MRNA nazorati - MRNA surveillance
mRNK kuzatuv mexanizmlari bor yo'llar sodiqligi va sifatini ta'minlash uchun organizmlar tomonidan foydalaniladi xabarchi RNK (mRNA) molekulalari. Hujayralar ichida bir qator kuzatuv mexanizmlari mavjud. Ushbu mexanizmlar mRNKning turli bosqichlarida ishlaydi biogenez to'g'ri ishlov berilmagan transkriptlarni aniqlash va buzish uchun yo'l.
Umumiy nuqtai
The tarjima ning xabarchi RNK transkriptlar oqsillarga molekulyar biologiyaning markaziy dogmasi. mRNK molekulalari mRNKning sifatiga o'girilishida xatolarni keltirib chiqarishi mumkin bo'lgan sodiqlik xatolariga moyil. oqsillar.[1] RNK nazorati mexanizmlari - hujayralar mRNK molekulalarining sifati va sodiqligini ta'minlash uchun foydalanadigan usullar.[2] Bunga odatda aberrant mRNA molekulasini markalash orqali erishiladi tanazzul turli xil endogen nukleazalar.[3]
mRNA nazorati hujjatlashtirilgan bakteriyalar va xamirturush. Yilda eukaryotlar, bu mexanizmlarning ikkalasida ham ishlashi ma'lum yadro va sitoplazma.[4] Yadroda mRNK molekulalarining ishonchliligi tekshiruvi sitoplazmada eksport qilinishidan oldin noto'g'ri ishlov berilgan transkriptlarning degradatsiyasiga olib keladi. Transkriptlar sitoplazmada bir marta qo'shimcha tekshiruvdan o'tkaziladi. Sitoplazmatik kuzatuv mexanizmlari mRNKni baholaydi stenogrammalar erta to'xtash kodonlari yo'qligi yoki mavjudligi uchun.[3][4]
Hozirda uchta kuzatuv mexanizmi faoliyat yuritishi ma'lum hujayralar: bema'nilik vositachiligidagi mRNA parchalanish yo'li (NMD); uzluksiz vositali mRNA parchalanish yo'llari (NSD); va taqiqlangan mRNA parchalanish yo'li (NGD).
Bema'nilik vositachiligidagi mRNA parchalanishi
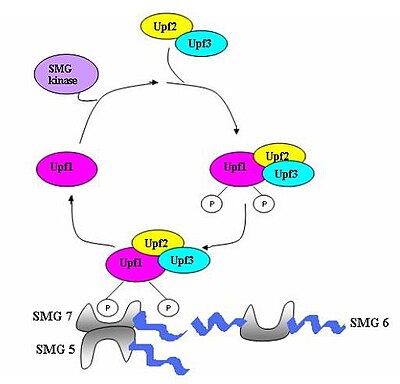
Umumiy nuqtai
Bema'ni vositachilik bilan parchalanish, muddatidan oldin tugatish kodonlarini (PTC) o'z ichiga olgan mRNA transkriptlarini aniqlash va parchalashda ishtirok etadi. PTClar turli xil mexanizmlar orqali hujayralarda paydo bo'lishi mumkin: urug'lanish DNKdagi mutatsiyalar; badandagi DNKdagi mutatsiyalar; xatolar transkripsiya; yoki transkripsiyadan keyingi mRNKni qayta ishlashdagi xatolar.[5][6] Ushbu mRNA transkriptlarini tanimaslik va parchalanmaslik organizm uchun zararli bo'lishi mumkin bo'lgan kesilgan oqsillarni ishlab chiqarishga olib kelishi mumkin. C-terminalining parchalanishiga olib keladi kesilgan polipeptidlar, NMD mexanizmi hujayralarni zararli narsalardan himoya qilishi mumkin dominant - salbiy va funktsiyani oshirish effektlar.[7] PTClar taxminan 30% ga tegishli meros qilib olingan kasalliklar; Shunday qilib, NMD yo'li organizmning umumiy omon qolishi va tayyorgarligini ta'minlashda muhim rol o'ynaydi[8][9]
Har xil oqsillardan (eRF1, eRF3, Upf1, Upf2 va Upf3) tashkil topgan kuzatuv kompleksi yig'ilib, mRNKni erta to'xtash kodonlari uchun tekshiradi.[5] Ushbu kompleksni yig'ish muddatidan oldin tarjimani to'xtatish bilan boshlanadi. Agar muddatidan oldin to'xtagan kodon aniqlansa, mRNK transkripti degradatsiyaga ishora qiladi - degradatsiya bilan aniqlashning birlashishi sodir bo'ladi.[3][10][11]
