Xameleyon - Chameleon
| Xameleyonlar | |
|---|---|
 | |
| Hind xameleyoni, Chamaeleo zeylanicus, yilda Mangaon, Maharashtra, Hindiston | |
| Ilmiy tasnif | |
| Qirollik: | Animalia |
| Filum: | Chordata |
| Sinf: | Reptiliya |
| Buyurtma: | Squamata |
| Suborder: | Iguaniya |
| Klade: | Akrodonta |
| Oila: | Chamaeleonidae Rafinesk, 1815 |
| Genera | |
 | |
| Chamaeleonidae ning mahalliy tarqalishi | |
Xameleyonlar yoki xameleyonlar (oila Chamaeleonidae) o'ziga xos va yuqori darajada ixtisoslashgan qoplama ning Eski dunyo kaltakesaklar 2015 yil iyun oyiga qadar tasvirlangan 202 tur bilan.[1] Ushbu turlar turli xil ranglarga ega va ko'plab turlar rangni o'zgartirish qobiliyatiga ega.
Xameleyonlar o'zlari bilan ajralib turadi zigodaktil oyoqlar; ularning juda keng, juda o'zgartirilmagan, tezda ekstrudirovka qilinadigan tillari; ularning chayqaladigan yurishi;[2] peshonalarida va tumshug'ida tepaliklar yoki shoxlar. Ko'pgina turlar, xususan, kattaroq turlari, shuningdek, a oldindan yaroqsiz quyruq. Xameleyonlarning ko'zlari mustaqil ravishda harakatchan, ammo yirtqich narsaga qarab, ular koordinatsiyada oldinga, hayvonga e'tibor qaratadilar. stereoskopik ko'rish.
Xameleonlar toqqa chiqish va vizual ov qilish uchun moslangan. Dan iborat bo'lgan issiq yashash joylarida yashaydilar yomg'ir o'rmoni ga cho'l Afrikada uchraydigan turli xil turlari bilan, Madagaskar, janubiy Evropa va janubiy Osiyo bo'ylab Shri-Lanka. Ular ham bo'lgan tanishtirdi ga Gavayi, Kaliforniya va Florida va ko'pincha uy hayvonlari sifatida saqlanadi.
Etimologiya

Inglizcha so'z xameleyon (/kəˈmiːlmenən/ kuh-MEEL-ee-un ) ning soddalashtirilgan imlosi Lotin xamaeleōn,[3] qarz olish Yunoncha gámias (hamailén),[4] a birikma ning gámáb (xamaí) "yerda"[5] va λέων (lén) "sher".[6][7][8]

Tasnifi
Chamaeleonidae oilasi ikkita kichik oilaga bo'lingan, Brukesiinae va Xamaeleoninae, 1986 yilda Klaver va Bohme tomonidan.[9] Ushbu tasnifga muvofiq, Brukesiinae avlodni o'z ichiga olgan Bruksiya va Remfoleon, shuningdek, keyinchalik nasl ulardan ajralib (Palleon va Rieppeleon ), esa Xamaeleoninae avlodni o'z ichiga olgan Bradypodion, Kalumma, Chamaeleo, Furcifer va Trioceros, shuningdek, keyinchalik nasl ulardan ajralib (Archaius, Nadzikambiya va Kinyongia ). O'sha vaqtdan boshlab, ushbu oilaviy belgining haqiqiyligi ko'p munozaralarga sabab bo'ldi,[10] ko'pgina filogenetik tadqiqotlar, Brookesiinae subfamily oilasining pigmentli xameleonlari emas degan tushunchani qo'llab-quvvatlaydi. monofiletik guruh.[11][12][13][14]
Ba'zi organlar ilgari ushbu subfamilial tasnifni dalillar tamoyilining yo'qligi asosida foydalanishni afzal ko'rishgan bo'lsa-da,[10] keyinchalik ushbu hokimiyat subfamilial bo'linishni tark etdi va endi Chamaeleonidae oilasi bilan hech qanday subfamilalarni tan olmadi.[15]
Biroq, 2015 yilda Glaw subfamilial bo'linishni faqat nasllarni joylashtirib qayta ishladi Bruksiya va Palleon ichida Brukesiinae subfamily, boshqa barcha nasllar joylashtirilgan Xamaeleoninae.[1]

Rang o'zgarishi
Ushbu bo'lim uchun qo'shimcha iqtiboslar kerak tekshirish. (2017 yil avgust) (Ushbu shablon xabarini qanday va qachon olib tashlashni bilib oling) |


Ba'zi xameleyon turlari o'zlarini o'zgartirishga qodir terining ranglanishi. Xameleyonning turli xil turlari ranglarini o'zgartirishga qodir naqsh pushti, ko'k, qizil, to'q sariq, yashil, qora, jigarrang, och ko'k, sariq, firuza va binafsha ranglarning kombinatsiyalari orqali.[16] Xameleyon terisi pigmentlarni o'z ichiga olgan yuzaki qatlamga ega va qatlam ostida hujayralar mavjud guanin kristallar. Xameleonlar guanin kristallari orasidagi bo'shliqni o'zgartirib rangini o'zgartiradi, bu esa terining rangini o'zgartiradigan kristallardan aks etgan nurning to'lqin uzunligini o'zgartiradi.
