Plazmalogen - Plasmalogen
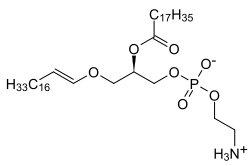
Plazmalogenlar ning subklassi efir fosfolipidlari odatda hujayra membranalarida uchraydi asabiy, immunitetga ega va yurak-qon tomir tizimlari. Ikkita turi mavjud efir fosfolipidlari, plazmanil va plazmenil. Plazalogenni o'z ichiga olgan plazmenil-fosfolipidlar va o'z o'rnida efir bog'lanishiga ega SN1 ga alkenil guruh. Plazmanil-fosfolipidlar, xususan, an efir S holatidagi bog'lanishN1 dan angacha alkil guruh.[1][2][3]
Sutemizuvchilarda SN1 pozitsiyasi odatda C16: 0, C18: 0 yoki C18: 1 yog'li spirtlaridan olinadi SN2 pozitsiyani ko'pincha ko'p to'yinmagan egallaydi yog 'kislotalari (PUFAlar). Sutemizuvchi plazalogenlarda mavjud bo'lgan eng keng tarqalgan bosh guruhlari etanolamin (belgilangan plazmeniletalominlar) yoki xolin (belgilangan plazmenilxolinlar).
Vazifalar
Plazmalogenlar insonning ko'plab to'qimalarida uchraydi, xususan asab, immun va yurak-qon tomir tizimlarida boyitiladi.[1][2][3] Insonning yurak to'qimalarida deyarli 30-40% xolin glitserofosfolipidlar plazmalogenlardir. Voyaga etgan odamning qalbidagi glitserofosfolipidlarning 32%, miyadagi 20% va miyelin qobig'ining 70% gacha etanolamin glitserofosfolipidlari plazalogenlar ekanligi yanada yorqinroq.[4]
Plazmalogenlarning funktsiyalari hali to'liq yoritilmagan bo'lsa ham, ular sutemizuvchilar hujayralarini zararli ta'siridan himoya qilishi mumkinligi isbotlangan. reaktiv kislorod turlari.[1][2][3] Bunga qo'shimcha ravishda, ular membrana dinamikasining signal beruvchi molekulalari va modulyatorlari sifatida ishtirok etgan.
Tarix
Plazmalogenlarni birinchi marta Feulgen va Voit 1924 yilda to'qima bo'limlarini o'rganish asosida tavsiflashgan.[1] Ular ushbu to'qima qismlarini kislota yoki bilan davolashdi simob xlorid yadroni bo'yash usulining bir qismi sifatida. Buning natijasida hosil olish uchun plazmalogen vinil-efir aloqasi uzildi aldegidlar. O'z navbatida, ikkinchisi a bilan reaksiyaga kirishdi fuksin -oltingugurt kislotasi bu yadro binoni usulida ishlatiladigan dog 'va hujayralar sitoplazmasi ichida rangli birikmalar paydo bo'lishiga olib keldi. Plazmalogenlarga ushbu rangli birikmalar hujayraning "plazmal" yoki ichki qismida bo'lganligi asosida nom berilgan.[1]
Biosintez

Plazalogenlarning biosintezi (PL) ning birikmasidan boshlanadi peroksizomal matritsa fermentlari GNPAT (glitseron fosfatatsil transferaza) va AGPS (alkil-glitseron fosfat sintaz) peroksizomal membrananing luminal tomonida.[5]Ushbu ikki ferment samaradorlikni oshirish uchun bir-biri bilan jismoniy ta'sir o'tkazishi mumkin. Shuning uchun, fibroblastlar AGPS faoliyatisiz GNPAT darajasi va faolligi pasaygan.[6][7]
Biyosintezning birinchi bosqichi GNPAT tomonidan katalizlanadi. Ushbu ferment asilatlar dihidroksietonfosfat (DHAP) sn-1 holatida. Buning ortidan AGPS tomonidan asil guruhining alkil guruhiga almashinishi kuzatiladi.[8]Keyin 1-alkil-DHAP asil / alkil-DHAP bilan 1-O-alkil-2-gidroksi-sn-glitserofosfat (GPA) ga kamayadi. reduktaza ikkala peroksizomal va endoplazmatik retikulum (ER) membranalar.[9]Boshqa barcha modifikatsiyalar ER-da sodir bo'ladi. U erda va asil guruh alkil / asil GPA bilan sn-2 holatiga joylashtiriladi asiltransferaza va fosfat guruhi a tomonidan olib tashlanadi fosfatidat kislota fosfataza hosil bo'lib, 1-O-alkil-2-atsil-sn-glitserol hosil qiladi.
