Biologik hayot aylanishi - Biological life cycle


Yilda biologiya, a biologik hayot aylanishi (yoki shunchaki hayot davrasi yoki hayot davrasi biologik kontekst aniq bo'lsa) - bu organizm boshlang'ich holatiga qaytib, shakllanadigan bir qator o'zgarishlar. "Kontseptsiya hayot tarixi bilan chambarchas bog'liq, rivojlanish va ontogenez, ammo yangilanishni ta'kidlashda ulardan farq qiladi. "[1][2] Shaklning o'zgarishi o'sishni o'z ichiga olishi mumkin, jinssiz ko'payish, yoki jinsiy ko'payish.
Ba'zi organizmlarda turlarning har xil "avlodlari" hayot aylanishi davomida bir-birining o'rnini egallaydi. Uchun o'simliklar va ko'p suv o'tlari, ikkita ko'p hujayrali bosqich mavjud va hayot aylanishi deb ataladi avlodlar almashinuvi. Atama hayot tarixi ko'pincha, ayniqsa, kabi organizmlar uchun ishlatiladi qizil suv o'tlari ikkita emas, balki uchta ko'p hujayrali bosqichga (yoki undan ko'p) ega.[3]
Jinsiy ko'payishni o'z ichiga olgan hayot tsikllari o'zgarishni o'z ichiga oladi gaploid (n) va diploid (2n) bosqichlari, ya'ni o'zgarishi ploidy ishtirok etadi. Diploid bosqichdan gaploid bosqichga qaytish uchun, mayoz sodir bo'lishi kerak. O'zgarishlar haqida ploidy, tsikllarning 3 turi mavjud:
- haplontik hayot aylanishi - gaploid bosqichi ko'p hujayrali va diploid bosqichi bitta hujayradan iborat bo'lib, meyoz "zigotik" hisoblanadi.
- diplomatik hayot aylanishi - diploid bosqichi ko'p hujayrali va gaploiddir jinsiy hujayralar hosil bo'ladi, mayoz "gametik".
- haplodiplontika hayot aylanishi (shuningdek, diplohaplontik, diplobiontik, yoki dibiontik hayot aylanishi) - ko'p hujayrali diploid va gaploid bosqichlari sodir bo'ladi, meyoz "sporik".
Tsikllar qachon farq qiladi mitoz (o'sish) sodir bo'ladi. Zigotik mayoz va gametik meyoz bitta mitoz bosqichga ega: mitoz paytida paydo bo'ladi n zigotik mayozdagi faza va 2 paytidan gametozli meyozda faza. Shuning uchun zigotik va gametik mayoz birgalikda "haplobiontik" deb nomlanadi (bitta mitotik faza, gapplontik bilan adashtirmaslik kerak). Boshqa tomondan, sporik mayozda mitoz ikki bosqichda, ham diploid, ham gaploid bosqichda bo'lib, "diplobiontik" deb nomlanadi (diplomatik bilan adashtirmaslik kerak).
Kashfiyot
O'rganish ko'payish va rivojlanish organizmlarda ko'plab botaniklar va zoologlar tomonidan olib borilgan.
Wilhelm Hofmeister buni namoyish etdi avlodlar almashinuvi o'simliklarni birlashtiradigan xususiyat bo'lib, ushbu natijani 1851 yilda nashr etdi (qarang) o'simliklarning jinsiyligi ).
Hayotiy tsikllarni tavsiflash uchun ishlatilgan ba'zi atamalar (haplobiont va diplobiont) dastlab Nils Svedelius tomonidan suv o'tlari uchun taklif qilingan va keyinchalik boshqa organizmlar uchun ishlatilgan.[4][5] Da ishlatiladigan boshqa atamalar (autogamiya va gamontogamiya) protist hayot tsikllari Karl Gottlieb Grell tomonidan kiritilgan.[6] Turli organizmlarning murakkab hayotiy tsikllarining tavsifi g'oyalarni rad etishga yordam berdi o'z-o'zidan paydo bo'ladigan avlod 1840 va 1850 yillarda.[7]
Gaplontik hayot aylanishi
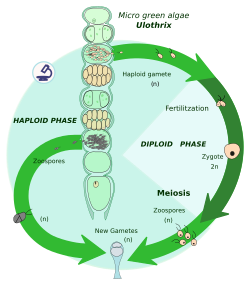
Zigotik mayoz - bu a mayoz a zigota darhol keyin karyogamiya, bu ikkitasining birlashishi hujayra yadrolari. Shunday qilib, organizm diploid fazasini tugatadi va bir nechta gaploid hujayralarni hosil qiladi. Ushbu hujayralar bo'linadi mitotik tarzda kattaroq, ko'p hujayrali shaxslarni yoki ko'proq gaploid hujayralarni hosil qilish uchun. Ushbu shaxslardan yoki hujayralardan qarama-qarshi ikkita jinsiy hujayralar turi (masalan, erkak va ayol) zigota bo'lib birlashadi.
