boshqacha. Dark deb nomlangan mevali uchish tizimida 8 ta subbirlik (PDB 4V4L) halqasi mavjud.[14] CED-4 deb nomlangan nematod apoptosomasi oktamerik, ammo ancha kichik (PDB 3LQQ) va u sitoxrom S ni bog'laydigan hududlarni o'z ichiga olmaydi.[15]
Faoliyat mexanizmi
Boshlash
Apoptosoma ta'sirining boshlanishi, ning birinchi qadamlariga to'g'ri keladi dasturlashtirilgan hujayra o'limi (PCD) yo'li. Hayvonlarda apoptoz bo'lishi mumkin katalizlangan ikki yo'lning birida; tashqi yo'l hujayradan tashqari ligandlarning transmembran retseptorlari bilan bog'lanishini o'z ichiga oladi ichki yo'l joy olish mitoxondriya.[16] Ushbu ichki yo'l ozodlikni o'z ichiga oladi sitoxrom S mitoxondriyadan va keyinchalik sitosolik oqsil bilan bog'lanishidan kelib chiqadi Apaf-1.[16][17] Sitokrom c chiqarilishi apoptosoma ta'sirini boshlash uchun zarurdir; ushbu versiya bir necha usul bilan tartibga solinadi, eng muhimi, aniqlash orqali kaltsiy ion darajalar.[16]
Sitoxrom c chiqarilishi
Sitoxrom c chiqarilishi ikki usuldan birida amalga oshirishni taklif qiladi. Birinchidan, o'tkazuvchanlik o'tish teshigi (PTP) mitoxondriya o'limga olib keladigan signalni qabul qilganda va chiqarilganda membranalararo kosmik oqsillar (12). PTP quyidagilardan iborat kuchlanishga bog'liq anion kanali (VDAC), ichki membrana oqsil adenin nukleotid translokatori (AdNT) va matritsa oqsili siklofilin D (CyD) [12]. Ushbu teshik mitoxondriyaning shishishiga va tashqi mitoxondriyal membrana yorilish uchun (Diamond & McCabe, 2007). Ushbu o'zgarish bilan o'tkazuvchanlik, oqsillar sitoxrom c kabi ajralib chiqadi sitozol (12). Ushbu o'zgarish, ehtimol, sabab bo'ladi mitokondriyal o'tkazuvchanlikning o'tishi (MPT), bu erda mitoxondrial transmembran potentsiali qulaydi va ATP ishlab chiqarish to'xtaydi (12). Ushbu usulning farmatsevtika agenti tomonidan inhibatsiyasi siklosporin A (CsA), ikkinchi yo'lni topishga olib boring (13). Sitoxrom v chiqarishning ikkinchi usuli PTP dan mustaqil bo'lib, faqat VDACni o'z ichiga oladi. Bcl-2 oilasining a'zolari pro-apoptotik oqsillar VDACning ochilishini keltirib chiqarishi mumkin (12). Bu esa, interembranalararo kosmik oqsillarning, shu jumladan sitoxrom c ning va keyinchalik MPT ning bir xil tarqalishiga olib keladi (12).
Apaf-1
a. Sitoxrom yo'qligi v
Sitokrom c bo'lmasa, Apaf-1 uning tarkibida mavjud monomerik shakl; deb o'ylashadi WD-40 domeni Apaf-1 ni an holatida ushlab, yana oqsilga o'ralgan holda qoling avtomatik taqiqlangan davlat.[16] Bundan tashqari, bir nechta sohalar shu qadar chambarchas bog'langanki, oqsil boshqa hech narsa bilan bog'lana olmaydi.[16] Bu orqali aniqlandi mass-spektrometriya avtoinhibitlangan yoki "qulflangan" holatda ADP Apaf-1 ning ATPase domeniga bog'langan.[16] Bunday holatda, bu protein yagona va har qanday kishini faollashtirishga qodir emas kaspalar.
