Gorgonopsiya - Gorgonopsia
| Gorgonopsianlar | |
|---|---|
 | |
| Skeletlari topildi Inostrancevia alexandri da Scienze musiqiy musiqasi, Trento, Italiya | |
| Ilmiy tasnif | |
| Qirollik: | Animalia |
| Filum: | Chordata |
| Klade: | Terapsida |
| Klade: | Neoterapsida |
| Klade: | Theriodontia Sili, 1895 |
| Klade: | †Gorgonopsiya Lydekker, 1890 |
| Genera | |
Gorgonopsiya (dan Qadimgi yunoncha γώνorγώνma'nosi "gorgon ", va ὄψ, "aspekt" ma'nosini anglatadi) yo'q bo'lib ketgan qoplama erta shamshir tishli terapevtiklar dan O'rta ga Yuqori Permiya. Ular uzun va tor bosh suyagi bilan ajralib turadi, shuningdek, yuqori va ba'zan pastki qism bilan o'ralgan it tishlari va tish kesuvchi qurol kesuvchi va pichoqlangan qurol sifatida ishlatilgan. Postkanin tishlari odatda kamayadi yoki yo'q. Katta o'ljani ovlash uchun, ular, ehtimol, pistirma va chekinish taktikasidan foydalanib, pistirma va zaiflashtiruvchi luqmani nishondan chiqarib olishdi va jarohati tugamasdan uni xavfsiz masofada kuzatib borishdi, natijada gorgonopsian hayvonni tortib olib, luqmani o'ldirish. Ular, ehtimol jag'ni echmasdan, 90 ° dan yuqori bo'lgan aql bovar qilmaydigan bo'yniga ega bo'lishlari kerak edi.
Vaqt o'tishi bilan ular hajmi kattalashib, O'rta Permiyadagi bosh suyagining 10-15 sm (4-6 dyuym) uzunligidan o'sib bordi. ayiq - Yuqori Permiyadagi 60 sm (2 fut) gacha bo'lgan nisbatlarga o'xshash. So'nggi gorgonopsians, Rubidjinalar, eng ko'p edi mustahkam guruhga kiradi va ayniqsa kuchli chaqishi mumkin. Ular to'liq bo'lgan deb o'ylashadi quruqlik va zamonaviy singari yer usti lokomotivlari diapazoni bilan yarim tik yurish bilan yurish mumkin edi timsohlar. Ular yirtqich narsalariga qaraganda epchilroq bo'lishlari mumkin, ammo inertsional bo'lishgan gomeotermalar dan ko'ra endotermlar zamonaviy sifatida trosefaliklar va sinodontlar va, ehtimol, nisbatan kam faol bo'lgan. Gorgonopsiyachilar tana haroratini ancha yuqori darajada ushlab tursalar-da, ularning mo'ynasi ham bo'ladimi, aniq emas, ter bezlari, yoki mo'ylovlar. Ularda umuman ko'proq narsa bor edi sudralib yuruvchi dan sutemizuvchi miya. Aksariyat turlar asosan bo'lishi mumkin kunduzgi, kun davomida faol, garchi ba'zilari bo'lishi mumkin edi krepuskulyar yoki tungi. Ularda bo'lgan deb o'ylashadi binokulyar ko'rish, a parietal ko'z, o'tkir hid hissi, funktsional vomeronazal organ ("Jakobsonning organi"), va ehtimol ibtidoiy quloq pardasi.
Asosiy terapevtik guruhlarning barchasi 265 million yil oldin "pelikozavr 'ajdod (barchasini o'z ichiga olgan aniqlanmagan guruh sinapsidlar terapevtik bo'lmagan). Pelikozavrlardan terapiyani qabul qilish O'rta Permiya tomonidan amalga oshirildi, chunki dunyo tobora quruqlashib bormoqda. Yo'qolib ketganidan so'ng, gorgonopsiyaliklar atrof-muhitning cho'qqisi yirtqichlariga aylanishdi dinosefaliklar va O'rta Permiydan keyin yirik terosefaliyalar. Permiya davrida bitta qit'aning mavjudligiga qaramay, Pangaeya, gorgonopsians faqat topilgan Karoo Supergroup (birinchi navbatda Janubiy Afrikada, shuningdek Tanzaniya, Zambiyada va Malavida), Moradi shakllanishi Niger va g'arbiy Rossiyaning, ehtimol Hindistondan ma'lum bo'lgan qoldiqlari bilan. Bu joylar mavsumiy yomg'ir yog'adigan yarim quruq hududlar bo'lgan. Gorgonopsian taksonlari juda oz farq qiladi va shuning uchun ko'plab turlar 19-asrning oxirlarida kashf etilganidan beri ular mayda va juda o'zgaruvchan farqlarga asoslanib nomlangan va qoplama bir necha taksonomik qayta ko'rib chiqilgan
Ular bir bosqichda yo'q bo'lib ketishdi Permiy-trias davridagi yo'q bo'lib ketish hodisasi Permianing oxirida sodir bo'lgan bo'lib, unda katta vulkanik faollik (hosil bo'ladigan) Sibir tuzoqlari ) va natijada katta boshoq paydo bo'ldi issiqxona gazlari harorat ko'tarilishi tufayli tez quruqlikka olib keldi, kislotali yomg'ir, tez-tez o'rmon yong'inlari va potentsial buzilishi ozon qatlami. Voqeadan omon qolgan yagona terapsid guruhi oxir-oqibat sutemizuvchilarga aylanib boradigan sinodontlar edi. Katta yirtqich nişler tomonidan qabul qilingan bo'lar edi arxhosaurs (ya'ni timsohlar va dinozavrlar ) ichida Mezozoy.
Tavsif

Anatomiya gorgonospianlar orasida juda oz farq qiladi.[1] Ko'pgina turlar noaniq mutanosiblik farqlari bilan ajralib turadi va shuning uchun kichikroq turlar aslida katta taksonlarning balog'at yoshiga yetmagan bolalarini aks ettirishi mumkin. Ta'kidlash joizki, qusish tumshug'ining uchida turlar orasida uning kengayish darajasi, shuningdek, 3 tizmaning joylashishi, tarqalish darajasi va shakli jihatidan farq qiladi.[2] Ular odatda uzun va tor bosh suyagiga ega.[3] Voyaga etmagan Rubidiya tumshug'i uzunroq bo'lganiga o'xshaydi.[4]

Goronopsian miyasi, boshqasutemizuvchi shakl terapevtiklar, ning kengayishi yo'q neokorteks, oldingi miyaga nisbatan katta orqa miyaga ega, katta epifizik asab (bilan mavjudotlarda mavjud parietal ko'z boshning tepasida), kattalashtirilgan gipofiz va umumiy cho'zilgan shakli; a-ga o'xshash narsa sudralib yuruvchi miya.[5] The braincase Bundan tashqari, sudralib yuruvchi, nisbatan kichikroq va unchalik qalin bo'lmagan.[6]:280 The flokulyus, lob serebellum, mutanosib ravishda katta va bilan bog'liq vestibulo-okulyar refleks (boshni siljitish paytida stablise nigohi). Ning yo'nalishiga qarab yarim dumaloq kanallar GPIT / RE / 7124 gorgonopsiya namunasining boshi (quloqda (erga parallel ravishda yo'naltirilgan bo'lishi kerak)) taxminan 41 ° ga egilib, 2 ko'zning ko'rish maydonlari orasidagi o'zaro to'qnashuvni oshirib, yaxshilaydi binokulyar ko'rish yirtqich uchun foydalidir. Yoki sudralib yuruvchilar yoki sutemizuvchilar, Yarim dumaloq kanallar tekis bo'lib, ehtimol ular orasida joylashgan edi opistotik (an ichki quloq suyak) va supraoksipital suyaklar.[5]
Boshqa perm terapevtlari singari, gorgonopsiyalar ham sutemizuvchilarning bir nechta xususiyatlarini, shu jumladan parazagital yurishni (oyoq-qo'llari vertikal yo'naltirilgan va umurtqa pog'onasiga parallel ravishda harakatlanadigan) rivojlangan. amfibiyalar va oldingi sinapsidlar, natijada quyruq kattaligi va falanj formulasi kamayadi[7] (sudralib yuruvchilarga o'xshash 2.3.4.5.3 bo'lgan bitta raqamga bo'g'inlar soni[3]), tolali lamellar kortikal suyak, a vaqtinchalik fenestra (teshik vaqtinchalik suyak ), chuqur o'rnatilgan tish va ikkilamchi tanglay (bu og'izni burun bo'shlig'idan ajratib turadi, ammo gorgonopsiya bunday bo'lmasligi mumkin).[7]

Ko'p sutemizuvchilar singari, gorgonopsiya ham bo'lgan heterodontlar, aniq belgilangan tish kesuvchi, itlar va postkanin tishlari gomologik bilan premolar va tishlar.[2] Ularning yuqori jag'ida 5 tishli tish bor edi (ko'pchilik uchun dastlabki 3 bir-birining o'lchamiga teng edi, oxirgi 2 qisqaroq edi) va pastki qismida 4 ta.[6]:17–18 Aksariyat gorgonopsiyalarda tish kesuvchi tishlar katta bo'lgan va yuqori itlar, xuddi keyingi davrlar singari qirg'oqqa cho'zilgan. qichitqi tishli mushuklar. Ba'zi gorgonopsianlar juda uzun bo'yli yuqori tishlarga ega edilar, masalan Inostransviya Va ularning ba'zilari pastki jag 'qismida og'iz yopilganda itning uchini qoplash uchun gardish bor edi. Qilichlar odatda pichoqlash yoki kesuvchi qurol sifatida ishlatilgan deb talqin etiladi, bu esa juda keng to'siqni talab qiladi. Ikkala yuqori va pastki tish itlari Rubidiya sho'ng'idi va hayvon bundan ham kattaroq bo'yniga muhtoj bo'lar edi.[8] Postkanin tishlari esa kattaligi va soni bo'yicha qisqartirildi; ko'p rubidjinlar (so'nggi gorgonopsianlar) pastki jagida postkaninlar bo'lmagan,[9] va Klelandiya ularga to'liq etishmadi.[4]