Butun tsiklda zigotlar yagona diploid hujayradir; mitoz faqat gaploid fazada sodir bo'ladi.
Mitoz natijasida hujayralar yoki hujayralar haplonts hisoblanadi, shuning uchun bu hayot tsikli haplontik hayot aylanishi deb ham ataladi. Haplonts:
- Yilda arxeplastidanlar: biroz yashil suv o'tlari (masalan, Xlamidomonalar, Zigema, Chara )[8]
- Yilda stramenopillar: biroz oltin suv o'tlari[8]
- Yilda alveolitlar: ko'p dinoflagellatlar, masalan, Ceratium, Gymnodinium, ba'zilari apikompleksanlar (masalan, Plazmodium )[9]
- Yilda rizariylar: biroz evgliflar,[10] astsetosporlar
- Yilda qazib oladi: biroz parabasalidlar[11]
- Yilda amebozoyanlar: Diktiosteliya[8]
- Yilda opisthokonts: eng qo'ziqorinlar (biroz xitridlar, zigomitsetlar, biroz ascomitsetlar, basidiomitsetalar )[8][12]:15
Diplomatik hayot aylanishi

Gametik meyozda darhol bo'linish o'rniga meiotik jihatdan gaploid hujayralarni hosil qilish uchun zigota bo'linadi mitotik tarzda ko'p hujayrali diploid individual yoki bir hujayrali diploid hujayralar guruhini ishlab chiqarish. Diploid hujayralar hujayralari keyinchalik gaploid hujayralarni hosil qilish uchun yoki meozga uchraydi jinsiy hujayralar. Gaploid hujayralar yana (mitoz bilan) bo'linib, ko'plab xamirturushlarda bo'lgani kabi ko'proq gaploid hujayralarni hosil qilishi mumkin, ammo gaploid fazasi hayot aylanish davrining asosiy bosqichi emas. Ko'pgina diplomlarda mitoz faqat diploid fazada sodir bo'ladi, ya'ni gametalar odatda tez shakllanadi va diploid zigotalar hosil qilish uchun birlashadi.
Butun tsiklda jinsiy hujayralar odatda bitta gaploid hujayralar bo'lib, mitoz odatda faqat diploid fazada sodir bo'ladi.
Diploid ko'p hujayrali shaxs diplontdir, shuning uchun gametli meyoz ham diplomatik hayot aylanishi deb ataladi. Diplomatlar:
- Yilda arxeplastidanlar: biroz yashil suv o'tlari (masalan, Cladophora glomerata,[13] Asetabulariya[8])
- Yilda stramenopillar: biroz jigarrang suv o'tlari (the Fucales ammo, ularning hayotiy tsiklini kuchli heteromorfik-diplohaplontik deb talqin qilish mumkin, juda past darajada gametofit fazasi bilan, gullarni o'simliklar ),[12]:207 biroz ksantofitlar (masalan, Vaucheria ),[12]:124 eng diatomlar,[11] biroz oomitsetlar (masalan, Saprolegnia, Plasmopara viticola ),[8] opalin,[11] biroz "heliozoans "(masalan, Aktinofrizlar, Aktinosferium )[11][14]
- Yilda alveolitlar: kirpiklar [11]
- Yilda qazib oladi: biroz parabasalidlar [11]
- Yilda opisthokonts: hayvonlar, biroz qo'ziqorinlar (masalan, ba'zilari ascomitsetlar )[8]
Haplodiplontik hayot aylanishi
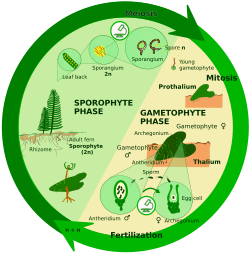
Sporik mayozda (shuningdek, uni vositachi meyoz deb ham atashadi), zigota mitotik bo'linib, ko'p hujayrali diploid hosil qiladi. sporofit. Sporofit mayoz orqali sporalarni hosil qiladi shuningdek keyin mitotik ishlab chiqaruvchi gaploid shaxslarni ajratib oling gametofitlar. Gametofitlar gametalarni mitoz orqali hosil qiladi. Ba'zi o'simliklarda gametofit nafaqat kichik o'lchamli, balki qisqa muddatli ham bo'ladi; boshqa o'simliklarda va ko'plab suv o'tlarida gametofit hayot aylanishining "dominant" bosqichidir.
