Janubiy Osiyo genetikasi va arxeogenetikasi - Genetics and archaeogenetics of South Asia
Janubiy Osiyo genetikasi va arxeogenetikasi ning o'rganilishi genetika va arxeogenetika ning Janubiy Osiyodagi etnik guruhlar. Ushbu guruhlarni aniqlashga qaratilgan genetik tarix. Janubiy Osiyoning geografik holati uning biologik xilma-xilligini erta tarqalishini o'rganish uchun muhim ahamiyatga ega anatomik jihatdan zamonaviy odamlar bo'ylab Osiyo.
Mitokondriyal DNKga asoslangan tadqiqotlar (mtDNA Turli xil Janubiy Osiyoning pastki populyatsiyalari bo'yicha genetik birlik haqida xabar berilgan.[1][2][3][4] Y xromosomalari o'zgarishi va DNKning avtosomal o'zgarishi asosida olib borilgan tadqiqotlar natijalari har xil edi, ammo ko'plab tadqiqotchilar barcha mtDNA turlarining filogenetik daraxtining ajdod tugunlarining ko'pi Janubiy Osiyoda kelib chiqqan deb ta'kidlaydilar. Yaqinda o'tkazilgan genom tadqiqotlari shuni ko'rsatadiki, aksariyat janubiy osiyoliklar ikkita asosiy ajdodlarning avlodlari, ulardan biri Janubiy Osiyo bilan cheklangan (Janubiy Hindistonning ajdodlari, dan kelib chiqqan IVC - odamlar va uzoq Janubiy Osiyo aholisi Andamancha ) va boshqa komponent (Shimoliy Hindistonning ajdodlari) IVC odamlari va Dasht odamlaridan kelib chiqqan bo'lib, uni boshqalar bilan yanada yaqinroq qilish Markaziy Osiyo, G'arbiy Osiyo va Evropa.[5][6][7] 2016 yilgi tadqiqot Hindiston materikida yana ikkita ajdod tarkibiy qismini aniqladi AAA va ATBAvstriya-Osiyo tillarida so'zlashadigan qabilalar va Tibet-Burman ma'ruzachilari uchun bu muhim. Tadqiqot shuningdek, aholisi Andaman orollari arxipelag aniq ajdodlarni shakllantiradi, "bu koansestral deb topilgan Okean populyatsiyalar ".[8] Genomik tadqiqotlar Janubiy Osiyoning genetik landshaftini G'arbiy Evroosiyo va Sharqiy Osiyo zamonaviy Janubiy Osiyoliklarni yaratish uchun mahalliy Janubiy Osiyo guruhlari bilan aralashgan ekzogen komponentlar.[9] Hindistonda aniqlangan Sharqiy Osiyo ajdodlari komponenti asosan Himoloy tog 'etaklaridagi va Hindistonning shimoliy-sharqiy qismidagi ma'lum populyatsiyalar bilan cheklangan.[10][11]
Barcha filogenetik daraxtning ajdodlar tuguni ekanligi aniqlandi mtDNA turlari (mitoxondriyal DNK haplogrouplari ) odatda Markaziy Osiyoda, G'arbiy Osiyoda va Evropada Janubiy Osiyoda nisbatan yuqori chastotalarda uchraydi. Ushbu umumiy ajdodlar tugunining taxmin qilingan farqi 50 ming yilga nisbatan bir oz kamroq sodir bo'lgan deb taxmin qilinadi.[12] Hindistonda onaning asosiy nasllari har xil M subkladlar, so'ngra R va U pastki chiziqlar. Ushbu mitoxondriyal haplogrouplarning birlashish vaqtlari bugungi kunga kelib taxminan 50,000 BP ga tenglashtirilgan.[12]
Tomonidan ifodalangan asosiy ota avlodlari Y xromosomalari haplogrouplar R1a1, R2, H, L va Haplogroup J2.[13] Ba'zi tadqiqotchilar Y-DNK Haplogroup R1a1 (M17) ning ekanligini ta'kidladilar avtonom Janubiy Osiyo kelib chiqishi.[14][15] Biroq, R1a1 uchun Markaziy Osiyo kelib chiqishi bo'yicha takliflar ham juda keng tarqalgan.[16][17]
Umumiy nuqtai
Afrikadan tashqaridagi barcha mtDNA va Y-xromosoma nasablari uchta asoschidan kelib chiqadi:
Ushbu oltita asoschining haplogrouplarini hozirgi kungi populyatsiyada topish mumkin Janubiy Osiyo. Bundan tashqari, mtDNA haplogroup M va Y-xromosoma haplogrouplar C va D bilan cheklangan sharq Janubiy Osiyo. Butun G'arb Evroosiyo populyatsiyalar mtDNA ning N va R gaplogruplaridan va Y-xromosomaning F gaplogruplaridan kelib chiqadi.[18]
Endikot va boshq. ushbu dalillar 65000 yil oldin a orqali Sharqiy Afrikadan bir marta ko'chib ketish gipotezasiga mos kelishini ta'kidlang janubiy qirg'oq yo'li, G'arbiy Evroosiyo nasablari Janubiy Osiyo nasablaridan Sharqiy / Shimoliy-Sharqiy Afrika va Janubiy Osiyo o'rtasida ajralib turadi.[19]
Janubiy osiyoliklarning aksariyat genom belgilarining barchasi chambarchas bog'liqdir G'arbiy Evroosiyoliklar va kelib chiqishi ham bo'lishi mumkin G'arbiy Osiyo yoki Janubiy Osiyo o'zi.[20]
mtDNA

Eng tez-tez mtDNA haplogrouplari Janubiy Osiyoda M, R va U (bu erda U R ning avlodi).[13]
Uzoq muddatli "raqib Y-xromosoma modeli" uchun bahslashib,[14] Stiven Oppengeymer bu Hindistonning kelib chiqishi ekanligi juda muhimdir Evroosiyo mtDNA u "Evroosiyo Evlari" deb ataydigan haplogrouplar. Oppengeymerning fikriga ko'ra, Markaziy Osiyoda, Yaqin Sharqda va Evropada odamlarning deyarli barcha nasl-nasablari Janubiy Osiyoda 50,000-100,000 yil oldin paydo bo'lgan atigi to'rtta mtDNA liniyalaridan kelib chiqqan.[21]
Macrohaplogroup M
The makrohaplogroup M proto-Osiyo nasablarining klasteri sifatida qaraladigan,[12] Janubiy Osiyo MtDNA ning 60% dan ortig'ini tashkil etadi.[22]
Hindistondagi M makrohaplotipi tarkibiga Sharqiy Osiyodagi boshqa sublinajlardan, xususan mongoloid populyatsiyalaridan tubdan farq qiluvchi ko'plab kichik guruhlar kiradi.[12] M filogenezining chuqur ildizlari, Janubiy Osiyodagi Janubiy Osiyodagi nasl-nasablarni boshqa Sharqiy Osiyodagi (Sharqiy Osiyoda va boshqa joylarda) taqqoslaganda, aniqrog'i Janubiy Osiyodagi ushbu sub-haplogruplarning kelib chiqishi "in-situ" kelib chiqishini taxmin qilmoqda. Ushbu chuqur ildiz otish nasablari tilga xos emas va Hindistondagi barcha til guruhlariga tarqalgan.[22]
Deyarli barcha zamonaviy Markaziy Osiyo MtDNA M nasablari Sharqiy Evroosiyoga tegishli (Mo'g'ul ) M haplogroupining Janubiy Osiyo subtiplari emas, balki hozirgi kundan katta miqyosda migratsiya yo'qligini ko'rsatadi Turkiy - O'rta Osiyoning so'zlovchi aholisi Hindistonga to'g'ri keldi. Evropaliklarda ha haplogroupning yo'qligi, uning janubiy osiyoliklar, sharqiy osiyoliklar va ba'zi bir Markaziy Osiyo populyatsiyalarida bir xil darajada yuqori chastotasi bilan taqqoslaganda, G'arbiy Evroosiyo janubiy osiyolik nasabiga qarama-qarshi.[12]
Janubiy va Janubi-g'arbiy Osiyodagi mavjud mtDNA chegaralarining aksariyati anatomik jihatdan zamonaviy odamlar tomonidan Evroosiyoning dastlabki joylashuvi paytida shakllangan bo'lishi mumkin.[23]
| Haplogroup | Muhim pastki qoplamalar | Populyatsiyalar |
|---|---|---|
| M2 | M2a, M2b | Shimoliy-g'arbiy qismida past darajadagi qit'a bo'ylab Bangladesh, Andra Pradesh, Tamilnadu va Shri-Lankaning qirg'oqlarida eng yuqori cho'qqisi |
| M3 | M3a | Hindistonning shimoli-g'arbiy qismida to'plangan Mumbay Parsees orasida eng yuqori ko'rsatkich |
| M4 | M4a | Pokiston, Kashmir va Andra Pradeshdagi cho'qqilar |
| M6 | M6a, M6b | Kashmir va Shri-Lankaning Bengal ko'rfazi sohillari yaqinida |
| M18 | Butun Janubiy Osiyo bo'ylab Rajastan va Andra-Pradeshda eng yuqori cho'qqisi | |
| M25 | Kerala va Maharashtrada o'rtacha tez-tez, ammo Hindistonning boshqa joylarida kamdan-kam uchraydi |
Macrohaplogroup R



The makrohaplogroup R (juda katta va eski bo'linma makrohaplogroup N ) shuningdek keng tarqalgan va Janubiy Osiyo MtDNA ning boshqa 40% ni tashkil qiladi. Uning juda qadimgi va eng muhim bo'linmasi haplogroup U deb, shu bilan birga G'arbiy Evroosiyo, Janubiy Osiyoga xos bo'lgan bir nechta subkladlarga ega.
