Asabiy tebranish - Neural oscillation - Wikipedia
Asabiy tebranishlar, yoki miya to'lqinlari, bu asabiy faoliyatning ritmik yoki takrorlanadigan naqshlari markaziy asab tizimi. Asab to'qimasi yaratishi mumkin salınımlı faoliyat ko'p jihatdan, yoki individual mexanizmlar tomonidan boshqariladi neyronlar yoki neyronlarning o'zaro ta'sirida. Ayrim neyronlarda tebranishlar yoki tebranishlar sifatida ko'rinishi mumkin membrana potentsiali yoki ning ritmik naqshlari sifatida harakat potentsiali, keyinchalik tebranuvchi aktivatsiyasini hosil qiladi post-sinaptik neyronlar. Darajasida asabiy ansambllar, ko'p sonli neyronlarning sinxronlashgan faolligi paydo bo'lishi mumkin makroskopik tebranishlari, bularni an kuzatilishi mumkin elektroansefalogramma. Neyronlar guruhlaridagi tebranuvchi faollik, odatda, neyronlarning o'zaro bog'liqliklaridan kelib chiqadi, natijada ularning otish rejimlarini sinxronlashtirishga olib keladi. Neyronlarning o'zaro ta'siri individual neyronlarning otish chastotasidan farqli ravishda chastotada tebranishlarni keltirib chiqarishi mumkin. Makroskopik asab salınımlarının taniqli misoli alfa faoliyati.
Nerv tebranishlari tadqiqotchilar tomonidan 1924 yildayoq kuzatilgan (tomonidan Xans Berger ). 50 yildan ko'proq vaqt o'tgach, ichki salınımlı xatti-harakatlar umurtqali hayvonlar neyronlarida uchradi, ammo uning funktsional roli hali ham to'liq tushunilmagan.[1] Asabiy tebranishlarning mumkin bo'lgan rollariga quyidagilar kiradi xususiyati majburiy, axborot uzatish mexanizmlari va ritmik vosita chiqishi avlodi. So'nggi o'n yilliklarda, ayniqsa, yutuqlar bilan ko'proq tushuncha hosil bo'ldi miya tasviri. Tadqiqotning asosiy yo'nalishi nevrologiya tebranishlar qanday vujudga kelishini va ularning qanday rollarni bajarishini aniqlashni o'z ichiga oladi. Miyadagi tebranish faolligi har xil darajada keng kuzatiladi tashkilot darajalari va asabiy ma'lumotlarni qayta ishlashda muhim rol o'ynaydi deb o'ylashadi. Ko'p sonli eksperimental tadqiqotlar asab tebranishlarining funktsional rolini qo'llab-quvvatlaydi; ammo birlashtirilgan talqin hali ham etishmayapti.

Tarix
Richard Katon quyonlar va maymunlarning miya yarim sharlarida elektr faolligini aniqladi va 1875 yilda o'z xulosalarini taqdim etdi.[2] Adolf Bek 1890 yilda uning quyonlar va itlarning miyasining o'z-o'zidan paydo bo'lgan elektr faolligi haqidagi kuzatuvlari nashr etilgan bo'lib, ular miyaning yuzasiga to'g'ridan-to'g'ri joylashtirilgan elektrodlar bilan aniqlangan yorug'lik ta'sirida o'zgargan ritmik tebranishlarni o'z ichiga olgan.[3] Xans Bergerdan oldin, Vladimir Vladimirovich Pravdich-Neminskiy birinchi hayvon EEG va uyg'ongan potentsial itning.[4]
Umumiy nuqtai
Nerv tebranishlari markaziy asab tizimida barcha darajalarda kuzatiladi va o'z ichiga oladi boshoqli poezdlar, mahalliy dala salohiyati va keng ko'lamli tebranishlar tomonidan o'lchanishi mumkin elektroensefalografiya (EEG). Umuman olganda, tebranishlarni ular bilan xarakterlash mumkin chastota, amplituda va bosqich. Ushbu signal xususiyatlari yordamida neyron yozuvlaridan olinishi mumkin vaqt chastotasini tahlil qilish. Katta miqyosli tebranishlarda amplituda o'zgarishlar a ichida sinxronizatsiya o'zgarishi natijasida kelib chiqadi deb hisoblanadi asab ansambli, shuningdek, mahalliy sinxronizatsiya deb nomlanadi. Mahalliy sinxronizatsiya bilan bir qatorda, uzoq nerv tuzilmalarining (yakka neyronlar yoki asab ansambllari) tebranuvchi faolligi ham sinxronlashi mumkin. Asabiy tebranishlar va sinxronizatsiya ma'lumot uzatish, idrok etish, motorni boshqarish va xotira kabi ko'plab bilim funktsiyalari bilan bog'liq.[5][6][7]
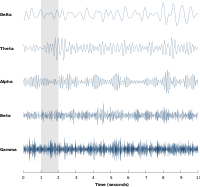
Neyronlarning tebranishlari katta neyron guruhlari tomonidan hosil qilingan nerv faoliyatida eng ko'p o'rganilgan. Katta miqyosdagi faoliyatni EEG kabi usullar bilan o'lchash mumkin. Umuman olganda, EEG signallari o'xshash keng spektral tarkibga ega pushti shovqin, shuningdek, ma'lum chastota diapazonlarida tebranish faolligini aniqlaydi. Birinchi kashf qilingan va eng taniqli chastota diapazoni alfa faoliyati (8–12 Hz )[8] dan aniqlanishi mumkin oksipital lob tinch uyg'onish paytida va bu ko'zlar yopilganda ko'payadi.[9] Boshqa chastota diapazonlari: delta (1-4 Hz), teta (4-8 Hz), beta-versiya (13-30 Hz), kam gamma (30-70 Hz) va yuqori gamma (70-150 Hz) chastota diapazonlari, bu erda gamma faolligi kabi tezroq ritmlar kognitiv ishlov berish bilan bog'liq. Darhaqiqat, EEG signallari uxlash vaqtida keskin o'zgaradi va tezroq chastotalardan alfa to'lqinlar kabi tobora sekinroq chastotalarga o'tishni ko'rsatadi. Aslida, turli xil uyqu bosqichlari odatda spektral tarkibi bilan tavsiflanadi.[10] Binobarin, asabiy tebranishlar kognitiv holatlar bilan bog'liq bo'lib, masalan xabardorlik va ong.[11][12]
Insonning miya faoliyatidagi asabiy tebranishlar asosan EEG yozuvlari yordamida tekshirilsa-da, ular invaziv qayd qilish texnikasi yordamida kuzatiladi. bitta birlik yozuvlar. Neyronlar ritmik naqshlarni yaratishi mumkin harakat potentsiali yoki boshoq. Ba'zi bir neyron turlari ma'lum chastotalarda otish xususiyatiga ega rezonatorlar.[13] Bursting ritmik pog'onaning yana bir shakli. Spiking naqshlari asosiy deb hisoblanadi ma'lumotlarni kodlash miyada. Salınımcılığını, shaklida ham kuzatilishi mumkin pastki ostidagi membrana potentsial tebranishlari (ya'ni harakat potentsiali bo'lmagan taqdirda).[14] Agar ko'plab neyronlar paydo bo'lsa sinxronizatsiya, ular tebranishlarni keltirib chiqarishi mumkin mahalliy dala salohiyati. Miqdoriy modellar yozib olingan ma'lumotlarda asabiy salınımların kuchini taxmin qilishlari mumkin.[15]
Asabiy tebranishlar odatda matematik doiradan o'rganiladi va "neyrodinamika" sohasiga tegishli bo'lib, kognitiv fanlar Bu tasvirlashda asab faoliyatining dinamik xususiyatiga katta e'tibor qaratadi miya funktsiya.[16] Bu miyani ko'rib chiqadi a dinamik tizim va foydalanadi differentsial tenglamalar asab faoliyati vaqt o'tishi bilan qanday rivojlanib borishini tasvirlash. Xususan, bu miya faoliyatining dinamik naqshlarini idrok va xotira kabi kognitiv funktsiyalar bilan bog'lashga qaratilgan. Juda mavhum shakl, asabiy tebranishlarni tahlil qilish mumkin analitik ravishda. Fiziologik jihatdan aniqroq sharoitda o'rganilganda, tebranish harakati odatda o'rganib o'rganiladi kompyuter simulyatsiyalari a hisoblash modeli.
Asabiy tebranishlarning funktsiyalari keng ko'lamli va turli xil tebranish faoliyat turlarida turlicha. Masalan, a kabi ritmik faoliyatni yaratish yurak urishi va asab bilan bog'lanish narsaning shakli va rangi kabi idrokdagi sezgir xususiyatlarning. Ko'pchilikda asabiy tebranishlar ham muhim rol o'ynaydi asab kasalliklari, paytida haddan tashqari sinxronizatsiya kabi soqchilik faoliyat epilepsiya yoki titroq bo'lgan bemorlarda Parkinson kasalligi. Tebranish harakati a kabi tashqi qurilmalarni boshqarish uchun ham ishlatilishi mumkin miya-kompyuter interfeysi.[17]
Fiziologiya
Tebranish harakati butun davomida kuzatiladi markaziy asab tizimi tashkilotning barcha darajalarida. Uch xil daraja keng miqyosda tan olingan: mikro miqyos (bitta neyronning faoliyati), mezo-shkalasi (mahalliy neyronlar guruhining faolligi) va makrosalasi (turli miya mintaqalarining faoliyati).[18]
Mikroskopik
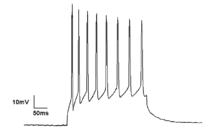
Neyronlar hosil qiladi harakat potentsiali elektr membrana potentsialining o'zgarishi natijasida paydo bo'ladi. Neyronlar boshoqli poezdlar deb nomlangan ketma-ketlikda bir nechta harakat potentsialini yaratishi mumkin. Ushbu boshoqli poezdlar uchun asosdir asab kodlash va miyada axborot uzatish. Spike poezdlari barcha turdagi naqshlarni yaratishi mumkin, masalan, ritmik pog'ona va yorilish va ko'pincha tebranish faolligini namoyish etadi.[19] Yagona neyronlarda tebranish faolligi ham kuzatilishi mumkin ostonadagi tebranishlar membrana potentsialida. Membrana potentsialidagi ushbu ritmik o'zgarishlar kritik chegaraga etib bormaydi va shuning uchun harakat potentsialiga olib kelmaydi. Ular sinxron kirishlardan postsinaptik potentsial yoki neyronlarning ichki xususiyatlaridan kelib chiqishi mumkin.