Gorgonopsians edi polifiodontlar va tishlar inson hayoti davomida doimiy ravishda o'sib bordi.[10] Ba'zi terapevtiklar singari, 1 ta funktsional it bor edi, muqarrar ravishda uzilib qolganida, uni o'rnini bosadigan boshqa bir it o'sayotgan edi. Jag'larning chap va o'ng tomonlari sinxron bo'lishi shart emas edi, shuning uchun, masalan, chap tomondagi 1-chi it o'ngdagi 1-chi tish o'sib ulgurgan paytda funktsional bo'lishi mumkin edi.[10] Bunday usul har doim funktsional itlar to'plamiga ega bo'lishi uchun o'ynab yurgan bo'lishi mumkin, chunki bitta yoki bitta bo'lmagan itlarga ega bo'lish ovga katta to'sqinlik qilishi mumkin va bunday katta tishlarni o'stirish uzoq vaqt talab qilishi mumkin. Boshqa tomondan, funktsional it odatda eng oldingi tish uyasida joylashganligi sababli (har ikkala rozetkada teng ravishda paydo bo'lish o'rniga), itni almashtirish sonli marta sodir bo'lishi mumkin va hayvon oxir-oqibat ushbu rozetkalarda bitta, doimiy ishlaydigan kinologiya to'plami. 1984 yilda ingliz paleontologlari Doris va Kennet Kermak itlar bosh suyagi kattaligiga mos ravishda o'sgan va hayvon o'sishi to'xtamaguncha uzilib qolgan va gorgonopsiyachilar ko'plab sutemizuvchilarda paydo bo'lgan tishlarni almashtirishning dastlabki versiyasini namoyish etishgan. Boshqa tishlarning tishlarini almashtirish naqshlari aniq emas.[8] Postkanin tishlari boshqa tishlarga qaraganda sekinroq almashtirildi, ehtimol ularning funktsional ahamiyati yo'q edi.[2]

Ilgari O'rta Permiyadagi gorgonopsidlar juda kichik bo'lgan, ularning bosh suyagi uzunligi 10-15 sm (4-6 dyuym) bo'lgan, keyinchalik ba'zi bir avlodlar uzunligi 60 sm (2 fut) gacha bo'lgan bosh suyaklari bilan katta, ayiqsimon o'lchamlarga ega bo'lishgan. Shunga qaramay, kichik gorgonopsiyachilar yo'q bo'lib ketguncha ko'p bo'lib qolishdi (garchi kichik turlar aslida boshqa taksonlarning balog'atga etmagan namunalarini aks ettirsa ham).[2]
Evteriodontlardan farqli o'laroq oksipital suyak (bosh suyagining orqa qismida) uchburchakdan farqli o'laroq to'rtburchaklar (balanddan kengroq) va konkav.[6]:279 7 bachadon bo'yni umurtqalari (bo'ynida) barchasi bir-birining o'lchamiga teng, oxirgisi bundan mustasno, qisqaroq va pastroq; 1 bor atlas va 1 o'qi. Qo'rqinchli tishli mushuklar singari, bo'yin yaxshi rivojlangan mushaklari bilan uzoqlashadi, bu ayniqsa, itlar hayvonga cho'ktirilganda foydalidir. Boshqa erta sinapsidlar singari, gorgonopsiyalarda ham bitta oksipital kondil va bachadon bo'yni umurtqasining artikulyatsiyasi (bo'g'imlari) umumiy sudraluvchi bo'lib, boshning yonma-yon harakatlanishiga imkon beradi, lekin yuqoriga va pastga harakatlarini cheklaydi. Oxirgi bachadon bo'yni shunga o'xshash shaklga ega orqa umurtqalari. Dorsallar g'altak shaklida bo'lib, barchasi bir-biriga o'xshash ko'rinadi. Spinous jarayonlar tsentradan keskin chiqib ketadi va old va orqa tomonlarida o'tkir keellar mavjud. Evteriodontlardan farqli o'laroq, gorgonopsiyalar farq qilmaganlar bel umurtqalari. Shunga qaramay, ushbu ketma-ketlikka teng keladigan dorsallar tik yo'naltirilgan zigopofazalarga ega bo'lgan qichitqi tishli mushuklarning lumbarlariga o'xshaydi, bu ayniqsa bellarini qashshoqlash paytida foydalidir. 3 bor sakral vertebra, va qatoriga biriktirilgan tos suyagi 1-umurtqa tomonidan. Tos suyagi sudraluvchi, ajratilgan ilium, iskiyum va pubis. The suyak suyagi biroz s-shaklga ega, va undan qisqa, lekin uzunroq va ingichka humerus. Ko'pchilik uchun tibia va fibula bir-biriga qattiq egilib, tibia fibulaga qaraganda mustahkamroq.[6]:291–299 To'piq va tovon suyaklari orasidagi bo'g'im biroz harakatchan bo'lgan bo'lishi mumkin. Ikkala qo'l va oyoq uchun 5-raqam raqamga biriktirilmagan karpus /tarsus, va buning o'rniga to'g'ridan-to'g'ri ulangan ulna / tovon suyagi.[11]
Taksonomiya
Tasnifi

Birinchi gorgonopsian qoldiqlari Bofort guruhi ning Karoo Supergroup Janubiy Afrikadan chiqib ketgan va sifatida tasniflangan Gorgonops torvus 1876 yilda ingliz biologi tomonidan Richard Ouen. Ism Gorgonops dan olingan Qadimgi yunoncha γώνorγώνma'nosi "gorgon ", va ὄψ, "tomon" ma'nosini anglatadi. Ouen ushbu va u supergrupda tasvirlangan boshqa bir necha taksonlar deb taxmin qildi sovuqqon sudralib yuruvchilar, shu bilan birga ularning tishlari go'shtli sutemizuvchilarnikiga o'xshash edi va shuning uchun hammasini yangi tanilganlar qatoriga kiritishni taklif qildilar. buyurtma Theriodontia (u joylashtirgan sinf Reptiliya). U Theriodontia-ni bo'linishga qaror qildi oilalar burun teshiklari anatomiyasiga asoslangan (suyak nariallari) - sutemizuvchilardek burun singari bosh suyagida 1 teshik bo'lganlar uchun, "Binarialia" sudralib yuruvchilar singari 2 teshikka ega bo'lganlar uchun va "Tectinarialia" Gorgonops chunki uning ochilishini qalin suyak tomi qoplagan[12] (tektus bu Lotin "yopiq, tom yopilgan, pastki" uchun).[13] 1890 yilda ingliz tabiatshunos Richard Lydekker qilingan Gorgonops The tur turlari Gorgonopsidae oilasiga mansub.[14] 1895 yilda ingliz paleontologi Garri Sili ishondi Gorgonops Theriodontia diagnostikasi bo'lgan vaqtinchalik suyakda (temporal fenestrae) ochilish yo'q edi va shuning uchun Gorgonopsidae Gorgonopsia-ga ko'tarilib, Theriodontia-dan ajralib chiqdi. U sudralib yuruvchilar va sutemizuvchilar xususiyatlarini o'z ichiga olgan barcha Janubiy Afrikadagi materiallarni "Theriosuchia" tartibiga kiritdi va Gorgonopsia va Theriodontia ni ko'rib chiqdi. suborderlar undan.[15] 1903 yilda amerikalik paleontolog Genri Feyrfild Osborn sudralib yuruvchilarning tasnifini to'liq qayta ishladi va ikkita katta guruhni o'rnatdi: Diapsida va Sinapsida,[16] va 1905 yilda Janubiy Afrikalik paleontolog Robert supurgi 3-guruhni yaratdi, Terapsida, "sutemizuvchilarga o'xshash sudralib yuruvchilar" ni, shu jumladan Theriodontia ni joylashtirish uchun. Shuningdek, u Sleylining da'vosiga qarshi chiqdi va ligaga tushib ketdi Gorgonops Theriodontia-ga qaytib keldi, lekin u uni yangi qurilgan kichik guruhiga joylashtirdi Terosefali, Gorgonopsiyani eritib yuborish.[17] 1913 yilda, ayniqsa deyarli to'liq tugallanganda G. torvus Muhtaram Jon H. Uayts tomonidan kashf etilgan bosh suyagi, Gorgonopsiyani qayta tikladi.[18]