R tarkibidagi eng muhim Janubiy Osiyo haplogrouplari:[23]
| Haplogroup | Populyatsiyalar |
|---|---|
| R2 | Sub qit'a bo'ylab keng tarqalgan |
| R5 | Hindistonning aksariyat qismi tomonidan keng tarqalgan. Hindistonning SW sohilidagi cho'qqilar |
| R6 | Hindiston bo'ylab past stavkalarda keng tarqalgan. Tamillar va Kashmirilar orasida eng yuqori cho'qqilar |
| V | Shimoli-g'arbiy shtatlarda joylashgan. Gujarat, Panjob va Kashmirdagi eng yuqori cho'qqilar, boshqa joylarda chastota kam. |
Haplogroup U
Haplogroup U macrohaplogroup R ning sub-haplogroupidir.[23] U haplogroupining tarqalishi M haplogroupi uchun aks ettirilgan tasvirdir: birinchisi shu paytgacha sharqiy osiyoliklar orasida ta'riflanmagan, ammo Evropa aholisi va Janubiy Osiyoliklar orasida tez-tez uchraydi.[24] Janubiy Osiyo U nasllari Evropadagi nasldan ancha farq qiladi va ularning umumiy ajdodga qo'shilishi ham taxminan 50 000 yillarga to'g'ri keladi.[1]
| Haplogroup | Populyatsiyalar |
|---|---|
| U2 * | (a parahaplogroup ) Janubiy Osiyoning shimoliy qismida maxsus ravishda kam tarqalgan. Shuningdek, u Saudiya Arabistonida uchraydi. |
| U2a | Pokiston va Shimoliy Shimoliy Hindistonda, shuningdek Karnatakada nisbatan yuqori zichlikni namoyish etadi, u erda u yuqori zichlikka etadi. |
| U2b | Uttar-Pradeshda eng yuqori kontsentratsiyaga ega, ammo boshqa ko'plab joylarda, ayniqsa Kerala va Shri-Lankada mavjud. U Ummonda ham uchraydi. |
| U2c | Bangladesh va G'arbiy Bengaliyada alohida ahamiyatga ega. |
| U2l | Uttar Pradesh, Shri-Lanka, Sind va Karnataka qismlarida maxsus yuqori konsentrasiyalarga (10% dan yuqori) etib boradigan Janubiy Osiyodagi U subkladalari orasida son jihatdan muhimroqdir. Ummonda ham bu ba'zi bir ahamiyatga ega. mtDNA haplogroup U2i Bamshadda "G'arbiy Evroosiyo" deb nomlangan va boshq. o'rganish, ammo "Sharqiy Evroosiyo (asosan Hindistonga xos)" Kivisildda va boshq. o'rganish. |
| U7 | ushbu haplogroup Gujarat, Panjob va Pokistonda katta ahamiyatga ega. Ushbu haplogrupning vatani Gujarot (eng yuqori chastota, 12%) va Eronni qamrab oladi, chunki u erdan uning chastotasi sharqqa ham, g'arbga ham keskin pasayadi. |
Y xromosoma
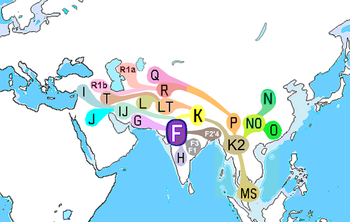
Janubiy Osiyo Y-xromosomasining asosiy DNK haplogrouplari H, J2, L, R1a1 va R2.[13] So'nggi stipendiyalarga ko'ra ularning geografik kelib chiqishi quyidagicha sanab o'tilgan:
| Janubiy Osiyoning asosiy Y-xromosoma nasllari: | H | J2 | L | R1a | R2 |
|---|---|---|---|---|---|
| Basu va boshq. (2003) | izohlarsiz | izohlarsiz | izohlarsiz | Markaziy Osiyo | izohlarsiz |
| Kivisild va boshq. (2003) | Hindiston | G'arbiy Osiyo | Hindiston | Janubiy va G'arbiy Osiyo | Janubiy-Markaziy Osiyo |
| Cordaux va boshq. (2004) | Hindiston | G'arbiy yoki Markaziy Osiyo | Yaqin Sharq | Markaziy Osiyo | Janubiy-Markaziy Osiyo |
| Sengupta va boshq. (2006) | Hindiston | Yaqin Sharq va Markaziy Osiyo | Janubiy Hindiston | Shimoliy Hindiston | Shimoliy Hindiston |
| Tansem va boshq. (2006) | Hindiston | Levant | Yaqin Sharq | Janubiy va Markaziy Osiyo | Janubiy va Markaziy Osiyo |
| Sahoo va boshq. (2006) | Janubiy Osiyo | Yaqin Sharq | Janubiy Osiyo | Janubiy yoki G'arbiy Osiyo | Janubiy Osiyo |
| Mirabal va boshq. (2009) | izohlarsiz | izohlarsiz | izohlarsiz | Shimoliy g'arbiy Hindiston yoki Markaziy Osiyo | izohlarsiz |
| Chjao va boshq. (2009) | Hindiston | Yaqin Sharq | Yaqin Sharq | Markaziy Osiyo yoki G'arbiy Evroosiyo | Markaziy Osiyo yoki G'arbiy Evroosiyo |
| Sharma va boshq. (2009) | izohlarsiz | izohlarsiz | izohlarsiz | Janubiy Osiyo | izohlarsiz |
| Thangaraj va boshq. (2010) | Janubiy Osiyo | Yaqin Sharq | Yaqin Sharq | Janubiy Osiyo | Janubiy Osiyo |
Haplogroup H
Haplogroup H (Y-DNK) Janubiy Osiyoda yuqori chastotada uchraydi. H bugungi kunda Janubiy Osiyodan tashqarida kamdan-kam uchraydi, ammo ular orasida keng tarqalgan Romanis, xususan, H-M82 kichik guruhi. H Evropaning qadimiy namunalarida ham keng tarqalgan va hozirgi kunda ham Evropaliklar va arablarda past chastotada uchraydi Levant. Haplogroup H ko'pincha populyatsiyalar orasida uchraydi Hindiston, Shri-Lanka, Nepal, Pokiston va Maldiv orollari. Uchala filial ham Haplogroup H (Y-DNK) Janubiy Osiyoda joylashgan.
Bu filiali Haplogroup F va pastga tushadi GHIJK oila. Haplogroup H Janubiy Osiyoda 30-40 ming yil oldin paydo bo'lgan deb ishoniladi.[25] Kirish joyi - Janubiy Osiyo, chunki u erda to'plangan. Bu Janubiy Osiyo paleolit aholisining asosiy Y-xromosoma haplogroupini ifodalaydi. Janubiy Osiyodagi ba'zi shaxslar H3 subkladiga (Z5857) nisbatan kamdan-kam uchraydigan ekanligi ko'rsatilgan.[25] Haplogroup H hech qachon ma'lum populyatsiyalar bilan chegaralanmaydi. Masalan, H ga hind-oriy kastalarining taxminan 28,8% i ega.[14][26] va qabilalarda taxminan 25-35%.[16][26]
Haplogroup J2
Haplogroup J2 Janubiy Osiyoda asosan J2a-M410 va J2b-M102 sifatida mavjud bo'lib, neolit davridan beri (9500 YBP).[27][28] J2 qoplamalar Shimoliy-G'arbiy va Janubiy Hindistonda eng yuqori chastotalarga ega[27] va Janubiy Hindiston kastalarida 19%, Shimoliy Hindiston kastalarida 11% va Pokistonda 12% da uchraydi.[14] Yilda Janubiy Hindiston, J2 ning mavjudligi o'rta kastalar orasida 21%, undan yuqori kastlar 18,6% va pastki kastlar orasida 14%.[14] Kast guruhlari orasida J2-M172 ning eng yuqori chastotasi tamil tilida kuzatiladi Vellalar Janubiy Hindistonning, 38,7%.[14] J2 qabilalarda ham mavjud[27] va Avstriya-Osiyo qabilalarida 11% chastotaga ega. Avstriya-Osiyo qabilalari orasida ustunlik J2 Lodxada (35%) uchraydi.[14] J2 Janubiy Hindistonda ham mavjud tepalik qabilasi Toda 38,46% da,[29] ichida Andh qabilasi ning Telangana 35,19% da[16] va Kol qabilasi ning Uttar-Pradesh 33,34% chastotada.[30] Haplogroup J-P209 Hindistonnikida keng tarqalganligi aniqlandi Shia musulmonlari, shundan 28,7% J haplogroupga tegishli bo'lib, J-M410 da 13,7%, J-M267 da 10,6% va J2b da 4,4% (Easvarxant 2009 yil ).
Yilda Pokiston, J2-M172 ning eng yuqori chastotalari orasida Parsis 38,89% da Dravidian Gapirmoqda Brahuis 28,18% va Makrani Balochlar 24% da.[31] Shuningdek, u 18,18% da sodir bo'ladi Makrani Siddis va 3% da Karnataka Siddis.[31][32]
J2-M172 umumiy chastotada 10,3% ni tashkil qiladi Sinhal xalqi ning Shri-Lanka.[33] Yilda Maldiv orollari, Maldiv aholisining 20,6 foizida haplogroup J2 aniqlangan.[34]
Haplogroup L
Doktorning so'zlariga ko'ra Spenser Uels, L-M20 kelib chiqishi Pomir tuguni mintaqa Tojikiston va ko'chib o'tdi Pokiston va Hindiston taxminan 30,000 yil oldin.[35][36][37] Biroq, boshqa ko'plab tadqiqotlarda a G'arbiy Osiyo L-M20 uchun kelib chiqishi va uning kengayishi bilan bog'liq Hind vodiysi (~ 7000 YBP) ga neolitik fermerlar.[31][38][16][26][39][40] Janubiy Osiyoda turli darajalarda topilgan L haplogroupining uchta kichik tarmog'i mavjud: L1-M76 (L1a1), L2-M317 (L1b) va L3-M357 (L1a2).[14]
Hindiston
Haplogroup L neolitik kengayish vaqtini ko'rsatadi.[41] Qopqoq hind populyatsiyasida umumiy chastotada mavjud. 7-15%.[14][16][42][26] Haplogroup L janubiy hind kastlari orasida yuqori chastotaga ega (taxminan 17-19%) va ba'zi kastalarda 68% gacha etadi Karnataka ammo shimoliy hind kastalarida biroz kam uchraydi (taxminan 5-6%).[14] Qaplogrup L borligi qabila guruhlari orasida juda kam uchraydi (taxminan 5,6-7%),[14][16][26] ammo o'rtacha darajadagi 14,6% kuzatilgan Chenchus.[33]
Mintaqaviy va ijtimoiy guruhlar orasida o'rtacha va yuqori chastotalar Konkanasta Braxminlarida (18,6%), Panjoblarda (12,1%), Gujaratisda (10,4%), Lambadilar (17.1%), Jats (36.8%)[33][36]
Pokiston
Pokistonda L-M20 ning L1-M76 va L3-M357 subkladalari umumiy chastotalarga mos ravishda 5,1% va 6,8% ga etadi.[14]Haplogroup L3 (M357) tez-tez uchraydi Burusho (taxminan 12%[43]) va Pashtunlar (taxminan 7%[43]). Uning eng yuqori chastotasini janubi-g'arbiy qismida topish mumkin Balujiston bo'ylab viloyat Makran sohil (28%) ga Hind daryosi delta. L3a (PK3) taxminan 23% da uchraydi Nuristani shimoli-g'arbda Pokiston.[43]
Qopqoq Pokiston aholisi o'rtasida o'rtacha taqsimotda mavjud (taxminan 14%).[44][45]
Shri-Lanka
Bir tadqiqotda sinhallarning 16% Haplogroup L-M20 musbat ekanligi aniqlandi.[46] Boshqa bir tadqiqotda 18% L1 ga tegishli ekanligi aniqlandi.[33]
Haplogroup R1a1
Janubiy Osiyoda R1a1 ko'pincha bir qator demografik guruhlarda yuqori chastotali kuzatilgan,[15][47][48] shuningdek, eng yuqori ko'rsatkich bilan STR ba'zilar uni kelib chiqish joyi deb bilishga olib keladigan xilma-xillik.[33][30][49]
R1a taxminan paydo bo'lgan bo'lsa-da. 22000[30] 25000 gacha[50] bir necha yil oldin, uning M417 (R1a1a1) subklade-si xilma-xil edi. 5800 yil oldin.[50] M417-subkladlarning R1-Z282 (shu jumladan R1-Z280) tarqalishi[51] Markaziy va Sharqiy Evropada va Osiyoda R1-Z93[51][50] R1a1a ning ichida diversifikatsiya qilinganligini ko'rsatadi Evroosiyo dashtlari yoki Yaqin Sharq va Kavkaz mintaqa.[51] Ushbu subkladlarning kelib chiqish joyi kelib chiqishi haqidagi munozarada rol o'ynaydi Hind-evropaliklar.