Ettita smg geni (smg1-7) va uchta UPF geni (Upf1-3) aniqlangan Saccharomyces cerevisiae va Caenorhabditis elegans NMD faoliyatiga hissa qo'shadigan muhim trans-ta'sir qiluvchi omillar sifatida.[12][13] Ushbu genlarning barchasi saqlanib qolgan yilda Drosophila melanogaster va boshqa sutemizuvchilar, ular NMDda ham muhim rol o'ynaydi. Eukaryotlarda NMD jarayonida saqlanadigan uchta komponent mavjud.[14] Bu Upf1 / SMG-2, Upf2 / SMG-3 va Upf3 / SMG-4 komplekslari. Upf1 / SMG-2 ko'p hujayrali organizmlarda fosfoprotein bo'lib, uning fosforillanish faolligi bilan NMDga hissa qo'shadi deb o'ylashadi. Biroq, hozirgi vaqtda oqsillarning o'zaro ta'siri va ularning NMDdagi rollari hozircha tortishuvlarga sabab bo'lgan.[11][12][14][15][16]
Sutemizuvchilardagi mexanizm
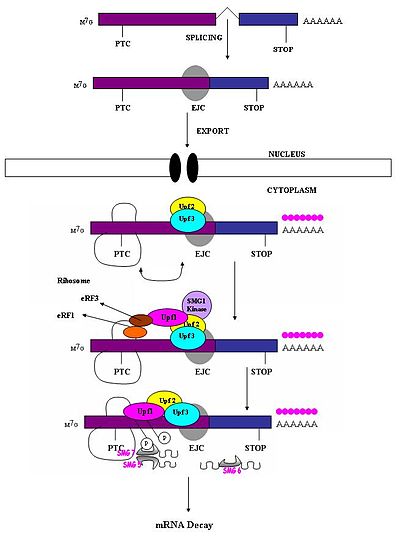
Vaqtdan oldin to'xtash kodoni odatdagi to'xtash kodonidan farq qilishi kerak, shunda faqat birinchisi NMD javobini keltirib chiqaradi. Bema'ni kodonning mRNK degradatsiyasini keltirib chiqarishi uning quyi oqim ketma-ketligi elementi va u bilan bog'liq oqsillarga nisbatan joylashishiga bog'liq ekanligi kuzatilgan.[1] Tadqiqotlar shuni ko'rsatdiki nukleotidlar oxirgi ekson-ekzon birikmasidan yuqori qismida 50-54 dan ortiq nukleotidlar parchalanish uchun mRNKni nishonga olishi mumkin.[1][4][5][6][7][17] Ushbu mintaqadan quyi oqimdagilar bunga qodir emaslar. Shunday qilib, bema'ni kodonlar oxirgisidan yuqorisida 50-54 dan ortiq nukleotid yotadi exon chegara, tabiiy to'xtash kodonlari terminal ekzonsida joylashgan.[18] Exon birlashma komplekslari (EJC) exon-exon chegaralarini belgilaydi. EJK - bu ko'paytirishli birikmalar bo'lib, ular birlashma birikmasidan yuqorida 20-24 nukleotid holatida birikish paytida yig'iladi.[19] Aynan ushbu EJC tabiiy to'xtatish kodonlaridan erta to'xtash kodonlarini ajratish uchun zarur bo'lgan pozitsiya ma'lumotlarini taqdim etadi. PTClarni tan olish ekzon-ekzon birikmalarining ta'riflariga bog'liq bo'lib tuyuladi. Bu splitseozomaning sutemizuvchilar NMDga aloqadorligini ko'rsatadi.[17][20] Tadqiqotlar sutemizuvchilarning NMD-ga splitseozomalar qo'shilish ehtimolini o'rganib chiqdi va bu ehtimollik ekanligini aniqladi.[18] Bundan tashqari, NMD mexanizmlari tabiiy ravishda intronlarni o'z ichiga olmaydigan genlardan hosil bo'ladigan bema'ni transkriptlar bilan faollashtirilmasligi kuzatilgan (ya'ni Histone H4, Hsp70, melanokortin-4-retseptorlari).[7]
Qachon ribosoma eRF1 va eRF3 tarjima omillari PTC ga etib boradi, ko'p proteinli ko'prik bo'lsa ham, saqlanib qolgan EJC komplekslari bilan o'zaro ta'sir qiladi.[21] Ning o'zaro ta'siri UPF1 tugatuvchi kompleks bilan va UPF2 / Saqlangan EJClarning UPF3-si juda muhimdir. Aynan shu o'zaro ta'sirlar mRNKni tezkor maqsadga yo'naltiradi yemirilish endogen nukleazalar tomonidan[18][21]

Umurtqasiz hayvonlardagi mexanizm