CDP-etanolamin yordamida a fosfotransferaza 1-O-alkil-2-asil-sn-GPEtn hosil qiladi. Elektron tashish tizimi va plazmaniletanolamin desaturaza ta'sirida alkil guruhining 1- va 2-pozitsiyalarida dehidrogenatsiyadan so'ng plazalogenlarning vinil efir aloqasi hosil bo'ladi va plazmaniletanolamin desaturazasiga mos keladigan oqsil aniqlanib, bakteriyalar va PEDS1 (TMEM189) tarkibida CarF deb nomlanadi. ) odamlarda (va hayvonlarda)[10],[11].Plasmenilxolin 1-O-alkil-2-atsil-sn-glitserindan hosil bo'ladi. xolin fosfotransferaza. Plazmenilxolin desaturaza yo'qligi sababli xolin plazmalogenlari faqat keyin hosil bo'lishi mumkin gidroliz ning etanolamin Xolin fosfotransferaza va CDP xolin bilan o'zgartirilishi mumkin bo'lgan PL-dan 1-O- (1Z-alkenil) -2-asil-sn-glitseringa.[12][13]
Patologiya
Peroksizom biogenezining buzilishi ko'pincha plazalogen biosintezi buzilganligi bilan ajralib turadigan autosomal retsessiv kasalliklardir. Bunday holatlarda plazalogen biosintezining dastlabki bosqichlari uchun zarur bo'lgan peroksizomal ferment GNPAT, faol bo'lmagan joyda sitoplazmasiga noto'g'ri joylashtirilgan. Bundan tashqari, genetik mutatsiyalar GNPAT yoki AGPS genlar plazmalogen etishmovchiligiga olib kelishi mumkin, bu esa rivojlanishiga olib keladi rizomel xondrodisplaziyasi punktatasi (RCDP) navbati bilan 2 yoki 3 turi.[14] Bunday hollarda ikkala nusxa GNPAT yoki AGPS Kasallik namoyon bo'lishi uchun gen mutatsiyaga uchragan bo'lishi kerak. Peroksizom biogenezi buzilishidan farqli o'laroq, RCDP2 va RCDP3 bemorlarida peroksizom yig'ilishining boshqa jihatlari odatiy holdir, chunki ular juda uzun zanjirli yog 'kislotalarini metabolizm qilish qobiliyatiga ega. Kuchli plazmalogen etishmovchiligi bo'lgan shaxslar tez-tez g'ayritabiiy nevrologik rivojlanishni, skeletning malformatsiyasini, nafas olishning buzilishini va kataraktni ko'rsatadilar.[15]
Plazmalogen darajasining etishmasligi patologiyani keltirib chiqaradi Zellveger sindromi.[13]
Plazmalogen-nokaut sichqonlar hibsga olish kabi o'xshash o'zgarishlarni ko'rsating spermatogenez, rivojlanishi katarakt va markaziy asab tizimidagi nuqsonlar miyelinatsiya.[16][17]
Yallig'lanish paytida
Yallig'lanish paytida neytrofil olinadi miyeloperoksidaza ishlab chiqaradi gipoxlorli kislota (HOCl). HOCl oksidlanishni keltirib chiqaradi xlorlash bilan reaksiyaga kirishib sn-1 zanjiridagi plazmalogenlarning vinil efir bog'lanish[18] Hozirgi vaqtda bir nechta tadqiqotchilar xlorli lipidlarning patologiyaga ta'sirini tekshirmoqdalar.