Gorgonopsians birinchi marta Rossiyada 1890-yillarda Sokolki hududida aniqlangan Shimoliy Dvina daryosi rus paleontologi nazorati ostida Sibirda Vladimir Proxorovich Amalitskii. Vafotidan keyingi nashrda u shunday tasvirlangan Inostrancevia alexandri, va u eng taniqli va eng katta gorgonopsianlardan biridir. O'shandan beri yana bir necha rus avlodlari tasvirlangan: Pravoslavleviya, Viatkogorgon, Suchogorgon, Leogorgon va Nochnitsa.[19] Afrikada gorgonopsians shuningdek, Karoo tashqarisida topilgan Ruhuhu vodiysi, Tanzaniya; The Yuqori Luangva vodiysi, Zambiya; va Chiveta, Malavi.[6]:7 Gorgonopsianlar ushbu 2 sohadan tashqarida ko'rinmaydi.[19] 1979 yilda xitoylik paleontolog Yang Zhonjian xitoylik gorgonopsiyani tasvirlab berdi "Wangwusaurus tayuensis "kech Permiyadagi tishlarga asoslangan Jiyuan shakllanishi,[20] ammo 1981 yilda paleoontologlar Denis Sigogneau-Rassel va Ai-Lin Sun tayinlangan materialni tasodifiy yig'ilish deb topdi, ularning ikkitasi ham Gorgonopsiya bilan uzoqdan o'xshashligi bor.[21] 2003 yilda hind paleontologlari Sanghamitra Rey va Sasvati Bandyopadxay Keyingi Permiyadagi bosh suyagi parchalarini tayinladilar. Kundaram shakllanishi o'rta gorgonopsianga,[22] garchi gorgonopsiya xususiyatlari ba'zi terefoseliylarda ham hujjatlashtirilgan.[19] 2008 yilda Kech Permiyadagi katta va ehtimol rubidjinli yuqori jag bo'lagi va iti aniqlandi Moradi shakllanishi Nigerda (bir necha past kenglikdagi kech perma tetrapodli shakllanishlaridan biri) va past kenglikdagi gorgonopsianing birinchi dalilidir.[23]
20-asrda Janubiy Afrikadagi nasllar soni tez sur'atlar bilan o'sib bordi, ular asosan Karumoterapistlar ustida olib borgan keng ko'lamli ishlari - 1897 yilda mamlakatda kariyerasini boshlaganidan 1951 yil vafotigacha - 57 gorgonopsianni ta'riflashga olib keldi. holotip namunalari va 29 avlod. Bromning ko'plab taksonlari keyinchalik bekor qilinadi.[24] Boshqa ko'plab zamonaviy ishchilar bitta namunalar asosida butunlay yangi turlarni yoki turlarni yaratdilar.[6]:57 Binobarin, Gorgonopsiya juda ko'p taksonomik tartibsizliklarga duch keldi va eng muammoli sinapsid guruhlaridan biri hisoblanadi. Boshsuyagi anatomiya taksonlar bo'yicha juda kam farq qiladiganligi sababli, ko'pchilik noaniq mutanosiblik farqlari, shu jumladan taniqli a'zolar asosida aniqlanadi. Nominal turlar asosan shaxsning yoshiga, shu jumladan o'zgaruvchanligi ma'lum bo'lgan xususiyatlar bilan ajralib turadi ko'z orbitasi postkanin tishlarining kattaligi, uzunligi va soni. Shunday qilib, ba'zi taksonlar bo'lishi mumkin sinonim bir-biri bilan va rivojlanishning turli bosqichlarini ifodalaydi.[25]
Klasni tashkil qilish bo'yicha birinchi urinishlar orasida ingliz zoologi tomonidan amalga oshirilgan Devid Meredith Seares Watson va amerikalik paleoontolog Alfred Romer 1956 yilda uni 20 oilaga ajratgan, ulardan 3 (Burnetiidae, Hipposauridae va Phthinosuchidae) a'zolari endi gorgonopsiya hisoblanmaydi.[4] 1970 yilda va yana 1989 yilda, asosan Afrika taksonlarini hisobga olgan holda, Sigogneau-Russell keng qamrovli nashr etdi monografiya Gorgonopsia bo'yicha (uni buzg'unchilik ) va faqat 2 ta oilani tan oldi: Watongiidae va Gorgonopidae. Vatongiya ko'chirildi Varanopidae 2004 yilda. U Gorgonopidae-ni 3 subfamilaga - Gorgonopsinae, Rubidgeinae va Inostranceviinae-ga ajratdi va avlodlar sonini 23 taga qisqartirdi.[26] 2002 yilda rus paleontologi Mixail Feodosevich Ivaxnenko rus taksilarini hisobga olgan holda, uning o'rniga Gorgonopsiyani suborder deb hisoblab, uni birlashtirdi. Dinosefali "Gorgodontia" tartibida. U Gorgonopsiyani superfamilalar "Gorgonopioidea" (Gorgonopidae, Cyonosauridae va Galesuchidae oilalari) va "Rubidgeoidea" (Rubidgeidae, Phtinosuchidae va Inostranceviidae). 2007 yilda biolog Eva V. I. Gebauer Gorgonopsiya (uning nomzodlik dissertatsiyasi) ga bag'ishlangan obzorida Ivaxnenkoning modelini Sigogno-Rassell foydasiga rad etdi,[6]:57 va rus naslidan tashqari 14 turga qisqartirildi: Alopozavr, Cyonosaurus, Aelurosaurus, Sauroctonus, Scylacognathus, Eoarctops, Gorgonops, Njalila, Lycaenops, Arktognat, Aelurognathus, Sikozavr, Klelandina va Rubidiya.[6]:244 Umuman olganda, Sigogneau-Russell modeli qo'llab-quvvatlanadi, ammo qaysi oilalarga qaysi nasllarni berish mumkinligi to'g'risida ozgina kelishuv mavjud emas.[4] 2015 yilda amerikalik paleoontolog Kristian F. Kammerer va uning hamkasblari qayta ko'rib chiqdilar Eriphostoma (bu noaniq termiodont deb nomlangan) gorgonopsiya,[27] va cho'kib ketgan Scylacognathus va keyingi yil Eoarctops unga.[4]
Birinchi filogeniya (oilaviy daraxt) Gorgonopsiya a'zolari 2016 yilda amerikalik paleoontolog Kristian F. Kammerer tomonidan Rubidjineni maxsus tekshirgan va ikkala subfamilyani va u tayinlagan 9 turni qayta tavsiflagan (sonini 36 turdan qisqartirgan) tomonidan nashr etilgan. Kammerer ham tirildi Dinogorgon, Leontosaurus, Ruhuhucerberus va Smilesaurus. Kammerer bunga amin emas edi Leontosaurus, Klelandina, Dinogorgonva Rubidiya barchasi bir xil taksoni ifodalaydimi yoki yo'qmi (buning uchun Dinogorgon bor ustuvorlik ), lekin u ularning barchasini tasniflashga qaror qildi qabila Rubidjini qo'shimcha tekshiruvni kutmoqda.[4] 2018 yilda Kammerer va rossiyalik paleoontolog Vladimir Masyutin yangi naslni aniqladilar Nochnitsa eng taniqli gorgonopsians sifatida va barcha rus taksonlari (bundan mustasno) Viatkogorgon, tashqi ko'rinishdagi) Afrika taksonlaridan butunlay alohida qoplama hosil qiladi.[19] Shuningdek, 2018 yilda paleobiolog Eva-Mariya Bendel, Kammerer va uning hamkasblari tirilishdi Kinariplar.[2]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Bendel va boshqalarning fikriga ko'ra Gorgonopsiya filogeniyasi. 2018 yil[2] |
Evolyutsiya
Sinapsida an'anaviy ravishda bazalga bo'lingan "Pelikozauriya "va Terapsidadan olingan. Birinchisiga sovuq qonli jonivorlar kiradi, ular yurishi keng tarqalgan va metabolizmning past darajasi Yuqori karbon. 20-asrning o'rtalaridan oxirigacha amerikalik paleontolog Everett C. Olson O'rta Permiyadagi sinapsid xilma-xilligini o'rgangan San-Anjelo, Gul tuvagi va Chickasha Shimoliy Amerikadagi shakllanishlar va pelikozavrlarning xilma-xilligi ushbu shakllanishlarda 6 dan 3 gacha kamayganligini va ular terapevt deb talqin qilgan bir nechta bo'lak namunalar bilan birga bo'lganligini ta'kidladilar. Keyin u smenaning O'rta Permiya davrida sodir bo'lishini taklif qildi (Olsonning yo'q bo'lib ketishi ); ammo, o'sha "terapsidlar" ning tasnifi va shakllanish yoshi shundan beri shubha ostiga olingan. Shunday qilib, pelikozavr darajasidan terapsid darajasiga moslashuvchan o'tishning aniq vaqti aniq emas, ammo 7 asosiy terapid guruhi (Biarmosuchiya, Dinosefali, Anomodontiya, Gorgonopsia, Therocephalia va Cynodontia) 265 million yil oldin mavjud bo'lgan.[28]
| Sinapsida[28] | |||||||||||||||||||||||||||||||||||||||
|

Permiya asta-sekin quritadigan va quritadigan bo'ldi. Yuqori karbon va quyi permda pelikozavrlar doimgiga o'xshash bo'lib qolgan ko'mir botqog'i ekvator yaqinidagi yashash joylari (paleoekvatorning har ikki tomonidan 10 ° masofada ma'lum bo'lgan qoldiqlar); shundan keyin taxminan 30 ° gacha bo'lgan keng cho'l bo'lib, u botqoqlarni mo''tadil mintaqalardan ajratib, qirg'oqqa qadar cho'zilgan. O'rta Permiyagacha ekvatorial o'rmonlar mavsumiy ho'l / quruq tizimga o'tdi, ammo botqoqlar Sharqiy Panjeya bo'ylab qirg'oq yo'llari orqali mo''tadil zonalarga bog'lanib, hozirgi Janubiy Afrikadan hozirgi Rossiyaga kontinental ko'chib o'tishga imkon berdi. . Terapsidlar ushbu mavsumiy nam / quruq landshaftda rivojlanib, mo''tadil zonalarga ham kengaygan ko'rinadi. Bu vaqtda sinapsidlar atrofdagi yagona yirik quruqlikdagi hayvonlar edi; va pelikozavrlar aridifikatsiyaga moslasha olmagan bo'lishi mumkin. Pelikozavrlar yo'q bo'lib ketish vaqtida terapevtiklar katta moslashuvchanlikni boshdan kechirdilar nurlanish (barcha yirtqich hayvonlar) yuqori Permiygacha davom etadi.[28]