Kabi organizmlarni o'z ichiga olgan tadqiqotlar S. cerevisiae, D. melanogaster va C. elegans o'z ichiga olgan PTC tan olinishini ko'rsatdi umurtqasizlar organizmlar ekzon-ekson chegaralarini o'z ichiga olmaydi.[20] Ushbu tadqiqotlar shuni ko'rsatadiki, umurtqasizlar NMD birikishidan mustaqil ravishda paydo bo'ladi. Natijada, exon-exon chegaralarini belgilash uchun mas'ul bo'lgan EJClar umurtqasizlar NMDda talab qilinmaydi.[3] PTClarning umurtqasizlarda normal to'xtash kodonlaridan qanday farqlanishini tushuntirish uchun bir nechta modellar taklif qilingan. Ulardan biri sutemizuvchilarning ekzon birikmalariga o'xshash funktsiyalarni bajaradigan quyi oqimdagi ketma-ketlik elementi bo'lishi mumkinligini ko'rsatadi.[11] Ikkinchi model mRNA-da keng tarqalgan xususiyat, masalan, 3 'poly-A quyruq, tanib olish uchun zarur bo'lgan pozitsion ma'lumotni taqdim etishi mumkin.[22] "Faux 3'UTR modeli" deb nomlangan yana bir model, tarjimaning muddatidan oldin tugatilishini odatdagi tugatishdan farq qilishi mumkin, chunki u o'ziga xos bo'lmagan xususiyatlar tufayli uning mavjudligini tan olishga imkon beradi.[3] Biroq, ushbu mexanizmlar hali aniq namoyish etilmagan.
O'simliklardagi mexanizm
O'simliklarda PTC ni tanib olishning ikkita mexanizmi mavjud: uning EJK dan masofasi (umurtqali hayvonlar singari) yoki poli-A dumidan. O'simliklardagi NMD mexanizmi 300 nt dan uzunroq bo'lgan 3'NTR o'z ichiga olgan mRNKlarning parchalanishini keltirib chiqaradi, shuning uchun uzunroq 3'NTR bo'lgan mRNKlarning nisbati umurtqali hayvonlarga qaraganda o'simliklarda ancha past bo'ladi.[23][24]
NMD oldini olish
Odatda bema'ni mutatsiyalarga ega mRNAlar NMD yo'llari orqali parchalanishga qaratilgan deb o'ylashadi. Ekzon birikmasiga qadar 50-54 nts 5 'gacha bo'lgan bu erta to'xtash kodonining mavjudligi tez yemirilish uchun omil bo'lib ko'rinadi; ammo, erta to'xtash kodoni bo'lgan ba'zi mRNK molekulalari aniqlash va parchalanishdan qochishga qodir ekanligi kuzatilgan.[17][25] Umuman olganda, ushbu mRNK molekulalari o'qish ramkasida juda erta to'xtash kodoniga ega (ya'ni PTC AUG-proksimal). Bu hozirgi qabul qilingan NMD modeliga zid keladigan ko'rinadi, chunki bu pozitsiya exon-exon birikmasidan sezilarli darajada 5 'dir.[26]
Bu g-globulinda ko'rsatilgan. Genning birinchi ekzoni boshida bema'ni mutatsiyani o'z ichiga olgan b-globulin mRNKlari NMD sezgir mRNK molekulalariga qaraganda ancha barqarordir. Aniqlanishdan saqlanishning aniq mexanizmi hozircha ma'lum emas. Ushbu barqarorlikda poli-A majburiy oqsil (PABP) rol o'ynaydi degan fikrlar mavjud.[27] AUG-proksimal PTC-lar yaqinida ushbu oqsilning mavjudligi, aks holda NMD sezgir mRNA-larning barqarorligini kuchaytirishi boshqa tadqiqotlarda ham isbotlangan. Ushbu himoya effekti faqat b-globulin promouteri bilan chegaralanmasligi kuzatilgan.[25] Bu shuni ko'rsatadiki, ushbu NMD oldini olish mexanizmi boshqa genlar uchun boshqa to'qima turlarida keng tarqalgan bo'lishi mumkin. Keyingi tadqiqotlar davomida NMD ning amaldagi modeli qayta ko'rib chiqilishi kerak bo'lishi mumkin.