Neyronik pog'onani ularning faoliyat uslublari bo'yicha tasniflash mumkin. Neyronlarning qo'zg'aluvchanligini I va II sinflarga bo'lish mumkin. I sinf neyronlar kirish kuchiga qarab o'zboshimchalik bilan past chastotali harakat potentsialini yaratishi mumkin, II sinf neyronlar ma'lum chastota diapazonida harakat potentsialini hosil qiladi, bu kirish kuchining o'zgarishiga nisbatan befarq.[13] II darajali neyronlar, shuningdek, membrana potentsialida pastki pol tebranishlarini namoyon etishga moyil.
Mezoskopik
Bir guruh neyronlar ham tebranuvchi faollikni vujudga keltirishi mumkin. Sinaptik o'zaro ta'sirlar natijasida turli xil neyronlarning otish naqshlari sinxronlashtirilishi mumkin va ularning ta'sir potentsiali tufayli elektr potentsialidagi ritmik o'zgarishlar qo'shiladi (konstruktiv aralashuv ). Boshqacha aytganda, sinxronlashtirilgan otishni o'rganish sxemalari boshqa kortikal sohalarga sinxronlashtirilgan kirishga olib keladi, bu esa katta amplituda tebranishlarni keltirib chiqaradi. mahalliy dala salohiyati. Ushbu keng ko'lamli tebranishlarni bosh terisi tashqarisida ham o'lchash mumkin elektroensefalografiya (EEG) va magnetoensefalografiya (MEG). Yagona neyronlar tomonidan ishlab chiqarilgan elektr potentsiallari bosh terisi tashqarisida to'planish uchun juda kichikdir va EEG yoki MEG faolligi har doim o'xshash fazoviy yo'nalishga ega bo'lgan minglab yoki millionlab neyronlarning sinxron faolligi yig'indisini aks ettiradi.[20] A neyronlari asab ansambli kamdan-kam hollarda olov bir vaqtning o'zida, ya'ni to'liq sinxronlashtiriladi. Buning o'rniga, otish ehtimoli ritmik ravishda modulyatsiya qilingan bo'lib, neyronlarning bir vaqtning o'zida otish ehtimoli yuqori, bu ularning o'rtacha faolligida tebranishlarni keltirib chiqaradi (sahifaning yuqori qismidagi rasmga qarang). Shunday qilib, ning chastotasi keng ko'lamli tebranishlarning individual neyronlarning otish tartibiga mos kelishi shart emas. Izolyatsiya qilingan kortikal neyronlar ma'lum sharoitlarda muntazam ravishda otishadi, ammo buzilmagan miyada kortikal hujayralar juda o'zgaruvchan sinaptik kirishlar bilan bombardimon qilinadi va odatda tasodifiy ko'rinadi. Ammo, agar neyronlarning katta guruhining ehtimoli umumiy chastotada ritmik ravishda modulyatsiya qilingan bo'lsa, ular o'rtacha maydonda tebranishlar hosil qiladi (shuningdek, sahifaning yuqori qismidagi rasmga qarang).[19] Asabiy ansambllar salınımlı faoliyatni yaratishi mumkin endogen ravishda qo'zg'atuvchi va inhibitor neyronlarning mahalliy o'zaro ta'siri orqali. Xususan, inhibitiv internironlar samarali qo'zg'alish uchun tor oynani yaratish va qo'zg'atuvchi neyronlarning otish tezligini ritmik ravishda modulyatsiya qilish orqali asab ansambli sinxroniyasini yaratishda muhim rol o'ynaydi.[21]
Makroskopik
Asabiy tebranish, shuningdek, struktura orqali bog'langan turli miya sohalari o'rtasidagi o'zaro ta'sirlardan kelib chiqishi mumkin yoqimli. Vaqt kechikmoqda bu erda muhim rol o'ynaydi. Miyaning barcha sohalari ikki tomonlama bog'langanligi sababli, miya sohalari orasidagi bu aloqalar shakllanadi mulohaza ko'chadan. Ijobiy mulohaza tsikllar chastotaning kechikish vaqti bilan teskari bog'liq bo'lgan joylarda tebranish faolligini keltirib chiqaradi. Bunday teskari aloqa tsiklining misoli talamus va korteks - the talamokortikal nurlanishlar. Ushbu talamokortikal tarmoq deb nomlanuvchi salınımsal faoliyatni yaratishga qodir takroriy talamo-kortikal rezonans.[22] Talamokortikal tarmoq avlodni yaratishda muhim rol o'ynaydi alfa faoliyati.[23][24] Butun miya tarmog'idagi modelda anatomik bog'lanish va miya hududlari orasidagi tarqalish kechikishi, tebranishlar beta chastota diapazoni gamma-diapazonda tebranib turadigan (mezoskopik darajada hosil bo'lgan) miya hududlarining pastki qismlarini qisman sinxronlashidan kelib chiqadi.[25]
Mexanizmlar
Neyron xususiyatlari
Olimlar ba'zi bir ichki narsalarni aniqladilar neyronal xususiyatlar membrana potentsial tebranishini hosil qilishda muhim rol o'ynaydi. Jumladan, kuchlanishli ionli kanallar harakat potentsialini yaratishda juda muhimdir. Ushbu ion kanallarining dinamikasi yaxshi yo'lga qo'yilgan Xojkin-Xaksli modeli Differentsial tenglamalar to'plami yordamida harakat potentsiallari qanday boshlanishi va tarqalishini tavsiflovchi. Foydalanish bifurkatsiya tahlili, ushbu neyronal modellarning turli xil salınımlı navlarini aniqlash mumkin, bu neyronlarning reaktsiyalari turlarini tasniflashga imkon beradi. Xodkin-Xaksli modelida aniqlangan neyronlar pog'onasining tebranuvchi dinamikasi empirik topilmalar bilan chambarchas mos keladi. Vaqti-vaqti bilan bosishdan tashqari, pastki ostidagi membrana potentsial tebranishlari, ya'ni rezonans harakat potentsialiga olib kelmaydigan xatti-harakatlar qo'shni neyronlarning sinxron faolligini osonlashtirish orqali tebranuvchi faollikka ham hissa qo'shishi mumkin.[26][27] Elektron yurak stimulyatori neyronlari singari markaziy naqsh generatorlari, kortikal hujayralar subtiplari boshoqlarning portlashlarini (boshoqlarning qisqa klasterlari) ritmik ravishda afzal qilingan chastotalarda yong'in chiqaradi. Bursting neyronlari sinxron tarmoq tebranishlari uchun yurak stimulyatori sifatida xizmat qilish imkoniyatiga ega va boshoq portlashlari neyron rezonansi asosida bo'lishi yoki kuchayishi mumkin.[19]
Tarmoq xususiyatlari
Neyronlarning ichki xususiyatlaridan tashqari, biologik asab tarmog'i xususiyatlari, shuningdek, tebranish faolligining muhim manbai hisoblanadi. Neyronlar muloqot qilish sinapslar orqali bir-birlari bilan va post-sinaptik neyronlarda boshoqli poezdlarning harakatlanish vaqtiga ta'sir qiladi. Ulanishning xususiyatlariga, masalan, ulanish kuchi, vaqtni kechiktirish va ulanishning bog'liqligi hayajonli yoki inhibitiv, o'zaro ta'sir qiluvchi neyronlarning boshoqli poezdlari aylanishi mumkin sinxronlashtirilgan.[28] Neyronlar mahalliy darajada bog'langan bo'lib, ular kichik guruhlarni hosil qiladi asabiy ansambllar. Ba'zi bir tarmoq tuzilmalari ma'lum chastotalarda tebranish faolligini rag'batlantiradi. Masalan, o'zaro bog'liq bo'lgan ikkita populyatsiya tomonidan hosil qilingan neyronal faollik inhibitiv va hayajonli hujayralar tomonidan tavsiflangan o'z-o'zidan tebranishlarni ko'rsatishi mumkin Uilson-Kovan modeli.