Mumkin bo'lgan kasallik havolalari
Plazmalogenni tahlil qilishning yaxshi usullarining etishmasligi olimlarga plazalogenning RCDP va Zellveger spektridan tashqari odam kasalliklarida qanday ta'sir qilishi mumkinligini baholashda qiyinchiliklarni keltirib chiqardi.[13] Odamlarda past plazmalogenlarning patologiyada ishtirok etishiga oid ba'zi dalillar mavjud bronxopulmoner displazi, bu muhim komplikasiyadir erta tug'ilish.[13] va bitta tadqiqot shuni ko'rsatadiki, odamlarda plazmalogen darajasi kamayadi KOAH chekmaydiganlarga nisbatan chekadiganlar. Inson va hayvonlardan miyada plazmalogenlarning miqdori kamayganligi to'g'risida ba'zi dalillar mavjud neyrodejenerativ kasalliklar shu jumladan Altsgeymer kasalligi, Parkinson kasalligi, Nemann-Pick kasalligi, C turi, Daun sindromi va skleroz, bu nedenselmi yoki o'zaro bog'liqmi, aniq emas.[13]
Evolyutsiya
Sutemizuvchilardan tashqari plazmalogenlar ham uchraydi umurtqasizlar va bitta hujayrali organizmlar protozoyanlar. Ular orasida bakteriyalar ular ko'plab anaerob turlarida, shu jumladan topilgan Klostridiya, Megasfera va Veillonella. Aerob bakteriyalar orasida plazalogenlar miksobakteriyalarda uchraydi va ularning vinil efir bog'lanishini hosil qilish uchun zarur bo'lgan plazmaniletanolamin desaturaza (CarF) va shu sababli plazmalogen odamlarda (va hayvonlarda) TMEM189 sifatida saqlanib qoladi.[19]. Plazmalogenlarning biosintetik yo'llari aerob va anaerob organizmlarda turlicha ekanligi asosida murakkab evolyutsion tarixga ega ekanligi isbotlangan.[20]
Yaqinda qizil qon tanachalari ekanligini isbotladilar odamlar va maymunlar (shimpanze, bonobos, gorilla va orangutanlar ) plazmalogen tarkibida farqlarga ega.[3] Umumiy RBC plazmalogen darajasi odamlarda bonobos, shimpanze yoki gorillalarga qaraganda pastroq, ammo orangutanlarga qaraganda yuqori ekanligi aniqlandi. Ushbu turlarning gen ekspression ma'lumotlari mualliflarning boshqa odam va buyuk maymun hujayralari va to'qimalari plazmalogen darajasida farq qiladi degan fikrlarini keltirib chiqardi. Ushbu potentsial farqlarning natijalari noma'lum bo'lsa-da, to'qima plazmalogenlaridagi turlarning xilma-xilligi organlarning funktsiyalari va ko'plab biologik jarayonlarga ta'sir qilishi mumkin.
Adabiyotlar
- ^ a b v d e Nagan, N .; Zoeller, R. A. (2001). "Plazmalogenlar: biosintez va funktsiyalari". Lipid tadqiqotida taraqqiyot. 40 (3): 199–229. doi:10.1016 / S0163-7827 (01) 00003-0. PMID 11275267.
- ^ a b v Gorgas, K .; Teygler, A .; Komljenovich, D. Faqat, W. W. (2006). "Efirda lipid etishmaydigan sichqon: plazmalogen funktsiyalarini kuzatish". Biochimica et Biofhysica Acta (BBA) - Molekulyar hujayralarni tadqiq qilish. 1763 (12): 1511–1526. doi:10.1016 / j.bbamcr.2006.08.038. PMID 17027098.
- ^ a b v d Mozer, A. B.; Steinberg, S. J.; Uotkins, P. A .; Mozer, X. V.; Ramasvami, K .; Zigmund, K. D .; Li, D. R .; Eli, J. J .; Rayder, O. A .; Hacia, J. G. (2011). "Inson va buyuk maymun eritrotsitlari plazalogen darajasi va tarkibi bilan farq qiladi". Sog'liqni saqlash va kasallikdagi lipidlar. 10: 101. doi:10.1186 / 1476-511X-10-101. PMC 3129581. PMID 21679470.
- ^ Farukki, A. A .; Horrocks, L. A. (2001). "Plazmalogenlar: normal va shikastlangan neyronlar va gliyalardagi membranalarning ishchi lipidlari". Neuroscientist: Neurobiology, Nevrologiya va Psixiatriyani keltiruvchi sharhlar jurnali. 7 (3): 232–245. doi:10.1177/107385840100700308. PMID 11499402. S2CID 85868497.
- ^ P. Brites, H.R.Voterxem, R.J. Sog'liqni saqlash va kasallikdagi plazalogenlarning sarson-sargardonligi, funktsiyalari va biosintezi, Biochim. Biofiz. Acta 1636 (2004) 219-231.
- ^ J. Biermann, VW. Faqat, R.J. Wanders, H. Van Den Bosch, Alkil-dihidroksiaseton fosfat sintaz va dihidroksieton fosfat asiltransferaza peroksizomalarda oqsil kompleksini hosil qiladi, Ev. J. Biokimyo. 261 (1999) 492-499
- ^ D. Hardeman, H. van den Bosch, Eter fosfolipid biosintezi topografiyasi, Biochim. Biofiz. Acta 1006 (1989) 1-8.