To'xtovsiz vositachilik qiladigan mRNA parchalanishi
Umumiy nuqtai
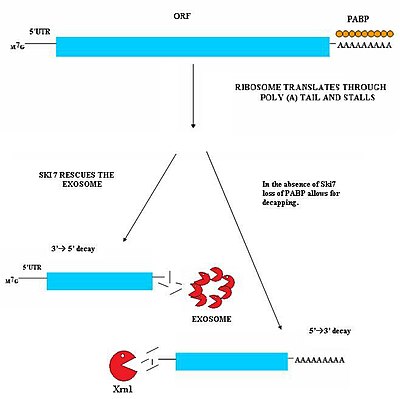
To'xtovsiz parchalanish (NSD) mRNA transkriptlarini aniqlash va parchalanishida ishtirok etadi, unda to'xtash kodoni yo'q.[29][30] Ushbu mRNA transkriptlari genning kodlash sohasidagi erta 3 'adenilatsiya yoki kriptik poliadenilatsiya signallari kabi turli xil mexanizmlardan kelib chiqishi mumkin.[31] Bu to'xtatish kodonining etishmasligi hujayralar uchun muhim muammoga olib keladi. MRNKni tarjima qiladigan ribosomalar oxir-oqibat 3'poly-A transkriptlar va stendlarning dumli qismiga aylanadi. Natijada, u mRNKni chiqara olmaydi.[32] Ribozomalar doimiy mRNK bilan bog'liq holda sekvestrga aylanishi mumkin va boshqa mRNA molekulalarini oqsillarga aylantirish mumkin bo'lmaydi. To'xtovsiz vositachilik bilan parchalanish bu muammoni to'xtab qolgan ribosomalarni bo'shatish va to'xtovsiz mRNKni hujayradagi nukleazalar bilan parchalanishi uchun belgilash orqali hal qiladi. To'xtovsiz vositachiligida parchalanish ikki xil yo'ldan iborat bo'lib, ular doimiy mRNKning parchalanishiga mos ravishda harakat qilishadi.[29][30]
Chang‘i7 yo‘li
Ushbu yo'l hujayrada Ski7 oqsili mavjud bo'lganda faol bo'ladi. Ski7 oqsili ribosomaning bo'sh joyiga bog'langan deb o'ylashadi. Ushbu bog'lanish ribosomani tiqilib qolgan mRNK molekulasini chiqarib tashlashga imkon beradi - bu hatto ribosomani bo'shatadi va boshqa transkriptlarni tarjima qilishga imkon beradi. Ski7 endi to'xtovsiz mRNA bilan bog'langan va aynan shu assotsiatsiya sitosolik tomonidan tan olinishi uchun uzluksiz mRNA ni maqsad qiladi. ekzozom. Ski7-ekzozoma kompleksi mRNK molekulasini tezda dezenil qiladi, bu esa ekzosomaning transkriptni 3 'dan 5' gacha parchalanishiga imkon beradi.[29][30]
Ski7 bo'lmagan yo'l
Xamirturushda NSD ning ikkinchi turi kuzatilgan. Ushbu mexanizmda Ski7 ning yo'qligi translyatsiya ribosomasi ta'sirida poli-A dumini bog'laydigan PABP oqsillarini yo'qotishiga olib keladi. Ushbu PABP oqsillarini olib tashlash keyinchalik himoya vositasini yo'qotishiga olib keladi 5'm7G qopqoq. Qopqoqning yo'qolishi transkriptning XrnI kabi endogen 5'-3 'ekzonukleaza bilan tezda degradatsiyasiga olib keladi.[30]
Yo'q, parchalanish

No-Go parchalanishi (NGD) - bu so'nggi kuzatuv mexanizmi.[33] Shunday qilib, hozirda u yaxshi tushunilmagan. NGD-ning haqiqiy maqsadlari yaxshi tushunilmagan bo'lsa-da, ular asosan ribosomalar tarjima paytida to'xtab qolgan mRNA transkriptlaridan iborat. Ushbu to'xtash joyiga turli xil omillar sabab bo'lishi mumkin, shu jumladan kuchli ikkilamchi tuzilmalar, bu translyatsiya texnikasini transkriptda pastga siljishini jismoniy to'sib qo'yishi mumkin.