Agar bir guruh neyronlar sinxronlashtirilgan tebranuvchi faollik bilan shug'ullanadigan bo'lsa, asab ansambli matematik ravishda bitta osilator sifatida ifodalanishi mumkin.[18] Turli xil nerv ansambllari uzoq masofali ulanishlar orqali bog'lanib, keyingi fazoviy miqyosda kuchsiz bog'langan osilatorlar tarmog'ini hosil qiladi. Zaif bog'langan osilatorlar bir qator dinamikani yaratishi mumkin, shu jumladan osilator faolligi.[29] Kabi turli xil miya tuzilmalari orasidagi uzoq muddatli aloqalar talamus va korteks (qarang talamokortikal tebranish ), cheklanganligi sababli vaqtni kechiktirishni o'z ichiga oladi o'tkazuvchanlik tezligi aksonlarning. Ko'pgina ulanishlar o'zaro bog'liq bo'lganligi sababli ular hosil bo'ladi orqaga qaytish ko'chadan tebranish faoliyatini qo'llab-quvvatlovchi. Ko'p kortikal sohalardan yozilgan tebranishlar hosil bo'lish uchun sinxronlashtirilishi mumkin keng miqyosli miya tarmoqlari yordamida dinamikasi va funktsional ulanishi o'rganilishi mumkin spektral tahlil va Grangerning sababi chora-tadbirlar.[30] Miya miqyosidagi keng ko'lamli faoliyat tarqatilgan ma'lumotni birlashtirish uchun zarur bo'lgan miya sohalari o'rtasida dinamik aloqalarni shakllantirishi mumkin.[12]
Neyromodulyatsiya
Tez to'g'ridan-to'g'ri qo'shimcha ravishda sinaptik o'zaro ta'sirlar tarmoq hosil qiluvchi neyronlar o'rtasida tebranuvchi faollik tartibga solinadi neyromodulyatorlar ancha sekin vaqt shkalasida. Ya'ni, ma'lum bir nörotransmitterlarning kontsentratsiyasi darajasi salınımlı faoliyat miqdorini tartibga solishi ma'lum. Masalan; misol uchun, GABA kontsentratsiyani induktsiya qilingan stimullarda tebranish chastotasi bilan ijobiy bog'liqligi ko'rsatilgan.[31] Bir qator yadrolar ichida miya sopi kabi nörotransmitterlarning kontsentratsiya darajalariga ta'sir qiluvchi miya bo'ylab tarqalgan proektsiyalar mavjud noradrenalin, atsetilxolin va serotonin. Ushbu neyrotransmitter tizimlari fiziologik holatga ta'sir qiladi, masalan. hushyorlik yoki qo'zg'alish va alfa faolligi kabi turli xil miya to'lqinlarining amplitudasiga aniq ta'sir ko'rsatadi.[32]
Matematik tavsif
Tebranishlarni ko'pincha matematikadan foydalanib tasvirlash va tahlil qilish mumkin. Matematiklar bir nechtasini aniqladilar dinamik ritmiklikni keltirib chiqaradigan mexanizmlar. Eng muhimlari orasida harmonik (chiziqli) osilatorlar, chegara davri osilatorlar va kechiktirilganmulohaza osilatorlar.[33] Garmonik tebranishlar tabiatda juda tez-tez uchraydi - masalan, tovush to'lqinlari, a harakati mayatnik va har qanday tebranishlar. Ular, odatda, jismoniy tizim a dan ozgina darajada buzilganda paydo bo'ladi minimal energiya holati va matematik jihatdan yaxshi tushuniladi. Vahima qo'zg'atadigan harmonik osilatorlar uyg'ongan EEGdagi alfa ritmini, shuningdek EEG uyqusidagi sekin to'lqinlarni va shpindellarni real ravishda simulyatsiya qiladi. Omadli EEG tahlili algoritmlar ana shunday modellarga asoslangan edi. Boshqa bir qator EEG komponentlari chegara davri yoki kechiktirilgan teskari tebranishlar bilan yaxshiroq tavsiflanadi. Limit-tsikl tebranishlari katta og'ishlarni ko'rsatadigan jismoniy tizimlardan kelib chiqadi muvozanat, kechiktirilgan teskari tebranishlar tizimning tarkibiy qismlari bir-birlariga vaqtni sezilarli darajada kechiktirgandan keyin ta'sir qilganda paydo bo'ladi. Chegara tsikli tebranishlari murakkab bo'lishi mumkin, ammo ularni tahlil qilish uchun kuchli matematik vositalar mavjud; kechiktirilgan teskari tebranishlar matematikasi taqqoslaganda ibtidoiy. Lineer osilatorlar va chegara tsikli osilatorlari kirish tebranishiga qanday javob berishlari jihatidan sifat jihatidan farq qiladi. Lineer osilatorda chastota ozmi-ko'pi doimiy, ammo amplitudasi juda katta farq qilishi mumkin. Limit tsiklli osilatorda amplituda ozmi-ko'pi doimiy bo'lishga intiladi, lekin chastota juda katta farq qilishi mumkin. A yurak urishi chegara tsikli tebranishining misoli, bu urishlar chastotasi keng o'zgarib turadi, shu bilan birga har bir individual urish bir xil miqdordagi qonni pompalamoqda.
Hisoblash modellari miya faoliyatida kuzatiladigan murakkab tebranish dinamikasini tavsiflash uchun turli xil abstraktsiyalarni qabul qilish. Ushbu sohada ko'plab modellar qo'llaniladi, ularning har biri abstraktsiyaning turli darajasida aniqlanadi va asab tizimlarining turli jihatlarini modellashtirishga harakat qiladi. Ular individual neyronlarning qisqa muddatli xatti-harakatlari modellaridan tortib, dinamikasi qanday bo'lgan modellarga qadar asab tizimlari individual neyronlarning o'zaro ta'siridan, xulq-atvorning to'liq quyi tizimlarni ifodalovchi mavhum nerv modullaridan qanday kelib chiqishi mumkinligi haqidagi modellardan kelib chiqadi.
Yagona neyron modeli

Biologik neyron modeli - bu asab hujayralari yoki neyronlarning xususiyatlarini matematik tavsiflash, bu uning biologik jarayonlarini aniq tasvirlash va bashorat qilish uchun mo'ljallangan. Eng muvaffaqiyatli va keng qo'llaniladigan neyronlarning modeli Hodkin-Xaksli modeli, ma'lumotlariga asoslangan kalmar ulkan akson. Bu neyronning elektr xususiyatlariga, xususan, hosil bo'lishi va tarqalishiga yaqinlashadigan chiziqli bo'lmagan oddiy differentsial tenglamalar to'plamidir. harakat potentsiali. Model juda aniq va batafsil va Xodkin va Xaksli ushbu ishi uchun fiziologiya yoki tibbiyot sohasida 1963 yil Nobel mukofotini oldi.
Xojkin-Xaksli modelining matematikasi juda murakkab va shunga o'xshash bir qancha soddalashtirish taklif qilingan FitzHugh-Nagumo modeli, Hindmarsh - Rose modeli yoki kondansatkichni almashtirish modeli[34] integratsiya va olov modelining kengaytmasi sifatida. Bunday modellar faqat asosiy neyron dinamikasini aks ettiradi, masalan, ritmik bosish va yorilish, lekin hisoblash samaradorligi yuqori. Bu a hosil qiluvchi ko'p sonli o'zaro bog'liq neyronlarni simulyatsiya qilishga imkon beradi neyron tarmoq.
Spiking modeli
Nerv tarmog'ining modeli fizik jihatdan bir-biriga bog'langan neyronlarning populyatsiyasini yoki kirishlari yoki signalizatsiya maqsadlari taniqli sxemani aniqlaydigan turli xil neyronlarning guruhini tavsiflaydi. Ushbu modellar neyron sxemalarining dinamikasi individual neyronlarning o'zaro ta'siridan qanday kelib chiqishini tasvirlashga qaratilgan. Neyronlarning mahalliy o'zaro ta'siri spiking faolligini sinxronlashiga olib kelishi va tebranish faoliyatining asosini tashkil qilishi mumkin. Xususan, o'zaro ta'sir qilish modellari piramidal hujayralar va inhibitor internironlar kabi miya ritmini yaratishi ko'rsatilgan gamma faolligi.[35] Xuddi shu tarzda, neyronlarning ta'sir etishmovchiligining fenomenologik modeli bilan neyron tarmoqlarini simulyatsiya qilish spontan keng polosali asab salınımlarını taxmin qilishi mumkinligi ko'rsatildi.[36]
Nerv massasi modeli

Neyron maydon modellari asab tebranishlarini o'rganishda yana bir muhim vosita bo'lib, makon va vaqtdagi o'rtacha otish tezligi kabi o'zgaruvchilar evolyutsiyasini tavsiflovchi matematik asosdir. Ko'p sonli neyronlarning faoliyatini modellashtirishda markaziy g'oya neyronlarning zichligini doimiylik chegarasiga etkazish, natijada fazoviy uzluksizlikka olib keladi. asab tarmoqlari. Ayrim neyronlarni modellashtirish o'rniga, ushbu yondashuv o'rtacha xossalari va o'zaro ta'sirlari bo'yicha neyronlar guruhiga yaqinlashadi. Bunga asoslanadi maydonga yaqinlashishni anglatadi, maydoni statistik fizika bu keng ko'lamli tizimlar bilan shug'ullanadi. Ushbu printsiplarga asoslangan modellar asab tebranishlari va EEG ritmlarining matematik tavsiflarini berish uchun ishlatilgan. Masalan, ular vizual gallyutsinatsiyalarni tekshirish uchun ishlatilgan.[38]
Kuramoto modeli

The Kuramoto modeli birlashtirilgan fazali osilatorlar[39] asab tebranishlari va sinxronizatsiyasini o'rganish uchun ishlatiladigan eng mavhum va asosiy modellardan biridir. U lokal tizim faoliyatini (masalan, bitta neyron yoki neyron ansambli) o'z doirasi orqali ushlaydi bosqich yolg'iz va shuning uchun tebranishlar amplitudasini e'tiborsiz qoldiradi (amplituda doimiy).[40] Ushbu osilatorlarning o'zaro ta'siri oddiy algebraik shakl (masalan, a sinus funktsiyasi) va birgalikda global miqyosda dinamik naqsh hosil qiladi. Kuramoto modeli osilatorli miya faoliyatini o'rganish uchun keng qo'llaniladi va uning neyrobiologik asosliligini oshiradigan bir qancha kengaytmalar taklif qilingan, masalan, mahalliy kortikal bog'lanishning topologik xususiyatlarini o'z ichiga olgan.[41] Xususan, o'zaro ta'sir qiluvchi neyronlar guruhining faoliyati qanday qilib sinxronlashishi va katta hajmdagi tebranishlarni hosil qilishi mumkinligini tasvirlaydi. Haqiqiy uzoq masofali kortikal ulanish va vaqtni kechiktiradigan o'zaro ta'sirlar bilan Kuramoto modelini ishlatadigan simulyatsiyalar tinch holatni takrorlaydigan sekin naqshli dalgalanmalar paydo bo'lishini ko'rsatadi QALIN yordamida o'lchanadigan funktsional xaritalar FMRI.[42]
Faoliyat shakllari
Ham bitta neyron, ham neyron guruhlari o'z-o'zidan tebranish faolligini hosil qilishi mumkin. Bundan tashqari, ular sezgir kirish yoki vosita chiqishiga tebranuvchi javoblarni ko'rsatishi mumkin. Ba'zi bir neyron turlari sinaptik kirish mavjud bo'lmaganda ritmik ravishda yonadi. Xuddi shu tarzda, miya bo'ylab faoliyat tebranish faolligini ochib beradi, sub'ektlar esa hech qanday faoliyat bilan shug'ullanmaydilar dam olish holati faoliyati. Ushbu davom etayotgan ritmlar sezgir kirish yoki vosita chiqishiga javoban har xil tarzda o'zgarishi mumkin. Tebranuvchi faollik chastota va amplituda o'sish yoki pasayish bilan javob berishi yoki vaqtni to'xtatishni ko'rsatishi mumkin, bu fazani qayta tiklash deb ataladi. Bundan tashqari, tashqi faoliyat doimiy faoliyat bilan umuman o'zaro aloqada bo'lmasligi mumkin, natijada qo'shimcha ta'sir ko'rsatadi.
