Lak operon - Lac operon
The laktoza operon (lac operon) - bu operon transport uchun zarur va metabolizm ning laktoza yilda E.coli va boshqa ko'plab narsalar ichak bakteriyalari. Garchi glyukoza ko'pgina bakteriyalar uchun afzal qilingan uglerod manbai hisoblanadi lak operon orqali glyukoza mavjud bo'lmaganda laktozani samarali hazm qilishga imkon beradi beta-galaktozidaza.[1] Genlarni tartibga solish ning lak operon aniq tushunilgan birinchi genetik tartibga solish mexanizmi edi, shuning uchun u eng yaxshi namunaga aylandi prokaryotik genlarni tartibga solish. Bu ko'pincha kirish qismida muhokama qilinadi molekulyar va uyali biologiya shu sababli darslar. Ushbu laktoza metabolizm tizimi tomonidan ishlatilgan Fransua Yakob va Jak Monod qanday qilib a biologik hujayra qaysi fermentni sintez qilishni biladi. Lak operonidagi ishlari ularga g'olib bo'ldi Fiziologiya bo'yicha Nobel mukofoti 1965 yilda.[1]
Bakterial operonlar polikistronik stenogrammalar bitta oqsil ishlab chiqarishga qodir mRNA stenogramma. Bunday holda, laktoza a sifatida talab qilinganida shakar bakteriya manbai, lak operonning uchta genini ifodalash va ularning keyingi oqsillarini tarjima qilish mumkin: lacZ, lacYva lacA. Ning gen mahsuloti lacZ bu b-galaktozidaza laktoza ajratadigan, a disaxarid, ichiga glyukoza va galaktoza. lacY kodlaydi Beta-galaktozid permeazasi, a membrana oqsili ichiga o'rnatilgan bo'ladi sitoplazmatik membrana ni yoqish uchun uyali transport hujayradan laktoza. Nihoyat, lacA kodlaydi Galaktozid atsetiltransferaza.


1: RNK polimeraza, 2: Repressor, 3: Promouter, 4: Operator, 5: Laktoza, 6: lacZ, 7: lacY, 8: lacA.
Laktoza mavjud bo'lmaganda yoki glyukoza kabi afzal energiya manbai mavjud bo'lganda fermentlarni ishlab chiqarish behuda bo'ladi. The lak operon hujayralar tomonidan kodlangan fermentlarni ishlab chiqarish uchun energiya sarflashini ta'minlash uchun ikki qismli boshqarish mexanizmidan foydalanadi lak operon faqat kerak bo'lganda.[2] Laktoza yo'q bo'lganda lak repressor, lacI, tomonidan kodlangan fermentlarning ishlab chiqarilishini to'xtatadi lak operon.[3] Lak repressor har doim ifoda etiladi, agar unga koordinatator bog'lanmasa. Boshqacha qilib aytganda, u faqat kichik molekulalarning koordinatori ishtirokida transkripsiyalanadi. Glyukoza mavjud bo'lganda katabolit faollashtiruvchi oqsil Fermentlarni ishlab chiqarish uchun zarur bo'lgan (CAP) harakatsiz bo'lib qoladi va EIIAGlc hujayra ichiga laktoza tashishining oldini olish uchun laktoza permeazasini o'chiradi. Ushbu ikki tomonlama boshqarish mexanizmi glyukoza va laktozaning ketma-ket ishlatilishini ikkita aniq o'sish bosqichida, ya'ni diauxie.
Tuzilishi

- The lak operon 3 dan iborat strukturaviy genlar va a targ'ibotchi, a terminator, regulyator va an operator. Uchta tuzilish genlari: lacZ, lacYva lacA.
- lacZ kodlaydi b-galaktozidaza (LacZ), hujayra ichidagi ferment bu yoriqlar disaxarid laktoza ichiga glyukoza va galaktoza.
- lacY kodlaydi Beta-galaktozid permeazasi (LacY), transmembran tarafdor bu nasoslar b-galaktozidlar laktoza, shu jumladan proton gradienti yordamida hujayra ichiga bir xil yo'nalishda. Permeaza hujayraning o'tkazuvchanligini oshiradi b-galaktozidlar.
- lacA kodlaydi b-galaktozid transatsetilaza (LacA), anni o'tkazadigan ferment atsetil guruhi atsetil-KoA dan b-galaktozidlarga qadar.
Faqat lacZ va lacY laktoza uchun zarur bo'lib ko'rinadi katabolizm.
Genetik nomenklatura
Uch harfli qisqartmalar, shu jumladan bakteriyalardagi fenotiplarni tavsiflash uchun ishlatiladi E. coli.