Haplodiplonts:
- Yilda arxeplastidanlar: qizil suv o'tlari (ikkita sporofit avlodiga ega), ba'zilari yashil suv o'tlari (masalan, Ulva ), quruqlikdagi o'simliklar[8]
- Yilda stramenopillar: eng jigarrang suv o'tlari[8]
- Yilda rizariylar: ko'p foraminiferanlar,[11] plazmodioforomitsetlar[8]
- Yilda amoebozoa: miksogastridlar
- Yilda opisthokonts: biroz qo'ziqorinlar (biroz xitridlar, biroz ascomitsetlar kabi pivo xamirturushlari )[8]
- Boshqa eukaryotlar: gappofitlar[11]
Ba'zi hayvonlarda a jinsni aniqlash tizimi deb nomlangan gaplodiploid, ammo bu haplodiplontik hayot aylanishi bilan bog'liq emas.
Vegetativ meyoz
Biroz qizil suv o'tlari (kabi Bonnemisoniya[15] va Lemaniya ) va yashil suv o'tlari (masalan Prasiola ) vegetativ mayozga ega, shuningdek kam uchraydigan hodisa bo'lgan somatik mayoz deb ham ataladi.[12]:82 Vegetativ meyoz haplodiplontikada, shuningdek diplomatik hayotda ham bo'lishi mumkin. Gametofitlar sporofitga yopishgan va uning bir qismi bo'lib qoladi. Vegetativ (reproduktiv bo'lmagan) diploid hujayralar vegetativ gaploid hujayralarni hosil qilib, meozga uchraydi. Ular ko'plab mitozlarga uchraydi va jinsiy hujayralar hosil qiladi.
Vegetativ diploidlanish deb ataladigan boshqa hodisa apomiksis, ba'zilarida uchraydi jigarrang suv o'tlari (masalan, Elachista stellaris).[16] O'simlikning gaploid qismidagi hujayralar o'z-o'zidan xromosomalarini ko'paytirib, diploid to'qima hosil qiladi.
Parazitar hayot aylanishi
Parazitlar bir yoki bir nechtasining ekspluatatsiyasiga bog'liq mezbonlar. Bir nechta xostlarni yuqtirishlari kerak bo'lganlar turlari ularning hayot tsikllarini yakunlash uchun murakkab yoki bilvosita hayot tsikllari deyiladi, bitta turga yuqadiganlar esa to'g'ridan-to'g'ri hayot aylanishiga ega.
Agar parazit hayot tsiklini yakunlash uchun ma'lum bir uy egasini yuqtirishga to'g'ri kelsa, u holda an deyiladi majburiy parazit o'sha uy egasi; ba'zan infektsiya bo'ladi fakultativ - parazit o'sha yashaydigan turga yuqtirmasdan yashashi va hayot aylanish jarayonini yakunlashi mumkin. Parazitlar ba'zan o'zlarining hayot tsikllarini yakunlay olmaydigan xostlarni yuqtirishadi; bu tasodifiy xostlar.
Parazitlar jinsiy yo'l bilan ko'payadigan xo'sh, aniqlovchi, yakuniy yoki birlamchi xost deb nomlanadi. O'rta xostlarda parazitlar ko'paymaydi yoki jinsiysiz ko'payadi, lekin parazit har doim bu turdagi xo'shda yangi bosqichga o'tib boradi. Ba'zi hollarda parazit xostga yuqadi, ammo rivojlanishni boshdan kechirmaydi, bu xostlar ma'lum paratenik[17] yoki xostlarni tashish. Paratenik xost parazitning aniq egasiga o'tishi ehtimolini oshirishda foydali bo'lishi mumkin. Masalan, mushuk o'pka qurti (Aelurostrongylus abstrusus) oraliq xost sifatida shilliqqurt yoki salyangozdan foydalanadi; birinchi bosqich lichinkasi mollyuskaga kirib, uchinchi darajali lichinkaga aylanadi, u aniq egasi - mushuk uchun yuqadi. Agar sichqon shilliqqani yeb qo'ysa, uchinchi bosqich lichinkasi sichqonchaning to'qimalariga kiradi, ammo hech qanday rivojlanmaydi.