- Tebranuvchi javoblar

The chastota t1 va t2 oralig'ida davom etayotgan tebranish faolligi oshadi.

The amplituda t1 va t2 orasida davom etayotgan tebranish faolligi oshadi.

The bosqich davom etayotgan tebranuvchi faollik t1 darajasida tiklanadi.

Faoliyat t1 va t2 oralig'ida davom etayotgan tebranish faolligiga chiziqli ravishda qo'shiladi.




Doimiy faoliyat
O'z-o'zidan paydo bo'ladigan faoliyat miya sezgir kirish yoki dvigatel chiqishi kabi aniq vazifa bo'lmagan taqdirda faoliyat va shu sababli dam olish holati faoliyati deb ham ataladi. Bu qo'zg'atilgan faoliyatga, ya'ni hissiy ogohlantirishlar yoki vosita reaktsiyalari bilan yuzaga keladigan miya faoliyatiga qarshi. Atama doimiy miya faoliyati ichida ishlatiladi elektroensefalografiya va magnetoensefalografiya a ishlov berish bilan bog'liq bo'lmagan signal komponentlari uchun rag'batlantirish yoki tana qismining harakatlanishi kabi aniq boshqa hodisalarning paydo bo'lishi, ya'ni shakllanmaydigan hodisalar uyg'ongan potentsial /uyg'otilgan dalalar yoki majburiy faoliyat. O'z-o'zidan paydo bo'ladigan faoliyat odatda qabul qilinadi shovqin agar kimdir rag'batlantirishni qayta ishlashga qiziqsa; ammo, o'z-o'zidan paydo bo'ladigan faollik miya rivojlanishi paytida, masalan, tarmoq shakllanishi va sinaptogenezda hal qiluvchi rol o'ynaydi. O'z-o'zidan paydo bo'ladigan faoliyat odamning hozirgi ruhiy holati to'g'risida ma'lumotga ega bo'lishi mumkin (masalan, hushyorlik, hushyorlik) va uxlashda tez-tez ishlatiladi. Kabi ba'zi bir salınımcı faoliyat turlari alfa to'lqinlari, o'z-o'zidan paydo bo'ladigan faoliyatning bir qismidir. Alfa faolligining quvvat o'zgarishini statistik tahlil qilish bimodal taqsimotni, ya'ni yuqori va past amplituda rejimni aniqlaydi va demak, dam olish holati nafaqat shovqin jarayon.[43] FMRI bo'lsa, o'z-o'zidan tebranishlar qon-kislorod darajasiga bog'liq (BOLD) signali, masalan, qolgan davlatlar tarmoqlari bilan bog'langan korrelyatsion naqshlarni ochib beradi standart tarmoq.[44] Dam olish holatidagi davlat tarmoqlarining vaqtinchalik evolyutsiyasi turli chastota diapazonlarida salınımlı EEG faolligining tebranishlari bilan bog'liq.[45]
Davomiy miya faoliyati idrok qilishda ham muhim rol o'ynashi mumkin, chunki u kiruvchi stimullar bilan bog'liq faoliyat bilan o'zaro ta'sir qilishi mumkin. Haqiqatdan ham, EEG tadqiqotlar shuni ko'rsatadiki, vizual idrok kortikal tebranishlarning fazasiga va amplitudasiga bog'liq. Masalan, vizual stimulyatsiya lahzasidagi alfa faolligining amplitudasi va fazasi, sub'ekt tomonidan kuchsiz stimul qabul qilinishini taxmin qiladi.[46][47][48]
Chastotaga javob
Kirishga javoban neyron yoki neyronlar ansambli tebranish chastotasini o'zgartirishi va shu bilan o'zgarishi mumkin stavka u ko'tariladi. Ko'pincha, neyronning otish tezligi uning qabul qilingan faoliyatiga bog'liq. Chastotani o'zgartirish, odatda, markaziy naqsh generatorlarida kuzatiladi va to'g'ridan-to'g'ri yurishdagi qadam chastotasi kabi vosita harakatlarining tezligiga bog'liq. Biroq, o'zgarishlar nisbiy turli miya sohalari orasidagi tebranish chastotasi unchalik keng tarqalgan emas, chunki tebranish faolligining chastotasi ko'pincha miya sohalari orasidagi kechikishlar bilan bog'liq.
Amplituda javob
Uyg'otilgan faoliyatning yonida, stimulni qayta ishlash bilan bog'liq bo'lgan asabiy faoliyat induktsiyaga olib kelishi mumkin. Induktsiya qilingan faollik stimullarni qayta ishlash yoki harakatga tayyorlash orqali kelib chiqadigan doimiy miya faoliyatidagi modulyatsiyani anglatadi. Demak, ular uyg'otilgan javoblardan farqli ravishda bilvosita javobni aks ettiradi. Induksion faoliyatning yaxshi o'rganilgan turi bu tebranish faolligining amplituda o'zgarishi. Masalan; misol uchun, gamma faolligi ko'pincha ob'ektni namoyish qilish paytida bo'lgani kabi, kuchaygan aqliy faoliyat paytida ortadi.[49] Induktsiya qilingan javoblar o'lchovlar bo'yicha har xil bosqichlarga ega bo'lishi va shuning uchun o'rtacha hisoblash paytida bekor qilinishi mumkinligi sababli, ularni faqat quyidagi usulda olish mumkin. vaqt chastotasini tahlil qilish. Induktsiya qilingan faoliyat odatda ko'plab neyronlarning faolligini aks ettiradi: tebranish faolligidagi amplituda o'zgarishlarni asab faoliyati sinxronlashidan kelib chiqadi, masalan, boshoq vaqtini sinxronlash yoki individual neyronlarning membrana potentsial tebranishlari. Shuning uchun tebranish faolligining oshishi ko'pincha hodisalar bilan bog'liq sinxronizatsiya, pasayishlar esa hodisalar bilan bog'liq desinxronizatsiya deb ataladi.[50]
Bosqichni tiklash
Fazni qayta tiklash neyron yoki neyronal ansamblga kirish davom etayotgan tebranishlar fazasini tiklaganda sodir bo'ladi.[51] Spike vaqti neyronlarning kiritilishiga moslashtirilgan bitta neyronlarda juda keng tarqalgan (neyron davriy kirishga javoban belgilangan kechikish bilan bosilib chiqishi mumkin, bu fazani qulflash deb ataladi)[13]) va shuningdek, neyronlarning fazalari bir vaqtning o'zida sozlanganda neyronal ansambllarda paydo bo'lishi mumkin. Bosqichni tiklash turli neyronlarni yoki turli miya mintaqalarini sinxronlashtirish uchun juda muhimdir[12][29] chunki boshoqlash vaqti boshqa neyronlarning faolligi bilan bog'liq bo'lishi mumkin.
Bosqichni tiklash, shuningdek, ishlatilgan atamani o'rganishga imkon beradi elektroensefalografiya va magnetoensefalografiya to'g'ridan-to'g'ri bog'liq bo'lgan miya faoliyatidagi javoblar uchun rag'batlantirish - tegishli faoliyat. Uyg'otilgan potentsial va voqea bilan bog'liq potentsial elektroansefalogrammadan stimul bilan blokirovka qilingan o'rtacha, ya'ni rag'batlantiruvchi taqdimot atrofida sobit kechikishlarda har xil sinovlarni o'rtacha hisoblash yo'li bilan olinadi. Natijada, har bir o'lchovda bir xil bo'lgan signal komponentlari saqlanib qoladi va qolganlari, ya'ni doimiy yoki o'z-o'zidan paydo bo'ladigan faoliyat o'rtacha hisoblanadi. Ya'ni, voqea bilan bog'liq potentsial faqat miya faoliyatidagi tebranishlarni aks ettiradi bosqich - rag'batlantirish yoki voqea uchun qulflangan. Uyg'otilgan faoliyat ko'pincha miyaning doimiy faoliyatidan mustaqil deb hisoblanadi, garchi bu doimiy munozara bo'lsa ham.[52][53]
Asimmetrik amplituda modulyatsiya
Yaqinda taklif qilinganidek, bosqichlar sinovlar davomida mos kelmasa ham, induktsiya qilingan faoliyat sabab bo'lishi mumkin voqea bilan bog'liq potentsial chunki davom etayotgan miya tebranishlari nosimmetrik bo'lmasligi mumkin va shuning uchun amplituda modulyatsiyalar o'rtacha siljishga olib kelmaydigan boshlang'ich siljishiga olib kelishi mumkin.[54][55] Ushbu model, voqea bilan bog'liq bo'lgan sekin reaktsiyalar, masalan, assimetrik alfa faolligi, dendritlardan oldinga va orqaga qarab tarqaladigan hujayra ichidagi oqimlarning assimetriyasi kabi miyaning assimetrik tebranish amplituda modulyatsiyasi natijasida kelib chiqishi mumkinligini anglatadi.[56] Ushbu taxminga ko'ra, dendritik oqimdagi nosimmetrikliklar EEG va MEG bilan o'lchanadigan tebranish faolligidagi nosimmetrikliklar keltirib chiqarishi mumkin, chunki piramidal hujayralardagi dendritik oqimlar odatda bosh terisida o'lchanadigan EEG va MEG signallarini hosil qiladi deb o'ylashadi.[57]
Funktsiya
Nerv sinxronizatsiyasi vazifa cheklovlari bilan modulyatsiya qilinishi mumkin, masalan diqqat, va rol o'ynaydi deb o'ylashadi xususiyati majburiy,[58] neyronal aloqa,[5] va motorni muvofiqlashtirish.[7] Neyronlarning tebranishlari eng dolzarb mavzuga aylandi nevrologiya 1990-yillarda Grey, Singer va boshqalar tomonidan miyaning vizual tizimini o'rganish tadqiqotlari qo'llab-quvvatlanganda paydo bo'ldi asab bilan bog'lanish gipoteza.[59] Ushbu g'oyaga ko'ra, neyronal ansambllarda sinxron tebranishlar ob'ektning turli xil xususiyatlarini ifodalovchi neyronlarni bog'laydi. Masalan, odam daraxtga qaraganida, daraxt tanasini ifodalovchi ingl. Korteks neyronlari va shu daraxt shoxlarini ifodalovchi sinxronlashganda daraxtning yagona vakili hosil bo'ladi. Ushbu hodisa eng yaxshi ko'rinishda mahalliy dala salohiyati mahalliy neyron guruhlarining sinxron faolligini aks ettiradigan, ammo u ham ko'rsatilgan EEG va MEG sinxron tebranuvchi faollik va idrok guruhlash kabi turli xil bilim funktsiyalari o'rtasidagi yaqin aloqalar uchun tobora ortib borayotgan dalillarni taqdim etadigan yozuvlar.[58]
Yurak stimulyatori
Hujayralar sinoatrial tugun, joylashgan o'ng atrium o'z-o'zidan yurakning depolyarizatsiya qilish daqiqada taxminan 100 marta. Yurakning barcha hujayralari yurak qisqarishini qo'zg'atadigan harakat potentsialini yaratish qobiliyatiga ega bo'lsa-da, sinoatrial tugun uni boshqa zonalarga nisbatan bir oz tezroq hosil qilganligi sababli, uni boshlaydi. Demak, bu hujayralar normal hosil qiladi sinus ritmi va yurak stimulyatori hujayralari deb ataladi, chunki ular to'g'ridan-to'g'ri boshqaradilar yurak urish tezligi. Tashqi asab va gormonal nazorat bo'lmasa, SA tugunidagi hujayralar ritmik ravishda bo'shatiladi. Sinoatrial tugun juda ko'p innervatsiya qilingan avtonom asab tizimi, yurak stimulyatori hujayralarining o'z-o'zidan otish chastotasini yuqoriga yoki pastga tartibga soladi.