Hindiston
Yilda Hindiston, ushbu haplogrupning yuqori foizlari kuzatiladi G'arbiy Bengal braxminlari (72%)[47] sharqda, Gujarat Lohanas (60%)[48] g'arbda, Xatrislar (67%)[48] shimolda, Iyengar Braxmanlar (31%) janubda.[47] Bundan tashqari, bir nechtasida topilgan Janubiy hind Dravidian -Gapirmoqda qabilalar shu jumladan Tamil Nadu kotalari (41%)[52] Chenchu (26%) va Valmikis Andxra-Pradesh[33] shuningdek Yadav va Kallar ning Tamil Nadu M17 ushbu janubiy hindular qabilalarida keng tarqalganligini ko'rsatmoqda.[33] Bundan tashqari, tadqiqotlar mintaqaviy xilma-xil guruhlarda yuqori foizlarni ko'rsatmoqda Manipuris (50%)[48] haddan tashqari Shimoliy Sharqqa va ular orasida Panjob (47%)[33] haddan tashqari Shimoliy G'arbga.
Pokiston
Pokistonda u Mohanna of orasida 71% uchraydi Sind viloyati janubda va 46% orasida Baltis ning Gilgit-Baltiston shimolga.[48]
Shri-Lanka
23% Sinhal xalqi 87 ta sub'ektning namunalaridan 2003 yilgi tadqiqotlarga ko'ra R1a1a (R-SRY1532) ijobiy ekanligi aniqlandi.[46]
Maldiv orollari
Yilda Maldiv orollari, Ning 23,8% Maldiv xalqi R1a1a (M17) musbat ekanligi aniqlandi.[34]
Nepal
Insonlar Teray Mintaqa, Nepal R1a1a ni 69% da ko'rsating.[53]
Haplogroup R2
Janubiy Osiyoda R2 va R2a nasab Hindistonda 10-15% atrofida va Shri-Lanka va Pokistonda 7-8%. R-M124 shaxslarining kamida 90% Janubiy Osiyoda joylashgan.[54] Shuningdek, bu haqida xabar berilgan Kavkaz va Markaziy Osiyo past chastotada. Mondal va boshqalarning genetik tadqiqotlari. 2017 haplogroup degan xulosaga keldi Haplogroup R2 Shimoliy Hindistonda paydo bo'lgan va Dasht ko'chishidan oldin mavjud bo'lgan.[55]
Hindiston
Mintaqaviy guruhlar orasida u orasida mavjud G'arb Bengaliyaliklar (23%), Nyu-Dehli Hindular (20%), Panjob (5%) va Gujaratlar (3%).[33] Qabila guruhlari orasida Karmalis of G'arbiy Bengal 100% eng yuqori ko'rsatkichni ko'rsatdi[15] dan so'ng Lodalar (43%)[56] sharqda esa Bhil ning Gujarat g'arbda 18%,[30] Tarus shimol uni 17% ko'rsatdi,[4] Chenchu va Pallan janubi mos ravishda 20% va 14% ni tashkil etdi.[15][14] Kast guruhlari orasida yuqori foizlar ko'rsatilgan Jaunpur Kshatriyalar (87%), kamma (73%), Bihar Yadav (50%), Xandayat (46%) va Kallar (44%).[15]
Shuningdek, bu ko'pchilikda sezilarli darajada yuqori Braxmin guruhlar, shu jumladan Panjob Brahminlari (25%), Bengaliyalik braxmanlar (22%), Konkanasta Braxmanlar (20%), Chaturvedis (32%), Bhargavas (32%), Kashmiri panditslari (14%) va Lingayat Braxmanlar (30%).[30][17][4][15]
Shimoliy Hindiston musulmonlarining chastotasi 19% (Sunniy ) va 13% (Shia ),[17] esa Dovudiy Bohra Muslim G'arbiy G'arot shtatida 16% va Mappila musulmonlari Janubiy Hindistonning chastotasi 5% ni tashkil qiladi.[57]
Pokiston
R2 haplogroupi 14 foizida uchraydi Burusho odamlar.[43] Orasida Xunza xalqi u 18% da topilgan Parsis uni 20% da ko'rsating. Shuningdek, shimoliy-sharqiy qismida joylashgan Afg'oniston.[iqtibos kerak ]
Shri-Lanka
Ning 38% Sinhal tili Shri-Lankaning 2003 yilgi tadqiqotlariga ko'ra R2 musbat ekanligi aniqlandi.[33]
Maldiv orollari
12% Maldiv xalqi Maldiv orollarida R2 borligi aniqlandi.[34]
Nepal
Nepalda R2 foizlar har xil tadqiqotlar ostida turli guruhlar ichida 2% dan 26% gacha. Yangilar odamlar esa 26% sezilarli darajada yuqori chastotani namoyish etadi Katmandu uni 10% da ko'rsating.
Janubiy Osiyo aholisi tarixini tiklash
The Hindiston genomining o'zgarishi bo'yicha konsortsium (2008), Janubiy Osiyo aholisini to'rt etnolingvistik guruhga ajratadi: Hind-evropa, Dravidian, Tibet-Burman va Avstriya-Osiyo.[58][59][60][61][62] Molekulyar antropologiya tadqiqotlarida uch xil turdagi markerlardan foydalaniladi: onadan naslga o'tgan va yuqori polimorfik bo'lgan mitoxondriyal DNK (mtDNA) o'zgarishi, erkak chiziqlar bo'ylab uniparental uzatishni o'z ichiga olgan Y xromosomalar o'zgarishi va DNKning avtosomal o'zgarishi.[4]:04
mtDNA o'zgarishi
MtDNA o'zgarishiga asoslangan tadqiqotlarning aksariyati til, kasta va qabila guruhlari bo'yicha Janubiy Osiyo populyatsiyasining genetik birligi haqida xabar bergan.[1][2][3] Ehtimol, M haplogroupi 78000 yil avvalgi migratsiya to'lqini bilan Janubiy yo'l bo'ylab Sharqiy Afrikadan Osiyoga olib kelingan.[1]
Ga binoan Kivisild va boshq. (1999), "Boshqa Evroosiyo populyatsiyalarida tasvirlangan nasl-nasablar bilan kichik bir-birining ustiga chiqib ketish, so'nggi immigratsiyalar onaning tug'ma tuzilishiga juda oz ta'sir qilganligini aniq ko'rsatib turibdi. genofond Janubiy osiyoliklar. Hindistonda mavjud bo'lgan farqlarga qaramay, bu populyatsiyalar cheklangan miqdordagi asoschilar nasabidan kelib chiqadi. Ushbu nasablar, ehtimol, Janubiy Osiyoga O'rta paleolit davrida, 48000 yil oldin Evropa va umuman olganda Qadimgi dunyo buzilishidan oldin kiritilgan. "[1] Basu va boshq. (2003), shuningdek, Hindistondagi ayollar nasablarining asosiy birligini ta'kidlaydi.[42]
Y Xromosomalarning o'zgarishi
Y xromosomalar o'zgarishiga asoslangan xulosalar mtDNA o'zgarishiga asoslangan holda turli xil bo'lgan. Esa Kivisild va boshq. (2003) Janubiy Osiyodagi erkak nasablarining qadimiy va umumiy genetik merosini taklif qiladi, Bamsad va boshq. (2001) janubiy osiyolik erkaklar nasablari va g'arbiy evrosiyoliklar o'rtasida yuqori tabaqa darajasiga mutanosiblikni taklif qiladi va janubiy Hindiston shtatlarining yuqori tabaqa aholisini yaqinroq joylashtiradi. Sharqiy evropaliklar.[63]
Basu va boshq. (2003) xulosasiga ko'ra, Avstriya-Osiyo qabilalari populyatsiyalari Hindistonga birinchi bo'lib shimoli-g'arbiy yo'lakdan, keyinchalik ularning bir qismi shimoliy-sharqiy yo'lak orqali kirib kelishgan.[42] Holbuki, Kumar va boshq. (2007) 25 ta Janubiy Osiyo avstro-osiyo qabilalarini tahlil qildi va Janubiy Osiyo avstro-osiyo populyatsiyasining sub-lingvistik guruhlari orasida kuchli otalik genetik aloqani topdi.[56] Mukherji va boshq. (2001) Pokiston va Shimoliy hindularni g'arbiy Osiyo va Markaziy Osiyo aholisi o'rtasida joylashtiradi,[64] Cordaux va boshq. (2004) hindlarning kast aholisi Markaziy Osiyo aholisiga yaqinroq ekanligini ta'kidlamoqda.[26] Sahoo va boshq. (2006) va Sengupta va boshq. (2006) shuni ko'rsatadiki, hind kast aholisi so'nggi paytlarda aralashmalarga duch kelmagan.[14][15] Sanghamitra Sahoo o'z tadqiqotini quyidagicha yakunlaydi:[15]
Hozirgi dalillarga asoslanib, ko'chmanchi qishloq xo'jaligi boshlangan davrda hindlarning aksariyat qismining otalik merosining kelib chiqishini Janubiy Osiyodan qidirib topish shart emas. Shimoliy g'arbiy koridor orqali birgalikda Hindistonga kelgan odamlar, til va qishloq xo'jaligining ko'p yillik kontseptsiyasi jiddiy tekshiruvga ega emas. So'nggi paytlarda J2, L, R1a va R2 haplogrouplarini hind kastalarining Janubiy Osiyodan tashqaridagi otalik nasablarining aksariyati uchun zamondosh kelib chiqishi bilan bog'lash to'g'risidagi da'volari rad etildi, ammo bizning topilmalarimiz F * haplogrouplarining mahalliy kelib chiqishini qo'llab-quvvatlamoqda. H. Boshqalar orasida faqat J2 Markaziy Osiyodan emas, balki G'arbiy Osiyodan bo'lgan so'nggi tashqi hissani ko'rsatadi. Gaplogrup chastotalarining joriy taqsimoti, nasablar bundan mustasno, asosan madaniy determinantlar emas, asosan geografik yo'naltirilgan. Ajablanarlisi shundaki, aynan Hindistonning shimoli-sharqida, sil kasalliklari guruhlari orasida genlar, madaniyat va tillar bo'yicha kuzatiladigan demik diffuziya bo'yicha aniq dalillar mavjud, ammo aftidan qishloq xo'jaligi.
Mondal va boshqalar tomonidan o'tkazilgan eng yaqin qo'shni tahlil. 2017 yil Hindistonning Y-avlodlari janubga yaqin degan xulosaga keldi Evropa populyatsiyalar va ikkala ilgari paydo bo'lgan Dasht migratsiyasi o'rtasidagi farqlanish vaqti. ":[65]
Ushbu natijalar shuni ko'rsatadiki, hind populyatsiyasida Evropaga oid ajdodlar kutilganidan ancha yoshi va murakkabroq bo'lishi mumkin va birinchi to'lqinli qishloq xo'jaligi dalillaridan kelib chiqishi yoki undan ham oldinroq bo'lishi mumkin.