Evolyutsiya
Hayotiy tsiklning ibtidoiy turida, ehtimol, jinssiz ko'payish bilan gaploid shaxslar bo'lgan.[11] Bakteriyalar va arxey shunga o'xshash hayot tsiklini namoyish eting va ba'zi ökaryotlar ham ko'rinadi (masalan, Kriptofit, Choanoflagellata, ko'p Evglenozoa, ko'p Amebozoa, ba'zilari qizil suv o'tlari, ba'zilari yashil suv o'tlari, nomukammal zamburug'lar, biroz rotifers va boshqa ko'plab guruhlar, albatta haploid emas).[18] Ammo, bu eukaryotlar, ehtimol ibtidoiy aseksual emas, balki jinsiy reproduktsiyasini yo'qotgan yoki u hali kuzatilmagan.[19][20] Ko'pgina eukaryotlar (shu jumladan hayvonlar va o'simliklar) jinssiz ko'payish, hayotiy tsikldagi fakultativ yoki majburiy bo'lishi mumkin, jinsiy ko'payish kamroq yoki tez-tez sodir bo'ladi.[21]
Shuningdek qarang
- Avlodlarning o'zgarishi - O'simliklar va suv o'tlarining ko'payish tsikli
- Apomiksis - Oddiy jinsiy ko'payishni urug'lanmasdan, jinssiz ko'payish bilan almashtirish
- Haplodiploidiya - Jinsiy xromosomalar to'plami soniga qarab belgilanadigan biologik tizim
- Paraseksual tsikl - Genetik materialni meyozsiz o'tkazish uchun nonseksual mexanizm
- Partenogenez - embrionlarning o'sishi va rivojlanishi urug'lanmasdan sodir bo'ladigan jinssiz ko'payishning tabiiy shakli
- Reproduktiv biologiya - ko'paytirishni o'rganadigan biologiya bo'limi
- Mitotik rekombinatsiya
Adabiyotlar
- ^ Grem Bell va Vassiliki Koufopanou (1991). "Kichik organizmlarda hayot tsiklining arxitekturasi". Falsafiy operatsiyalar: Biologiya fanlari. 332 (1262): 81–89. doi:10.1098 / rstb.1991.0035. JSTOR 55494.
- ^ Rodriges, Juliani Kola Fernandes; Godinyo, Xoseane Lima Prado; de Souza, Uanderli (2014). "Inson patogen tripanosomatidlari biologiyasi: epidemiologiya, hayot aylanishi va ultrastruktura". Leyshmaniya va Trypanosoma oqsillari va proteomikasi. Subcellular Biokimyo. Springer Niderlandiya. 74: 1–42. doi:10.1007/978-94-007-7305-9_1. ISBN 978-94-007-7304-2. PMID 24264239.
- ^ Dikson, P.S. 1973 yil. Rodofit biologiyasi. Oliver va Boyd. ISBN 0 05 002485 X
- ^ C. Skottsberg (1961), "Nils Eberxard Svedelius. 1873-1960", Qirollik jamiyati a'zolarining biografik xotiralari, 7: 294–312, doi:10.1098 / rsbm.1961.0023
- ^ Svedelius, N. 1931 yil. Rodofitsiyada yadro fazalari va almashinuvi. In: Beihefte zum Botanischen Centralblatt. Tarmoq 48/1: 38-59.
- ^ L. Margulis (1996), "Evkaryoning kelib chiqishidagi arxeoal-eubakterial qo'shilish: hayotning filogenetik tasnifi", PNAS, 93 (3): 1071–1076, doi:10.1073 / pnas.93.3.1071, PMC 40032, PMID 8577716
- ^ Moselio Schaechter (2009). Mikrobiologiya entsiklopediyasi. Akademik matbuot. 4-jild, p. 85.
- ^ a b v d e f g h men j k Diaz Gonsales, TE, C. Fernandes-Karvaxal Alvares va J.A. Fernandes Prieto. (2004). Kurso-Botanika. Xijon: Trea. Onlayn material: Botanika: Ciclos biológicos de vegetales (Vegetativ hayot tsikllari, ispan tilida). Biología de Organismos y Sistemas Departamento, Universidad de Oviedo.