Markaziy naqsh generatori
Neyronlarning sinxron otilishi ham ritmik harakatlar uchun davriy motor buyruqlarining asosini tashkil etadi. Ushbu ritmik natijalar tarmoqni tashkil etuvchi o'zaro ta'sir qiluvchi neyronlar guruhi tomonidan ishlab chiqariladi markaziy naqsh generatori. Markaziy naqsh ishlab chiqaruvchilari - bu faollashtirilganida, aniq vaqt ma'lumotlarini olib boruvchi sezgir yoki pasayuvchi yozuvlar bo'lmaganda ritmik vosita naqshlarini ishlab chiqaradigan neyronlarning sxemalari. Examples are yurish, nafas olish va suzish,[60] Most evidence for central pattern generators comes from lower animals, such as the lamprey, but there is also evidence for spinal central pattern generators in humans.[61][62]
Information processing
Neuronal spiking is generally considered the basis for information transfer in the brain. For such a transfer, information needs to be coded in a spiking pattern. Different types of coding schemes have been proposed, such as rate coding va temporal coding. Neural oscillations could create periodic time windows in which input spikes have larger effect on neurons, thereby providing a mechanism for decoding temporal codes.[63]
Idrok
Synchronization of neuronal firing may serve as a means to group spatially segregated neurons that respond to the same stimulus in order to bind these responses for further joint processing, i.e. to exploit temporal synchrony to encode relations. Purely theoretical formulations of the binding-by-synchrony hypothesis were proposed first,[64] but subsequently extensive experimental evidence has been reported supporting the potential role of synchrony as a relational code.[65]
The functional role of synchronized oscillatory activity in the brain was mainly established in experiments performed on awake kittens with multiple electrodes implanted in the visual cortex. These experiments showed that groups of spatially segregated neurons engage in synchronous oscillatory activity when activated by visual stimuli. The frequency of these oscillations was in the range of 40 Hz and differed from the periodic activation induced by the grating, suggesting that the oscillations and their synchronization were due to internal neuronal interactions.[65] Similar findings were shown in parallel by the group of Eckhorn, providing further evidence for the functional role of neural synchronization in feature binding.[66] Since then, numerous studies have replicated these findings and extended them to different modalities such as EEG, providing extensive evidence of the functional role of gamma oscillations in visual perception.
Gilles Laurent and colleagues showed that oscillatory synchronization has an important functional role in odor perception. Perceiving different odors leads to different subsets of neurons firing on different sets of oscillatory cycles.[67] These oscillations can be disrupted by GABA blocker pikrotoksin,[68] and the disruption of the oscillatory synchronization leads to impairment of behavioral discrimination of chemically similar odorants in bees[69] and to more similar responses across odors in downstream β-lobe neurons.[70] Recent follow-up of this work has shown that oscillations create periodic integration windows for Kenyon cells in the insect mushroom body, such that incoming spikes from the antennal lobe are more effective in activating Kenyon cells only at specific phases of the oscillatory cycle.[63]
Neural oscillations are also thought be involved in the sense of time[71] and in somatosensory perception.[72] However, recent findings argue against a clock-like function of cortical gamma oscillations.[73]
Motor coordination
Oscillations have been commonly reported in the motor system. Pfurtscheller and colleagues found a reduction in alfa (8–12 Hz) and beta-versiya (13–30 Hz) oscillations in EEG activity when subjects made a movement.[50][74] Using intra-cortical recordings, similar changes in oscillatory activity were found in the motor cortex when the monkeys performed motor acts that required significant attention.[75][76] In addition, oscillations at spinal level become synchronised to beta oscillations in the motor cortex during constant muscle activation, as determined by cortico-muscular coherence.[77][78][79] Likewise, muscle activity of different muscles reveals inter-muscular coherence at multiple distinct frequencies reflecting the underlying neural circuitry da ishtirok etish motor coordination.[80][81]
Recently it was found that cortical oscillations propagate as travelling waves across the surface of the motor cortex along dominant spatial axes characteristic of the local circuitry of the motor cortex.[82] It has been proposed that motor commands in the form of travelling waves can be spatially filtered by the descending fibres to selectively control muscle force.[83] Simulations have shown that ongoing wave activity in cortex can elicit steady muscle force with physiological levels of EEG-EMG coherence.[84]
Oscillatory rhythms at 10 Hz have been recorded in a brain area called the pastki zaytun, which is associated with the cerebellum.[14] These oscillations are also observed in motor output of physiological titroq[85] and when performing slow finger movements.[86] These findings may indicate that the human brain controls continuous movements intermittently. In support, it was shown that these movement discontinuities are directly correlated to oscillatory activity in a cerebello-thalamo-cortical loop, which may represent a neural mechanism for the intermittent motor control.[87]
Xotira
Neural oscillations, in particular theta activity, are extensively linked to memory function. Theta rhythms are very strong in rodent hippocampi and entorhinal cortex during learning and memory retrieval, and they are believed to be vital to the induction of uzoq muddatli kuchaytirish, a potential cellular mechanism for learning and memory. Birlashma between theta and gamma activity is thought to be vital for memory functions, including episodic memory.[88][89] Tight coordination of single-neuron spikes with local theta oscillations is linked to successful memory formation in humans, as more stereotyped spiking predicts better memory.[90]
Sleep and consciousness
Sleep is a naturally recurring state characterized by reduced or absent ong and proceeds in cycles of rapid eye movement (REM) va non-rapid eye movement (NREM) sleep. Sleep stages are characterized by spectral content of EEG: for instance, stage N1 refers to the transition of the brain from alpha waves (common in the awake state) to theta waves, whereas stage N3 (deep or slow-wave sleep) is characterized by the presence of delta waves. The normal order of sleep stages is N1 → N2 → N3 → N2 → REM.[iqtibos kerak ]
Rivojlanish
Neural oscillations may play a role in neural development. Masalan, retinal waves are thought to have properties that define early connectivity of circuits and synapses between cells in the retina.[91]
Patologiya

Specific types of neural oscillations may also appear in pathological situations, such as Parkinson kasalligi yoki epilepsiya. These pathological oscillations often consist of an aberrant version of a normal oscillation. For example, one of the best known types is the spike and wave oscillation, which is typical of generalized or absence epileptic seizures, and which resembles normal sleep spindle oscillations.
Tremor
A tremor is an involuntary, somewhat rhythmic, muscle contraction and relaxation involving to-and-fro movements of one or more body parts. It is the most common of all involuntary movements and can affect the hands, arms, eyes, face, head, vocal cords, trunk, and legs. Aksariyat titroq qo'llarda sodir bo'ladi. In some people, tremor is a symptom of another neurological disorder. Many different forms of tremor have been identified, such as muhim titroq yoki Parkinsonian tremor. It is argued that tremors are likely to be multifactorial in origin, with contributions from neural oscillations in the central nervous systems, but also from peripheral mechanisms such as reflex loop resonances.[92]
Epilepsiya
Epilepsy is a common chronic neurological disorder characterized by soqchilik. These seizures are transient signs and/or symptoms of abnormal, excessive or hypersynchronous neuronal activity miyada.[93]
Thalamocortical dysrhythmia
In thalamocortical dysrhythmia (TCD), normal thalamocortical resonance is disrupted. The thalamic loss of input allows the frequency of the thalamo-cortical column to slow into the theta or delta band as identified by MEG and EEG by machine learning.[94] TCD can be treated with neurosurgical methods like thalamotomy.
Ilovalar
Clinical endpoints
Neural oscillations are sensitive to several drugs influencing brain activity; accordingly, biomarkerlar based on neural oscillations are emerging as secondary endpoints in clinical trials and in quantifying effects in pre-clinical studies. These biomarkers are often named "EEG biomarkers" or "Neurophysiological Biomarkers" and are quantified using Quantitative electroencephalography (qEEG). EEG biomarkers can be extracted from the EEG using the open-source Neurophysiological Biomarker Toolbox.
Miya-kompyuter interfeysi
Neural oscillation has been applied as a control signal in various brain–computer interfaces (BCIs).[95] For example, a non-invasive BCI can be created by placing electrodes on the scalp and then measuring the weak electric signals. Although individual neuron activities cannot be recorded through non-invasive BCI because the skull damps and blurs the electromagnetic signals, oscillatory activity can still be reliably detected. The BCI was introduced by Vidal in 1973[96] as challenge of using EEG signals to control objects outside human body.