— Mondal va boshq. 2017 yil
Avtosomal DNKning o'zgarishi
AASI-ANI-ASI
Avtosomal DNK o'zgarishiga asoslangan tadqiqotlar natijalari ham har xil. 500000 dan ortiq biallelik avtosomal markerlardan foydalangan holda (2009) o'tkazilgan katta tadqiqotda Reyx zamonaviy Janubiy Osiyo aholisi genotsial jihatdan ajralib turuvchi ajdodlar populyatsiyasining Xolotsen davridan keyingi davrga kelib qo'shilishi natijasida kelib chiqqan deb taxmin qildi. Bu ikki "rekonstruksiya qilingan" qadimiy populyatsiyalarni u "Ancestral South Indian" (ASI) va "Ancestral North Indians" (ANI) deb atagan. Reyxning so'zlariga ko'ra: "ANI ajdodlari hind-evropada Dravidian ma'ruzachilariga qaraganda ancha yuqori bo'lib, ajdodlar ASI ANI bilan aralashmasidan oldin Dravid tilida gaplashgan bo'lishi mumkin". ANI genetik jihatdan O'rta Sharq aholisi, Markaziy Osiyo va Evropaliklarga yaqin bo'lsa, ASI subkontinent tashqarisidagi guruhlar bilan chambarchas bog'liq emas. "ASI" qadimiy DNK mavjud emasligi sababli, mahalliy aholi Andamancha Onge ASI (nomukammal) ishonchli vakili sifatida ishlatiladi (Reyx va boshq. fikriga ko'ra, Andamanliklar, ulardan ajralib tursalar ham, ASIga eng yaqin tirik aholi hisoblanadi). Reyx va boshqalarning fikriga ko'ra, ANI va ASI ajdodlari subkontinent bo'ylab (Hindistonning shimoliy va janubiy qismida) turli xil nisbatlarda uchraydi va "ANI ajdodlari Hindistonda 39-71% oralig'ida va an'anaviy yuqori qismida yuqori kast va hind-evropa ma'ruzachilari. "[66]
Moorjani va boshq. 2013 yilda ASI biron bir tirik guruh bilan chambarchas bog'liq bo'lmasa-da, "mahalliy Andaman orollari aholisi bilan (uzoqdan) qarindosh" ekanligini ta'kidlagan. Moorjani va boshq. shuningdek, ASI bilan bog'liq bo'lgan populyatsiyadan Andamanese genlarning kirib borishini taklif qiladi. Tadqiqot natijalariga ko'ra "hinduevropa yoki dravid tillarida gaplashadigan deyarli barcha guruhlar G'arbiy Evroosiyoliklar bilan PCA (" hind klini "deb nomlanadi) bilan bog'liqlik gradyenti bo'ylab yotadi".[67]
2013 yilda bitta nukleotidli polimorfizm (SNP) yordamida o'tkazilgan tadqiqot shuni ko'rsatadiki, Andaman xalqi (Onge) genomi Janubiy Osiyoliklarga qaraganda boshqa Okeanik negrito guruhlariga yaqinroq.[68]
Basu va boshqalarning fikriga ko'ra. 2016 yilda o'tkazilgan keyingi tahlil shuni ko'rsatdiki, materik hind populyatsiyasining genomik tuzilishi to'rtta ajdod tarkibiy qismlarining hissalari bilan izohlanadi. ANI va ASI-dan tashqari, Basu va boshqalar. al (2016) materik Hindistondagi ikkita ajdod tarkibiy qismlarini aniqladi, ular avstro-osiyo tilida so'zlashadigan qabilalar va Tibet-burman tilida so'zlashuvchilar uchun asosiy hisoblanadi, ular AAA ("Ancestral Austro-Osiyo" uchun) va ATB ("Ancestral Tibeto" uchun) -Burman ") navbati bilan. Tadqiqot shuningdek, aholisi Andaman orollari arxipelag aniq ajdodlarni shakllantiradi, "bu koansestral deb topilgan Okean populyatsiyalar ".[8]
ANI va ASI nasablari orasidagi qo'shilish klini c davriga to'g'ri keladi. 4.2-1.9 kya Moorjani va boshq. (2013), hind bronza davriga to'g'ri keladi va mualliflar tomonidan deurbanizatsiya jarayoni bilan bog'liq Hind vodiysi tsivilizatsiyasi va aholi boshlang'ich hind temir davrida Gangetik tizimga o'tadi.[6] Basu va boshq. (2003) "Dravidian qabilalari hind-evropada so'zlashadigan ko'chmanchilar kelguniga qadar butun Hindistonda keng tarqalgan" degan fikrni va "asoschilar va drift effektlarini keltirib chiqaradigan populyatsiyalarning bo'linishi bilan shakllanishi zamonaviy genetik tuzilmalarda o'z izlarini qoldirdi" populyatsiyalar ".[42] Genetika bo'yicha mutaxassis P.P.Majumder (2010) yaqinda Reyx va boshq. (2009) mtDNA va Y-DNK yordamida oldingi tadqiqotlar bilan juda mos keladi:[69]
O'rta Osiyo aholisi hind genofondiga, xususan shimoliy hind genofondiga katta hissa qo'shgan bo'lishi kerak edi va muhojirlar go'yoki Hindistonga hozirgi Afg'oniston va Pokiston orqali ko'chib kelishgan. Foydalanish mitoxondrial DNK Turli xil tadqiqotlar natijalariga ko'ra varyasyon ma'lumotlari shuni ko'rsatdiki, Markaziy Osiyo va Pokiston populyatsiyalari shimoliy hind populyatsiyalari bilan genetik farqlanishning eng past koeffitsientini, janubiy hind populyatsiyalari bilan yuqori darajadagi farqni va shimoliy-sharqiy hind populyatsiyalari bilan eng yuqori ko'rsatkichni ko'rsatmoqda. Shimoliy hind populyatsiyalari genetik jihatdan Hindistonning boshqa geografik mintaqalari aholisiga qaraganda Markaziy Osiyoliklarga yaqinroqdir .... Yuqoridagi topilmalarga muvofiq, 500.000 dan ortiq biallelik autosomal markerlardan foydalangan holda o'tkazilgan so'nggi tadqiqotlar shimoldan janubiy gradientni hind populyatsiyasining Evropaning g'arbiy qismiga genetik yaqinligini topdi. Ushbu xususiyat, ehtimol, g'arbiy Evroosiyo genofondidan kelib chiqqan nasablarning nisbati bilan bog'liq bo'lishi mumkin, bu tadqiqot shuni ko'rsatdiki, Hindistonning shimoliy qismida yashovchi populyatsiyalarda janubiy Hindistonda yashovchilarga qaraganda ko'proq.
Chaubey va boshq. 2015 yilda Sharqiy Osiyo ajdodlarining o'ziga xos komponenti aniqlandi, asosan Himoloy etaklarida va Hindistonning shimoliy-sharqiy qismida ma'lum populyatsiyalar bilan cheklangan. Komponentning eng yuqori chastotasi shimoli-sharqdagi Tibet-Birma tilida so'zlashuvchi guruhlar orasida kuzatilgan va Androman populyatsiyasida ham aniqlangan, bu erda Austroasiatik ma'ruzachilar katta ishtirok etishgan. Himolay tog 'etaklarida va Hindistonning markaziy-janubiy qismida yashovchi ba'zi bir etnik guruhlar bundan mustasno, hind-evropa va dravid tilida so'zlashuvchilarda umuman yo'qligi aniqlandi.[10] Tadqiqotchilar shuni ta'kidladilarki, o'rganilgan Andaman guruhlarida o'lchangan Sharqiy Osiyo ajdodi (Xan vakili) aslida Andamanaliklarning melaneziyaliklar va Malayziya Negritoslari bilan yaqinligini (haqiqiy Sharqiy Osiyo aralashmasi o'rniga) aks ettirishi mumkin, deb taxmin qilishdi.[10] Chaubey va boshqalarning oldingi tadqiqotlari kabi. Andamanliklar, melaneziyaliklar va boshqa negrito guruhlari (shuningdek, janubiy osiyoliklar) o'rtasida "chuqur umumiy nasab" taklif qildi,[10] va Janubi-Sharqiy Osiyo negritoslari va melaneziyaliklar (shuningdek, Andamanliklar) Sharqiy Osiyoliklar bilan yaqinligi.[68]
Lazaridis va boshq. (2016) eslatmalarida "Dasht bilan bog'liq populyatsiyalarning Janubiy Osiyodagi demografik ta'siri, chunki Mala, janubiy hind Dalit Shimoliy Hindiston (ANI) minimal ajdodlari bilan "hind klinasi" bo'ylab bu nasl-nasabga ega bo'lganlar ~ 18% dasht bilan bog'liq ajdodlar, esa Kalash Pokistonning ~ 50% dasht bilan bog'liq ajdodlari bor degan xulosaga kelishdi. "[70] Lazaridis va boshqalarning 2016 yilgi tadqiqotida Janubiy Osiyoliklarning dasht bilan bog'liq aralashmasi (6,5-50,2%) taxmin qilingan. Lazaridis va boshq. "kelajakdagi tadqiqotlarning foydali yo'nalishi - bu dasht populyatsiyalari, shuningdek Markaziy Osiyo aholisi (Eronning sharqi va dashtning janubi) dan qadimgi DNKni yanada kengroq tanlab olishdir. bu erda va Janubiy Osiyoda to'g'ridan-to'g'ri hududdagi aholi o'zgarishi traektoriyasini aniqlash uchun ko'rib chiqilganlar.