- ^ A.J. Jigarrang, F. Snayder, Alkildihidroksiaseton-P sintaz. Solubilizatsiya, qisman tozalash, yangi tahlil usuli va ping-pong mexanizmi uchun dalillar, J. Biol. Kimyoviy. 257 (1982) 8835–8839
- ^ P.F. Jeyms, A. Leyk, A.K. Xajra, L.K. Larkins, M. Robinson, F.G. Buchanan, RA Zoeller, asil / alkil-dihidroksia-ton-fosfat reduktaza faolligi etishmovchiligi bo'lgan hayvon hujayralari mutanti. Eter bilan bog'langan va diatsil glitserolipidlarning biosinteziga ta'siri, J. Biol. Kimyoviy. 272 (1997) 23540–23546
- ^ Gallego-García A, Monera-Girona AJ, Pajares-Martines E, Bastida-Martines E, Peres-Castanño R, Iniesta AA, Fontes M, Padmanabhan S, Elías-Arnanz M. "Bakterial nur reaktsiyasi inson uchun yetim desaturazani aniqlaydi. plazmalogen sintezi "(2019) Ilm-fan 366 (6461) 128-132, doi: 10.1126 / science.aay1436, PMID = 31604315
- ^ Verner, ER; Keller, MA; Dengizchi, S; Lackner, K; Koch, J; Hermann, M; Coassin, S; Golderer, G; Verner-Felmayer, G; Zoeller, RA; Xulo, N; Berger, J; Watschinger, K (2020 yil 7 aprel). " TMEM189 gen plazmaniletanolamin desaturazani kodlaydi, bu esa o'ziga xos vinil efir qo'shaloq bog'lanishini plazmalogenlarga kiritadi ". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 117 (14): 7792–7798. doi:10.1073 / pnas.1917461117. PMID 32209662.
- ^ T.C. Li, plazalogenlarning biosintezi va mumkin bo'lgan biologik funktsiyalari, Biochim. Biofiz. Acta 1394 (1998) 129-145
- ^ a b v d e Braverman, NE; Moser, AB (sentyabr, 2012). "Sog'liqni saqlash va kasallikdagi plazalogen lipidlarining funktsiyalari". Biochimica et Biofhysica Acta (BBA) - Kasallikning molekulyar asoslari. 1822 (9): 1442–52. doi:10.1016 / j.bbadis.2012.05.008. PMID 22627108.
- ^ Wanders, R .; Waterham, H. (2006). "Peroksizomal kasalliklar: yagona peroksizomal ferment etishmovchiligi". Biochimica et Biofhysica Acta (BBA) - Molekulyar hujayralarni tadqiq qilish. 1763 (12): 1707–20. doi:10.1016 / j.bbamcr.2006.08.010. PMID 17055078.
- ^ Rhizomelic Chondrodysplasia Punctata Type 1. Mualliflar Braverman NE, Moser AB, Steinberg SJ. Tahrirlovchilar: Pagon RA, Adam MP, Bird TD, Dolan CR, Fong CT, Smith RJH, Stephens K, muharrirlar. GeneReviews manbasi [Internet]. Sietl (WA): Vashington universiteti, Sietl; 1993-2014. 2001 yil 16-noyabr
- ^ Gorgas K, Teigler A, Komljenovich D, Just WW. Efirda lipid etishmaydigan sichqon: plazalogen funktsiyalarini kuzatish. Biochim Biophys Acta. 2006 yil dekabr; 1763 (12): 1511-26
- ^ Rodemer C, Thai TP, Brugger B, Kaercher T, Verner H, Nave KA, Wieland F, Gorgas K, Just WW. Eter lipid biosintezini faolsizlantirish erkaklarning bepushtligini, ko'z rivojlanishidagi nuqsonlarni va sichqonlardagi optik asab gipoplaziyasini keltirib chiqaradi. 2003 yil 1-avgust; 12 (15): 1881-95.
- ^ Albert, Kerolin J.; Krouli, Jan R.; Xsu, Fong-Fu; Thukkani, Arun K.; Ford, Devid A. (2001-06-29). "Myeloperoksidaza tomonidan ishlab chiqarilgan reaktiv xlorlovchi turlar, plazmalogenlarning vinil efir bog'lanishiga yo'naltirilgan. 2-XLOROXEXADEKANALNI ISTANFAK QILISh". Biologik kimyo jurnali. 276 (26): 23733–23741. doi:10.1074 / jbc.M101447200. ISSN 0021-9258. PMID 11301330.
- ^ Gallego-García A, Monera-Girona AJ, Pajares-Martines E, Bastida-Martines E, Peres-Castanño R, Iniesta AA, Fontes M, Padmanabhan S, Elías-Arnanz M. "Bakterial nur reaktsiyasi inson uchun yetim desaturazani aniqlaydi. plazmalogen sintezi "(2019) Ilm-fan 366 (6461) 128-132, doi: 10.1126 / science.aay1436, PMID = 31604315
- ^ Goldfine, H. (2010). "Plazalogenlarning evolyutsiyada paydo bo'lishi, yo'q bo'lib ketishi va paydo bo'lishi". Lipid tadqiqotida taraqqiyot. 49 (4): 493–498. doi:10.1016 / j.plipres.2010.07.003. PMID 20637230.
Tashqi havolalar
- Plazmalogenlar AQSh Milliy tibbiyot kutubxonasida Tibbiy mavzu sarlavhalari (MeSH)