After the BCI challenge, in 1988, alpha rhythm was used in a brain rhythm based BCI for control of a physical object, a robot.[97][98] Alpha rhythm based BCI was the first BCI for control of a robot.[99][100]In particular, some forms of BCI allow users to control a device by measuring the amplitude of oscillatory activity in specific frequency bands, including mu va beta-versiya ritmlar.
Misollar
A non-inclusive list of types of oscillatory activity found in the central nervous system:
- Delta wave
- Theta wave
- Alpha wave
- Mu wave
- Beta wave
- Gamma wave
- PGO waves
- Sleep spindle
- Thalamocortical oscillations
- Subthreshold membrane potential oscillations
- Bursting
- Cardiac cycle
- Epileptik tutilish
- Mathematical modeling of electrophysiological activity in epilepsy
- Sharp wave–ripple complexes
Shuningdek qarang
- Kibernetika
- Dinamik tizimlar nazariyasi
- Neurocybernetics
- Systems neuroscience
- EEG analysis
- Oscillatory neural network
- ThetaHealing
Adabiyotlar
- ^ Llinás RR (2014). "Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective". Frontiers in Cellular Neuroscience. 8: 320. doi:10.3389/fncel.2014.00320. PMC 4219458. PMID 25408634.
- ^ "Caton, Richard - The electric currents of the brain". echo.mpiwg-berlin.mpg.de. Olingan 2018-12-21.
- ^ Coenen A, Fine E, Zayachkivska O (2014). "Adolf Beck: a forgotten pioneer in electroencephalography". Journal of the History of the Neurosciences. 23 (3): 276–86. doi:10.1080/0964704x.2013.867600. PMID 24735457. S2CID 205664545.
- ^ Pravdich-Neminsky VV (1913). "Ein Versuch der Registrierung der elektrischen Gehirnerscheinungen". Zentralblatt für Physiologie. 27: 951–60.
- ^ a b Fries P (October 2005). "A mechanism for cognitive dynamics: neuronal communication through neuronal coherence". Kognitiv fanlarning tendentsiyalari. 9 (10): 474–80. doi:10.1016/j.tics.2005.08.011. PMID 16150631. S2CID 6275292.
- ^ Fell J, Axmacher N (February 2011). "The role of phase synchronization in memory processes". Tabiat sharhlari. Nevrologiya. 12 (2): 105–18. doi:10.1038/nrn2979. PMID 21248789. S2CID 7422401.
- ^ a b Schnitzler A, Gross J (April 2005). "Normal and pathological oscillatory communication in the brain". Tabiat sharhlari. Nevrologiya. 6 (4): 285–96. doi:10.1038/nrn1650. PMID 15803160. S2CID 2749709.
- ^ Foster JJ, Sutterer DW, Serences JT, Vogel EK, Awh E (July 2017). "Alpha-Band Oscillations Enable Spatially and Temporally Resolved Tracking of Covert Spatial Attention". Psixologiya fanlari. 28 (7): 929–941. doi:10.1177/0956797617699167. PMC 5675530. PMID 28537480.
- ^ Berger H, Gray CM (1929). "Uber das Elektroenkephalogramm des Menschen". Arch Psychiat Nervenkr. 87: 527–570. doi:10.1007/BF01797193. S2CID 10835361.
- ^ Dement W, Kleitman N (November 1957). "Cyclic variations in EEG during sleep and their relation to eye movements, body motility, and dreaming". Electroencephalography and Clinical Neurophysiology. 9 (4): 673–90. doi:10.1016/0013-4694(57)90088-3. PMID 13480240.
- ^ Engel AK, Singer W (January 2001). "Temporal binding and the neural correlates of sensory awareness". Kognitiv fanlarning tendentsiyalari. 5 (1): 16–25. doi:10.1016/S1364-6613(00)01568-0. PMID 11164732. S2CID 11922975.
- ^ a b v Varela F, Lachaux JP, Rodriguez E, Martinerie J (April 2001). "The brainweb: phase synchronization and large-scale integration". Tabiat sharhlari. Nevrologiya. 2 (4): 229–39. doi:10.1038/35067550. PMID 11283746. S2CID 18651043.
- ^ a b v Izhikevich EM (2007). Dynamical systems in neuroscience. Cambridge, Massachusetts: The MIT Press.
- ^ a b Llinás R, Yarom Y (July 1986). "Oscillatory properties of guinea-pig inferior olivary neurones and their pharmacological modulation: an in vitro study". Fiziologiya jurnali. 376: 163–82. doi:10.1113/jphysiol.1986.sp016147. PMC 1182792. PMID 3795074.
- ^ Mureşan RC, Jurjuţ OF, Moca VV, Singer W, Nikolić D (March 2008). "The oscillation score: an efficient method for estimating oscillation strength in neuronal activity". Neyrofiziologiya jurnali. 99 (3): 1333–53. doi:10.1152/jn.00772.2007. PMID 18160427.
- ^ Burrow T (1943). "The neurodynamics of behavior. A phylobiological foreword". Ilmiy falsafa. 10 (4): 271–288. doi:10.1086/286819. S2CID 121438105.
- ^ Vansteensel MJ, Pels EG, Bleichner MG, Branco MP, Denison T, Freudenburg ZV, et al. (November 2016). "Fully Implanted Brain-Computer Interface in a Locked-In Patient with ALS". Nyu-England tibbiyot jurnali. 375 (21): 2060–2066. doi:10.1056/NEJMoa1608085. hdl:1874/344360. PMC 5326682. PMID 27959736.
- ^ a b Haken H (1996). Principles of brain functioning. Springer. ISBN 978-3-540-58967-9.
- ^ a b v Wang XJ (July 2010). "Neurophysiological and computational principles of cortical rhythms in cognition". Physiological Reviews. 90 (3): 1195–268. doi:10.1152/physrev.00035.2008. PMC 2923921. PMID 20664082.
- ^ Nunez PL, Srinivasan R (1981). Electric fields of the brain: The neurophysics of EEG. Oksford universiteti matbuoti. ISBN 9780195027969.
- ^ Cardin JA, Carlén M, Meletis K, Knoblich U, Zhang F, Deisseroth K, et al. (Iyun 2009). "Driving fast-spiking cells induces gamma rhythm and controls sensory responses". Tabiat. 459 (7247): 663–7. Bibcode:2009Natur.459..663C. doi:10.1038/nature08002. PMC 3655711. PMID 19396156.
- ^ Llinás R, Ribary U, Contreras D, Pedroarena C (November 1998). "The neuronal basis for consciousness". London Qirollik Jamiyatining falsafiy operatsiyalari. B seriyasi, Biologiya fanlari. 353 (1377): 1841–9. doi:10.1098/rstb.1998.0336. PMC 1692417. PMID 9854256.
- ^ Bollimunta A, Mo J, Schroeder CE, Ding M (March 2011). "Neuronal mechanisms and attentional modulation of corticothalamic α oscillations". Neuroscience jurnali. 31 (13): 4935–43. doi:10.1523/JNEUROSCI.5580-10.2011. PMC 3505610. PMID 21451032.
- ^ Suffczynski P, Kalitzin S, Pfurtscheller G, Lopes da Silva FH (December 2001). "Computational model of thalamo-cortical networks: dynamical control of alpha rhythms in relation to focal attention". International Journal of Psychophysiology. 43 (1): 25–40. doi:10.1016/S0167-8760(01)00177-5. PMID 11742683.
- ^ Cabral J, Luckhoo H, Woolrich M, Joensson M, Mohseni H, Baker A, et al. (2014 yil aprel). "Exploring mechanisms of spontaneous functional connectivity in MEG: how delayed network interactions lead to structured amplitude envelopes of band-pass filtered oscillations". NeuroImage. 90: 423–35. doi:10.1016/j.neuroimage.2013.11.047. PMID 24321555.
- ^ Llinás RR (December 1988). "The intrinsic electrophysiological properties of mammalian neurons: insights into central nervous system function". Ilm-fan. 242 (4886): 1654–64. Bibcode:1988Sci...242.1654L. doi:10.1126/science.3059497. PMID 3059497.
- ^ Llinás RR, Grace AA, Yarom Y (February 1991). "In vitro neurons in mammalian cortical layer 4 exhibit intrinsic oscillatory activity in the 10- to 50-Hz frequency range". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 88 (3): 897–901. Bibcode:1991PNAS...88..897L. doi:10.1073/pnas.88.3.897. PMC 50921. PMID 1992481.
- ^ Zeitler M, Daffertshofer A, Gielen CC (June 2009). "Asymmetry in pulse-coupled oscillators with delay" (PDF). Jismoniy sharh E. 79 (6 Pt 2): 065203. Bibcode:2009PhRvE..79f5203Z. doi:10.1103/PhysRevE.79.065203. hdl:1871/29169. PMID 19658549.
- ^ a b Pikovsky A, Rosenblum M, Kurths J (2001). Synchronization: a universal concept in nonlinear sciences. Kembrij universiteti matbuoti. ISBN 978-0-521-53352-2.
- ^ Andrea Brovelli, Steven L. Bressler and their colleagues, 2004
- ^ Muthukumaraswamy SD, Edden RA, Jones DK, Swettenham JB, Singh KD (May 2009). "Resting GABA concentration predicts peak gamma frequency and fMRI amplitude in response to visual stimulation in humans". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 106 (20): 8356–61. Bibcode:2009PNAS..106.8356M. doi:10.1073/pnas.0900728106. PMC 2688873. PMID 19416820.
- ^ Moruzzi G, Magoun HW (November 1949). "Brain stem reticular formation and activation of the EEG". Electroencephalography and Clinical Neurophysiology. 1 (4): 455–73. doi:10.1016/0013-4694(49)90219-9. PMID 18421835.
- ^ Buzsáki G, Draguhn A (June 2004). "Neuronal oscillations in cortical networks". Ilm-fan. 304 (5679): 1926–9. Bibcode:2004Sci...304.1926B. doi:10.1126/science.1099745. PMID 15218136. S2CID 8002293.