b. Sitoxrom v
Sitoxrom c Apaf-1 ning WD-40 domeniga ulanadi.[16] Bu "qulf" ni chiqarishga imkon beradi, ya'ni bu domen endi avtoinhib qilinmaydi.[1][16] Biroq, KARTA va NB-ARC domenlari avtoinhiblangan holatda qoladi.[16] CARD domeni bu qulfdan faqat Apaf-1 (d) ATP / ATP bilan bog'langanda chiqariladi; ATP ulanganda, CARD domeni Caspase-9 bilan bog'lanishiga ruxsat beriladi.[1][16] ADP ATPase domenida bo'lsa, oligomerizatsiya inhibe qilinadi. Shunday qilib, ATP ning bog'lanishi, shuningdek, Apaf-1 ning oligomerizatsiyasini beradi olti burchakli quyi oqimdagi kaspazni faollashtirish uchun zarur bo'lgan tuzilish.[1][7][16] Mutatsiyalar ATPase domen oqsilni harakatsiz qiladi; ammo, ushbu ADP-ATP almashinuvini boshqarish usuli aniq emas.[1][7][16]Oligomerizatsiya faqat 7 ta sitoxrom s molekulalari, 7 ta Apaf-1 oqsillari va etarli (d) ATP / ATP mavjud bo'lganda sodir bo'lishi mumkin.[7] ATPase domeni quyidagilarga tegishli AAA + ATPase oilasi; bu oila boshqa ATPase domenlariga ulanish va hexa- yoki heptamerlarni hosil qilish qobiliyati bilan mashhur.[16] Keyinchalik g'ildirak konstruktsiyasida joylashtirilgan ettita Apaf-1 molekulasi mavjud bo'lganda, apoptosoma faol deb hisoblanadi, bu yo'nalish NB-ARC domenlari markazda joylashgan.[1][16]
Faol apoptosoma harakati
Ushbu funktsional apoptosoma keyinchalik kaspaz 9 platformasini faollashtirishi mumkin.[1][16]Caspase 9 a sifatida mavjud zimogen sitozolda va hujayralardagi 20 nM da topilgan deb o'ylashadi.[16] Faol bo'lish uchun zimogenni ajratish shart emasligi ma'lum bo'lsa ham,[16] procaspase-9 faolligi bir marta bo'linib bo'lgandan keyin sezilarli darajada oshishi mumkin.[13] Birinchi gipoteza shundaki, apoptosoma uchun dimerizatsiya parchalanishdan oldin ikkita kaspaza 9 molekulasidan; bu gipotezani Reidl va Salvasen 2007 yilda qo'llab-quvvatlagan. Ikkinchisi, kaspaz 9 hali ham monomerik shaklda bo'lsa, bo'linish sodir bo'ladi.[13][16] Har holda, kaspaz 9 faollashuvi to'liq kaspaz kaskadining faollashishiga va keyinchalik hujayralar o'limiga olib keladi. Kaspaza kaskadini faollashtiradigan multimerik oqsil kompleksining evolyutsion sababi sitoxrom c ning tasodifan apoptozga olib kelmasligini ta'minlashdir.[7]
Tadqiqot yo'nalishlari
Mutatsiyalar paydo bo'lganda nima bo'ladi?