[33] Dom34 / Hbs1 to'xtab qolgan ribosomalar joylashgan joy yaqinida bog'lanib, komplekslarni qayta ishlashini osonlashtirishi mumkin.[34] Ba'zi hollarda stenogramma stend joyi yonida endonukleolitik usulda ham kesilgan; ammo mas'ul endonukleazning kimligi bahsli bo'lib qolmoqda. The parchalangan Keyin mRNA molekulalari 3 'dan 5' gacha bo'lgan shaklda va Xrn1 tomonidan 5 'dan 3' gacha bo'lgan shaklda ekzosoma tomonidan to'liq parchalanadi.[33]Hozirgi vaqtda ushbu jarayon mRNKni ribosomalardan qanday chiqarishi ma'lum emas, ammo Hbs1 Ski7 vositachiligidagi NSDda ribosomalar chiqarilishida aniq rol o'ynaydigan Ski7 oqsillari bilan chambarchas bog'liq. Hbs1 NGD da xuddi shunday rol o'ynashi mumkinligi taxmin qilingan.[5][35]
Evolyutsiya
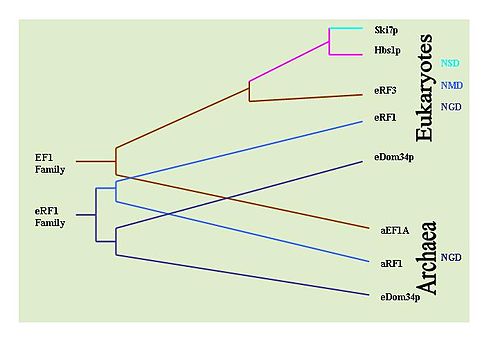
Ni aniqlash mumkin evolyutsion har bir mexanizmga taalluqli bo'lgan asosiy proteinlarning saqlanishini kuzatish orqali ushbu mexanizmlarning tarixi. Masalan: Dom34 / Hbs1 NGD bilan bog'langan;[33] Ski7 NSD bilan bog'liq;[29] va eRF oqsillari NMD bilan bog'liq.[6] Shu maqsadda keng qamrovli Portlash turli xil organizmlarda oqsillarning tarqalishini aniqlash uchun izlanishlar olib borildi. NGD Hbs1 va NMD eRF3 faqat eukaryotlarda bo'lganligi aniqlandi. Biroq, NGD Dom34 eukaryotlarda va universaldir arxey. Bu shuni ko'rsatadiki, NGD birinchi bo'lib rivojlangan mRNA kuzatuv mexanizmi bo'lgan. NSD Ski7 oqsillari xamirturush turlari bilan cheklangan ko'rinadi, bu esa NSD eng so'nggi rivojlangan kuzatuv mexanizmi ekanligini ko'rsatmoqda. Bu sukut bo'yicha NMD ni ikkinchi rivojlangan kuzatuv mexanizmi sifatida qoldiradi.[36]
Adabiyotlar
- ^ a b v Amrani N, Sachs MS, Jacobson A (iyun 2006). "Erta bema'nilik: mRNA parchalanishi tarjima muammosini hal qiladi". Tabiat sharhlari. Molekulyar hujayra biologiyasi. 7 (6): 415–25. doi:10.1038 / nrm1942. PMID 16723977.
- ^ Mur MJ (sentyabr 2005). "Tug'ilishdan o'limgacha: eukaryotik mRNKlarning murakkab hayoti". Ilm-fan. 309 (5740): 1514–8. Bibcode:2005 yil ... 309.1514M. doi:10.1126 / science.1111443. PMID 16141059.
- ^ a b v d e Amrani N, Ganesan R, Kervestin S, Mangus DA, Ghosh S, Jacobson A (Noyabr 2004). "Soxta 3'-UTR aberrant tugatishni rag'batlantiradi va bema'nilik vositachiligidagi mRNA parchalanishini keltirib chiqaradi". Tabiat. 432 (7013): 112–8. Bibcode:2004 yil Tabiat. 432..112A. doi:10.1038 / tabiat03060. PMID 15525991.
- ^ a b v Fasken MB, Corbett AH (iyun 2005). "Jarayon yoki nobud bo'lish: mRNA biogenezidagi sifat nazorati". Tabiatning strukturaviy va molekulyar biologiyasi. 12 (6): 482–8. doi:10.1038 / nsmb945. PMID 15933735.