Xameleonlardagi rang o'zgarishi funktsiyalarga ega kamuflyaj, lekin ko'pincha ijtimoiy signalizatsiya va harorat va boshqa sharoitlarga reaktsiyalarda. Ushbu funktsiyalarning nisbiy ahamiyati sharoitga, shuningdek turga qarab farq qiladi. Rang o'zgarishi xameleonning fiziologik holati va boshqa xameleyonlarga niyatlari to'g'risida signal beradi.[17][18] Chunki xameleyonlar ektotermik, ularning rangini o'zgartirishlarining yana bir sababi - bu tana haroratini tartibga solish, yoki yorug'likni yutish uchun qorong'i rangga va haroratni ko'tarish uchun issiqlikka, yoki yorug'lik va issiqlikni aks ettirish uchun engil rangga, shu bilan tanadagi haroratni barqarorlashtiradi yoki pasaytiradi.[19] Xameleonlar boshqa xameleonlarga tajovuz ko'rsatganda yorqinroq ranglarni namoyish etadi,[20] va ular topshirganda yoki "voz kechganda" quyuq ranglar.[21] Ba'zi turlar, xususan Madagaskar va tropik o'rmonlarda yashovchi ba'zi afrikalik nasllar ko'k rangga ega lyuminestsentsiya ularning bosh suyagida sil kasalligi, suyaklardan kelib chiqqan va ehtimol signalizatsiya rolini bajargan.[22]
Kabi ba'zi turlari Smitning mitti xameleyoni, kamuflyaj uchun ranglarini ularga tahdid soladigan ma'lum yirtqich turlarining (qush yoki ilon) ko'rinishiga mos ravishda sozlang.[23]
Cho'lda yashovchi Namaqua xameleyoni rang o'zgarishini yordam sifatida ham qo'llaydi termoregulyatsiya, salqinroq tongda qora rangga aylanib, issiqlikni samaraliroq yutadi, so'ngra kunning issiq paytida yorug'likni aks ettirish uchun ochroq kul rang bo'ladi. U ikkala rangni bir vaqtning o'zida ko'rsatishi mumkin, o'murtqa chapdan o'ngdan o'ngga ajratilgan.[iqtibos kerak ]
Rangni o'zgartirish mexanizmi
Uzoq vaqt davomida xameleyonlar terisiga pigment o'z ichiga olgan organoidlarning tarqalishi bilan rangini o'zgartiradi deb o'ylar edilar. Biroq, 2014 yilda panter xameleonlari bo'yicha olib borilgan tadqiqotlar shuni ko'rsatdiki, pigment harakati faqat mexanizmning bir qismini anglatadi.[24]
Xameleonlarning terisida ularning rangini va termoregulyatsiyasini boshqaradigan ikkita ustma-ust qatlamlar mavjud. Yuqori qatlam guanin nanokristallarining panjarasini o'z ichiga oladi va bu panjarani hayajonga solib, nanokristallar orasidagi masofani boshqarish mumkin, bu esa o'z navbatida yorug'likning qaysi to'lqin uzunliklari aks etishi va ular yutilishiga ta'sir qiladi. Panjarani hayajonlantirish nanokristallar orasidagi masofani oshiradi va teri yorug'likning uzunroq to'lqin uzunligini aks ettiradi. Shunday qilib, tinch holatda kristallar ko'k va yashil ranglarni aks ettiradi, lekin hayajonlangan holatda sariq, to'q sariq, yashil va qizil kabi uzunroq to'lqin uzunliklari aks etadi.[25]
Xameleyon terisida ba'zi sariq pigmentlar mavjud bo'lib, ular bo'shashgan kristall panjarada aks ettirilgan ko'k rang bilan birlashganda, ularning xameleyonlari bo'shashgan holatida ko'p uchraydigan xarakterli yashil rangga ega bo'ladi. Xameleyon rang palitrasi evolyutsiya va atrof-muhit orqali rivojlandi. O'rmonda yashovchi xameleyonlar asosiy, jigarrang va charred palitraga ega bo'lgan cho'l yoki savanada yashovchilarga nisbatan ancha aniqroq va rang-barang palitraga ega.[26]
Evolyutsiya

Eng qadimgi tasvirlangan xameleyon Anqingosaurus brevicephalus O'rta paleotsendan (taxminan 58,7-61,7) mya ) Xitoy.[27]
Boshqa xameleyon qoldiqlari orasida Chamaeleo karoliquarti Chexiya va Germaniyaning Quyi Miosen (taxminan 13–23 mya) dan va Chamaeleo intermedius Keniyaning yuqori miosen (taxminan 5–13 mya) dan.[27]
Xameleyonlar, ehtimol, bundan ancha kattaroqdir, ehtimol ular umumiy ajdodlari bilan bo'lishishadi Iguanidlar va agamidalar 100 mya dan ortiq (agamidalar bir-biriga yaqinroq). Qoldiqlar Afrikada, Evropada va Osiyoda topilganligi sababli, xameleyonlar bugungi kunga qaraganda ancha keng tarqalgan.
Garchi bugungi kunda xameleyonlarning deyarli yarmi Madagaskarda yashasa ham, bu xameleyonlarning u erdan kelib chiqishi mumkinligi haqida taxmin qilish uchun hech qanday asos yo'q.[28] Darhaqiqat, yaqinda xameleyonlar Afrikaning materik qismida paydo bo'lganligi isbotlangan.[14] Ma'lum bo'lishicha, materikdan Madagaskarga ikki xil okean ko'chishi bo'lgan. Xameleonlarning xilma-xil spetsifikatsiyasi Oligotsen davri bilan birga bo'lgan ochiq yashash joylarining (savanna, o'tloq va hetland) ko'payishini to'g'ridan-to'g'ri aks ettirganligi haqida nazariya qilingan. Monofil oilani bir nechta tadqiqotlar qo'llab-quvvatlaydi.[29]
Daza va boshq. (2016), ehtimol, kichkina (burun teshigining uzunligi 10,6 mm) tasvirlangan yangi tug'ilgan chaqaloq bo'r davrida saqlanib qolgan kaltakesak (Albian -Senomiyalik chegara) kehribar Myanma. Mualliflarning ta'kidlashicha, kaltakesak "bosh suyagining kalta va kengligi, katta orbitalari, cho'zilgan va mustahkam til jarayoni, parallel chekkalari bilan frontal, boshlang'ich prefrontal boshlig'i, gijjalar kamaygan, retroartikulyar jarayon yo'q, prekakral vertebra soni past (15 dan 17 gacha) va juda yuqori kalta, bukilgan dum »; mualliflar bu xususiyatlarni kaltakesakning Chamaeleonidae bilan bog'lanishidan dalolat beradi deb hisoblashgan. Mualliflar tomonidan o'tkazilgan filogenetik tahlillar shuni ko'rsatdiki, kaltakesak a ildiz -xamaeleonid.[30] Biroq, Matsumoto & Evans (2018) ushbu namunani qayta izohladi albanerpetontid amfibiya.[31]
Xameleonlarda rang o'zgarishining aniq evolyutsion tarixi hali ham noma'lum bo'lsa-da, xameleyon rangining o'zgarishi tarixining allaqachon aniq o'rganilgan bir tomoni bor: signal samaradorligining ta'siri. Signalning samaradorligi yoki uning fonida signalni qanchalik yaxshi ko'rish mumkinligi to'g'ridan-to'g'ri xameleon displeylarining spektral fazilatlari bilan bog'liqligi ko'rsatilgan.[32] O'rganish xameleoni bo'lgan mitti xameleyonlar o'rmonlardan o'tloqgacha va butazorgacha turli xil yashash joylarini egallaydi. Yorug'roq joylardagi xameleonlar yorqinroq signallarni, ammo qorong'i joylardagi xameleonlar o'zlarining kelib chiqishiga nisbatan qarama-qarshi signallarni taqdim etishga moyil ekanliklari namoyish etildi. Ushbu topilma signal samaradorligi (va shu bilan yashash joyi) xameleon signalizatsiyasi evolyutsiyasiga ta'sir qilganligini ko'rsatadi. Styuart-Foks va boshq. kripsis uchun tanlov signal samaradorligi uchun tanlov kabi muhim ahamiyatga ega emasligi mantiqiy ekanligiga e'tibor bering, chunki signallar faqat qisqacha ko'rsatiladi; xameleonlar har doim tovushsiz, sirli ranglarda bo'ladi.[32]
Tavsif



Xameleyonlar kattaligi va tana tuzilishi jihatidan juda katta farq qiladi, maksimal uzunligi erkaklarda 15 mm (0,59 dyuym) gacha. Bruksiya mikrasi (dunyodan biri eng kichik sudralib yuruvchilar ) erkakda 68,5 sm (27,0 dyuym) gacha Furcifer oustaleti.[33] Ko'pchilik bosh yoki yuz bezaklariga ega, masalan burun chiqindilari yoki shoxga o'xshash proektsiyalar Trioceros jacksonii yoki shunga o'xshash boshlarning tepasida katta tepaliklar Chamaeleo kaliptratusi. Ko'p turlari mavjud jinsiy dimorfik va erkaklar odatda ayol xameleyonlarga qaraganda ancha bezatilgan.