Patxak va boshq. 2018 yil yakunlariga ko'ra Hind-evropa ma'ruzachilar Gangetik tekisliklar va Dravidian ma'ruzachilar muhim ahamiyatga ega Yamnaya erta-o'rta bronza davri (Steppe_EMBA) ajdodlari, ammo o'rta bronza davridagi Dasht (Steppe_MLBA) ajdodlari yo'q. Boshqa tomondan, "Shimoliy-G'arbiy Hindiston va Pokiston" aholisi (PNWI) Yamnaya (Steppe_EMBA) ajdodlari bilan bir qatorda muhim Steppe_MLBA ajdodlarini ko'rsatdilar. Tadqiqotda ta'kidlanishicha, qadimgi Janubiy Osiyo namunalarida Steppe_EMBA (yoki Yamnaya) ga nisbatan Steppe_MLBA sezilarli darajada yuqori bo'lgan. Tadqiqot shuningdek, Rors ANI uchun proksi sifatida ishlatilishi mumkin.[71]
Devid Reyx uning 2018 yilgi kitobida Biz kimmiz va qanday qilib bu erga keldik 2016 yilgi tahlillarda ASIda eronlik dehqonlar tomonidan kelib chiqadigan ajdodlar tarkibiy qismi (ularning nasablarining taxminan 25%), qolgan 75% esa Janubiy Osiyodagi ovchilarni yig'uvchilaridan olinganligi aniqlandi. Uning qo'shimcha qilishicha, ASI ilgari tashkil topgan Janubiy Osiyoning mahalliy ovchilarni yig'ishi ehtimoldan yiroq, ammo qishloq xo'jaligini butun Janubiy Osiyoda tarqatish uchun mas'ul bo'lgan aholi. ANI misolida Eron dehqonlarining nasablari 50% ni tashkil qiladi, qolganlari Yamnaya bilan bog'liq bo'lgan dasht guruhlaridan.[72]
Narasimxon va boshq. (2018), xuddi shunday, ANI va ASI miloddan avvalgi 2-ming yillikda shakllangan degan xulosaga kelish.[73] Ulardan oldin AASI aralashmasi (qadimgi ajdodlar janubiy hindu, ya'ni ovchilarni yig'uvchilar Andamanliklar bilan umumiy ildizni baham ko'rishgan); va Hindistonga etib kelgan Eron qishloq xo'jaligi dehqonlari Miloddan avvalgi 4700-3000 yillarda va "miloddan avvalgi 4-ming yillikda Hind vodiysiga etib borgan bo'lishi kerak".[73] Narasimxon va boshqalarning fikriga ko'ra, hind vodiysi tsivilizatsiyasiga xos bo'lgan ushbu aralash aholi miloddan avvalgi 2-ming yillikda shakllangan "ANI va ASIga katta nisbatlarda hissa qo'shgan". ANI "aralashmasidan hosil bo'lganHind daryosibilan bog'liq guruhlar "va dashtdan kelgan migrantlar, ASI esa tashqaridan tuzilgan"Hind daryosibog'liq guruhlar "janubga ko'chib, mahalliy ovchilar bilan aralashgan. ASI aholisining ajdodlari AASI dan o'rtacha 73% va Eronga tegishli dehqonlar orasida 27% ni tashkil etgan. Narasimxon va boshqalar. Hind periferiyasi guruhi har doim AASI va Eronning dehqonchilik bilan bog'liq nasablarining bir xil ikki proksimal manbalari aralashmasi; "AASI nasabiga ega bo'lgan ~ 42% AASI ajdodiga ega bo'lgan Hind atrofi shaxslaridan biri va ~ 14-18% AASI ajdodiga ega bo'lgan boshqa ikki shaxs" bilan (ularning nasl-nasabining qolgan qismi Eron qishloq xo'jaligi bilan bog'liq aholidan bo'lganligi bilan).[73] Mualliflarning ta'kidlashicha, AASI mahalliy ovchilarni yig'adiganlar Sharqiy Osiyo, Onge (Andaman) va Avstraliyaning tub aholisi ajdodlari bir-biridan ajratib turadigan davrda ajralib chiqqan turli xil filialni anglatadi. "Hozirgi sharqiy va janubiy osiyoliklarning barcha ajdodlari (G'arbiy Evroosiyoga tegishli janubiy osiyoliklarga qo'shilishidan oldin) yagona sharqqa tarqalishidan kelib chiqadi, bu qisqa vaqt ichida AASIga olib keladigan nasllarni keltirib chiqardi. , Sharqiy Osiyoliklar, Onge va Avstraliyaliklar.[73]
Yelmen va boshqalarning genetik tadqiqotlari. (2019) shuni ko'rsatadiki, zamonaviy Janubiy Osiyo aholisi odatda G'arbiy Evroosiyoliklarga eng yaqin. Ular zamonaviy janubiy osiyoliklar asosan mahalliy Janubiy Osiyo genetik komponenti va keyinchalik paydo bo'lgan G'arbiy-Evroosiyo komponentining aralashmasi (G'arbiy Osiyo va g'arbiy Dashtlardan olingan) degan xulosaga kelishdi. Mualliflar, shuningdek, mahalliy Janubiy Osiyoning genetik komponenti Andamanaliklardan ajralib turishini va shuning uchun Andamanaliklar nomukammal proksi ekanligini ta'kidlaydilar. Ushbu komponent (Andamanese Onge tomonidan namoyish etilganda) Hindistonning shimoliy qismida aniqlanmadi Gujarati va shu tariqa Janubiy Hindiston qabilasi deb taxmin qilinadi Paniya odamlar (asosan ASI ajdodlari guruhi) zamonaviy janubiy osiyoliklarda "mahalliy Janubiy Osiyo" komponenti uchun Andamaniklardan (Onge) ko'ra yaxshiroq proksi sifatida xizmat qilishlari mumkin edi, chunki Paniya to'g'ridan-to'g'ri Janubiy Osiyodan kelib chiqqan (uzoqdan emas) ular bilan Onge kabi bog'liqdir).[9]
Ikki genetik tadqiqotlar (Shinde va boshq. 2019 va Narasimhan va boshq., 2019) Hind vodiysi tsivilizatsiyasidan qolgan (bronza asri Shimoliy G'arbiy Hindiston va Sharqiy Pokistonning ba'zi qismlari) qoldiqlarini tahlil qilib, ularni ajdodlar aralashmasi bilan topdilar: Shinde va boshq. ularning namunalarida genomning taxminan 50-98% i eronlik dehqonlar bilan bog'liq bo'lgan xalqlardan va ularning genomining 2-50% dan janubiy osiyolik ovchilarni yig'ishgan va o'zlarining nasablarini Andamanliklar bilan birlashtirgan eronliklarga tegishli. ajdodlari o'rtacha ustunlik qiladi. Narasimxon va boshqalar tomonidan tahlil qilingan namunalar. 45-82% eronlik dehqonlar bilan bog'liq nasabga va 11-50% AASI (yoki Andamanikka bog'liq ovchi-terib ajdodlari) bo'lgan. Ikkala tadqiqotning tahlil qilingan namunalarida hech kimga kam bo'lmagan "Dasht ajdodlari "Hindistonga keyingi Hind-Evropa migratsiyasi bilan bog'liq bo'lgan tarkibiy qism. Mualliflar ushbu ajdodlarning tegishli miqdori individual ravishda sezilarli darajada o'zgarib turishini aniqladilar va Hindiston aholisi tarixini to'liq tasavvur qilish uchun ko'proq namunalar olish kerak degan xulosaga kelishdi.[74][75]
Kasta guruhlari va qabilalar o'rtasidagi genetik masofa
Watkins va boshq. (2005) va Kivisild va boshq. (2003) avtosomal markerlarga asoslanib hind kastalari va qabila populyatsiyalari umumiy nasabga ega degan xulosaga kelishdi.[33][76] Reddi va boshq. (2005) janubning kast guruhlari bo'yicha bir xil allel chastotalarini taqsimlanishini aniqladi Andxra-Pradesh, ammo kasta guruhlari va qabilalar o'rtasidagi genetik masofa sezilarli darajada katta genetik izolyatsiya qabilalar va kastalarning.[77]
Visvanatan va boshqalar. (2004) janubiy Hindistonning qabilaviy populyatsiyalari orasidagi genetik tuzilish va yaqinliklarga oid tadqiqotda shunday xulosaga keldi: "Genetik farqlash yuqori bo'lgan va genetik masofalar geografik masofalar bilan sezilarli darajada bog'liq emas edi. Shuning uchun genetik drift, ehtimol janubiy hindu qabilaviy populyatsiyalarida kuzatilgan genetik o'zgarishlarning shakllarini shakllantirishda muhim rol o'ynagan. Aks holda, aholi o'rtasidagi munosabatlarni tahlil qilish shuni ko'rsatdiki, fenotipik xususiyatlaridan qat'iy nazar barcha hind va janubiy osiyo populyatsiyalari bir-biriga o'xshashdir va afrikaliklarga nisbatan yaqinlikni ko'rsatmaydi. Xindistonlik ayrim guruhlarning afrikaliklarga fenotipik o'xshashligi degan xulosaga keldik bunday qilma ushbu guruhlar o'rtasidagi yaqin munosabatlarni aks ettiradi, ammo yaxshiroq tushuntiriladi yaqinlashish."[78]
2011 yilda chop etilgan tadqiqot Amerika inson genetikasi jurnali[5] hindlarning ajdodlari tarkibiy qismlari ilgari o'ylanganiga qaraganda ancha murakkab demografik tarix natijasidir. According to the researchers, South Asia harbours two major ancestral components, one of which is spread at comparable frequency and genetic diversity in populations of Central Asia, West Asia and Europe; the other component is more restricted to South Asia. However, if one were to rule out the possibility of a large-scale Indo-Aryan migration, these findings suggest that the genetic affinities of both Indian ancestral components are the result of multiple gene flows over the course of thousands of years.[5]
A genetic study published in 2020 in the Journal ScienceDirect by Rowold et al. found that the great majority of Hind xalqi ("caste people"), which make up about 91,4%, are closely related to West-Eurasians (Evropaliklar va O'rta sharqliklar ). Northern Africans (samplified by Misrliklar ) were part of this cluster but shifted towards Saxaradan Afrikaliklar. Sharqiy osiyoliklar va Janubi-sharqiy osiyoliklar were most distant to both West-Eurasians and Sub-Saharan Africans and formed a completely separate cluster. Interesingly, non-caste populations of India (samplified by the Soliga xalqi, lardan biri Adivasi ) which make up about 8,6% of Indians population, were found to be shifted towards Sub-Saharan Africans and share specific alleles. A link to certain Oceanians was also detected, suggesting ancient gene-flow.[79]
Moreover, the presence of these Y-STR profiles in several sub-Saharan populations and conspicuous absence from the other Eurasian collections suggest a unique genetic connection between Indian tribal groups and sub-Saharan Africans.
— Rowold et al. 2020 yil
Shuningdek qarang
- Arxeogenetika
- Janubiy Osiyodagi etnik guruhlar
- List of ethnolinguistic regions of South Asia
- Peopling of India
- Janubiy Osiyo populyatsiyalaridagi Y-DNK haplogrouplari
- Gujarati odamlari bo'yicha genetik tadqiqotlar
Adabiyotlar
- ^ a b v d e Kivisild T, Kaldma K, Metspalu M, Parik J, Papiha S, Villems R (1999). "The Place of the Indian Mitochondrial DNA Variants in the Global Network of Maternal Lineages and the Peopling of the Old World". Genomic Diversity. 135–152 betlar. doi:10.1007/978-1-4615-4263-6_11. ISBN 978-1-4613-6914-1.CS1 maint: ref = harv (havola)
- ^ a b Baig MM, Khan AA, Kulkarni KM (September 2004). "Mitochondrial DNA diversity in tribal and caste groups of Maharashtra (India) and its implication on their genetic origins". Inson genetikasi yilnomalari. 68 (Pt 5): 453–60. doi:10.1046/j.1529-8817.2004.00108.x. PMID 15469422. S2CID 23032872.
- ^ a b Singh AK (2007). Science & Technology For Upsc. Tata McGraw-Hill ta'limi. p. 595. ISBN 978-0-07-065548-5. Arxivlandi asl nusxasidan 2014 yil 3 yanvarda. Olingan 24 may 2016.
- ^ a b v d Tripathy, Vikal; Nirmala, A.; Reddy, B. Mohan (4 September 2017). "Trends in Molecular Anthropological Studies in India". Xalqaro genetika jurnali. 8 (1–2): 1–20. doi:10.1080/09723757.2008.11886015. S2CID 12763485.