- ^ a b v d e f g Chang YF, Imom JS, Uilkinson MF (2007). "Bema'nilik vositachiligida parchalanadigan RNKni kuzatish yo'li". Biokimyo fanining yillik sharhi. 76: 51–74. doi:10.1146 / annurev.biochem.76.050106.093909. PMID 17352659.
- ^ a b v Rehwinkel J, Raes J, Izaurralde E (2006 yil noyabr). "Bema'nilik vositachiligidagi mRNA parchalanishi: maqsadli genlar va effektorlarning funktsional diversifikatsiyasi". Biokimyo fanlari tendentsiyalari. 31 (11): 639–46. doi:10.1016 / j.tibs.2006.09.005. PMID 17010613.
- ^ a b v Maquat LE (2004 yil fevral). "Bema'nilik vositachiligidagi mRNA parchalanishi: qo'shilish, tarjima va mRNP dinamikasi". Tabiat sharhlari. Molekulyar hujayra biologiyasi. 5 (2): 89–99. doi:10.1038 / nrm1310. PMID 15040442.
- ^ Xolbruk JA, Neu-Yilik G, Xentze MW, Kulozik AE (2004 yil avgust). "Klinikaga bema'nilik vositachiligida buzilish yaqinlashmoqda". Tabiat genetikasi. 36 (8): 801–8. doi:10.1038 / ng1403. PMID 15284851.
- ^ Mendell JT, Sharifi NA, Meyers JL, Martinez-Murillo F, Dietz HC (oktyabr 2004). "Bema'nilik nazorati sutemizuvchilar transkriptlarining turli sinflarini ifodalashni tartibga soladi va genomik shovqinni o'chiradi". Tabiat genetikasi. 36 (10): 1073–8. doi:10.1038 / ng1429. PMID 15448691.
- ^ Lejeune F, Maquat LE (iyun 2005). "Nonsens vositachiligida mRNA parchalanishi va sutemizuvchi hujayralardagi mRNK qo'shilishining mexanik aloqalari". Hujayra biologiyasidagi hozirgi fikr. 17 (3): 309–15. doi:10.1016 / j.ceb.2005.03.002. PMID 15901502.
- ^ a b v Conti E, Izaurralde E (iyun 2005). "Bema'nilik vositachiligidagi mRNA parchalanishi: molekulyar tushunchalar va turlar bo'yicha mexanik o'zgarishlar". Hujayra biologiyasidagi hozirgi fikr. 17 (3): 316–25. doi:10.1016 / j.ceb.2005.04.005. PMID 15901503.
- ^ a b Cali BM, Kuchma SL, Latham J, Anderson P (1999 yil fevral). "Caenorhabditis elegans-da mRNA nazorati uchun smg-7 kerak". Genetika. 151 (2): 605–16. PMC 1460488. PMID 9927455.
- ^ Yamashita A, Kashima I, Ohno S (dekabr 2005). "SMG-1ning bema'nilik vositachiligidagi mRNA yemirilishidagi roli". Biochimica et Biofhysica Acta (BBA) - Oqsillar va Proteomikalar. 1754 (1–2): 305–15. doi:10.1016 / j.bbapap.2005.10.002. PMID 16289965.
- ^ a b Kim YK, Furic L, Desgroseillers L, Maquat LE (yanvar 2005). "Sutemizuvchi Staufen1 mRNK parchalanishini keltirib chiqarish uchun Upf1 ni o'ziga xos mRNA 3'UTRsga yollaydi". Hujayra. 120 (2): 195–208. doi:10.1016 / j.cell.2004.11.050. PMID 15680326.
- ^ Longman D, Plasterk RH, Johnstone IL, Cáceres JF (may 2007). "Mexanik tushunchalar va C. elegans NMD yo'lidagi ikkita yangi omilni aniqlash". Genlar va rivojlanish. 21 (9): 1075–85. doi:10.1101 / gad.417707. PMC 1855233. PMID 17437990.
- ^ Gatfield D, Unterholzner L, Ciccarelli FD, Bork P, Izaurralde E (avgust 2003). "Drozofilada bema'nilik vositachiligidagi mRNK yemirilishi: xamirturush va sutemizuvchilar yo'llari kesishmasida". EMBO jurnali. 22 (15): 3960–70. doi:10.1093 / emboj / cdg371. PMC 169044. PMID 12881430.