Odatda asirda yoki uy hayvonlari sifatida saqlanadigan xameleyon turlarining odatiy o'lchamlari quyidagicha:
| Ilmiy nomi | Umumiy ism | Uzunlik (erkak) | Uzunlik (ayol) | Rang | Umr (yillar) |
|---|---|---|---|---|---|
| Chamaeleo kaliptratusi | Yopiq xameleyon | 35-60 sm | 25-33 sm | Yashil va och ranglar | taxminan 5 |
| Trioceros jacksonii | Jeksonning xameleyoni | 23-33 sm | 25-33 sm | Yashil va och ranglar | 5–10 |
| Furcifer pardalis | Panter xameleon | 38-53 sm | 23-33 sm | To'q ranglar | taxminan 5 (tug'ish ayollari uchun 2-3) |
| Rieppeleon brevicaudatus | Soqolli pigmentli xameleyon | 5-8 sm | 5-8 sm | Jigarrang, bej, yashil | taxminan 3-5 |
| Remfoleon spektri | Spektral pigmentli xameleyon | 8-10 sm | 5-10 sm | Sarg'ish va kulrang | 3–5 |
| Remfoleon vaqtinchalik | Usambara pigme xameleonini qoqdi | 6-10 sm | 5-9 sm | Kulrang va jigarrang | 5–11 |
Xameleonlarning oyoqlari juda moslashgan daraxtlar harakati va shunga o'xshash turlari Chamaeleo namaquensis er usti odatini ikkinchidan qabul qilganlar, ozgina o'zgarishsiz bir xil oyoq morfologiyasini saqlab qolishdi. Har bir oyoqda beshta aniq ajralib turadigan barmoqlar ikkita hayratga birlashtirilgan. Har bir moziydagi barmoqlar ikki yoki uch kishidan iborat tekislangan guruhga bog'lanib, har bir oyoqqa a beradi qisqich - tashqi ko'rinish kabi. Old oyoqlarda, tashqi, lateral, guruhda ikkita barmoq bor, ichki qismida, medial, guruhda uchta mavjud. Orqa oyoqlarda bu tartibga solish teskari bo'lib, medial guruh ikki barmoqni va lateral guruhni uchta o'z ichiga oladi. Ushbu ixtisoslashgan oyoqlar xameleyonlarni tor yoki qo'pol novdalar ustiga mahkam ushlashga imkon beradi. Bundan tashqari, toqqa chiqishda po'stlog'i kabi sirtlarni ushlab turish uchun har bir oyoq barmog'i o'tkir tirnoq bilan jihozlangan. Xameleyonlarning oyoqlariga murojaat qilish odatiy holdir didaktil yoki zigodaktil garchi ikkala atama ham to'liq qoniqtirmasa ham, ikkalasi ham umuman boshqa oyoqlarni tasvirlashda ishlatiladi, masalan, to'tiqushlarning zigodaktil oyoqlari yoki yalqovlar yoki tuyaqushlarning didaktil oyoqlari, ularning hech biri xameleon oyoqlariga o'xshamaydi. "Zigodaktil" xameleon oyoqlari anatomiyasini tavsiflovchi xususiyatga ega bo'lsa-da, ularning oyoq tuzilishi bu atama birinchi marta qo'llanilgan to'tiqushnikiga o'xshamaydi. Didaktikaga kelsak, xameleyonlar har oyog'ida ikkita emas, beshta barmoqqa ega.
Ba'zi xameleyonlarda umurtqa pog'onasi bo'ylab cho'zilgan mayda boshoqchalar tepasi bor proksimal quyruqning bo'yniga qismi; boshoqlarning kattaligi ham, kattaligi ham turlar va individlar orasida turlicha. Ushbu boshoqlar xameleyonning aniq tasavvurlarini buzishga yordam beradi, bu esa orqa fonga o'tishga yordam beradi.
Sezgilar
Xameleyonlar har qanday sudralib yuruvchilarning eng ko'ziga ega. Yuqori va pastki ko'z qovoqlari birlashtirilib, faqat ko'z qorachig'i ko'rishi uchun etarlicha teshikcha mavjud. Har bir ko'z mustaqil ravishda burilish va diqqatni jamlashi mumkin, bu esa xameleyonga bir vaqtning o'zida ikki xil ob'ektni kuzatishga imkon beradi. Bu ularga tanalari atrofida 360 daraja ko'rish qobiliyatini to'liq beradi. Yirtqich hayvon monokulyar yordamida joylashgan chuqurlik hissi, emas stereopsis.[34] Xameleyonlar sudralib yuruvchilar uchun juda yaxshi ko'rish qobiliyatiga ega bo'lib, ularga 5-10 metr masofadan kichik hasharotlarni ko'rishga imkon beradi.[iqtibos kerak ] Darhaqiqat, xameleonlar har qanday umurtqali hayvonlar orasida eng katta kattalashtirishga (har bir o'lchovga) ega.[35]
Yoqdi ilonlar, xameleyonlarning tashqi yoki o'rtasi yo'q quloq, shuning uchun na quloq ochiladi, na quloq pardasi bor. Biroq, xameleyonlar kar emas: ular 200-600 Hz oralig'idagi tovush chastotalarini aniqlay olishadi.[36]
Xameleyonlar ham ko'rinadigan, ham ko'rishlari mumkin ultrabinafsha nur.[37] Ultraviyole nurga uchragan xameleonlar ijtimoiy xulq-atvor va faollik darajasining oshganligini ko'rsatadi, ko'proq ovqatlanishga moyil bo'ladi va ko'payish ehtimoli ko'proq, chunki bu ijobiy ta'sir ko'rsatadi epifiz bezi.