- ^ a b v Metspalu M, Romero IG, Yunusbayev B, Chaubey G, Mallick CB, Hudjashov G, et al. (2011 yil dekabr). "Shared and unique components of human population structure and genome-wide signals of positive selection in South Asia". Amerika inson genetikasi jurnali. 89 (6): 731–44. doi:10.1016/j.ajhg.2011.11.010. PMC 3234374. PMID 22152676.
- ^ a b Moorjani P, Thangaraj K, Patterson N, Lipson M, Loh PR, Govindaraj P, et al. (2013 yil sentyabr). "Genetic evidence for recent population mixture in India". Amerika inson genetikasi jurnali. 93 (3): 422–38. doi:10.1016/j.ajhg.2013.07.006. PMC 3769933. PMID 23932107.
- ^ Silva M, Oliveira M, Vieira D, Brandão A, Rito T, Pereira JB, et al. (2017 yil mart). "A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals". BMC evolyutsion biologiyasi. 17 (1): 88. doi:10.1186/s12862-017-0936-9. PMC 5364613. PMID 28335724.
- ^ a b Basu A, Sarkar-Roy N, Majumder PP (February 2016). "Genomic reconstruction of the history of extant populations of India reveals five distinct ancestral components and a complex structure". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 113 (6): 1594–9. Bibcode:2016PNAS..113.1594B. doi:10.1073/pnas.1513197113. PMC 4760789. PMID 26811443.
- ^ a b Yelmen, Burak; Mondal, Mayux; Marnetto, Davide; Pathak, Ajai K.; Montinaro, Francesco; Gallego Romero, Irene; Kivisild, Toomas; Metspalu, Mait; Pagani, Luca (1 August 2019). "Ancestry-Specific Analyses Reveal Differential Demographic Histories and Opposite Selective Pressures in Modern South Asian Populations". Molekulyar biologiya va evolyutsiya. 36 (8): 1628–1642. doi:10.1093/molbev/msz037. ISSN 0737-4038. PMC 6657728. PMID 30952160.
- ^ a b v d Chaubey, Gyaneshwer (January 2015). "Sharqiy Osiyo ajdodlari Hindistonda" (PDF). Indian Journal of Physical Anthropology and Human Genetics. 34 (2): 193–199.
Here the analysis of genome wide data on Indian and East/Southeast Asian demonstrated their restricted distinctive ancestry in India mainly running along the foothills of Himalaya and northeastern part.
- ^ Chaubey, Gyaneshver; Kadian, Anurag; Bala, Saroj; Rao, Vadlamudi Raghavendra (10 June 2015). "Genetic Affinity of the Bhil, Kol and Gond Mentioned in Epic Ramayana". PLOS ONE. 10 (6): e0127655. Bibcode:2015PLoSO..1027655C. doi:10.1371/journal.pone.0127655. ISSN 1932-6203. PMC 4465503. PMID 26061398. S2CID 848806.
- ^ a b v d e Kivisild T, Papiha SS, Rootsi S, Parik J, Kaldma K, Reidla M, et al. (2000), An Indian Ancestry: a Key for Understanding Human Diversity in Europe and Beyond (PDF), McDonald Institute Monographs, arxivlandi (PDF) asl nusxasidan 2006 yil 19 fevralda, olingan 11 noyabr 2005
- ^ a b v "Y Haplogroups of the World, 2005, McDonald" (PDF). Arxivlandi asl nusxasi (PDF) 2004 yil 28 iyulda. Olingan 24 oktyabr 2006.
- ^ a b v d e f g h men j k l m n Sengupta S, Jivotovskiy LA, King R, Mehdi SQ, Edmonds CA, Chow CE va boshq. (2006 yil fevral). "Hindistondagi yuqori aniqlikdagi y-xromosoma tarqalishining qutbliligi va vaqtliligi mahalliy va ekzogen kengayishlarni aniqlaydi va Markaziy Osiyo chorvadorlarining ozgina genetik ta'sirini ochib beradi". Amerika inson genetikasi jurnali. 78 (2): 202–21. doi:10.1086/499411. PMC 1380230. PMID 16400607.
- ^ a b v d e f g h Sahoo S, Singh A, Himabindu G, Banerji J, Sitalaximi T, Gaikvad S va boshq. (January 2006). "Hindistondagi Y xromosomalari tarixi: demik diffuziya stsenariylarini baholash". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 103 (4): 843–8. Bibcode:2006 yil PNAS..103..843S. doi:10.1073 / pnas.0507714103. PMC 1347984. PMID 16415161.
- ^ a b v d e f Thanseem I, Thangaraj K, Chaubey G, Singh VK, Bhaskar LV, Reddy BM, et al. (2006 yil avgust). "Hindistonning quyi kastalari va qabilaviy guruhlari o'rtasidagi genetik yaqinlik: Y xromosoma va mitoxondriyal DNK xulosasi". BMC Genetika. 7: 42. doi:10.1186/1471-2156-7-42. PMC 1569435. PMID 16893451.
- ^ a b v Zhao Z, Khan F, Borkar M, Herrera R, Agrawal S (2009). "Presence of three different paternal lineages among North Indians: a study of 560 Y chromosomes". Inson biologiyasi yilnomalari. 36 (1): 46–59. doi:10.1080/03014460802558522. PMC 2755252. PMID 19058044.
- ^ Endicott, Metspalu & Kivisild 2007, p. 231.
- ^ Endicott, Metspalu & Kivisild 2007, 234–235 betlar.
- ^ Das, Ranajit; Upadhyai, Priyanka (25 November 2016). "Tracing the biogeographical origin of South Asian populations using DNA SatNav". bioRxiv: 089466. doi:10.1101/089466. S2CID 88966532.
Our hypothesis is supported by archaeological, linguistic and genetic evidences that suggest that there were two prominent waves of immigrations to India. A majority of the Early Caucasoids were proto-Dravidian language speakers that migrated to India putatively ~ 6000 YBP.
- ^ Oppenheimer 2003[sahifa kerak ]
- ^ a b Puente XS, Velasco G, Gutiérrez-Fernández A, Bertranpetit J, King MC, López-Otín C (January 2006). "Comparative analysis of cancer genes in the human and chimpanzee genomes". BMC Genomics. 7: 15. doi:10.1186/1471-2164-7-15. PMC 1382208. PMID 16438707.
- ^ a b v Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (2004 yil avgust). "Janubiy va janubi-g'arbiy Osiyodagi mtDNA chegaralarining aksariyati anatomik jihatdan zamonaviy odamlar tomonidan Evroosiyoning dastlabki joylashuvi paytida shakllangan bo'lishi mumkin". BMC Genetika. 5: 26. doi:10.1186/1471-2156-5-26. PMC 516768. PMID 15339343.
- ^ Kivisild, T .; Bamshad, M.J.; Kaldma, K .; Metspalu, M .; Metspalu, E .; Reyda, M.; Laos, S.; Parik, J .; Watkins, W.S.; Dixon, M.E.; Papiha, S.S.; Mastana, S.S.; Mir, M.R.; Ferak, V.; Villems, R. (November 1999). "Deep common ancestry of Indian and western-Eurasian mitochondrial DNA lineages". Hozirgi biologiya. 9 (22): 1331–1334. doi:10.1016/s0960-9822(00)80057-3. PMID 10574762. S2CID 2821966.
- ^ a b "Y-DNA Haplogroup H and its Subclades – 2015". Arxivlandi asl nusxasidan 2015 yil 1-noyabrda. Olingan 11 oktyabr 2015.
- ^ a b v d e f Cordaux R, Aunger R, Bentley G, Nasidze I, Sirajuddin SM, Stoneking M (February 2004). "Hind kastasi va urug 'urug' nasablarining mustaqil kelib chiqishi". Hozirgi biologiya. 14 (3): 231–5. doi:10.1016 / j.cub.2004.01.024. PMID 14761656. S2CID 5721248.
- ^ a b v Singh, Sakshi; Singx, Ashish; Rajkumar, Raja; Sampath Kumar, Katakam; Kadarkarai Samy, Subburaj; Nizamuddin, Sheikh; Singh, Amita; Ahmed Sheikh, Shahnawaz; Peddada, Vidya; Khanna, Vinee; Veeraiah, Pandichelvam; Pandit, Aridaman; Chaubey, Gyaneshver; Singx, Lalji; Thangaraj, Kumarasamy (12 January 2016). "Dissecting the influence of Neolithic demic diffusion on Indian Y-chromosome pool through J2-M172 haplogroup". Ilmiy ma'ruzalar. 6 (1): 19157. Bibcode:2016 yil NatSR ... 619157S. doi:10.1038 / srep19157. ISSN 2045-2322. PMC 4709632. PMID 26754573.
- ^ Errera, Rene J.; Garcia-Bertrand, Ralph (2018). Ota-bobolarning DNKsi, odamning kelib chiqishi va migratsiyasi. Akademik matbuot. p. 250. ISBN 978-0-12-804128-4.
- ^ Arunkumar G, Soria-Hernanz DF, Kavitha VJ, Arun VS, Syama A, Ashokan KS, et al. (2012). "Population differentiation of southern Indian male lineages correlates with agricultural expansions predating the caste system". PLOS ONE. 7 (11): e50269. Bibcode:2012PLoSO...750269A. doi:10.1371/journal.pone.0050269. PMC 3508930. PMID 23209694.
- ^ a b v d e Sharma S, Rai E, Sharma P, Jena M, Singh S, Darvishi K, et al. (Yanvar 2009). "The Indian origin of paternal haplogroup R1a1* substantiates the autochthonous origin of Brahmins and the caste system". Inson genetikasi jurnali. 54 (1): 47–55. doi:10.1038 / jhg.2008.2. PMID 19158816. S2CID 22162114.
- ^ a b v Qamar R, Ayub Q, Mohyuddin A, Helgason A, Mazhar K, Mansoor A, et al. (2002 yil may). "Y-chromosomal DNA variation in Pakistan". Amerika inson genetikasi jurnali. 70 (5): 1107–24. doi:10.1086/339929. PMC 447589. PMID 11898125.
- ^ Shah AM, Tamang R, Moorjani P, Rani DS, Govindaraj P, Kulkarni G, et al. (2011 yil iyul). "Indian Siddis: African descendants with Indian admixture". Amerika inson genetikasi jurnali. 89 (1): 154–61. doi:10.1016 / j.ajhg.2011.05.030. PMC 3135801. PMID 21741027.
- ^ a b v d e f g h men j k Kivisild (2003)
- ^ a b v Pijpe, Xeren; de Voogt, Alex; van Oven, Mannis; Henneman, Peter; van der Gaag, Kristiaan J.; Kayser, Manfred; de Knijff, Peter (21 March 2013). "Indian ocean crossroads: Human genetic origin and population structure in the maldives". Amerika jismoniy antropologiya jurnali. 151 (1): 58–67. doi:10.1002/ajpa.22256. ISSN 0002-9483. PMC 3652038. PMID 23526367.
- ^ Uells, Spenser (2007). Chuqur ajdodlar: Genografik loyiha ichida. Vashington, Kolumbiya: National Geographic. ISBN 978-1426201189.
- ^ a b Mahal DG, Matsoukas IG (20 September 2017). "Y-STR Gaplogroupining Jat populyatsiyasidagi xilma-xilligi bir necha xil qadimiy kelib chiqishlarini ochib beradi". Genetika chegaralari. 8: 121. doi:10.3389 / fgene.2017.00121. PMC 5611447. PMID 28979290.