O'rta Permiya bo'ylab ko'pincha ulkan dinozefaliyalar ularning ekotizimlarining dominant hayvonlari edi. Ular fotoalbomlardan yo'qoladi Wordian, oldin Kapitaniyada ommaviy qirilish hodisasi xitoyliklarni vujudga keltirgan vulkanik faollik Emeishan tuzoqlari. Ularning yo'q bo'lishining aniq sababi noaniq, ammo ularning o'rnini gorgonopsiya va dicynodonts (bu kattaligi juda ko'payishni boshlagan) va kichikroq tsefoseliylar.[29] Rubidjenlar eng so'nggi va natijada eng katta va og'ir qurilgan gorgonopsian guruhi edi.[4]
Paleobiologiya
Tishlash

Gorgonopsians, ehtimol, faol yirtqichlar edi. Gorgonopsiyaliklar orasida, ayniqsa, bosh suyagini asosiy qurol sifatida ishlatadigan ulkan makropredatorlarnikiga o'xshab qurilgan bosh suyagi bor. mosasaurlar yoki ba'zilari teropod dinozavrlar. Uzunroq itlar va juda ozroq tishlash kabi kamroq kuchli gorgonopsians Smilesaurus yoki InostransviyaBuning o'rniga, ehtimol, ularning itlarini qirg'in qilish uchun ishlatgan, shundoqqina tishli mushuklarga o'xshash. Ning postkaninlari Klelandina o'rniga pichoqqa o'xshash dicynodontslardan farqli o'laroq silliq tizma bilan almashtirildi keratinli va u asosan yirtqichlardan ketib qolgan bo'lishi mumkin. Gorgonopsian taksonlari bir-birlari bilan bir vaqtda yashagan - bir vaqtning o'zida 7 ga yaqin va ba'zi rubidjinlarda postkaninlar borligi, boshqa ba'zi zamonaviylar esa ular amaliyot qilganligini anglatmaydi. joyni ajratish va turli xil o'lja buyumlarini ta'qib qildilar.[4]
Uzaygan itlar odatda ularning ov qilish taktikasida muhim rol o'ynagan deb o'ylashadi. Goronopsian jag 'menteşe er-xotin bo'g'inli va bir oz harakatlanuvchi va aylanadigan suyaklardan tashkil topgan bo'lib, ular og'zini nihoyatda keng ochishlariga imkon bergan bo'lardi - ehtimol, 90 ° dan oshiqroq - jag'ning echimiga ehtiyoj qolmasdan.[30] Shu bilan bir qatorda, har qanday shamshir tishli sinapsid guruhi orasida shamshirlar birinchi navbatda evolyutsiyasi tufayli rivojlangan deb taxmin qilingan jinsiy tanlov (juftlashishning bir shakli sifatida displey ) ba'zi zamonaviy kiyik turlarida bo'lgani kabi, ammo sinerzali tirik sinapsid yirtqichlarning etishmasligi sababli buni sinash qiyin. Qilich tishli mushuklarda uzoq sabrli ("dirk-tishli") taksonlar bo'lgan deb o'ylashadi ta'qib ovchilari, ammo qisqa tishli ("scimitar-tishli") taksonlar bo'lgan deb o'ylashadi pistirma yirtqichlari.[31] Kirli tishli mushuklar orasida bu yirtqichlar o'ljani urishtirgandan keyin tomoqqa yaxshi o'ralgan holda o'ldirilgan deb taxmin qilinmoqda, ammo gorgonopsiyachilar sudraluvchilarning jag'lari va tishlari bilan qurollangan holda, tishlamoq joyi bilan unchalik aniq bo'lmagan bo'lishi mumkin. Buning o'rniga, gorgonopsiyachilar tishlash va orqaga chekinish taktikasini qo'llaganlar: gorgonopsian o'z karerini pistirmaga olib, undan katta va zaiflashtiruvchi luqma olib chiqib ketar, so'ngra jarohatlariga berilmasdan qochib qutulishga urinayotganda unga ergashar edi. qotillik. Postkaninlar kamaytirilganligi yoki umuman yo'qligi sababli, go'sht tana go'shtidan kuch bilan yirtilib, butunlay yutib yuborilgan bo'lar edi.[30]

Gorgonopsians, boshqa dastlabki yirtqich hayvonlar va timsohlar bilan birgalikda, asosan, pterygoid va o'sha yirtqichni tishlamoqchi bo'lgan "Kinetik-Inertial tizim" (KI) ga tayanganlar. temporal mushaklar foydalanib, jag'larni tezda qisib qo'ydi impuls va kinetik energiya jabrlanuvchini tortish uchun jag'lar va tishlarning. Sutemizuvchilarning yirtqich hayvonlari, shu jumladan qasr tishli mushuklar, asosan, "Statik bosim tizimiga" (SP) tayanadi, bu erda temporalis va masseter mushaklari o'ljani o'ldirish uchun kuchli luqma kuchini ishlab chiqarish.[9] Temporalis va masseter faqat sutemizuvchilardan ajralgan, gorgonopsiyalar esa mushaklarning pastki qismidan cho'zilib ketgan. bosh suyagi tomi, orqaga skuamozal suyak (Bosh suyagining orqa qismida) va yonoq suyaklari bo'ylab; yonoqlar tomonidan langarga qo'yilgan qism jag 'suyagini stabillashtirgan va uni yopish paytida yonma-yon harakatlanishiga imkon bergan va bu tishlashda juda muhim bo'lgan bo'lishi mumkin, chunki itlar uzoqlashishi bilan tandemda yonoq suyaklari kuchayadi.[6]:278
Kabi kichik gorgonopsianlar Cyonosaurus (bu aslida boshqa turdagi balog'atga etmagan bolani anglatishi mumkin), nazokatli bosh suyaklari va qilichlari bo'lgan va xuddi shunday harakat qilgan bo'lishi mumkin shoqollar va tulkilar. Kattaroq gorgonopsianlar, masalan Gorgonops, kuchli pterygoidlarni va kuchli KI ısırmasını qo'llab-quvvatlaydigan kuchli yonib ketgan yonoqlari bilan uzoq muddatli mustahkam burunlari bor edi. O'rtacha Arktognat qutiga o'xshash bosh suyagi va natijada kuchli tumshug'i bor edi, bu kuchli egilish va burish harakatlariga imkon beradi va KI va SP tishlash elementlarining kombinatsiyasini beradi. Hatto katta gorgonopsianlar, masalan Arktops, oldingi sfenekodont singari qisqaroq va qavariq tumshug'i bor edi Dimetrodon, va keng japdan yopilgan jag'larni tezda qisib qo'yishi mumkin edi (bu uzun itlar uchun zarur bo'lar edi). Bundan ham kattaroq Rubidjinada nihoyatda kuchli, og'ir qurilgan, dumaloq suyaklari, keng tumshuqlari, kuchli yonoqlari va nihoyatda uzun tishlari bor edi; Qilichlar Rubidgea atrox tishlaridan uzunroq Tiranozavr.[9]
Sutemizuvchilarning yirtqich hayvonlaridan farqli o'laroq, gorgonopsiya (va tereosefali) postkaninlarni kamaytirgan yoki umuman etishmayotgan edi, va ehtimol jag 'suyakni ochish uchun zarur bo'lgan siljish bosimini o'tkaza olmadi ilik. Sinapsidlarda suyak iligi bo'lmaganligi sababli, ulardan foydalanishning iloji yo'q edi.[32]
Joylashtirish
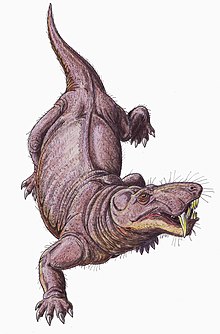
Gorgonopsianlar qat'iyan qabul qilingan deb hisoblanadi quruqlik.[33] Gorgonopsianlar timsohlar ishlatganiga o'xshash tik yurish bilan harakatlana olishgan, deb o'ylashadi, oyoq-qo'llar, kaltakesaklarning keng yurishida bo'lgani kabi gorizontal ravishda qarama-qarshi bo'lib, deyarli vertikal holda joylashgan. The glenoid bo'shliq elkama pichog'ida dumaloq tomoni kuchli burchakli, shuning uchun oyoq-qo'llar oldinga siljish harakati cheklangan va ular qisqa qadam uzunligiga ega bo'lishi mumkin. Qatlam uzunligini oshirish uchun kaltakesaklar ko'pincha tizmalarini yonma-yon harakatlantiradi, lekin vertikal yo'naltirilgan faset qo'shimchalari gorgonopsianlarda umurtqalarni bog'lash umurtqa pog'onasini yanada qattiqroq va barqaror qilib, bunday harakatni og'irlashtirgan bo'lar edi.[6]:259–260