Tana tabiiy ishlashi uchun apoptoz zarur bo'lsa, apoptosoma yo'lining mutatsiyalari tanadagi katastrofik ta'sirlarni va o'zgarishlarni keltirib chiqaradi. Hujayra yo'lining mutatsiyalari hujayra o'limini kuchaytirishi yoki tanadagi juda ko'p miqdordagi kasallikni keltirib chiqaradigan hujayralarni o'lishiga yo'l qo'ymasligi mumkin. Mutatsiyaga uchragan apoptoz yo'llari juda ko'p va apoptosoma faolligi yo'qligi sababli saraton kasalligidan kelib chiqadi, Altsgeymer juda ko'p apoptosoma faolligi tufayli kasallik va boshqa ko'plab neyrodejenerativ kasalliklar Parkinson kasallik va Xantington kasalligi.[18] Altsgeymer, Parkinson va Xantington singari neyrodejenerativ kasalliklar yoshga bog'liq kasalliklar bo'lib, hujayralar o'lib ketadigan apoptozning ko'payishini o'z ichiga oladi, ular hali ham ishlashga qodir yoki to'qimalarning ishlashiga yordam beradi. Apaf-1-ALT - bu prostata bezi saratonida topilgan, 339-1248 qoldiqlari bo'lmagan Apaf-1 mutantidir. Apoptosomaning so'nggi tizimli tadqiqotlari shuni isbotlaydiki, Apaf-1-ALT apoptosoma hosil qila olmaydi, chunki u yig'ilish uchun asosiy tarkibiy qismlarni o'tkazib yuboradi.[1]
Saraton kasalligini keltirib chiqaradigan Apoptozning repressiyasi
Hujayra ichidagi genetik va biokimyoviy anormalliklar, odatda tanani tartibsiz hujayralar faoliyati va rivojlanishidan xalos qilish uchun dasturlashtirilgan hujayralarni o'limiga olib keladi; ammo saraton hujayralari mutatsiyalarga ega bo'lib, ular apoptozni bostirishga va omon qolishga imkon beradi. Ushbu repressiya qilingan PCD yo'llarini normal PCD ni targ'ib qilish uchun giper stimulyatsiya orqali faollashtirish uchun ionlashtiruvchi nurlanish kabi kimyoviy terapiya ishlab chiqilgan.[19]
Apoptozdagi P53 mutatsiyalari
P53 o'sma vazifasini bajaradi supressor saratonni oldini olishda ishtirok etadigan va tabiiy ravishda apoptotik yo'llarda paydo bo'ladigan. P53 hujayralarni apoptozga olib keladi va hujayraning keyingi bo'linishini buzadi, shuning uchun bu hujayraning saraton kasalligini oldini oladi (16). Saraton kasalligining aksariyat qismida bu mutatsiyaga uchragan p53 yo'lidir, natijada disfunktsional hujayralarni to'xtatish qobiliyati yo'q. P53 funktsiyasi, shuningdek, p53 genining mutatsiyalari uzoq umr ko'radigan hayvonlarni hosil qiluvchi dominant-salbiy shakllarning namoyon bo'lishiga olib keladigan cheklangan umr uchun javobgar bo'lishi mumkin. Masalan, eksperimentda C. elegans, p53 mutantlarining umr ko'rish davomiyligi avtofagiyaga bog'liq ekanligi aniqlandi.[19] Boshqa bir tajribada Drozofiliya p53 mutatsiyasi kattalar hayotiga ijobiy va salbiy ta'sir ko'rsatdi, bu jinsiy farqlash, PCD va qarish o'rtasidagi bog'liqlikni keltirib chiqardi.[19] P53 ning hayot davomiyligiga qanday ta'sir qilishini aniqlash kelajakdagi tadqiqotlar uchun muhim yo'nalish bo'ladi.
Saratonni davolash uchun apoptozomani maqsad qilish
Apoptozni inhibe qilish saraton kasalligining asosiy xususiyatlaridan biridir, shuning uchun apoptozomani hosil qilish va kaspazalarni faollashtirish uchun ushbu inhibatsiyani boshqarish va engish usullarini topish yangi saratonni davolashda muhim ahamiyatga ega.[20] To'g'ridan-to'g'ri apoptosoma faollashuvini keltirib chiqarish qobiliyati saratonni davolashda juda muhimdir, chunki yuqtirilgan saraton genlari yo'q qilinmaydi, saratonning davomi hosil bo'ladi. Apoptozomani tashqi stimul bilan faollashtirish orqali apoptoz paydo bo'lishi va mutatsiyaga uchragan hujayralardan xalos bo'lishi mumkin. Hozirda bunga erishish uchun ko'plab yondashuvlar, jumladan, rekombinant biomolekulalar, antisens strategiyalar, gen terapiyasi va klassik organik vositalar qo'llanilmoqda. kombinatorial kimyo inson kasalliklarida ortiqcha yoki etishmayotgan hujayralar o'limini to'g'irlash bo'yicha yondashuvda o'ziga xos apoptotik regulyatorlarni maqsad qilish.[18]