- ^ a b v Nagy E, Maquat LE (iyun 1998). "Intron o'z ichiga olgan genlar tarkibidagi tugatish-kodon pozitsiyasi qoidasi: bema'nilik RNKning ko'pligiga ta'sir qilganda". Biokimyo fanlari tendentsiyalari. 23 (6): 198–9. doi:10.1016 / S0968-0004 (98) 01208-0. PMID 9644970.
- ^ a b v Aktyor TJ, Mills DJ, Horton AA (1979 yil iyun). "DAB tomonidan qo'zg'atilgan gepatomalardan mikrosomal fraktsiya va ajratib olingan mikrosomal lipidlarning lipid peroksidatsiyasi". Britaniya saraton jurnali. 39 (6): 773–8. doi:10.1128 / mcb.18.9.5272. PMC 109113. PMID 9710612.
- ^ Neu-Yilik G, Gehring NH, Thermann R, Frede U, Xentze MW, Kulozik AE (2001 yil fevral). "Bezovta vositachiligi bilan parchalanish qobiliyatiga ega bo'lgan inson beta-globin mRNPlarini aniqlashda qo'shilish va 3 'ni shakllantirish". EMBO jurnali. 20 (3): 532–40. doi:10.1093 / emboj / 20.3.532. PMC 133467. PMID 11157759.
- ^ a b Behm-Ansmant I, Gatfild D, Rehwinkel J, Hilgers V, Izaurralde E (mart 2007). "Bema'nilik vositachiligidagi mRNA parchalanishida sitoplazmatik poli (A) bilan bog'langan oqsil 1 (PABPC1) uchun saqlanib qolgan rol". EMBO jurnali. 26 (6): 1591–601. doi:10.1038 / sj.emboj.7601588. PMC 1829367. PMID 17318186.
- ^ a b Kashima I, Yamashita A, Izumi N, Kataoka N, Morishita R, Hoshino S, Ohno M, Dreyfuss G, Ohno S (2006 yil fevral). "Yangi SMG-1-Upf1-eRF1-eRF3 kompleksining (SURF) ekzon birikmasi kompleksiga ulanishi Upf1 fosforillanishini va bema'nilik vositachiligidagi mRNA parchalanishini keltirib chiqaradi". Genlar va rivojlanish. 20 (3): 355–67. doi:10.1101 / gad.1389006. PMC 1361706. PMID 16452507.
- ^ Palanisvami V, Moraes KC, Uilush CJ, Uilush J (may 2006). "Nukleofosmin poliadenilatlanish jarayonida tanlab mRNKga yotqiziladi". Tabiatning strukturaviy va molekulyar biologiyasi. 13 (5): 429–35. doi:10.1038 / nsmb1080. PMC 2811576. PMID 16604083.
- ^ Shvarts AM, Komarova TV, Skulachev M.V., Zvereva AS, Doroxov I, Atabekov JG (2006 yil dekabr). "O'simliklar mRNKlarining barqarorligi 3'-tarjima qilinmagan mintaqaning uzunligiga bog'liq". Biokimyo. Biokimiya. 71 (12): 1377–84. doi:10.1134 / s0006297906120145. PMID 17223792.
- ^ Nyiko T, Kerényi F, Szabadkai L, Benkovics AH, mayor P, Sonkoly B, Mérai Z, Barta E, Niemiec E, Kufel J, Silhavy D (iyul 2013). "O'simliklarning bema'ni vositachiligida mRNK parchalanishi turli xil avtoregulyatsiya davrlari tomonidan boshqariladi va EJCga o'xshash kompleks tomonidan chaqirilishi mumkin". Nuklein kislotalarni tadqiq qilish. 41 (13): 6715–28. doi:10.1093 / nar / gkt366. PMC 3711448. PMID 23666629.
- ^ a b Inacio A, Silva AL, Pinto J, Ji X, Morgado A, Almeyda F, Faustino P, Lavinha J, Liebhaber SA, Romão L (iyul 2004). "Boshlanish kodoniga yaqin bo'lgan bema'ni mutatsiyalar to'liq bema'nilik vositachiligidagi mRNA parchalanishini keltirib chiqarmaydi". Biologik kimyo jurnali. 279 (31): 32170–80. doi:10.1074 / jbc.m405024200. PMID 15161914.