Goronopsian yelka qo'shilishi juda g'ayrioddiy konfiguratsiyaga ega. Yelka bilan bog'langan humerus boshi glenoiddan uzunroq, shuning uchun u bo'shliqqa kira olmadi. Binobarin, ular katta xaftaga massasi bilan biriktirilgan bo'lishi mumkin, bunda humerus glenoid ustida siljish harakatini amalga oshiradi. Bu nazariy jihatdan humerus va glenoid orasidagi burchakni hayvonga qaragan holda 80-145 ° gacha o'zgartirishi mumkin. Agar burchak pastki uchida bo'lsa (ya'ni bir oz bosh bilan burilgan bo'lsa), bu juda mustahkam bo'g'in bo'lib, deltalar old tomondan katta kuch sarflash, masalan, kurashayotgan o'ljani qoqish yoki tana go'shtini yulib olish paytida tana go'shtini ushlab turish. Agar humerus yuqoriroq burchak ostida joylashgan bo'lsa (pastga qarab), bu oldinga va orqaga (uzun o'qi bo'ylab) kengaytirilgan kengaytmaga va shu tariqa hujumda yoki qisqa ta'qibda ko'proq qadam uzunligiga imkon berishi mumkin edi. Yelka pichog'i hayvonning yon tomonlariga kengayadi (yon tomonga chiqib turadi), shuningdek deltalar uchun katta biriktirma beradi. Hammasi uchun ishlab chiqarilgan biriktirma mavjud edi skapulohumeral mushaklar, ayniqsa deltalar. Old oyoqlarni kengaytirganda, deltalar humerusning old tomonini (old chetini) ko'targan bo'lishi mumkin va coracobrachialis mushaklari orqa tomonni pastga tushirdi (orqa chekka). Old oyoqni tortib olayotganda ko'krak mushaklari oldingi chetini pastga itarib yuborgan bo'lishi mumkin va subkapularis mushak orqa chetini yuqoriga tortdi.[6]:260–264
Tos suyagi qo'shilishi odatdagiga ega koptok va rozetkaning qo'shilishi konfiguratsiya. Bir oz yassilangan femur boshi nazariy jihatdan bunga mos kelishi mumkin edi kestirib uyasi keng burchak ostida. 1982 yilda paleontolog Tom S. Kemp dastlabki termodontlar, shu jumladan gorgonopsiyalar, femurni keng yurishda gorizontal burchak ostida, shuningdek tik yurishda vertikal burchak ostida joylashtirishi mumkin deb taxmin qildi. U bu jonzotlarning lokomotor odatlarini timsohlarga taqqosladi, ular qisqa masofada yurishdan foydalanadilar, ammo uzoqroq masofalarga yugurish yoki harakat qilish paytida tik turishga o'tadilar. Garchi kestirib Sauroctonus o'rtasida anatomik oraliq bo'lib ko'rinadi Dimetrodon va sutemizuvchilar - bilan ilium quyruqlardan ko'ra bosh tomon yo'nalishda ko'proq kengaygan va pubis biroz qisqargan - puboischiofemoralis mushaklari (katta mushak, faqat tos suyagidan femurgacha o'tadigan sudralib yuruvchilar tomonidan olib boriladi) pubisning pastki qismiga juda bog'langan va iskiyum, bu kuchli ishlab chiqarishga imkon bergan bo'lar edi qo'shimchalar kuch (oyoqlarni tanaga yaqinlashtirish), keng yurishda foydali. Bundan tashqari, gorgonopsiyachilar, asosan, mushaklar bilan kurashayotgan o'lja bilan kurashayotganda shug'ullangan deb o'ylash mumkin.[6]:264–270 Bilagi zo'r suyaklar bilan taqqoslaganda qisqaroq bo'lib, gorgonopsiyalar uzoq masofalarga yugurish uchun yaxshi moslashmaganligini anglatadi.[6]:298–299

Oyoqlarning erga qanday joylashtirilganligi to'g'risida gorgonopsianlar ektataksoniya (oxirgi raqam eng katta vaznga ega), gomopodiya (oyoq izlari va qo'l izlari bir xil ko'rinishda) mavjud bo'lgan yagona terapevtlardir. yarim o'simlik (ma'lum darajada oyoqlar erga tekis qilib qo'yilgan).[34] Ushbu moslashuvlar gorgonopsianlarni o'ljalariga qaraganda tezroq va epchilroq qilgan bo'lishi mumkin.[30] Gorgonopsians juda chaqqon raqamlarga ega edi, bu ikkala qo'l va oyoq uchun tushunish qobiliyatini ko'rsatib turibdi, ehtimol ular haddan tashqari yuk ko'tarilishining oldini olish uchun kurashayotgan o'lja bilan kurashish, va natijada ular jabrlanuvchiga singib ketayotganda itlarning sinishi yoki sinishi.[11]
Sezgilar

Ko'proq olingan termiodontlardan farqli o'laroq, ammo bugungi kunda ba'zi bir ektotermik jonzotlar singari, barcha gorgonopsiyaliklar boshning tepasida pineal ko'zga ega edilar, bu esa kunduzgi yorug'likni aniqlash uchun ishlatiladi (va shunday qilib, optimal harorat faol bo'lishi kerak). Ehtimol, boshqa termodontlar buni endotermiya evolyutsiyasi tufayli yo'qotgan bo'lishi mumkin, ichki sezgir retinal ganglion hujayralari Ko'zlarda - rang ko'rishni yo'qotish va tungi hayotga o'tish bilan yoki ikkalasi bilan birgalikda.[35] Tungi xatti-harakatlar azaldan sutemizuvchilardan kelib chiqqan deb taxmin qilingan (tungi tiqilish ), lekin katta orbitaning hajmi va mavjudligi sklerotik halqalar Karbonifergacha cho'zilgan ko'plab dastlabki sinapsidlarda kam nurli sharoitda chiqib ketish qobiliyati ancha oldin rivojlangan degan fikrni bildiradi. Ushbu jihatlarga asoslanib, Sauroctonus parringtoni bo'lishi mumkin edi mezopik ko'rish va Cyonosaurus skotopik yoki fotopik ko'rish.[36] Kichkintoylar uchun sklerotik halqalarning diametrlari Viatkogorgon mutanosib ravishda katta bo'lib, ichki diametri 1,5 sm (0,59 dyuym) va tashqi diametri 2,3 sm (0,91 dyuym), orbitaning o'zi uchun 2,8 sm (1,1 dyuym) diametriga nisbatan, bu asosan tungi ekskursiyalarni amalga oshirganligini anglatadi.[37] Gorgonopsianlar orasida rubidjin Klelandina g'ayrioddiy kichkina sklerotik halqalarga ega, bu uning fotopopik ko'rish qobiliyatiga ega ekanligini va qat'iy bo'lganligini ko'rsatadi kunduzgi; Kammerer rubidjinlar o'rtasida bo'linishni taqsimlashni taklif qildi (chunki bu hududda 7 xil takson mavjud edi), qisman kunning turli vaqtlarida faol bo'lgan har xil turlar shaklini oldi, ammo faqat sklerotik halqalar Klelandina Ushbu subfamila orasida aniqlangan, bu gipotezani juda spekulyativ qiladi.[4]
Gorgonopsianlar pelikozavrlar singari juda qisqa burun bo'shlig'iga ega, ammo uning orqasida mo'l-ko'l bo'ylama tizmalar mavjud ichki burun teshiklari (burun bo'shlig'ini tomoq bilan bog'laydigan); chunki ular orqali nafas oladigan havo o'tmagan bo'lar edi, ular odatda hidlovchi turbinalar deb talqin qilinadi va gorgonopsiyachilarga ancha rivojlangan hid bilish hissi bergan bo'lar edi.[38] Gorgonopsians a vomeronazal organ ("Jeykobsonning organi") - ning bir qismi aksessuarlarning hidlash tizimi - tagiga joylashtirilgan bo'lar edi burun septum; dicynodonts va trocephaliansdan farqli o'laroq, organni og'iz bilan bog'laydigan kanal bor edi, bu uning gorgonopsiyalarda funktsionalligini ko'rsatmoqda.[39]

Dastlabki termodontlar (jumladan, gorgonopsiyalar) an quloq pardasi, avvalgi pelikozavrlardan farqli o'laroq, chunki to'rtburchak suyagi (jag 'menteşesinde) va pterigoid suyagi (da tomoq ) kamayishni boshlagan, bu kvadratni mustaqil ravishda bir darajaga tebranishiga imkon beradi. Bu havo bilan eshitiladigan tovushlarni past darajadagi tovushlarni aniqlashga imkon bergan bo'lishi mumkin amplituda 1 mm dan kam (0,039 dyuym), lekin quloq pardasi qo'llab-quvvatlagan bo'lar edi xaftaga yoki ligamentlar suyak o'rniga. To'g'ri bo'lsa, u holda tishdan keyingi suyaklar (bu erta sutemizuvchilarda hosil bo'lgan o'rta quloq suyaklar) tish tishidan (jag 'suyagidan) ajralishi kerak edi; gorgonopsian fotoalbomlari postdentary-dentary aloqaning kamayganligini ko'rsatmoqda. Gorgonopsiya uchun, muqobil ravishda, kvadratning izolyatsiyasi sabab bo'lishi kerak deb taxmin qilingan streptostil ravishda (aylanadigan kvadrat) eshitish qobiliyatini engillashtirish uchun emas, balki bo'shliqni oshirish uchun.[7]
Termoregulyatsiya