Umuman olganda anti-apoptotik oqsillarni yuqori darajada tartibga solish apoptozning oldini olishga olib keladi, uni inhibitorlar hal qilishi mumkin, antipoptotik oqsillarni pastga tushishi esa apoptozning induktsiyasiga olib keladi, bu esa ularni bog'lab turadigan va o'zgartirishga qodir aktivatorlar tomonidan qaytariladi. faoliyat. Apoptozga asoslangan davolashda muhim maqsad molekulasi Bcl-2 dori dizayni uchun.[18] Bcl-2 saratonni inhibe qiluvchi apoptozni keltirib chiqaradigan birinchi onkogen edi. U o'smalarda ifodalangan va kimyoviy terapiyaga chidamli.[18] Olimlar, Bcl-2 anti-apoptotik oqsillar bilan biriktiruvchi depressorlarni inhibe qiladi va to'g'ridan-to'g'ri faollashtiruvchilar bilan ta'sir o'tkazish uchun erkin qoldiradi. Bax va Bak.[18]
Saratonni davolash uchun mo'ljallangan boshqa molekulaga kaspazlar oilasi va ularning regulyatorlari kiradi. Kaspaza faolligining inhibatsiyasi neyrodejenerativ kasalliklar, qon tomirlari, yurak xuruji va jigar shikastlanishlari, shu jumladan inson kasalliklarida hujayralar o'limini bloklaydi. Shuning uchun kaspaz inhibitörleri qon tomir va boshqa inson kasalliklarini davolashni ta'minlaydigan istiqbolli farmakologik vositadir. Hozirgi vaqtda klinikgacha bo'lgan ba'zi bir kaspaz inhibitörleri mavjud, ular ba'zi bir neyrodejenerativ kasalliklarning ta'sirini qaytarish haqida umidvor dalillarni ko'rsatmoqdalar. Yaqinda o'tkazilgan bir tadqiqotda tadqiqotchilar qayta tiklanadigan narsani ishlab chiqdilar kaspaz-3 inhibitori M-826 deb nomlangan va uni sichqoncha modelida sinab ko'rgan, bu erda miya to'qimalariga zarar yetgan. Bundan tashqari, u Xantington kasalligi bilan sichqonchada sinovdan o'tkazildi va inhibitor bu kaspaz inhibitori bo'yicha keyingi tadqiqotlar uchun istiqbolli ta'sirlarni ko'rsatadigan striatlangan neyron o'limining oldini oldi.[18]
Apoptosoma kompleksi molekulyar terapiya uchun yangi potentsial maqsadlarni ochib berdi
Apaf1 / caspase-9 apoptosoma shakllanishi apoptotik kaskadda hal qiluvchi voqea hisoblanadi. Faol apoptosoma kompleksining shakllanishiga to'sqinlik qiladigan yoki barqarorlashtiradigan yangi potentsial dori-darmonlarni aniqlash juda yoki etarli bo'lmagan apoptoz bilan tavsiflangan kasallikni davolash uchun ideal strategiyadir.[18] Yaqinda taurin mitoxondriyal disfunktsiyani oldini olmasdan Apaf1 / caspase-9 apoptosoma shakllanishini inhibe qilish qobiliyati tufayli kardiyomiyotsitlarda ishemiya keltirib chiqaradigan apoptozning oldini oladi. Taurin apoptosoma hosil bo'lishini inhibe qilishi mumkin bo'lgan mexanizm, apoptosomaning asosiy tarkibiy qismi bo'lgan kaspaz-9 ekspresiyasini kamaytirishga qodir ekanligi aniqlandi. Shu bilan birga, Aparf1 va caspase-9 ning apoptosomadan tashqari mustaqil rollarga ega ekanligini ko'rsatadigan tadqiqotlar mavjud, shuning uchun ularning darajasini o'zgartirish hujayra funktsiyasini ham o'zgartirishi mumkin. Shunday qilib, eksperimental ma'lumotlarning rag'batlantirilishiga qaramay, bir nechta muammolar echimini topmayapti va klinik amaliyotda eksperimental dorilarni qo'llashni cheklaydi.[18]
Apoptosoma inhibitörlerinin kashf etilishi apoptoz vositachiligidagi kasallikni davolash uchun yangi terapevtik vositani beradi. Apoptosoma tarkibiy qismlarining transkripsiyaviy darajasini o'zgartirmasdan hujayra ichidagi oqsil-oqsil ta'sirida ta'sir o'tkazib, apoptosoma barqarorligini va faolligini inhibe qila oladigan yangi birikmalar alohida ahamiyatga ega.[18] Apoptosomaning so'nggi tizimli tadqiqotlari apoptosomaga asoslangan terapiyani loyihalash uchun qimmatli vositalarni taqdim etishi mumkin.[1][13]
Shuningdek qarang
Adabiyotlar
- ^ a b v d e f g h men j k l m n o p q r s t siz v w Yuan S, Yu X, Topf M, Ludtke SJ, Vang X, Akey CW. "Apoptosoma-procaspase-9 CARD kompleksining tuzilishi." Tuzilishi. 2010 yil may; 18 (5): 571-83.