- ^ Spencer Wells (2003), The Journey of Man. A Genetic Odyssey. New Delhi: Penguin Books India, p. 167
- ^ Zhao Z, Khan F, Borkar M, Herrera R, Agrawal S (2009). "Presence of three different paternal lineages among North Indians: a study of 560 Y chromosomes". Inson biologiyasi yilnomalari. 36 (1): 46–59. doi:10.1080/03014460802558522. PMC 2755252. PMID 19058044.
- ^ McElreavey K, Quintana-Murci L (2005). "Hindiston vodiysining populyatsion genetik nuqtai nazari, yagona meros qilib olingan mermerlar orqali". Inson biologiyasi yilnomalari. 32 (2): 154–62. doi:10.1080/03014460500076223. PMID 16096211. S2CID 109014.
- ^ Thangaraj K, Naidu BP, Crivellaro F, Tamang R, Upadhyay S, Sharma VK, et al. (2010 yil dekabr). "The influence of natural barriers in shaping the genetic structure of Maharashtra populations". PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO ... 515283T. doi:10.1371 / journal.pone.0015283. PMC 3004917. PMID 21187967.
- ^ Thangaraj K, Naidu BP, Crivellaro F, Tamang R, Upadhyay S, Sharma VK, et al. (2010 yil dekabr). Cordaux R (ed.). "The influence of natural barriers in shaping the genetic structure of Maharashtra populations". PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO ... 515283T. doi:10.1371 / journal.pone.0015283. PMC 3004917. PMID 21187967.
- ^ a b v d Basu A, Mukherjee N, Roy S, Sengupta S, Banerjee S, Chakraborty M, et al. (2003 yil oktyabr). "Ethnic India: a genomic view, with special reference to peopling and structure". Genom tadqiqotlari. 13 (10): 2277–90. doi:10.1101 / gr.1413403. PMC 403703. PMID 14525929.CS1 maint: ref = harv (havola)
- ^ a b v d Firasat S, Khaliq S, Mohyuddin A, Papaioannou M, Tyler-Smith C, Underhill PA, Ayub Q (January 2007). "Pokistonning Patan aholisiga cheklangan yunonlarning hissasi to'g'risida Y-xromosoma dalillari". Evropa inson genetikasi jurnali. 15 (1): 121–6. doi:10.1038 / sj.ejhg.5201726. PMC 2588664. PMID 17047675.CS1 maint: ref = harv (havola)
- ^ Qamar 2002 yil.
- ^ Mcelreavey 2005.
- ^ a b Kivisild T, Rootsi S, Metspalu M, Metspalu E, Parik J, Kaldma K, Usanga E, Mastana S, Papiha SS, Villems R (2003). "The Genetics of Language and Farming Spread in India" (PDF). In Bellwood P, Renfrew C (ed.). Dehqonchilik / til tarqalishi gipotezasini o'rganish. McDonald Institute for Archaeological Research, Cambridge, United Kingdom. 215-222 betlar.
- ^ a b v Sengupta va boshq. (2005)[to'liq iqtibos kerak ]
- ^ a b v d e Underhill PA, Myres NM, Rootsi S, Metspalu M, Jivotovskiy LA, King RJ va boshq. (2010 yil aprel). "R1a haplogroupi tarkibidagi Evropa va Osiyo Y xromosomalarining muzlikdan keyingi koankestrini ajratish". Evropa inson genetikasi jurnali. 18 (4): 479–84. doi:10.1038 / ejhg.2009.194. PMC 2987245. PMID 19888303.CS1 maint: ref = harv (havola)
- ^ Mirabal S, Regueiro M, Cadenas AM, Cavalli-Sforza LL, Underhill PA, Verbenko DA, et al. (Oktyabr 2009). "Y-chromosome distribution within the geo-linguistic landscape of northwestern Russia". Evropa inson genetikasi jurnali. 17 (10): 1260–73. doi:10.1038 / ejhg.2009.6. PMC 2986641. PMID 19259129.
- ^ a b v Underhill 2014.
- ^ a b v Pamjav 2012 yil.
- ^ ArunkumarG 2012.
- ^ Fornarino va boshq. (2009)
- ^ Manoukian, Jean-Grégoire (2006), "A Synthesis of Haplogroup R2 – 2006 Arxivlandi 1 oktyabr 2015 da Orqaga qaytish mashinasi."
- ^ Mondal, Mayux; Bergstrem, Anders; Syu, Yali; Kalafell, Franchesk; Laayuni, Xofid; Kasals, Ferran; Majumder, Parfa P.; Tayler-Smit, Kris; Bertranpetit, Jaume (1 May 2017). "Turli hind populyatsiyalarining Y-xromosoma ketma-ketliklari va Andamanliklarning ajdodlari". Inson genetikasi. 136 (5): 499–510. doi:10.1007 / s00439-017-1800-0. hdl:10230/34399. ISSN 1432-1203. PMID 28444560. S2CID 3725426.
- ^ a b Kumar V, Reddy AN, Babu JP, Rao TN, Langstieh BT, Thangaraj K, et al. (2007 yil mart). "Y-xromosoma dalillari Avstriya-Osiyo populyatsiyasining umumiy ota merosidan dalolat beradi". BMC evolyutsion biologiyasi. 7: 47. doi:10.1186/1471-2148-7-47. PMC 1851701. PMID 17389048.
- ^ Eaaswarkhanth M, Haque I, Ravesh Z, Romero IG, Meganathan PR, Dubey B, et al. (2010 yil mart). "Hindiston musulmonlari populyatsiyasida Saxara va Yaqin Sharq nasablari izlari". Evropa inson genetikasi jurnali. 18 (3): 354–63. doi:10.1038 / ejhg.2009.168. PMC 2859343. PMID 19809480.
- ^ "The Place of the Indian mtDNA Variants in the Global Network of Maternal Lineages and the Peopling of the Old World" (PDF). Arxivlandi (PDF) asl nusxasidan 2012 yil 8 martda. Olingan 28 sentyabr 2011.
- ^ "Ethnologue report for Indo-European". Ethnologue.com. Arxivlandi asl nusxasidan 2012 yil 15 oktyabrda. Olingan 24-noyabr 2015.
- ^ Baldi P (1990). Linguistic Change and Reconstruction Methodology. Valter de Gruyter. p. 342. ISBN 978-3-11-011908-4.
- ^ Burling (2003), 174–178 betlar.
- ^ Bradley (2012) notes, MK in the wider sense including the Munda languages of eastern South Asia is also known as Austroasiatic.Languages and Language Families in China Arxivlandi 2017 yil 30-aprel kuni Orqaga qaytish mashinasi
- ^ Bamshad M, Kivisild T, Watkins WS, Dixon ME, Ricker CE, Rao BB, et al. (Iyun 2001). "Genetic evidence on the origins of Indian caste populations". Genom tadqiqotlari. 11 (6): 994–1004. doi:10.1101/gr.GR-1733RR. PMC 311057. PMID 11381027.
- ^ Mukherjee N, Nebel A, Oppenheim A, Majumder PP (December 2001). "High-resolution analysis of Y-chromosomal polymorphisms reveals signatures of population movements from Central Asia and West Asia into India". Genetika jurnali. 80 (3): 125–35. doi:10.1007 / BF02717908. PMID 11988631. S2CID 13267463.
- ^ Mondal, Mayux; Bergstrem, Anders; Syu, Yali; Kalafell, Franchesk; Laayuni, Xofid; Kasals, Ferran; Majumder, Parfa P.; Tayler-Smit, Kris; Bertranpetit, Jaume (25 April 2017). "Turli hind populyatsiyalarining Y-xromosoma ketma-ketliklari va Andamanliklarning ajdodlari". Inson genetikasi. 136 (5): 499–510. doi:10.1007 / s00439-017-1800-0. hdl:10230/34399. PMID 28444560. S2CID 3725426.
- ^ Reich D, Thangaraj K, Patterson N, Price AL, Singh L (September 2009). "Reconstructing Indian population history". Tabiat. 461 (7263): 489–94. Bibcode:2009Natur.461..489R. doi:10.1038/nature08365. PMC 2842210. PMID 19779445.
- ^ Moorjani P, Thangaraj K, Patterson N, Lipson M, Loh PR, Govindaraj P, et al. (2013 yil sentyabr). "Genetic evidence for recent population mixture in India". Amerika inson genetikasi jurnali. 93 (3): 422–38. doi:10.1016/j.ajhg.2013.07.006. PMC 3769933. PMID 23932107.
- ^ a b Chaubey, Gyaneshver; Endicott, Phillip (June 2013). "The Andaman Islanders in a Regional Genetic Context: Reexamining the Evidence for an Early Peopling of the Archipelago from South Asia". Inson biologiyasi. 85 (1–3): 153–172. doi:10.3378/027.085.0307. PMID 24297224. S2CID 7774927.
- ^ Majumder PP (February 2010). "The human genetic history of South Asia". Hozirgi biologiya. 20 (4): R184-7. doi:10.1016/j.cub.2009.11.053. PMID 20178765. S2CID 1490419.
- ^ Iqtibos xatosi. Qanday tuzatish kerakligini ichki sharhga qarang.[tekshirish kerak ]
- ^ Pathak, Ajai K.; Kadian, Anurag; Kushniarevich, Alena; Montinaro, Francesco; Mondal, Mayux; Ongaro, Linda; Singx, Manvendra; Kumar, Pramod; Ray, Niraj; Parik, Juri; Metspalu, Ene; Rootsi, Siiri; Pagani, Luka; Kivisild, Toomas; Metspalu, Mait; Chaubey, Gyaneshver; Villems, Richard (December 2018). "The Genetic Ancestry of Modern Indus Valley Populations from Northwest India". Amerika inson genetikasi jurnali. 103 (6): 918–929. doi:10.1016/j.ajhg.2018.10.022. PMC 6288199. PMID 30526867.
- ^ Reyx, Devid (2018). Who We Are and How We Got Here: Ancient DNA and the new science of the human past. Oksford. ISBN 978-0-19-257040-6. Olingan 2 mart 2020.
- ^ a b v d Narasimxon va boshq. 2018 yil, p. 15.
- ^ Shinde V, Narasimhan VM, Rohland N, Mallick S, Mah M, Lipson M, Nakatsuka N, Adamski N, Broomandkhoshbacht N, Ferry M, Lawson AM, Michel M, Oppenheimer J, Stewardson K, Jadhav N, Kim YJ, Chatterjee M, Munshi A, Panyam A, Waghmare P, Yadav Y, Patel H, Kaushik A, Thangaraj K, Meyer M, Patterson N, Rai N, Reich D (September 2019). "An Ancient Harappan Genome Lacks Ancestry from Steppe Pastoralists or Iranian Farmers". Hujayra. 179 (3): 729–735.e10. doi:10.1016/j.cell.2019.08.048. PMC 6800651. PMID 31495572.
- ^ Narasimhan VM, Patterson N, Moorjani P, Rohland N, Bernardos R, Mallick S, et al. (Sentyabr 2019). "Janubiy va Markaziy Osiyoda odam populyatsiyasining shakllanishi". Ilm-fan. 365 (6457): eaat7487. doi:10.1126 / science.aat7487. PMC 6822619. PMID 31488661.