Oldingi pelikozavrlar va terapevtiklar o'rtasida katta anatomik siljish yuz berdi, bu o'sish bilan bog'liq deb taxmin qilingan. metabolizm va kelib chiqishi uy sharoitida davolanish (yuqori tana haroratini saqlab turish). Ikkilamchi tanglay evolyutsiyasi va og'izning burun bo'shlig'idan ajralishi yuqori darajadagi shamollatish samaradorligini oshirishi mumkin. aerob faoliyat; gorgonopsians suyakli ikkilamchi tanglayga ega emas edi, lekin, ehtimol, yumshoq to'qimalardan biri bo'lishi mumkin edi, ammo shunda ham, uning o'rniga ovqatlanish xatti-harakatlari bo'lishi mumkin edi. Skeletning qayta tashkil etilishi (tarqalishdan parazagittal yurishga qadar) mavjudligini ko'rsatuvchi postulyatsiya qilingan diafragma va shu bilan aerob faolligi uchun shamollatish yaxshilandi; ammo buning o'rniga tezlanishni yoki chaqqonlikni oshirish mumkin edi, bu timsohlar singari intensiv aerobik faollikka teng kelmaydi. Fibrous lamellar cortical bone, which all early therapsids had, would indicate an increased growth rate, but this may not be linked to metabolic rate.[40]
Modern large reptiles naturally give off body heat at a slower rate than smaller ones, and are considered "inertial homeotherms", but they maintain a low body temperature of 25–30 °C (77–86 °F). If therapsids required a higher body temperature of 35–40 °C (95–104 °F), they would either have needed to have been endotermlar (generated their own body heat) or have had greater control over heat loss (that is, better homeothermy). The parasagittal gait may have aided the latter, as it would have kept most of the body off the ground as well as allowed blood to stay in the abdomen instead of having to circulate through the appendages, both of which would reduce heat transfer to the ground and stabalise asosiy harorat. The reduced tail would have also reduced the total surface area of the animal, further minimising heat loss.[41] Among therapsids, only eutheriodonts (not gorgonopsians) have respiratory burun turbinalari, which help retain moisture while breathing in large quantities of air, and its evolution is typically associated with the beginning of "mammalian" oxygen consumption rates and the origins of endothermy.[38]
If gorgonopsians were inertial homeotherms, it is not impossible that they had hair. The snout is typically riddled with foramina (small holes which confer with blood vessels), which could potentially point to the existence of loose skin (as opposed to scales), hair, various teri bezlari (kabi ter bezlari ) va whiskers; however, some reptiles present a similar patterning of foramina, which are instead related to dental development rather than skin.[42]
Palaeopathology
The anterolateral aspect of the left radius (a forearm bone) of the gorgonopsian specimen NHCC LB396 presents a circular bony lesion, featuring irregular-to-radial spikes made of cortical bone surrounded by a thin layer of subperiosteal bone, which grew rapidly over a single growing season. Bu mos keladi periostit most likely stemming from subperiosteal gematoma. This specific condition as well as the fast growth rate are more reminiscent of mammals and dinosaurs than crocodilians or kaltakesaklarni kuzatish. Among early synapsids, the only other pathology noted is osteomiyelit in several pelycosaur groups.[43]
The labial (tongue) side of the tooth root of a functional canine of RB382 presents as many as 8 lesions, clustering along the midline of the tooth, which resemble miniature teeth with a pulpa, dentin, and a thin emal qoplama. They are roughly circular—with diameters varying from 0.3–3.9 mm (0.012–0.154 in)–though they become less circular at around the middle point of the root until passing the cervix of the tooth. This is roughly consistent with the human ailment odontoma, the most frequent type of odontogenic tumour, which previously only extended a few million years back in the fossil record. At 255 million years old, RB382 presents the oldest known case of odontoma.[44]
Paleoekologiya

Following the extinction of the dinocephalians and (in South Africa) the basal therocephalians Scylacosauridae va Likosuchidae, gorgonopsians evolved from small and uncommon forms into large tepalik yirtqichlari. Through the Middle to Upper Permian, in South Africa the dicynodonts were the most common animals, whereas the pareiasaurs Deltavjatiya va Skutozavr were the most abundant in the gorgonopsian-bearing Russian formations.[45] During the Upper Permian, the South African Beaufort Group was a semi-arid cold steppe featuring large, vaqtinchalik (temporary) rivers and floodplains draining water sources much farther north into the Karoo Sea, with some occurrences of toshqin toshqinlari after sudden, heavy rainfall;[46][47] ning taqsimlanishi karbonatlar is consistent with present-day kalisiya deposits which form in climates with an average temperature of 16–20 °C (61–68 °F) and 100–500 mm (3.9–19.7 in) of seasonal rainfall.[47] The Salarevskian Formation in western Russia was also probably deposited in a semi-arid environment with highly seasonal rainfall, and featured hygrophyte va halofit plants in coastal areas, as well as more drought-resistant ignabargli daraxtlar balandliklarda.[48] The Moradi Formation was an arid desert, primarily dominated by the reptile Moradisaurus and the pareiasaur Bunostegos, which likely fed on conifers.[23]
Gorgonopsians went extinct at the end of the Upper Permian during the Permiy-trias davridagi yo'q bo'lib ketish hodisasi, which was primarily caused by several series of major volcanic activity; the extinction over the Permian–Triassic border was primarily driven by volcanism which has since formed the Sibir tuzoqlari. The resultant massive spike in issiqxona gazlari caused rapid aridification due to: temperature spike (as much as 8–10°C at the equator, with average equatorial temperatures of 32–35 °C (90–95 °F) at the beginning of the Triassic), kislotali yomg'ir (bilan pH as low as 2 or 3 during eruption and 4 globally, and the subsequent dearth of forests for the first 10 million years of the Triassic), frequent o'rmon yong'inlari (though they were already rather common throughout the Permian), and potential breakdown of the ozon qatlami (possibly briefly increasing UV nurlanishi bombardment by 400% at the equator and 5000% at the poles).[49] Among therapsids, large herbivorous anomodonts managed to cross the Permian–Triassic boundary and survived until the Karnay ichida Yuqori trias, but only small-bodied species of cynodonts survived the extinction event. The nişler they left open were eventually filled by the arxhosaurs (including crocodiles and dinozavrlar ) during the early stages of the Trias.[50]
Shuningdek qarang
Izohlar
- ^ ce, serebellum; cnI, hid hid; cnV +vcm—trigeminal asab va vena capitis medialis; cnVI, abducens nerve; cnVII, yuz nervi; cnIX-XI, glossofaringeal va vagoaccessory nerves; cnXII, gipoglossal asab; en, epiphyseal nerve; fb, oldingi miya; fcl, flocculus; ibic, internal branch of the internal carotid; lob, left xushbo'y lampochka; ob, olfactory bulb; ot, xushbo'y hidli trakt; pg, gipofiz; pgll, pituitary gland lateral lobes; pf, pontine flexure; rob, right olfactory bulb; vc, vidian canal; vc=spa, vidian canal where the sfenopalatin arteriyasi passes; vcd, vena capitis dorsalis
Adabiyotlar
- ^ Angielchik, Kennet D.; Kammerer, Christian F. (2018). "Non-Mammalian synapsids: the deep roots of the mammalian family tree". Zakosda Frank E.; Asher, Robert J. (eds.). Sutemizuvchilar evolyutsiyasi, xilma-xilligi va sistematikasi. Berlin: De Gruyter. ISBN 9783110275902.