- ^ a b v d e f g h men j k l m Acehan D, Jiang X, Morgan DG, Heuser JE, Van X, Akey CW (fevral 2002). "Apoptosomaning uch o'lchovli tuzilishi: montaj, prokaspaza-9 ni bog'lash va faollashtirish uchun ta'sirlari". Molekulyar hujayra. 9 (2): 423–32. doi:10.1016 / s1097-2765 (02) 00442-2. PMID 11864614.
- ^ T. F. Reubold; S. Vohlgemut; S. Eshenburg (2011). "To'liq uzunlikdagi Apaf-1 kristalli tuzilishi: o'lim signalining apoptoz mitoxondriyal yo'lida qanday o'tishi". Tuzilishi. 19 (8): 1074–1083. doi:10.1016 / j.str.2011.05.013. PMID 21827944.
- ^ Tsujimoto Y (1998 yil noyabr). "Bcl-2 oilaviy oqsillarining apoptozdagi o'rni: apoptosomalarmi yoki mitoxondriyami?". Hujayralar uchun genlar. 3 (11): 697–707. doi:10.1046 / j.1365-2443.1998.00223.x. PMID 9990505.
- ^ Pan, G.; O'Rourke, K .; Diksit, V. M. (1998). "Caspase-9, Bcl-XL va Apaf-1 uchlamchi kompleks hosil qiladi". Biologik kimyo jurnali. 273 (10): 5841–845. doi:10.1074 / jbc.273.10.5841. PMID 9488720.
- ^ Xu, Ding; Spenser; Nunez (1998). "WD-40 takroriy mintaqasi" Apaf-1 "o'zini o'zi birlashtirishi va" Prokaspaza-9 "ni faollashtirishni tartibga soladi". Biologik kimyo jurnali. 273 (50): 33489–3494. doi:10.1074 / jbc.273.50.33489. PMID 9837928.
- ^ a b v d e f g h Zou H, Li Y, Liu X, Van X (1999 yil aprel). "APAF-1. sitoxrom c multimerik kompleksi - bu prokaspaza-9 ni faollashtiradigan funktsional apoptosoma". Biologik kimyo jurnali. 274 (17): 11549–56. doi:10.1074 / jbc.274.17.11549. PMID 10206961.
- ^ Jia L, Srinivasula SM, Liu FT, Newland AC, Fernandes-Alnemri T, Alnemri ES, Kelsey SM (iyul 2001). "Apaf-1 oqsilining etishmasligi odamning leykemik hujayralarida sitoxrom c ga bog'liq apoptozga qarshilik ko'rsatadi". Qon. 98 (2): 414–21. doi:10.1182 / qon.v98.2.414. PMID 11435311.
- ^ Bitzer M, Armeanu S, Prinz F, Ungerechts G, Wybranietz V, Spiegel M, Bernlöhr C, Cecconi F, Gregor M, Neubert WJ, Schulze-Osthoff K, Lauer UM (avgust 2002). "Sendai virusi yuqtirilgan hujayralarida Caspase-8 va Apaf-1 mustaqil kaspaz-9 faollashuvi". Biologik kimyo jurnali. 277 (33): 29817–9824. doi:10.1074 / jbc.M111898200. PMID 12021264.