Oziqlantirish
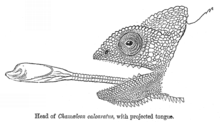




Barcha xameleyonlar birinchi navbatda hasharotlar tomonidan oziqlanadigan ballistik jihatdan loyihalash ularning uzun tillar uzoqroqda joylashgan o'ljani qo'lga kiritish uchun ularning og'zidan.[38] Xameleonlarning tillari odatda tanasining uzunligidan bir yarim-ikki baravar ko'p deb o'ylansa (ularning uzunligi dumidan tashqari), yaqinda xameleyonlar (ikkala kichik turlar va bir xil turdagi mayda shaxslar) yaqinda topilgan mutanosib ravishda katta til apparatlari, ularning katta o'xshashlariga qaraganda.[39] Shunday qilib, kichikroq xameleyonlar ko'pgina tadqiqotlar va tillarning uzunligini taxmin qilishning predmeti bo'lgan katta xameleyonlarga qaraganda o'z tillarini uzoqroq masofada proektsiyalashga qodir va tillarini tana uzunligidan ikki baravar ko'proq proektsiyalashi mumkin.[40]
Xameleyonning til apparati yuqori darajada o'zgartirilganlardan iborat hyoid suyaklar, til mushaklari va kollagen elementlar.[41][42][39][43] Giyoid suyagi uzun bo'yli, parallel qirrali proektsiyaga ega bo'lib, entoglossal jarayon deb ataladi, uning ustida quvurli mushak, tezlashtiruvchi mushak o'tiradi.[39][43][41][42] Tezlashtiruvchi mushak entoglossal jarayon atrofida qisqaradi va entoglossal jarayon bilan tezlashtiruvchi mushak o'rtasida joylashgan kollagen elementlarning yuklanishi orqali ham to'g'ridan-to'g'ri, ham til proektsiyasini kuchaytirish uchun ishni yaratishga mas'uldir.[38][39][41][42] Tilni tortib oluvchi mushak, gigogloss, gyoid va tezlashtiruvchi mushakni bir-biriga bog'lab turadi va til proektsiyasidan so'ng tilni og'ziga qaytarish uchun javobgardir.[38][39][43][41]
Tilni proektsiyalash o'ta yuqori ko'rsatkichlarda sodir bo'ladi, o'ljaga 0,07 soniya ichida yetib boradi,[41][42][44]41 dan yuqori tezlikda ishga tushirildi g.[44] The kuch 3000 Vt dan oshishi ma'lum bo'lgan til ishga tushirildi−1, mushak ishlab chiqarishga qodir bo'lgan narsadan oshib ketadi, bu esa tilni proektsiyalash uchun elastik quvvat kuchaytirgichi mavjudligini ko'rsatadi.[42] Shunday qilib, til apparatlaridagi elastik elementlarning orqaga tortilishi umumiy til proektsiyasining katta foizlari uchun javobgardir.
Tilni proektsiyalash mexanizmiga elastik orqaga tortish mexanizmini qo'shilishining natijalaridan biri bu tilni tortib olishga nisbatan til proektsiyasining nisbiy termal befarqligi bo'lib, u faqat mushaklarning qisqarishi bilan quvvatlanadi va juda termik sezgir bo'ladi.[44][45] Boshqasi esa ektotermik hayvonlar tana haroratining pasayishi bilan sustlashadi, mushaklarining qisqarish tezligining pasayishi tufayli xameleyonlar tana haroratini past bo'lgan taqdirda ham o'z tillarini yuqori darajada proektsiyalashga qodir.[44][45] Xameleonlarda tilni tortib olishning termal sezgirligi muammo emas, chunki xameleyonlar til bilan aloqa qilgandan keyin ularning o'ljasini ushlab turish mexanizmining juda samarali mexanizmiga ega, shu jumladan ho'l yopishish va bir-biriga yopishish kabi sirt hodisalari. assimilyatsiya.[46] Shunday qilib, til proektsiyasining termal befarqligi xameleyonlarning tana haroratini xatti-harakatlari bilan ko'tarishidan oldin sovuq ertalab samarali ovqatlanishiga imkon beradi. termoregulyatsiya, qachon boshqasi simpatik kaltakesaklar turlari hanuzgacha harakatsiz, ehtimol vaqtincha ularning termal kengayishi joy Natijada.[44]
Suyaklar
Xameleonlarning ayrim turlarida suyaklari bor, ular ostida porlaydilar ultrabinafsha nur, shuningdek, nomi bilan tanilgan biogen lyuminestsentsiya.[47] Ba'zi 31 xil turlari Kalumma xameleonlar, barchasi mahalliy Madagaskar, ushbu lyuminestsentsiyani namoyish etdi KT tekshiruvi.[48] Suyaklar yorqin moviy nur sochdi va xameleyonning to'rtta terisi orqali porlashi ham mumkin edi.[48] Yuzning boshqa porlashi aniqlandi, aks holda ular nuqta sifatida tanilgan sil kasalligi yuz suyaklarida.[47] Yorqinlik paydo bo'ladi oqsillar, pigmentlar, xitin va xameleyon skeletini tashkil etuvchi boshqa materiallar,[47] ehtimol xameleyonlarga rangni o'zgartirish qobiliyatiga xalaqit bermaydigan ikkilamchi signalizatsiya tizimini berish va jinsiy tanlov.[47]
Tarqatish va yashash muhiti

Xameleyonlar asosan materikda yashaydilar Saxaradan Afrikaga va Madagaskar orolida, garchi bir nechta turlari yashaydi shimoliy Afrika, janubiy Evropa (Ispaniya, Italiya, Gretsiya), Yaqin Sharq, Janubiy Hindiston, Shri-Lanka, va bir nechta kichik orollar g'arbiy Hind okeani. Yirtqich populyatsiyalar mavjud pardali va Jeksonniki xameleyonlar Gavayi, va yakson Jekson xameleonlarining ajratilgan cho'ntaklari haqida xabar berilgan Kaliforniya, Florida va Texas.