- ^ Watkins WS, Prasad BV, Naidu JM, Rao BB, Bhanu BA, Ramachandran B, et al. (2005 yil noyabr). "Diversity and divergence among the tribal populations of India". Inson genetikasi yilnomalari. 69 (Pt 6): 680–92. doi:10.1046/j.1529-8817.2005.00200.x. PMID 16266407. S2CID 31907598.
- ^ Reddy BM, Naidu VM, Madhavi VK, Thangaraj LK, Kumar V, Langstieh BT, et al. (2005 yil dekabr). "Microsatellite diversity in Andhra Pradesh, India: genetic stratification versus social stratification". Inson biologiyasi. 77 (6): 803–23. doi:10.1353/hub.2006.0018. PMID 16715839. S2CID 18446485.
- ^ Vishwanathan H, Deepa E, Cordaux R, Stoneking M, Usha Rani MV, Majumder PP (March 2004). "Genetic structure and affinities among tribal populations of southern India: a study of 24 autosomal DNA markers". Inson genetikasi yilnomalari. 68 (Pt 2): 128–38. doi:10.1046/j.1529-8817.2003.00083.x. PMID 15008792. S2CID 24230856.
- ^ "The Y-chromosome of the Soliga, an ancient forest-dwelling tribe of South India". Gene: X. 5: 100026. 1 December 2020. doi:10.1016/j.gene.2019.100026. ISSN 2590-1583.
Qo'shimcha o'qish
- Allikas A, Ord D, Kurg R, Kivi S, Ustav M (June 2001). "Roles of the hinge region and the DNA binding domain of the bovine papillomavirus type 1 E2 protein in initiation of DNA replication". Viruslarni o'rganish. 75 (2): 95–106. doi:10.1016/S0168-1702(01)00219-2. PMID 11325464.CS1 maint: ref = harv (havola)
- Behar DM, Garrigan D, Kaplan ME, Mobasher Z, Rosengarten D, Karafet TM, et al. (2004 yil mart). "Ashkenazi yahudiylari va mezbon yahudiy bo'lmagan Evropa populyatsiyalaridagi Y xromosomalarining o'zgaruvchanligi". Inson genetikasi. 114 (4): 354–65. doi:10.1007 / s00439-003-1073-7. PMID 14740294. S2CID 10310338.
- Bhattacharyya NP, Basu P, Das M, Pramanik S, Banerjee R, Roy B, et al. (1999 yil avgust). "Negligible male gene flow across ethnic boundaries in India, revealed by analysis of Y-chromosomal DNA polymorphisms". Genom tadqiqotlari. 9 (8): 711–9. doi:10.1101/gr.9.8.711 (inactive 25 September 2020). PMID 10447506.CS1 maint: DOI 2020 yil sentyabr holatiga ko'ra faol emas (havola)
- Cann RL (March 2001). "Genetic clues to dispersal in human populations: retracing the past from the present". Ilm-fan. 291 (5509): 1742–8. Bibcode:2001Sci...291.1742C. doi:10.1126/science.1058948. PMID 11249820. S2CID 19367408.CS1 maint: ref = harv (havola)
- Cinnioğlu C, King R, Kivisild T, Kalfoğlu E, Atasoy S, Cavalleri GL va boshq. (2004 yil yanvar). "Anadolida qazib olinadigan Y-xromosoma haplotipi qatlamlari". Inson genetikasi. 114 (2): 127–48. doi:10.1007 / s00439-003-1031-4. PMID 14586639. S2CID 10763736.
- Das B, Chauhan PS, Seshadri M (October 2004). "Minimal sharing of Y-chromosome STR haplotypes among five endogamous population groups from western and southwestern India". Inson biologiyasi. 76 (5): 743–63. doi:10.1353/hub.2005.0003. PMID 15757245. S2CID 12835244.
- Hemphill BE, Christensen AF (3 November 1994). The Oxus Civilization as a Link between East and West: A Non-Metric Analysis of Bronze Age Bactrain Biological Affinities. Madison, Viskonsin. p. 13.CS1 maint: ref = harv (havola) (paper read at the South Asia Conference)
- Jobling MA, Tyler-Smith C (August 2003). "The human Y chromosome: an evolutionary marker comes of age". Tabiat sharhlari. Genetika. 4 (8): 598–612. doi:10.1038/nrg1124. PMID 12897772. S2CID 13508130.
- Kivisild T, Rootsi S, Metspalu M, Mastana S, Kaldma K, Parik J, et al. (2003 yil fevral). "Qadimgi ko'chmanchilarning genetik merosi hind qabilalari va kast populyatsiyalarida saqlanib qolgan". Amerika inson genetikasi jurnali. 72 (2): 313–32. doi:10.1086/346068. PMC 379225. PMID 12536373.CS1 maint: ref = harv (havola)
- Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (2004 yil avgust). "Janubiy va janubi-g'arbiy Osiyodagi mtDNA chegaralarining aksariyati anatomik jihatdan zamonaviy odamlar tomonidan Evroosiyoning dastlabki joylashuvi paytida shakllangan bo'lishi mumkin". BMC Genetika. 5: 26. doi:10.1186/1471-2156-5-26. PMC 516768. PMID 15339343.
- Patowary A, Purkanti R, Singh M, Chauhan RK, Bhartiya D, Dwivedi OP, et al. (2012 yil iyul). "Systematic analysis and functional annotation of variations in the genome of an Indian individual". Inson mutatsiyasi. 33 (7): 1133–40. doi:10.1002/humu.22091. PMID 22461382. S2CID 11466942.
- Rootsi S, Magri C, Kivisild T, Benuzzi G, Help H, Bermisheva M, et al. (2004 yil iyul). "Phylogeography of Y-chromosome haplogroup I reveals distinct domains of prehistoric gene flow in europe". Amerika inson genetikasi jurnali. 75 (1): 128–37. doi:10.1086/422196. PMC 1181996. PMID 15162323.
- Qamar R, Ayub Q, Mohyuddin A, Helgason A, Mazhar K, Mansoor A, et al. (2002 yil may). "Y-chromosomal DNA variation in Pakistan". Amerika inson genetikasi jurnali. 70 (5): 1107–24. doi:10.1086/339929. PMC 447589. PMID 11898125.
- Semino O, Magri C, Benuzzi G, Lin AA, Al-Zahery N, Battaglia V, et al. (2004 yil may). "Y-xromosoma haplogrouplari E va J ning kelib chiqishi, diffuziyasi va differentsiatsiyasi: Evropaning neolitlanishi va O'rta er dengizi mintaqasidagi keyinchalik migratsion hodisalar to'g'risida xulosalar". Amerika inson genetikasi jurnali. 74 (5): 1023–34. doi:10.1086/386295. PMC 1181965. PMID 15069642.
Manbalar
- Indian Genome Variation Consortium (April 2008). "Genetic landscape of the people of India: a canvas for disease gene exploration". Genetika jurnali. 87 (1): 3–20. doi:10.1007/s12041-008-0002-x. PMID 18560169. S2CID 21473349.
- Endicott P, Metspalu M, Kivisild T (2007). "Genetic evidence on modern human dispersals in South Asia: Y chromose and mitochondrial DNA perspectives". In Michael D. Petraglia, Bridget Allchin (eds.). Janubiy Osiyoda inson populyatsiyasining rivojlanishi va tarixi. Springer. 201-228 betlar. ISBN 978-1-4020-5561-4.
- Hemphill B, Lukacs J, Kennedy K (1991). "Biological Adaptations and Affinities of Bronze Age Harappans". In Meadow RH (ed.). Harappa excavations 1986–1990: a multidisciplinary approach to third millennium urbanism. pp. 137–82. ISBN 978-0-9629110-1-9.CS1 maint: ref = harv (havola)
- Kennedy KA (1984). "A Reassessment of the Theories of Racial Origins of the People of the Indus Valley Civilization from Recent Anthropological Data". In Kennedy KA, Possehl GL (eds.). Studies in the Archaeology and Palaeoanthropology of South Asia. Atlantic Highlands, NJ: Humanities Press. pp. 99–107.CS1 maint: ref = harv (havola)
- Kennedy KA (1995). "Have Aryans been identified in the prehistoric skeletal record from South Asia?". Jorj Erdosida (tahrir). The Indo-Aryans of Ancient South Asia. Valter de Gruyter. 49-54 betlar. ISBN 978-3-11-014447-5.
- Kivisild T (2000b). The origins of southern and western Eurasian populations: an mtDNA study (PDF). Tartu University, Estonia.CS1 maint: ref = harv (havola) (PhD)
- Mascarenhas DD, Raina A, Aston CE, Sanghera DK (2015). "Genetic and Cultural Reconstruction of the Migration of an Ancient Lineage". BioMed Research International. 2015: 651415. doi:10.1155/2015/651415. PMC 4605215. PMID 26491681.
- Narasimxan VM, Entoni D, Mallori J, Reyx D (2018). "Janubiy va Markaziy Osiyoning genomik shakllanishi". bioRxiv 10.1101/292581. doi:10.1101/292581. S2CID 89658279. Iqtibos jurnali talab qiladi
| jurnal =(Yordam bering) - Oppenheimer S (2003). The Real Eve: Modern Man's Journey out of Africa. Nyu-York: Kerrol va Graf nashriyotlari. ISBN 978-0-7867-1192-5.CS1 maint: ref = harv (havola)
- Pamjav (December 2012), "Brief communication: New Y-chromosome binary markers improve phylogenetic resolution within haplogroup R1a1", Amerika jismoniy antropologiya jurnali, 149 (4): 611–615, doi:10.1002 / ajpa.22167, PMID 23115110, S2CID 4820868
- Renfrew C, Boyle K, eds. (2000a). An Indian Ancestry: a Key for Understanding Human Diversity in Europe and Beyond (PDF). ISBN 978-1-902937-08-3.CS1 maint: ref = harv (havola)
- Underhill PA (2003). "Inferring human history: clues from Y-chromosome haplotypes". Kantitativ biologiya bo'yicha sovuq bahor porti simpoziumlari. 68: 487–93. doi:10.1101/sqb.2003.68.487. PMID 15338652. S2CID 15527457.
- Underhill PA, Poznik GD, Rootsi S, Järve M, Lin AA, Wang J, et al. (Yanvar 2015). "Y-xromosoma haplogroupi R1a ning filogenetik va geografik tuzilishi". Evropa inson genetikasi jurnali. 23 (1): 124–31. doi:10.1038 / ejhg.2014.50. PMC 4266736. PMID 24667786.
- Wells S (2003). Inson sayohati: Genetik Odisseya. Prinston universiteti matbuoti.
Tashqi havolalar
- Introduction to Haplogroups and Haplotypes, Mark A. Jobling, University of Leicester. [1]
- Journey of Man: Peopling of the World, Bradshaw Foundation, in association with Stiven Oppengeymer.
- Indian Genome Variation Database Genomika va integral biologiya instituti
- List of R2 frequency
- Maldiv orollari
| Sub-mavzular | |
|---|---|
| Genetika tarixi mintaqalar bo'yicha | |
| Populyatsiya genetikasi guruh bo'yicha |
|
| |