- ^ Cejnar P, Vyšata O, Kukal J, Beránek M, Vališ M, Procházka A (April 2020). "Simple capacitor-switch model of excitatory and inhibitory neuron with all parts biologically explained allows input fire pattern dependent chaotic oscillations". Ilmiy ma'ruzalar. 10 (1): 7353. Bibcode:2020NatSR..10.7353C. doi:10.1038/s41598-020-63834-7. PMC 7192907. PMID 32355185.
- ^ Whittington MA, Traub RD, Kopell N, Ermentrout B, Buhl EH (December 2000). "Inhibition-based rhythms: experimental and mathematical observations on network dynamics". International Journal of Psychophysiology. 38 (3): 315–36. CiteSeerX 10.1.1.16.6410. doi:10.1016/S0167-8760(00)00173-2. PMID 11102670.
- ^ Goldental A, Vardi R, Sardi S, Sabo P, Kanter I (2015). "Broadband macroscopic cortical oscillations emerge from intrinsic neuronal response failures". Frontiers in Neural Circuits. 9: 65. arXiv:1511.00235. doi:10.3389/fncir.2015.00065. PMC 4626558. PMID 26578893.
- ^ Wendling F, Bellanger JJ, Bartolomei F, Chauvel P (October 2000). "Relevance of nonlinear lumped-parameter models in the analysis of depth-EEG epileptic signals". Biologik kibernetika. 83 (4): 367–78. doi:10.1007/s004220000160. PMID 11039701. S2CID 8751526.
- ^ Bressloff PC, Cowan JD (2003) Spontaneous pattern formation in primary visual cortex. In: J Hogan, AR Krauskopf, M di Bernado, RE Wilson (Eds.), Nonlinear dynamics and chaos: where do we go from here?
- ^ Kuramoto Y (1984). Chemical Oscillations, Waves, and Turbulence. Dover Publications.
- ^ Ermentrout B (1994). "An introduction to neural oscillators". In F Ventriglia (ed.). Neural Modeling and Neural Networks. pp. 79–110.
- ^ Breakspear M, Heitmann S, Daffertshofer A (2010). "Generative models of cortical oscillations: neurobiological implications of the kuramoto model". Frontiers in Human Neuroscience. 4: 190. doi:10.3389/fnhum.2010.00190. PMC 2995481. PMID 21151358.
- ^ Cabral J, Hugues E, Sporns O, Deco G (July 2011). "Role of local network oscillations in resting-state functional connectivity". NeuroImage. 57 (1): 130–139. doi:10.1016/j.neuroimage.2011.04.010. PMID 21511044. S2CID 13959959.
- ^ Freyer F, Aquino K, Robinson PA, Ritter P, Breakspear M (July 2009). "Bistability and non-Gaussian fluctuations in spontaneous cortical activity". Neuroscience jurnali. 29 (26): 8512–24. doi:10.1523/JNEUROSCI.0754-09.2009. PMC 6665653. PMID 19571142.
- ^ Fox MD, Raichle ME (September 2007). "Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging". Tabiat sharhlari. Nevrologiya. 8 (9): 700–11. doi:10.1038/nrn2201. PMID 17704812. S2CID 15979590.
- ^ Laufs H, Krakow K, Sterzer P, Eger E, Beyerle A, Salek-Haddadi A, Kleinschmidt A (September 2003). "Electroencephalographic signatures of attentional and cognitive default modes in spontaneous brain activity fluctuations at rest". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 100 (19): 11053–8. Bibcode:2003PNAS..10011053L. doi:10.1073/pnas.1831638100. PMC 196925. PMID 12958209.
- ^ Mathewson KE, Gratton G, Fabiani M, Beck DM, Ro T (March 2009). "To see or not to see: prestimulus alpha phase predicts visual awareness". Neuroscience jurnali. 29 (9): 2725–32. doi:10.1523/JNEUROSCI.3963-08.2009. PMC 2724892. PMID 19261866.
- ^ Busch NA, Dubois J, VanRullen R (June 2009). "The phase of ongoing EEG oscillations predicts visual perception". Neuroscience jurnali. 29 (24): 7869–76. doi:10.1523/jneurosci.0113-09.2009. PMC 6665641. PMID 19535598.
- ^ van Dijk H, Schoffelen JM, Oostenveld R, Jensen O (February 2008). "Prestimulus oscillatory activity in the alpha band predicts visual discrimination ability". Neuroscience jurnali. 28 (8): 1816–23. doi:10.1523/jneurosci.1853-07.2008. PMC 6671447. PMID 18287498.
- ^ Tallon-Baudry C, Bertrand O (April 1999). "Oscillatory gamma activity in humans and its role in object representation". Kognitiv fanlarning tendentsiyalari. 3 (4): 151–162. doi:10.1016/S1364-6613(99)01299-1. PMID 10322469. S2CID 1308261.
- ^ a b Pfurtscheller G, Lopes da Silva FH (November 1999). "Event-related EEG/MEG synchronization and desynchronization: basic principles". Clinical Neurophysiology. 110 (11): 1842–57. doi:10.1016/S1388-2457(99)00141-8. PMID 10576479. S2CID 24756702.
- ^ Tass PA (2007). Phase resetting in medicine and biology: stochastic modelling and data analysis. Berlin Geydelberg: Springer-Verlag. ISBN 978-3-540-65697-5.
- ^ Makeig S, Westerfield M, Jung TP, Enghoff S, Townsend J, Courchesne E, Sejnowski TJ (January 2002). "Dynamic brain sources of visual evoked responses". Ilm-fan. 295 (5555): 690–4. Bibcode:2002Sci...295..690M. doi:10.1126/science.1066168. PMID 11809976. S2CID 15200185.
- ^ Mäkinen V, Tiitinen H, May P (February 2005). "Auditory event-related responses are generated independently of ongoing brain activity". NeuroImage. 24 (4): 961–8. doi:10.1016/j.neuroimage.2004.10.020. PMID 15670673. S2CID 16210275.
- ^ Nikulin VV, Linkenkaer-Hansen K, Nolte G, Lemm S, Müller KR, Ilmoniemi RJ, Curio G (May 2007). "A novel mechanism for evoked responses in the human brain". Evropa nevrologiya jurnali. 25 (10): 3146–54. doi:10.1111/j.1460-9568.2007.05553.x. PMID 17561828. S2CID 12113334.
- ^ Mazaheri A, Jensen O (July 2008). "Asymmetric amplitude modulations of brain oscillations generate slow evoked responses". Neuroscience jurnali. 28 (31): 7781–7. doi:10.1523/JNEUROSCI.1631-08.2008. PMC 6670375. PMID 18667610.
- ^ Mazaheri A, Jensen O (2008). "Rhythmic pulsing: linking ongoing brain activity with evoked responses". Frontiers in Human Neuroscience. 4: 177. doi:10.3389/fnhum.2010.00177. PMC 2972683. PMID 21060804.
- ^ Hamalainen M, Hari R, Ilmoniemi RJ, Knuutila J, Lounasmaa OV (1993). "Magnetoencephalography - Theory, instrumentation, and applications to noninvasive studies of the working human brain". Rev Mod Phys. 65 (2): 413–497. Bibcode:1993RvMP...65..413H. doi:10.1103/RevModPhys.65.413.
- ^ a b Singer W (1993). "Synchronization of cortical activity and its putative role in information processing and learning". Annual Review of Physiology. 55: 349–74. doi:10.1146/annurev.ph.55.030193.002025. PMID 8466179.
- ^ Singer W, Gray CM (1995). "Visual feature integration and the temporal correlation hypothesis". Nevrologiyani yillik sharhi. 18: 555–86. CiteSeerX 10.1.1.308.6735. doi:10.1146/annurev.ne.18.030195.003011. PMID 7605074.
- ^ Marder E, Bucher D (November 2001). "Central pattern generators and the control of rhythmic movements". Hozirgi biologiya. 11 (23): R986-96. doi:10.1016/S0960-9822(01)00581-4. PMID 11728329. S2CID 1294374.
- ^ Dimitrijevic MR, Gerasimenko Y, Pinter MM (November 1998). "Evidence for a spinal central pattern generator in humans". Nyu-York Fanlar akademiyasining yilnomalari. 860 (1): 360–76. Bibcode:1998NYASA.860..360D. doi:10.1111/j.1749-6632.1998.tb09062.x. PMID 9928325. S2CID 102514.
- ^ Danner SM, Hofstoetter US, Freundl B, Binder H, Mayr W, Rattay F, Minassian K (March 2015). "Human spinal locomotor control is based on flexibly organized burst generators". Miya. 138 (Pt 3): 577–88. doi:10.1093/brain/awu372. PMC 4408427. PMID 25582580.
- ^ a b Gupta N, Singh SS, Stopfer M (December 2016). "Oscillatory integration windows in neurons". Tabiat aloqalari. 7: 13808. Bibcode:2016NatCo...713808G. doi:10.1038/ncomms13808. PMC 5171764. PMID 27976720.
- ^ Milner PM (November 1974). "A model for visual shape recognition". Psixologik sharh. 81 (6): 521–35. doi:10.1037/h0037149. PMID 4445414.
- ^ a b Gray CM, König P, Engel AK, Singer W (March 1989). "Oscillatory responses in cat visual cortex exhibit inter-columnar synchronization which reflects global stimulus properties". Tabiat. 338 (6213): 334–7. Bibcode:1989Natur.338..334G. doi:10.1038/338334a0. PMID 2922061. S2CID 4281744.
- ^ Eckhorn R, Bauer R, Jordan W, Brosch M, Kruse W, Munk M, Reitboeck HJ (1988). "Coherent oscillations: a mechanism of feature linking in the visual cortex? Multiple electrode and correlation analyses in the cat". Biologik kibernetika. 60 (2): 121–30. doi:10.1007/BF00202899. PMID 3228555. S2CID 206771651.
- ^ Wehr M, Laurent G (November 1996). "Odour encoding by temporal sequences of firing in oscillating neural assemblies". Tabiat. 384 (6605): 162–6. Bibcode:1996Natur.384..162W. doi:10.1038/384162a0. PMID 8906790. S2CID 4286308.