Xameleyonlar barcha turlarda yashaydi tropik va tog ' yomg'ir o'rmonlari, savannalar va ba'zan cho'llar va dashtlar. Chamaeleoninae oilasidan odatdagi xameleyonlar daraxt, odatda daraxtlarda yoki butalarda yashaydi, garchi bir nechtasi (xususan Namaqua xameleyoni ) qisman yoki katta darajada quruqlik. Brookesiinae subfamilyasidan ko'p turlar, ular nasabga kiradi Bruksiya, Rieppeleon va Remfoleon, o'simlik ostida kam yoki tuproqda yashaydi barg axlati. Xameleonlarning ko'plab turlari yo'qolib ketish xavfi ostida. Xameleon sonining kamayishi yashash joylarining yo'qolishi bilan bog'liq.[49]
Ko'paytirish


Xameleyonlar asosan tuxumdon, ba'zi birlari bilan ovoviviparous.
Tuxumdon turlari uch-olti hafta o'tgach tuxum qo'yadi ko'paytirish. 10-30 sm dan (4-12 dyuymgacha), turiga qarab chuqur qazib, tuxum qo'yadi va tuxum qo'yadi. Debriyajning o'lchamlari turlarga qarab juda farq qiladi. Kichik Bruksiya turlari faqat ikki-to'rt tuxum qo'yishi mumkin, katta bo'lsa pardali xameleyonlar (Chamaeleo kaliptratusi) 20–200 (pardali xameleonlar) va 10–40 (pantera xameleonlar) tuxumlarini debriyaj qilishlari ma'lum bo'lgan. Debriyajning o'lchamlari bir xil turlar orasida juda katta farq qilishi mumkin. Tuxumlar, odatda, turlarga qarab, to'rt oydan 12 oygacha tug'iladi. Ning tuxumlari Parson xameleyoni (Calumma parsonii), asirlikda kamdan-kam uchraydigan turlarning paydo bo'lishiga 24 oydan ko'proq vaqt ketadi deb ishoniladi.[50]
Xameleonlar moslashuvchan qobiqli tuxum qo'yadi, ular inkubatsiya paytida atrof-muhit xususiyatlariga ta'sir qiladi. Tuxum massasi inkubatsiya paytida Xameleyondan omon qolganlarni farqlashda eng muhim hisoblanadi. Tuxum massasining ko'payishi harorat va suv salohiyatiga bog'liq bo'ladi.[51] Xameleyon tuxumidagi suv potentsialining dinamikasini tushunish uchun tuxum qobig'iga bosimni hisobga olish juda muhimdir, chunki butun inkubatsiya davrida tuxum qobig'ining bosimi suv bilan bog'liqlikda muhim rol o'ynaydi. [52]
The ovoviviparous kabi turlar, masalan Jeksonning xameleyoni (Trioceros jacksonii) beshdan etti oygacha homiladorlik davriga ega. Har bir yosh xameleyon uning sarig'i sumkasining yopishqoq shaffof membranasida tug'iladi. Ona har bir tuxumni novdaga bosadi, u erda u yopishadi. Membrana yorilib, yangi chiqqan xameleyon o'zini ozod qiladi va o'zini ov qilish va yirtqichlardan yashirish uchun ko'tariladi. Ayol bitta homiladorlikdan 30 taga qadar yosh bolaga ega bo'lishi mumkin.[53]
Parhez
Xameleyonlar odatda ovqatlanadilar hasharotlar, ammo oddiy xameleyon kabi kattaroq turlar boshqa kaltakesaklar va yoshlarni ham olishi mumkin qushlar.[54]:5 Parhez turlarini quyidagi misollardan ko'rish mumkin:
- Pardali xameleyon, Chamaeleo kaliptratusi dan Arabiston, hasharotlidir, ammo boshqa suv manbalari mavjud bo'lmaganda barglarni iste'mol qiladi. Bu dietada saqlanishi mumkin kriketlar.[55] Ular kuniga 15-50 tagacha katta kriket yeyishlari mumkin.
- Jeksonning xameleyoni (Trioceros jacksonii ) Keniyadan va Tanzaniyaning shimoliy qismidan chumolilar, kapalaklar, tırtıllar, salyangozlar, qurtlar, kaltakesaklar, gekkoslar, amfibiyalar va boshqa xameleonlar, shuningdek barglar, mayin kurtaklar va mevalar kabi o'simlik moddalarini iste'mol qiladi. Uni karam, karahindiba barglari, marul, banan, pomidor, olma, kriket va mum qurtlari kabi aralash dietada saqlash mumkin.[53]
- Evropa, Shimoliy Afrika va Yaqin Sharqning umumiy xameleyoni, Chamaeleo xamaeleon, asosan yeydi ari va mantis; shunday artropodlar uning dietasining to'rtdan uch qismini tashkil qiladi.[54]:5 Ba'zi ekspertlar oddiy xameleyonni faqat kriket bilan boqmaslik kerakligini maslahat berishadi; bu dietaning yarmidan ko'pini tashkil qilishi kerak, qolgan qismi esa aralashtiriladi mum qurtlari, yomg'ir qurtlari, chigirtkalar, chivinlar va yashil barglar, jo'xori va mevalar kabi o'simlik materiallari.[54]:5–6
- Harorat iste'mol qilingan ovqat miqdoriga ta'sir qiladi.