- ^ Wolf BB, Schuler M, Li V, Eggers-Sedlet B, Lee V, Tailor P, Fitzgerald P, Mills GB, Green DR (sentyabr 2001). "Apoptotik proteaz faollashtiruvchi omil-1 faolligi kamayganligi yoki yo'qligi sababli tuxumdon saraton hujayralari sathida sitokrom c-ga bog'liq kaspaz faollashuvi nuqsonli". Biologik kimyo jurnali. 276 (36): 34244–51. doi:10.1074 / jbc.M011778200. PMID 11429402.
- ^ Belmoxar CA, Hillion J, Dudognon C, Fiorentino S, Flexor M, Lanotte M, Segal-Bendirdjian E (Avgust 2003). "Apoptoz uchun apoptozomadan mustaqil yo'l. APAF-1 nuqsonlari va biologik natijalarini biokimyoviy tahlil qilish". Biologik kimyo jurnali. 278 (32): 29571–80. doi:10.1074 / jbc.M302924200. PMID 12773531.
- ^ a b v d e f g Ceconi F, Ferraro E, Fuoco C, Strappazzon F (2010). "Apoptosoma tuzilishi va regulyatsiyasi". Apoptosoma: 27–39. doi:10.1007/978-90-481-3415-1_2. ISBN 978-90-481-3414-4.
- ^ a b v d e Yuan S, Yu X, Asara JM, Heuser JE, Ludtke SJ va Akey CW. "Holo-Apoptosoma: Prokaspaza-9 ning faollashishi va Caspase-3 bilan o'zaro ta'siri." Tuzilishi. 2011 yil avgust; 19 (8): 1084-1096.
- ^ a b Yuan S, Yu X, Topf M, Dorstyn L, Kumar S, Ludtke SJ va Akey CW. "Drosophila apoptosomasining tuzilishi 6.9 Å piksellar sonida." Tuzilishi. 2011 yil yanvar; 19 (1): 128-140.
- ^ S. Qi; Y. Pang; Q. Xu; Q. Lyu; H. Li; Y. Chjou; T. U; Q. Liang; Y. Lyu; X. Yuan; G. Luo; H. Li; J. Vang; N. Yan; Y. Shi (2010). "Caenorhabditis elegans apoptosomasining kristalli tuzilishida CED-4 ning oktamerik birikmasi aniqlanadi". Hujayra. 141 (3): 446–457. doi:10.1016 / j.cell.2010.03.017. PMID 20434985.
- ^ a b v d e f g h men j k l m n o p q r Riedl SJ, Salvesen GS (2007 yil may). "Apoptosoma: hujayra o'limining signal beruvchi platformasi". Tabiat sharhlari. Molekulyar hujayra biologiyasi. 8 (5): 405–13. doi:10.1038 / nrm2153. PMID 17377525.
- ^ Diamond & McCabe (2007). Mitoxondriya va o'simlik dasturlashtirilgan hujayralar o'limi. In: Logan DC (Ed.), O'simlik mitoxondriyasi. Yillik o'simliklarni ko'rib chiqish 2007 yil, 31: 308-334.
- ^ a b v d e f g h men D'Amelio M, Tino E, Cecconi F (2008 yil aprel). "Apoptosoma: paydo bo'layotgan tushunchalar va giyohvand moddalarni ishlab chiqarishda yangi potentsial maqsadlar". Farmatsevtika tadqiqotlari. 25 (4): 740–51. doi:10.1007 / s11095-007-9396-z. PMC 2279152. PMID 17674158.
- ^ a b v Shen J, Minora J (2009 yil dekabr). "Qarishdagi hujayralar o'limi va apoptoz va umr ko'rishning regulyatsiyasi". Kashfiyot tibbiyoti. 8 (43): 223–6. PMID 20040274.
- ^ Fischer U, Yanssen K, Schulze-Osthoff K (2007). "Apoptozga asoslangan zamonaviy terapiya: saraton kasalligiga qarshi davo?". BioDrugs: Klinik immunoterapiya, biofarmatsevtika va gen terapiyasi. 21 (5): 273–97. doi:10.2165/00063030-200721050-00001. PMID 17896835.