- ^ a b v d e f Bendel, Eva-Maria; Kammerer, Kristian F.; Kardjilov, Nikolay; Fernandez, Vinsent; Fröbisch, Jörg (2018). "Cranial anatomy of the gorgonopsian Cynariops robustus based on CT-reconstruction". PLOS ONE. 13 (11): e0207367. doi:10.1371/journal.pone.0207367. PMC 6261584. PMID 30485338.
- ^ a b Swinton, W. E. (1954). Fossil amphibians and reptiles. Britaniya muzeyi (Tabiat tarixi). p. 33.
- ^ a b v d e f g h men j Kammerer, Christian F. (2016). "Rubidjinalar sistematikasi (Therapsida: Gorgonopsia)". PeerJ. 4: e1608. doi:10.7717/peerj.1608. PMC 4730894. PMID 26823998.
- ^ a b Arauxo, Rikardo; Fernandez, Vinsent; Polsin, Maykl J.; Fröbisch, Jörg; Martins, Rui M. S. (2017). "Goronopsian paleobiologiyasi va evolyutsiyasi jihatlari: bazraniyum, oksiput, suyak labirintasi, qon tomirlari va neyroanatomiyadan tushunchalar". PeerJ. 5: e3119. doi:10.7717 / peerj.3119. PMC 5390774. PMID 28413721.
- ^ a b v d e f g h men j k l m Gebauer, E. V. I. (2007). Phylogeny and Evolution of the Gorgonopsia with a Special Reference to the Skull and Skeleton of GPIT/RE/7113 (PhD). Eberhard-Karls University of Tübingen.
- ^ a b v Laurin, M. (1998). "New data on the cranial anatomy of Lycaenops (Synapsida, Gorgonopsidae), and reflections on the possible presence of streptostyly in gorgonopsians". Umurtqali hayvonlar paleontologiyasi jurnali. 18 (4): 765–776. doi:10.1080/02724634.1998.10011105.
- ^ a b Kermack, D. M.; Kermack, K. A. (1984). "Dentitions, Tooth-Replacement and Jaw Articulation". Sutemizuvchilar belgilarining evolyutsiyasi. 71-79 betlar. doi:10.1007/978-1-4684-7817-4_5. ISBN 978-1-4684-7817-4.
- ^ a b v Van Valkenburg, B.; Jenkins, I. (2002). "Permo-trias va senozoy sinapsid yirtqichlari tarixidagi evolyutsion naqshlar". Paleontologik jamiyat hujjatlari. 8: 273–275. doi:10.1017/S1089332600001121.
- ^ a b Kermack, K. A. (1956). "Tooth replacement in mammal-like reptiles of the suborders Gorgonopsia and Therocephalia". London B Qirollik jamiyati falsafiy operatsiyalari: Biologiya fanlari. 240 (670): 95–133. doi:10.1098/rstb.1956.0013.
- ^ a b Kümmell, S. B.; Frey, E. (2014). "Range of Movement in Ray I of Manus and Pes and the Prehensility of the Autopodia in the Early Permian to Late Cretaceous Non-Anomodont Synapsida". PLOS ONE. 9 (12): e113911. doi:10.1371/journal.pone.0113911. PMC 4269487. PMID 25517726.
- ^ Ouen, R. (1986). Descriptive and illustrated catalogue of the fossil Reptilia of South Africa in the collection of the British museum. Britaniya muzeyi (Tabiat tarixi). 27-29 betlar.
- ^ tectus in Charlton T. Lewis (1891) Lotin tilining boshlang'ich lug'ati, Harper & Brothers
- ^ Lydekker, R. (1890). Catalogue of the fossil Reptilia and Amphibia in the British Museum (Natural history) Part IV. Britaniya muzeyi (Tabiat tarixi). p. 111.
- ^ Seli, H. G. (1895). "Researches on the structure, organization, and classification of the fossil reptilia.—Part IX. section 1. On the Therosuchia". Tabiiy tarix yilnomalari va jurnali. 13 (6): 375. doi:10.1080/00222939408677718.
- ^ Osborn, H. F. (1903). "The Reptilian Subclasses Diapsida and Synapsida and the Early History of the Diaptosauria. Volume I. Part VIII" (PDF): 451–466. Iqtibos jurnali talab qiladi
| jurnal =(Yordam bering) - ^ Supurgi, R. (1905). "On the use of the term anomodont". Albani muzeyining yozuvlari. 1: 266–269.
- ^ Supurgi, R. (1913). "On the Gorgonopsia, a Suborder of the Mammal-like Reptiles". London zoologik jamiyati materiallari. 1913 (2): 225–230. doi:10.1111/j.1469-7998.1913.tb07574.x.
- ^ a b v d Kammerer, C. F.; Masyutin, V. (2018). "Gorgonopsian therapsids (Nochnitsa gen. nov va Viatkogorgon) from the Permian Kotelnich locality of Russia". PeerJ. 6: e4954. doi:10.7717 / peerj.4954. PMC 5995105. PMID 29900078.
- ^ Young, C. C. (1979). "A Late Permian fauna from Jiyuan, Henan". Vertebrata PalAsiatica. 17 (2): 99–113.
- ^ Sigogneau-Russell, D.; Sun, A.-L. (1981). "A brief review of Chinese synapsids". Geobios. 14 (2): 276. doi:10.1016/S0016-6995(81)80012-5.
- ^ "Late Permian vertebrate community of the Pranhita–Godavari valley, India". Iqtibos jurnali talab qiladi
| jurnal =(Yordam bering) - ^ a b Smiley, T. M.; Sidor, C. A .; Maga, A .; Ide, O. (2008). "The vertebrate fauna of the Upper Permian of Niger. VI. First evidence of a gorgonopsian therapsid". Umurtqali hayvonlar paleontologiyasi jurnali. 28 (2): 543–547. doi:10.1671/0272-4634(2008)28[543:TVFOTU]2.0.CO;2.
- ^ Wyllie, A. (2003). "A review of Robert Broom's therapsid holotypes: Have they survived the test of time?" (PDF). Paleontologia Africana. 39 (39): 2.