- Ba'zi xameleyonlar shunga o'xshash panter xameleon ning Madagaskar ularni tartibga solish D3 vitamini o'zlarining ta'siriga tushib, hasharotlarning parhezi kambag'al manba bo'lgan darajalar quyosh nuri undan beri UV nurlari komponent ichki ishlab chiqarishni ko'paytiradi.[56]
Yirtqichlik va mudofaa

Xameleyonlar boshqa hayvonlarga o'lja bo'lib, iste'mol qilinadi. Qushlar va ilonlar eng muhimini anglatadi yirtqichlar kattalar xameleonlari va umurtqasiz hayvonlar, ayniqsa chumolilar, xameleon tuxumlari va balog'atga etmagan bolalarga yuqori yirtqich bosim o'tkazmoqda.[57] Xameleyonlarning yirtqichlardan qochib qutulishi va ularga ishonishi ehtimoldan yiroq emas kripsis ularning asosiy himoyasi sifatida.[58] Xameleyonlar o'zlarining ranglarini ham, naqshlarini ham o'zgarishi mumkin (har xil darajada) atrofdagilarga o'xshab ketishi yoki tana chizig'ini buzishi va potentsial dushmanning ko'zidan yashirin qolishi mumkin. Faqatgina aniqlangan taqdirda, xameleyonlar faol ravishda o'zini himoya qiladi. Ular mudofaa holatini egallaydilar, tajovuzkorga kattaroq ko'rinish uchun yon tomondan yassilangan tanani namoyish qiladilar, ochiq og'iz bilan ogohlantiradilar va agar kerak bo'lsa, kurashish uchun oyoq va jag'lardan foydalanadilar.[59] Ba'zan vokalizatsiya tahdid ekraniga qo'shiladi.[57]
Parazitlar
Xameleyonlar parazitlangan tomonidan nematod qurtlar, shu jumladan ip qurtlari (Filarioidea ). Ip qurtlari kabi hasharotlarni tishlash orqali yuqishi mumkin Shomil va chivinlar. Boshqa yumaloq qurtlar yumaloq qurt tuxumlari bilan ifloslangan oziq-ovqat orqali yuqadi; lichinkalari devoridan o'tib ketadi ichak qon oqimiga.[60]
Xameleyonlar bir nechta protozoy parazitlariga, masalan Plazmodium sabab bo'ladi bezgak, Tripanozoma sabab bo'ladi uyqu kasalligi va Leyshmaniya sabab bo'ladi leyshmanioz.[61]
Xameleyonlar tomonidan parazitizmga uchragan koksidiya,[61] shu jumladan nasl turlari Xoleoimeriya, Eimeria va Isospora.[62]
Adabiyotlar
- ^ a b Glaw, F. (2015). "Xameleyonlarning taksonomik tekshiruvi ro'yxati (Squamata: Chamaeleonidae)". Umurtqali hayvonlar zoologiyasi. 65 (2): 167–246.
- ^ Edmonds, Patrisiya (2015 yil sentyabr). "Haqiqiy ranglar". National Geographic: 98.
- ^ xameleyon. Charlton T. Lyuis va Charlz Short. Lotin lug'ati kuni Perseus loyihasi.
- ^ gámias. Liddel, Genri Jorj; Skott, Robert; Yunoncha-inglizcha leksikon da Perseus loyihasi.
- ^ gámáb yilda Liddel va Skott.
- ^ λέων yilda Liddel va Skott.
- ^ "Xameleyon". Dictionary.com.
- ^ Xarper, Duglas. "xameleyon". Onlayn etimologiya lug'ati.
- ^ Klaver, C .; Böhme, W. (1986). "Filippeniya va Chamaeleonidae (Sauria) klassifikatsiyasi, gemipenis morfologiyasiga alohida murojaat qilish bilan". Bonner Zoologische Monographien. 22: 1–64.
- ^ a b Tilberi, Kolin (2010). Afrika xameleonlari, Evropa, Yaqin Sharq va Osiyo xameleonlarini o'z ichiga olgan Atlas. Frankfurt: Chimaira nashri. ISBN 978-3899734515.
- ^ Taunsend, T .; Larson, A. (2002). "Chamaeleonidae (Reptilia, Squamata) da molekulyar filogenetik va mitoxondriyal genomik evolyutsiya". Molekulyar filogenetik va evolyutsiyasi. 23 (1): 22–36. doi:10.1006 / mpev.2001.1076. PMID 12182400.
- ^ Raxvorti, C. J .; Forstner, M. R. J .; Nussbaum, R. A. (2002). "Okean tarqalishi bilan xameleyon nurlanishi" (PDF). Tabiat. 415 (6873): 784–787. Bibcode:2002 yil natur.415..784R. doi:10.1038 / 415784a. hdl:2027.42/62614. PMID 11845207. S2CID 4422153.
- ^ Taunsend, T. M .; Tolley, K. A .; Glav, F .; Bohme, V.; Vents, M. (2011). "Afrikadan sharqqa qarab: Seyshel orollariga tarqaladigan paleourrent vositasida xameleyon tarqalishi". Biologik xatlar. 7 (2): 225–228. doi:10.1098 / rsbl.2010.0701. PMC 3061160. PMID 20826471.
- ^ a b Tolley, K. A .; Taunsend, T. M .; Vents, M. (2013). "Xameleyonlarning katta miqyosdagi filogeniyasi afrikalik kelib chiqishi va evosenning xilma-xilligini anglatadi". Qirollik jamiyati materiallari B. 280 (1759): 20130184. doi:10.1098 / rspb.2013.0184. PMC 3619509. PMID 23536596.
- ^ Tilberi, Kolin (2014). "Chamaeleonidae sistematikasiga umumiy nuqtai". Tolleyda, Kristal A.; Herrel, Entoni (tahrir). Xameleonlar biologiyasi. Berkli: Kaliforniya universiteti matbuoti. 151–174 betlar. ISBN 9780520276055.
- ^ Xameleyonlar. National Geographic Explorer
- ^ Styuart-Foks, D.; Moussalli, A. (2008). "Ijtimoiy signalizatsiya uchun tanlov xameleyon rangining o'zgarishini keltirib chiqaradi". PLOS biologiyasi. 6 (1): e25. doi:10.1371 / journal.pbio.0060025. PMC 2214820. PMID 18232740.
- ^ Xarris, Tom. "Hayvonlarning kamuflyaji qanday ishlaydi". Qanday narsalar ishlaydi. Olingan 2006-11-13.
- ^ Kuk, Mariya. "Xameleyonlarning moslashuvi". Ilm-fan. Olingan 15 iyun 2020.
- ^ Ligon, Rassel A.; McGraw, Kevin J. (2013). "Xameleyonlar musobaqalar paytida ranglarning murakkab o'zgarishi bilan aloqa qilishadi: tananing turli mintaqalari turli xil ma'lumotlarni etkazishadi". Biologiya xatlari. 9 (6): 20130892. doi:10.1098 / rsbl.2013.0892. PMC 3871380. PMID 24335271.
- ^ Ligon, Rassell A (2014). "Mag'lubiyatga uchragan xameleonlar dominantlar xavfini kamaytirish uchun dyadik nizolar paytida dinamik ravishda qorayadi" Xulq-atvor ekologiyasi va sotsiobiologiyasi. 68 (6): 1007–1017. doi:10.1007 / s00265-014-1713-z. S2CID 18606633.
- ^ Devid Protsel, Martin Xess, Mark D. Sherz, Martina Shvager, Anuk van’t Padje, Frank Glav (2018). "Xameleonlarda suyakka asoslangan keng tarqalgan lyuminestsentsiya". Ilmiy ma'ruzalar. 8 (698): 698. Bibcode:2018 yil NatSR ... 8..698P. doi:10.1038 / s41598-017-19070-7. PMC 5768862. PMID 29335580.CS1 maint: mualliflar parametridan foydalanadi (havola)
- ^ Yosh, Emma (2008) Xameleyonlar kamuflyajni yirtqichlarning ko'rish qobiliyatiga moslashtiradi. Yangi olim
- ^ Taysier, Jeremie; Saenko, Suzanna V.; van der Marel, Dirk; Milinkovich, Mishel C. (2015). "Fotonik kristallar xameleyonlarda faol rang o'zgarishini keltirib chiqaradi." Tabiat aloqalari (6).