- ^ Silva AL, Pereyra FJ, Morgado A, Kong J, Martins R, Faustino P, Libeber SA, Romão L (dekabr 2006). "Kanonik UPF1-ga bog'liq bema'nilik vositachiligidagi mRNA parchalanishi ketma-ketlik kontekstidan mustaqil ravishda qisqa o'qish doirasini o'z ichiga olgan transkriptlarda inhibe qilinadi". RNK. 12 (12): 2160–70. doi:10.1261 / rna.201406. PMC 1664719. PMID 17077274.
- ^ Silva AL, Ribeiro P, Inacio A, Liebhaber SA, Romão L (mart 2008). "Poli (A) bilan bog'langan oqsilning muddatidan oldin tugagan kodonga yaqinligi sutemizuvchilarning bema'nilik vositasi bo'lgan mRNK yemirilishini inhibe qiladi". RNK. 14 (3): 563–76. doi:10.1261 / rna.815108. PMC 2248256. PMID 18230761.
- ^ Garneau NL, Wilushz J, Wilusz CJ (2007 yil fevral). "MRNA parchalanishining magistral yo'llari va shosseslari". Tabiat sharhlari. Molekulyar hujayra biologiyasi. 8 (2): 113–26. doi:10.1038 / nrm2104. PMID 17245413.
- ^ a b v d van Hoof A, Frischmeyer PA, Dietz HC, Parker R (2002 yil mart). "Tugatish kodoniga ega bo'lmagan mRNKlarning ekzosoma vositasida tan olinishi va degradatsiyasi". Ilm-fan. 295 (5563): 2262–4. doi:10.1126 / science.1067272. PMID 11910110.
- ^ a b v d Frischmeyer PA, van Hoof A, O'Donnell K, Gerrerio AL, Parker R, Dietz HC (mart 2002). "Tugatish kodonlari bo'lmagan transkriptlarni yo'q qiladigan mRNA kuzatuv mexanizmi". Ilm-fan. 295 (5563): 2258–61. Bibcode:2002 yil ... 295.2258F. doi:10.1126 / science.1067338. PMID 11910109.
- ^ Temperley RJ, Seneca SH, Tonska K, Bartnik E, Bindoff LA, Lightowlers RN, Chrzanowska-Lightowlers ZM (sentyabr 2003). "Patogen mtDNA mikrodeletsiyasini o'rganish natijasida odam mitoxondriyasida tarjimaga bog'liq bo'lgan dedenilatsiyaning parchalanish yo'li aniqlanadi". Inson molekulyar genetikasi. 12 (18): 2341–8. doi:10.1093 / hmg / ddg238. PMID 12915481.
- ^ Karzay AW, Roche ED, Sauer RT (iyun 2000). "SsrA-SmpB tizimi oqsillarni belgilash, degradatsiyalash va ribosomalarni qutqarish". Tabiatning strukturaviy biologiyasi. 7 (6): 449–55. doi:10.1038/75843. PMID 10881189.
- ^ a b v d Doma MK, Parker R (2006 yil mart). "Eukaryotik mRNKlarning endonukleolitik parchalanishi, tarjimani cho'zishda stendlar bilan". Tabiat. 440 (7083): 561–4. Bibcode:2006 yil natur.440..561D. doi:10.1038 / tabiat04530. PMC 1839849. PMID 16554824.
- ^ Kobayashi K, Kikuno I, Kuroha K, Saito K, Ito K, Ishitani R, Inada T, Nureki O (oktyabr 2010). "Pelota va GTP bilan bog'langan EF1a kompleksi tomonidan mRNA nazorati uchun tizimli asos". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 107 (41): 17575–9. Bibcode:2010PNAS..10717575K. doi:10.1073 / pnas.1009598107. PMC 2955123. PMID 20876129.
- ^ Gril M, Chaylet M, van Tilberg H (mart 2008). "Dom34 xamirturushining tuzilishi: tarjimani tugatish omiliga bog'liq oqsil va Er-1 parchalanishiga aralashadi". Biologik kimyo jurnali. 283 (11): 7145–54. doi:10.1074 / jbc.M708224200. PMID 18180287.
- ^ a b Atkinson GC, Baldauf SL, Hauryliuk V (oktyabr 2008). "To'xtovsiz, ishlamaslik va bema'nilik vositachiligidagi mRNK yemirilishining evolyutsiyasi va ularning tugash omillaridan kelib chiqqan komponentlar". BMC evolyutsion biologiyasi. 8: 290. doi:10.1186/1471-2148-8-290. PMC 2613156. PMID 18947425.