Bunga misollar:
- Lak (laktoza ishlatish qobiliyati),
- Uning (gistidin aminokislotasini sintez qilish qobiliyati)
- Mot (suzish harakati)
- SmR (antibiotikga qarshilik streptomitsin )
Lak holatida yovvoyi turdagi hujayralar Lakdir+ va laktozani uglerod va energiya manbai sifatida ishlatishga qodir, Lak esa− mutant hosilalari laktoza ishlata olmaydi. Xuddi shu uchta harf odatda ma'lum bir fenotipga aloqador genlarni belgilash uchun ishlatiladi (kichik harflar bilan, kursiv bilan), bu erda har bir turli genlar qo'shimcha harflar bilan qo'shimcha ravishda ajralib turadi. The lak fermentlarni kodlovchi genlar lacZ, lacYva lacA. To'rtinchi lak gen lacI, laktoza repressorini kodlash - "men" degan ma'noni anglatadi induktivlik.
Ularning birini farqlashi mumkin tizimli fermentlarni kodlovchi genlar va gen ekspressioniga ta'sir qiluvchi oqsillarni kodlovchi regulyativ genlar. Amaldagi foydalanish fenotipik nomenklaturani oqsillarga nisbatan kengaytiradi: shuning uchun LacZ oqsil mahsulotidir lacZ gen, b-galaktozidaza. Genlar bo'lmagan turli xil qisqa ketma-ketliklar, shuningdek, gen ekspressioniga ta'sir qiladi lak targ'ibotchi, lac p, va lak operator, lac o. Bu qat'iy standart foydalanish bo'lmasa-da, mutatsiyalar ta'sir qiladi lac o deb nomlanadi lac ov, tarixiy sabablarga ko'ra.
Tartibga solish
Maxsus boshqarish lak genlar mavjudligiga bog'liq substrat bakteriyalarga laktoza. Laktoza uglerod manbai sifatida mavjud bo'lmaganda, oqsillar bakteriya tomonidan ishlab chiqarilmaydi lak genlar an operon; ya'ni ular xromosomaga bevosita tutashgan bir yo'nalishga yo'naltirilgan va bitta transkripsiya qilingan polikistronik mRNK molekulasi. Barcha genlarning transkripsiyasi fermentning bog'lanishidan boshlanadi RNK polimeraza (RNAP), a DNK bilan bog'lovchi oqsil, ma'lum bir DNK bog'lanish joyiga bog'langan, targ'ibotchi, darhol yuqori oqim genlarning. RNK polimerazaning promotor bilan bog'lanishiga yordam beradi lager - bog'langan katabolit faollashtiruvchi oqsil (CAP, shuningdek, cAMP retseptorlari oqsili deb ham ataladi).[4] Biroq, lacI gen (tartibga soluvchi gen lak operon) RNAPni operon operatoriga ulanishini to'sadigan oqsil ishlab chiqaradi. Ushbu oqsilni faqat qachon olib tashlash mumkin alloaktoza unga bog'laydi va uni inaktiv qiladi. Tomonidan hosil bo'lgan oqsil lacI gen lak repressor sifatida tanilgan. Bu tartibga solishning turi lak operon salbiy induktsiya deb ataladi, ya'ni gen regulyator omil tomonidan o'chiriladi (lak repressor), agar ba'zi bir molekula (laktoza) qo'shilmasa. Borligi sababli lak repressor oqsili, o'rnini bosuvchi gen muhandislari lacZ boshqa genga ega bo'lgan gen eksperimental bakteriyalarni agarda mavjud bo'lgan laktoza bilan agarda o'sishi kerak bo'ladi. Agar shunday qilmasalar, ular ifoda etishga harakat qilayotgan gen ifoda etilmaydi, chunki repressor oqsili hali ham RNAPni promotorga bog'lanishini va genni transkripsiyasini to'xtatmoqda. Repressor chiqarilgandan so'ng, RNAP uchta genni ham transkripsiyalashga kirishadi (lacZYAmRNKga MRNA zanjiridagi uchta genning har biri o'ziga xos xususiyatga ega Shine-Dalgarno ketma-ketligi, shuning uchun genlar mustaqil ravishda tarjima qilinadi.[5] Ning DNK ketma-ketligi E. coli lak operon, lacZYA mRNA, va lacI genlar mavjud GenBank (ko'rinish).