- ^ National Geographic (2015) Xameleyonlarning rangli tili
- ^ Styuart-Foks, Devi; Moussalli, Adnan (2008-01-29). "Ijtimoiy signalizatsiya uchun tanlov xameleyon rangining o'zgarishini keltirib chiqaradi". PLOS biologiyasi. 6 (1): e25. doi:10.1371 / journal.pbio.0060025. ISSN 1545-7885. PMC 2214820. PMID 18232740.
- ^ a b Maisano, Jessie (2003 yil 27 avgust). "Chamaeleo calyptratus, pardali xameleyon". Digimorf. Ostindagi Texas universiteti. Olingan 10 yanvar, 2012.
- ^ Tolli, Kristal; Burger, Marius (2007). Janubiy Afrikaning xameleyonlari. Struik. 26-28 betlar. ISBN 978-1-77007-375-3.
- ^ Tolley, Kristal A.; Herrel, Entoni (2013 yil 16-noyabr). Xameleonlar biologiyasi. California Press universiteti. ISBN 9780520276055. Olingan 1 noyabr 2017 - Google Books orqali.
- ^ Daza, Xuan D.; Stenli, Edvard L.; Vagner, Filipp; Bauer, Aaron M.; Grimaldi, Devid A. (2016). "O'rta bo'r davridagi kehribar tosh qoldiqlari tropik kaltakesaklarning o'tmishdagi xilma-xilligini yoritmoqda". Ilmiy yutuqlar. 2 (3): e1501080. Bibcode:2016SciA .... 2E1080D. doi:10.1126 / sciadv.1501080. PMC 4783129. PMID 26973870.
- ^ Ryoko Matsumoto; Syuzan E. Evans (2018). "Sharqiy Osiyodan kelgan albanerpetontid amfibiyalarining birinchi yozuvi (Amfibiya: Albanerpetontidae)". PLOS ONE. 13 (1): e0189767. Bibcode:2018PLoSO..1389767M. doi:10.1371 / journal.pone.0189767. PMC 5752013. PMID 29298317.
- ^ a b Styuart-Foks, D.; Mussalli, Adnan; Whiting, Martin J. (2007). "Ijtimoiy signallarda tabiiy selektsiya: signal samaradorligi va xameleyon displeyining ranglanishi evolyutsiyasi". Amerikalik tabiatshunos. 170 (6): 916–930. doi:10.1086/522835. PMID 18171173. S2CID 21716855.
- ^ Glav, Frank; Vens, Migel (1994). Madagaskar amfibiyalari va sudralib yuruvchilar uchun dala qo'llanmasi (2-nashr). Kyoln: Verlags GbR. ISBN 978-3-929449-01-3.
- ^ Ott, M.; Sheeffel, F.; Kirmse, W. (1998). "Binokulyar ko'rish va yirtqich xameleyonlarda yashash". Qiyosiy fiziologiya A. 182 (3): 319–330. doi:10.1007 / s003590050182. S2CID 19988312.
- ^ Ott, M. va F. Sheffel. (1995). Xameleyondagi salbiy quvvatli ob'ektiv. Tabiat 373: 692-694
- ^ Le Berre va Bartlett, p. 31
- ^ "Chamaeleon News, 2004 yil avgust". Chameleonnews.com. Asl nusxasidan arxivlandi 2008 yil 22 yanvar. Olingan 1 noyabr 2017.CS1 maint: BOT: original-url holati noma'lum (havola)
- ^ a b v Higham, T. E.; Anderson, C. V. (2014), "Xameleyonlarning funktsiyasi va moslashuvi", Tolleyda, K. A.; Herrel, A. (tahr.), Xameleonlar biologiyasi, Berkli, Kaliforniya: Kaliforniya universiteti matbuoti, 63-83 betlar
- ^ a b v d e Anderson, C. V.; Sheridan, T .; Deban, S. M. (2012). "Xameleyonlarda ballistik til apparatining masshtablanishi". Morfologiya jurnali. 273 (11): 1214–1226. doi:10.1002 / jmor.20053. PMID 22730103. S2CID 21033176.
- ^ Anderson, Kristofer V. (2009) Rhamfolon spinosusi videoni oziqlantirish. chamaeleonidae.com
- ^ a b v d e Herrel, A .; Meyers, J. J .; Nishikava, K. C .; De Vree, F. (2001). "Xameleonlardagi giologik til apparati morfologiyasi va gistokimyosi". Morfologiya jurnali. 249 (2): 154–170. doi:10.1002 / jmor.1047. PMID 11466743. S2CID 3246256.
- ^ a b v d e de Groot, J. H .; van Liuven, J. L. (2004). "Xameleyon tilidagi elastik proektsiya mexanizmi uchun dalillar". London Qirollik jamiyati materiallari B. 271 (1540): 761–770. doi:10.1098 / rspb.2003.2637. PMC 1691657. PMID 15209111.
- ^ a b v Anderson, C. V.; Higham, T. E. (2014), "Xameleyon anatomiyasi", Tolleyda, K. A.; Herrel, A. (tahr.), Xameleonlar biologiyasi, Berkli, Kaliforniya: Kaliforniya universiteti matbuoti, 7-55 betlar
- ^ a b v d e Anderson, C. V.; Deban, S. M. (2010). "Xameleonlardagi balistik til proektsiyasi past haroratda yuqori ko'rsatkichlarni saqlaydi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 107 (12): 5495–5499. Bibcode:2010PNAS..107.5495A. doi:10.1073 / pnas.0910778107. PMC 2851764. PMID 20212130.
- ^ a b Anderson, C. V.; Deban, S. M. (2012). "Dvigatelni boshqarishga issiqlik ta'siri va in vitro xameleonlardagi ballistik til apparatining mushak dinamikasi ". Eksperimental biologiya jurnali. 215 (24): 4345–4357. doi:10.1242 / jeb.078881. PMID 23125336.
- ^ Herrel, A .; Meyers, J. J .; Aerts, P .; Nishikava, K. C. (2000). "Xameleyonlarda o'lja prexanitsiyasining mexanikasi" (PDF). Eksperimental biologiya jurnali. 203 (Pt 21): 3255-33263. PMID 11023845.