- ^ Kammerer, C. F. (2013). "Theriodontia: Introduction". Sinapsidaning dastlabki evolyutsion tarixi. Umurtqali hayvonlarning paleobiologiyasi va paleoantropologiyasi. Springer. 165–169 betlar. doi:10.1007/978-94-007-6841-3_10. ISBN 978-94-007-6841-3.
- ^ Sigogneau-Rassell, Denis (1989). Wellnhofer, Piter (tahrir). Theriodontia I: Ftinosuchiya, Biarmosuchia, Eotitanosuchia, Gorgonopsia. Paleoherpetologiya entsiklopediyasi. 17 B / I. Shtutgart: Gustav Fischer Verlag. ISBN 978-3437304873.
- ^ Kammerer, C. F.; Smit, R. M. H.; Day, M. O .; Rubidge, B. S. (2015). "New information on the morphology and stratigraphic range of the mid‐Permian gorgonopsian Eriphostoma microdon Broom, 1911". Paleontologiyadagi hujjatlar. 1 (2): 201–221. doi:10.1002 / spp2.1012.
- ^ a b v Kemp, T. S. (2006). "The origin and early radiation of the therapsid mammal‐like reptiles: a palaeobiological hypothesis". Evolyutsion biologiya jurnali. 19 (4): 1231–1247. doi:10.1111 / j.1420-9101.2005.01076.x. PMID 16780524. S2CID 3184629.
- ^ Day, M. O .; Ramezani, J .; Bowring, S. A.; va boshq. (2015). "Qachon va qanday qilib er yuzidagi Permiyadagi ommaviy qirg'in sodir bo'lgan? Janubiy Afrikaning Karoo havzasi tetrapod yozuvlaridan dalillar". Qirollik jamiyati materiallari B. 282 (1811): 20150834. doi:10.1098 / rspb.2015.0834. PMC 4528552. PMID 26156768.
- ^ a b v Anton, Maurisio (2013). Sabertoth. Bloomington, Indiana: Indiana universiteti matbuoti. 204–207 betlar. ISBN 9780253010421.
- ^ Randau, M.; Carbone, C.; Turvey, S. T. (2013). "Canine Evolution in Sabretoothed Carnivores: Natural Selection or Sexual Selection?". PLOS ONE. 8 (8): e72868. doi:10.1371/journal.pone.0072868. PMC 3738559. PMID 23951334.
- ^ Cruikshank, A. R. I. (1973). "The Mode of Life of Gorgonopsians" (PDF). Palaeontologica Africana. 15 (1–2): 65–67.
- ^ Kriloff, A .; Jermen, D .; Kanovil, A .; Vinsent, P .; Sache, M .; Laurin, M. (2008). "Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference". Evolyutsion biologiya jurnali. 21 (3): 807–826. doi:10.1111 / j.1420-9101.2008.01512.x. PMID 18312321.
- ^ Marchetti, L.; Klayn, X.; Buchvits, M.; va boshq. (2019). "Permian-Triassic vertebrate footprints from South Africa: Ichnotaxonomy, producers and biostratigraphy through two major faunal crises". Gondvana tadqiqotlari. 72: 159. doi:10.1016/j.gr.2019.03.009.
- ^ Benoit, J .; Abdala, F.; Manger, P. R.; Rubidge, B. S. (2016). "The Sixth Sense in Mammalian Forerunners: Variability of the Parietal Foramen and the Evolution of the Pineal Eye in South African Permo-Triassic Eutheriodont Therapsids". Acta Palaeontologica Polonica. 61 (4): 777–789. doi:10.4202/app.00219.2015. S2CID 59143925.
- ^ Angielczyk, K. D.; Schmitz, L. (2014). "Nocturnality in synapsids predates the origin of mammals by over 100 million years". Qirollik jamiyati materiallari B. 281 (1793). doi:10.1098/rspb.2014.1642. PMC 4173690. PMID 25186003.
- ^ Kammerer, Kristian F.; Masyutin, Vladimir (2018). "Gorgonopsian therapsids (Nochnitsa gen. nov va Viatkogorgon) from the Permian Kotelnich locality of Russia". PeerJ. 6: e4954. doi:10.7717 / peerj.4954. PMC 5995105. PMID 29900078.
- ^ a b Hillenius, W. J. (1992). Nasal turbinates and the evolution of mammalian endothermy (PhD). Oregon shtat universiteti. 74-76 betlar. hdl:1957/35986.
- ^ Hillenius, W. J. (2000). "Septomaxilla of nonmammalian synapsids: Soft‐tissue correlates and a new functional interpretation". Morfologiya jurnali. 245 (1): 29–50. doi:10.1002/1097-4687(200007)245:1<29::AID-JMOR3>3.0.CO;2-B.
- ^ Kemp, T. S. (2006). "The origin of mammalian endothermy: a paradigm for the evolution of complex biological structure". Linnean Jamiyatining Zoologik jurnali. 147 (4): 473–488. doi:10.1111/j.1096-3642.2006.00226.x.
- ^ Turner, J. S.; Tracey, C. R. (1986). "Body Size, Homeothermy and the Control of Body Heat in Mammal-like Reptiles". Sut emizuvchilarga o'xshash sudralib yuruvchilarning ekologiyasi va biologiyasi (PDF). Smithsonian Institution Press.
- ^ Van Velan, L. (1960). "Therapsids as Mammals". Evolyutsiya. 14 (3): 304–313. doi:10.2307/2405973. JSTOR 2405973.
- ^ Kato, K. M.; Rega, E. A.; Sidor, C. A .; Huttenlocker, A. K. (2020). "Investigation of a bone lesion in a gorgonopsian (Synapsida) from the Permian of Zambia and periosteal reactions in fossil non-mammalian tetrapods". Qirollik jamiyatining falsafiy operatsiyalari B. 375 (1793). doi:10.1098/rstb.2019.0144. PMC 7017433. PMID 31928188.
- ^ Whitney, M. R.; Mose, L.; Sidor, C. A. (2017). "Odontoma in a 255-Million-Year-Old Mammalian Forebear". JAMA Onkologiya. 3 (7): 998–1000. doi:10.1001/jamaoncol.2016.5417. PMC 5824274. PMID 27930769.
- ^ Kammerer, Kristian F.; Masyutin, Vladimir (2018). "A new therocephalian (Gorynychus masyutinae gen. va boshqalar. nov.) from the Permian Kotelnich locality, Kirov Region, Russia". PeerJ. 6: e4933. doi:10.7717/peerj.4933. PMC 5995100. PMID 29900076.
- ^ Uilson, A .; Flint, S.; Payenberg, T.; Tohver, E. (2014). "Architectural Styles and Sedimentology of the Fluvial Lower Beaufort Group, Karoo Basin, South Africa". Cho'kindi tadqiqotlar jurnali. 84 (4): 326–348. doi:10.2110/jsr.2014.28.
- ^ a b Smith, R. M. H. (1987). "Morphology and depositional history of exhumed Permian point bars in the southwestern Karoo, South Africa". Cho'kindi tadqiqotlar jurnali. 57 (1): 19–20. doi:10.1306/212F8A8F-2B24-11D7-8648000102C1865D.
- ^ Yakimenko, E.; Inozemtsev, S. A.; Naugolnykh, S. V. "Upper Permian paleosols (Salarevskian Formation) in the central part of the Russian Platform: Paleoecology and paleoenvironment". 21 (1): 110–119. Iqtibos jurnali talab qiladi
| jurnal =(Yordam bering) - ^ Benton, M. J. (2018). "Hyperthermal-driven mass extinctions: killing models during the Permian–Triassic mass extinction". Qirollik jamiyatining falsafiy operatsiyalari A. 376 (2130). doi:10.1098/rsta.2017.0076. PMC 6127390. PMID 30177561.
- ^ Sookias, R. B.; Butler, R. J.; Benson, R. B. J. (2012). "Rise of dinosaurs reveals major body-size transitions are driven by passive processes of trait evolution". Qirollik jamiyati materiallari B. 279 (1736): 2180–2187. doi:10.1098/rspb.2011.2441. PMID 22298850. S2CID 1231897.
Qo'shimcha o'qish
- Bakker, R.T. (1986), Dinozavrlarning bid'atlari, Kensington Publishing Corp.
- Cox, B. and Savage, R.J.G. and Gardiner, B. and Harrison, C. and Palmer, D. (1988) The Marshall illustrated encyclopedia of dinosaurs & prehistoric animals, 2nd Edition, Marshall Publishing.
- Fenton, C.L.and Fenton, M.A. (1958) Fotoalbomlar kitobi, Ikki karra nashr.
- Hore, P.M. (2005), The Dinosaur Age, Issue #18. National Dinosaur Museum.
- Ward, P.D. (2004), Gorgon, Viking Pengueni.
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||