- ^ Sinden, R. E.; Xartli, R. H. "Bezgak parazitlarining meiotik bo'linishini aniqlash". Protozoologiya jurnali.
- ^ Lahr DJ, Parfrey LW, Mitchell EA, Katz LA, Lara E (iyul 2011). "Amyobaning iffati: ameboid organizmlarda jinsiy aloqaga oid dalillarni qayta baholash". Proc. Biol. Ilmiy ish. 278 (1715): 2083–6. doi:10.1098 / rspb.2011.0289. PMC 3107637. PMID 21429931.
- ^ a b v d e f g h men Ruppert, E. E., Fox, R. S., & Barnes, R. D. (2004). Umurtqasizlar zoologiyasi: funktsional evolyutsion yondashuv. Belmont, Kaliforniya: Tomas-Bruks / Koul, 26-bet.
- ^ a b v d van den Hoek, C., D.G. Mann va X.M. Jahns (1995). Yosunlar: fizikaga kirish. Kembrij universiteti matbuoti (623 bet).
- ^ O. P. Sharma. Yosunlar darsligi, p. 189
- ^ Bell, G. (1989). Protozoa jinsi va o'limi. Kembrij universiteti matbuoti, p. 11, [1].
- ^ Salvador Soler, Noemi; Gomes Garreta, Ameliya; Antonia Ribera Siguan, M. (2009 yil 1-avgust). "Pirat yarim orolidan Bonnemaisonia asparagoides va Bonnemaisonia clavata (Bonnemaisoniales, Rhodophyta) hayot tarixidagi somatik meyoz". Evropa pikologiya jurnali. 44 (3): 381–393. doi:10.1080/09670260902780782.
- ^ Lyuis, RJ (1996). "Jigarrang suv o'tlari xromosomalari". Fikologiya. 35 (1): 19–40. doi:10.2216 / i0031-8884-35-1-19.1.
- ^ Shmidt va Roberts. 1985. Parazitologiya asoslari 3-nashr. Times Mirror / Mosby kollejining nashriyoti
- ^ Xeyvud, P .; Magee, P.T. (1976). "Protistlarda meyoz. Yosunlar, zamburug'lar va protozoalardagi meyozning ba'zi tarkibiy va fiziologik jihatlari". Bakteriologik sharhlar. 40 (1): 190–240. doi:10.1128 / mmbr.40.1.190-240.1976. PMC 413949. PMID 773364.
- ^ Shehre-Banoo Malik; Artur V.Paytling; Lauren M. Stefaniak; Andrew M. Schurko va John M. Logsdon, Jr (2008). "Konservalangan meiotik genlarning inventarizatsiyasi jinsiy aloqada dalillar keltiradi Trichomonas vaginalis". PLOS ONE. 3 (8): e2879. Bibcode:2008PLoSO ... 3.2879M. doi:10.1371 / journal.pone.0002879. PMC 2488364. PMID 18663385.
- ^ Spayder, D .; Lukash, J .; Elias, M. (2015). "Jinsiy aloqa - bu eukaryotik hayotning hamma joyda tarqalgan, qadimiy va o'ziga xos xususiyati". PNAS. 112 (29): 8827–8834. doi:10.1073 / pnas.1501725112. PMC 4517231. PMID 26195746.
- ^ Shon, I .; Martens, K .; van Deyk, P. (2009). Yo'qotilgan jinsiy aloqa: Partenogenezning evolyutsion biologiyasi. Springer. ISBN 9789048127702.
Qo'shimcha o'qish
- Hayotiy tsikllar: Evolyutsion biologning mulohazalari. Jon Tayler Bonner. Princeton University Press, Princeton (Nyu-Jersi).
- Valero, M (1992). "Hayot tsikllarida gaploid va диплоид fazalarining almashinuvi evolyutsiyasi". Ekologiya va evolyutsiya tendentsiyalari. 7 (1): 25–29. doi:10.1016 / 0169-5347 (92) 90195-soat. PMID 21235940.
- Mable, B .; Otto, S. (1998). "Gaploid va diploid fazali hayot tsikllari evolyutsiyasi" (PDF). BioEssays. 20 (6): 453–462. doi:10.1002 / (sici) 1521-1878 (199806) 20: 6 <453 :: aid-bies3> 3.0.co; 2-n.