- ^ a b v d Protsel, Dovud; Xess, Martin; Sherz, Mark D .; Shvager, Martina; Padje, Anouk yo'q; Glav, Frank (2018-01-15). "Xameleonlarda suyakka asoslangan keng tarqalgan lyuminestsentsiya". Ilmiy ma'ruzalar. 8 (1): 698. Bibcode:2018 yil NatSR ... 8..698P. doi:10.1038 / s41598-017-19070-7. ISSN 2045-2322. PMC 5768862. PMID 29335580.
- ^ a b Elaina Zachos (2018-01-18). "Xameleyon suyaklari zulmatda, hatto teri orqali ham porlaydi". National Geographic. Olingan 2018-08-03.
- ^ "Habitatning yo'qolishi va parchalanishi Tanzaniyada xameleon sonini kamaytiradi". Phys.org. Olingan 1 noyabr 2017.
- ^ Glav, Frank; Vens, Migel (1994). Madagaskar amfibiyalari va sudralib yuruvchilar uchun dala qo'llanmasi (2-nashr). Kyoln: Verlags GbR. ISBN 978-3-929449-01-3.
- ^ Diaz-Paniagua, Kuadrado, Inkubatsion sharoitlarning inkubatsiya muvaffaqiyatiga, embrionning rivojlanishiga va oddiy xameleyon tuxumlarining inkubatsiya fenotiplariga ta'siri., 429-440 betlar[to'liq iqtibos kerak ]
- ^ Endryus, Yopiq xameleyonning o'sishi va ishlashiga inkubatsiya haroratining ta'siri (Chemaeleo calyptratus), Journal of Experimental Zooly, 435–446 betlar[to'liq iqtibos kerak ]
- ^ a b "Afrika yomg'ir o'rmoni". Jeksonning xameleyoni. Toronto hayvonot bog'i. Arxivlandi asl nusxasi 2011 yil 11-noyabrda. Olingan 9 yanvar, 2012.
- ^ a b v Dever, Jennifer (2007 yil 5-dekabr). "Umumiy xameleyon" (PDF). usfca.edu. Arxivlandi asl nusxasi (PDF) 2015 yil 3 fevralda. Olingan 9 yanvar, 2012.
- ^ "Sudralib yuruvchilar va amfibiyalar: yopiq xameleyon". Smithsonian National Zoological Park. Arxivlandi asl nusxasi 2011-12-17. Olingan 9 yanvar, 2012.
- ^ Karsten, K. B.; Fergyuson G. V.; Chen T. C .; Holick M. F. (2009). "Panther xameleonlari, Furcifer pardalis, xun vitaminlari D3 holatiga qarab ultrabinafsha nurlar ta'siriga ta'sir qiladi". Fiziol. Biokimyo. Zool. 82 (3): 218–25. doi:10.1086/597525. PMID 19335229. S2CID 205990383.
- ^ a b Styuart-Foks, D (2014). Xameleyonning xatti-harakatlari va ranglarning o'zgarishi. K. A. Tolley va A. Herrel (Eds.), Xameleonlar biologiyasi (115-130-betlar). Berkli: Kaliforniya universiteti matbuoti.
- ^ Measey, G. J., Raselimanana, A., va Herrel, A. (2014). Xameleyonlarning ekologiyasi va hayot tarixi. K. A. Tolley va A. Herrel (Eds.), Xameleonlar biologiyasi (85–114-betlar). Berkli: Kaliforniya universiteti matbuoti.
- ^ Berg, Filipp; Berg, Jessica; Berg, Rayner (2020). "Boomslang, Dispholidus typus va flap-necked xameleon, Chamaeleo dilepis" o'rtasidagi yirtqich hayvonlarning o'zaro ta'siri. Afrika ekologiya jurnali. doi:10.1111 / aje.12782.
- ^ Le Berre va Bartlett, p. 110
- ^ a b Le Berre va Bartlett, p. 109
- ^ Sloboda, Mixal; Modri, Devid (2006). "Chameeleo calyptratus (Sauria: Chamaeleonidae), xameleonlardan emeriya koksidiyalarini taksonomik qayta ko'rib chiqish bilan yangi xoleoeimeria turlari (Apicompleksa: Eimeriidae)". Folia Parasitologica. 53 (2): 91–97. doi:10.14411 / fp.2006.012. PMID 16898122.
Bibliografiya
- Le Berre, Fransua; Bartlett, Richard D. (2009) Xameleyon uchun qo'llanma. Barronning ta'lim seriyalari. 3-nashr. ISBN 0764141422.
Qo'shimcha o'qish
- "Olimlar Madagaskar xameleyonini oxirgi marta 100 yil oldin ko'rgan". Associated Press. 30 oktyabr 2020 yil.
- Anderson, C. V.; Deban, S. M. (2010). "Xameleonlardagi balistik til proektsiyasi past haroratda yuqori ko'rsatkichlarni saqlaydi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 107 (12): 5495–5499. Bibcode:2010PNAS..107.5495A. doi:10.1073 / pnas.0910778107. PMC 2851764. PMID 20212130.
- Anderson, C. V.; Deban, S. M. (2012). "Dvigatelni boshqarishga issiqlik ta'siri va in vitro xameleonlardagi ballistik til apparatining mushak dinamikasi ". Eksperimental biologiya jurnali. 215 (24): 4345–4357. doi:10.1242 / jeb.078881. PMID 23125336.
- Anderson, C. V.; Sheridan, T .; Deban, S. M. (2012). "Xameleyonlarda ballistik til apparatining masshtablanishi". Morfologiya jurnali. 273 (11): 1214–1226. doi:10.1002 / jmor.20053. PMID 22730103. S2CID 21033176.
- Devison, Linda J. Xameleyonlar: ularni parvarish qilish va ko'paytirish. Hancock House Publishers, 1997 yil.
- de Groot, J. H .; van Liuen, J. L. (2004). "Xameleyon tilidagi elastik proektsion mexanizm uchun dalillar.". London Qirollik jamiyati materiallari B. 271 (1540): 761–770. doi:10.1098 / rspb.2003.2637. PMC 1691657. PMID 15209111.
- de Vosjoli, Filippe. Xameleyonlarga zaruriy yordam. Advanced Vivarium Systems, 2004 yil.
- Herrel, A .; Meyers, J. J .; Nishikava, K. C .; De Vree, F. (2001). "Xameleyonlardagi giologik til apparati morfologiyasi va gistoximyasi". Morfologiya jurnali. 249 (2): 154–170. doi:10.1002 / jmor.1047. PMID 11466743. S2CID 3246256.