- ^ MacLeod K, Laurent G (November 1996). "Distinct mechanisms for synchronization and temporal patterning of odor-encoding neural assemblies". Ilm-fan. 274 (5289): 976–9. Bibcode:1996Sci...274..976M. doi:10.1126/science.274.5289.976. PMID 8875938. S2CID 10744144.
- ^ Stopfer M, Bhagavan S, Smith BH, Laurent G (November 1997). "Impaired odour discrimination on desynchronization of odour-encoding neural assemblies". Tabiat. 390 (6655): 70–4. Bibcode:1997Natur.390...70S. doi:10.1038/36335. PMID 9363891. S2CID 205024830.
- ^ MacLeod K, Bäcker A, Laurent G (October 1998). "Who reads temporal information contained across synchronized and oscillatory spike trains?". Tabiat. 395 (6703): 693–8. Bibcode:1998Natur.395..693M. doi:10.1038/27201. PMID 9790189. S2CID 4424801.
- ^ Buhusi CV, Meck WH (October 2005). "What makes us tick? Functional and neural mechanisms of interval timing". Tabiat sharhlari. Nevrologiya. 6 (10): 755–65. doi:10.1038/nrn1764. PMID 16163383. S2CID 29616055.
- ^ Ahissar E, Zacksenhouse M (2001). Temporal and spatial coding in the rat vibrissal system. Prog Brain Res. Progress in Brain Research. 130. 75-87 betlar. doi:10.1016/S0079-6123(01)30007-9. ISBN 9780444501103. PMID 11480290.
- ^ Burns SP, Xing D, Shapley RM (June 2011). "Is gamma-band activity in the local field potential of V1 cortex a "clock" or filtered noise?". Neuroscience jurnali. 31 (26): 9658–64. doi:10.1523/jneurosci.0660-11.2011. PMC 3518456. PMID 21715631.
- ^ Pfurtscheller G, Aranibar A (June 1977). "Event-related cortical desynchronization detected by power measurements of scalp EEG". Electroencephalography and Clinical Neurophysiology. 42 (6): 817–26. doi:10.1016/0013-4694(77)90235-8. PMID 67933.
- ^ Murthy VN, Fetz EE (December 1996). "Oscillatory activity in sensorimotor cortex of awake monkeys: synchronization of local field potentials and relation to behavior". Neyrofiziologiya jurnali. 76 (6): 3949–67. doi:10.1152/jn.1996.76.6.3949. PMID 8985892.
- ^ Sanes JN, Donoghue JP (May 1993). "Oscillations in local field potentials of the primate motor cortex during voluntary movement". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 90 (10): 4470–4. Bibcode:1993PNAS...90.4470S. doi:10.1073/pnas.90.10.4470. PMC 46533. PMID 8506287.
- ^ Conway BA, Halliday DM, Farmer SF, Shahani U, Maas P, Weir AI, Rosenberg JR (December 1995). "Synchronization between motor cortex and spinal motoneuronal pool during the performance of a maintained motor task in man". Fiziologiya jurnali. 489 ( Pt 3) (3): 917–24. doi:10.1113/jphysiol.1995.sp021104. PMC 1156860. PMID 8788955.
- ^ Salenius S, Portin K, Kajola M, Salmelin R, Hari R (June 1997). "Cortical control of human motoneuron firing during isometric contraction". Neyrofiziologiya jurnali. 77 (6): 3401–5. doi:10.1152/jn.1997.77.6.3401. PMID 9212286. S2CID 2178927.
- ^ Baker SN, Olivier E, Lemon RN (May 1997). "Coherent oscillations in monkey motor cortex and hand muscle EMG show task-dependent modulation". Fiziologiya jurnali. 501 ( Pt 1) (1): 225–41. doi:10.1111/j.1469-7793.1997.225bo.x. PMC 1159515. PMID 9175005.
- ^ Boonstra TW, Danna-Dos-Santos A, Xie HB, Roerdink M, Stins JF, Breakspear M (December 2015). "Muscle networks: Connectivity analysis of EMG activity during postural control". Ilmiy ma'ruzalar. 5: 17830. Bibcode:2015NatSR...517830B. doi:10.1038/srep17830. PMC 4669476. PMID 26634293.
- ^ Kerkman JN, Daffertshofer A, Gollo LL, Breakspear M, Boonstra TW (June 2018). "Network structure of the human musculoskeletal system shapes neural interactions on multiple time scales". Ilmiy yutuqlar. 4 (6): eaat0497. Bibcode:2018SciA....4..497K. doi:10.1126/sciadv.aat0497. PMC 6021138. PMID 29963631.
- ^ Rubino D, Robbins KA, Hatsopoulos NG (December 2006). "Propagating waves mediate information transfer in the motor cortex". Tabiat nevrologiyasi. 9 (12): 1549–57. doi:10.1038/nn1802. PMID 17115042. S2CID 16430438.
- ^ Heitmann S, Boonstra T, Gong P, Breakspear M, Ermentrout B (2015). "The rhythms of steady posture: Motor commands as spatially organized oscillation patterns". Neurocomputing. 170: 3–14. doi:10.1016/j.neucom.2015.01.088.
- ^ Heitmann S, Boonstra T, Breakspear M (October 2013). "A dendritic mechanism for decoding traveling waves: principles and applications to motor cortex". PLOS hisoblash biologiyasi. 9 (10): e1003260. Bibcode:2013PLSCB...9E3260H. doi:10.1371/journal.pcbi.1003260. PMC 3814333. PMID 24204220.
- ^ Allum JH, Dietz V, Freund HJ (May 1978). "Neuronal mechanisms underlying physiological tremor". Neyrofiziologiya jurnali. 41 (3): 557–71. doi:10.1152/jn.1978.41.3.557. PMID 660226.
- ^ Vallbo AB, Wessberg J (September 1993). "Organization of motor output in slow finger movements in man". Fiziologiya jurnali. 469: 673–91. doi:10.1113/jphysiol.1993.sp019837. PMC 1143894. PMID 8271223.
- ^ Gross J, Timmermann L, Kujala J, Dirks M, Schmitz F, Salmelin R, Schnitzler A (February 2002). "The neural basis of intermittent motor control in humans". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 99 (4): 2299–302. Bibcode:2002PNAS...99.2299G. doi:10.1073/pnas.032682099. PMC 122359. PMID 11854526.
- ^ Buszaki G (2006). Rhythms of the brain. Oksford universiteti matbuoti.
- ^ Nyhus E, Curran T (June 2010). "Functional role of gamma and theta oscillations in episodic memory". Neuroscience and Biobehavioral Reviews. 34 (7): 1023–35. doi:10.1016/j.neubiorev.2009.12.014. PMC 2856712. PMID 20060015.
- ^ Rutishauser U, Ross IB, Mamelak AN, Schuman EM (April 2010). "Human memory strength is predicted by theta-frequency phase-locking of single neurons" (PDF). Tabiat. 464 (7290): 903–7. Bibcode:2010Natur.464..903R. doi:10.1038/nature08860. PMID 20336071. S2CID 4417989.
- ^ Feller MB (July 2009). "Retinal waves are likely to instruct the formation of eye-specific retinogeniculate projections". Asab rivojlanishi. 4: 24. doi:10.1186/1749-8104-4-24. PMC 2706239. PMID 19580682.
- ^ McAuley JH, Marsden CD (August 2000). "Physiological and pathological tremors and rhythmic central motor control". Miya. 123 ( Pt 8) (8): 1545–67. doi:10.1093/brain/123.8.1545. PMID 10908186.
- ^ Shusterman V, Troy WC (June 2008). "From baseline to epileptiform activity: a path to synchronized rhythmicity in large-scale neural networks". Jismoniy sharh E. 77 (6 Pt 1): 061911. Bibcode:2008PhRvE..77f1911S. doi:10.1103/PhysRevE.77.061911. PMID 18643304.
- ^ Vanneste S, Song JJ, De Ridder D (March 2018). "Thalamocortical dysrhythmia detected by machine learning". Tabiat aloqalari. 9 (1): 1103. Bibcode:2018NatCo...9.1103V. doi:10.1038/s41467-018-02820-0. PMC 5856824. PMID 29549239.
- ^ Birbaumer N (November 2006). "Breaking the silence: brain-computer interfaces (BCI) for communication and motor control". Psychophysiology. 43 (6): 517–32. doi:10.1111/j.1469-8986.2006.00456.x. PMID 17076808.
- ^ Vidal JJ (1973). "Toward direct brain-computer communication". Biofizika va bioinjiniring yillik sharhi. 2: 157–80. doi:10.1146/annurev.bb.02.060173.001105. PMID 4583653.
- ^ Bozinovski S, Sestakov M, Bozinovska L (November 1988). "Using EEG alpha rhythm to control a mobile robot.". Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society. New Orleans: IEEE. pp. 1515–1516. doi:10.1109/IEMBS.1988.95357. ISBN 0-7803-0785-2. S2CID 62179588.
- ^ Bozinovski S (August 1990). "Mobile robot trajectory control: From fixed rails to direct bioelectric control.". In Kaynak O (ed.). Proceedings of the IEEE International Workshop on Intelligent Motion Control. 2. Istanbul: IEEE. pp. 463–467. doi:10.1109/IMC.1990.687362. S2CID 60642344.
- ^ Lebedev M (2016). "Augmentation of sensorimotor functions with neural prostheses". Opera Medica et Physiologica. 2 (3–4): 211–227. doi:10.20388/OMP.003.0035 (inactive 2020-10-21).CS1 maint: DOI inactive as of October 2020 (havola)
- ^ Lebedev MA, Nicolelis MA (April 2017). "Brain-Machine Interfaces: From Basic Science to Neuroprostheses and Neurorehabilitation". Physiological Reviews. 97 (2): 767–837. doi:10.1152/physrev.00027.2016. PMID 28275048.
Qo'shimcha o'qish
- Buzsáki G (2006). Rhythms of the Brain. Oksford universiteti matbuoti. ISBN 978-0-19-530106-9.
- Freeman W (1975). Mass Action in the Nervous System. Akademik matbuot. ISBN 978-0124120471. Arxivlandi asl nusxasi on 2015-07-05.