Birinchi nazorat mexanizmi hujayra ichidagi laktoza ta'sirini tartibga soluvchi javobdir tartibga soluvchi oqsil deb nomlangan laktoza repressori ishlab chiqarishga xalaqit berish b-galaktozidaza laktoza yo'q bo'lganda. The lacI repressor uchun genlarni kodlash yaqin joylashgan lak operon va har doim ifoda etiladi (konstitutsiyaviy). Agar o'sish muhitida laktoza etishmayotgan bo'lsa, repressor boshlanishiga yaqin promotorning pastki qismida qisqa DNK ketma-ketligi bilan juda qattiq bog'lanadi. lacZ deb nomlangan lac operatori. Operator bilan bog'laydigan repressor RNAPning promotorga ulanishiga xalaqit beradi va shuning uchun LacZ va LacY kodlash mRNA juda past darajada amalga oshiriladi. Hujayralar laktoza ishtirokida o'stirilganda, laktozadan hosil bo'lgan laktoza metaboliti allolaktoza deb ataladi. lacZ gen, repressor bilan bog'lanib, allosterik siljishni keltirib chiqaradi. Shunday qilib o'zgartirilgan, repressor operator bilan bog'lana olmaydi va RNAP-ga transkripsiyasini o'tkazishga imkon beradi lak genlar va shu bilan kodlangan oqsillarning yuqori darajalariga olib keladi.
Ikkinchi nazorat mexanizmi glyukozaga javob bo'lib, u ishlatadi katabolit faollashtiruvchi oqsil (CAP) homodimeri glyukoza yo'q bo'lganda b-galaktosidaza ishlab chiqarishni sezilarli darajada oshiradi. Tsiklik adenozin monofosfat (cAMP) - bu tarqalish darajasi glyukoza bilan teskari proportsional bo'lgan signal molekulasi. U CAP bilan bog'lanadi, bu esa o'z navbatida CAPni CAP bog'lanish joyiga bog'lashga imkon beradi (quyida keltirilgan diagrammada chap tarafdagi promotorning tepasida 16 bp DNK ketma-ketligi, transkripsiyani boshlash joyining yuqorisida 60 bp).[6] bu DNK bilan bog'lanishda RNAPga yordam beradi. Glyukoza yo'q bo'lganda, cAMP kontsentratsiyasi yuqori va CAP-cAMP ning DNK bilan bog'lanishi b-galaktozidaza hosil bo'lishini sezilarli darajada oshiradi va hujayraga imkon beradi. gidroliz laktoza va ajralib chiqadigan galaktoza va glyukoza.
Yaqinda induktorni chiqarib tashlash ekspresiyasini blokirovka qilish uchun ko'rsatildi lak glyukoza mavjud bo'lganda operon. Glyukoza hujayra ichiga PEP ga bog'liq bo'lgan fosfotransferaza tizimi. Ning fosfat guruhi fosfoenolpiruvat umumiy PTS (fosfotransferaza tizimi) oqsillari HPr va EIA va glyukozaga xos bo'lgan PTS oqsillari EIIA dan tashkil topgan fosforillanish kaskadi orqali uzatiladi.Glc va EIIBGlc, EII glyukoza tashuvchisi sitoplazmik domeni. Glyukozani tashish uning EIIB tomonidan fosforillanishi bilan kechadiGlc, fosfat guruhini boshqa PTS oqsillaridan, shu jumladan EIIA dan tozalashGlc. EIIA ning fosforlanmagan shakliGlc ga bog'laydi lak permease va uni hujayraga laktoza olib kelishining oldini oladi. Shuning uchun ham glyukoza, ham laktoza mavjud bo'lsa, glyukoza tashilishi ning induktorining transportini bloklaydi lak operon.[7]
Repressor tuzilishi

The lak repressor bir xil subbirliklarga ega bo'lgan to'rt qismli oqsil, tetramer. Har bir bo'linmada DNK bilan bog'lanish qobiliyatiga ega bo'lgan spiral-burilish-spiral (HTH) motifi mavjud. Repressor bog'langan operator uchastkasi teskari takroriy simmetriya bilan DNK ketma-ketligi. Operatorning ikkita DNK yarim joylari birgalikda repressorning ikkita bo'linmasiga bog'lanadi. Repressorning boshqa ikkita bo'linmasi ushbu modelda hech narsa qilmasa ham, bu xususiyat ko'p yillar davomida tushunilmagan.
Oxir-oqibat ikkita qo'shimcha operator ishtirok etishi aniqlandi lak tartibga solish.[8] Bittasi (O3) O ning yuqorisida -90 bp atrofida yotadi1 oxirida lacI gen va boshqa (O2) O ning pastki qismida taxminan +410 bp1 ning dastlabki qismida lacZ. Ushbu ikkita sayt dastlabki ishda topilmadi, chunki ular ortiqcha funktsiyalarga ega va individual mutatsiyalar repressiyaga unchalik ta'sir qilmaydi. Ikkala O ga bitta mutatsiyalar2 yoki O3 faqat 2 dan 3 martagacha effektlarga ega. Biroq, ularning ahamiyati ikkala O da nuqsonli er-xotin mutant ekanligi bilan namoyon bo'ladi2 va O3 dramatik ravishda repressiya qilingan (taxminan 70 baravar).
Amaldagi modelda, lak repressor bir vaqtning o'zida ikkala asosiy operator O bilan bog'langan1 va ikkalasiga ham2 yoki O3. Aralashgan DNK kompleksdan chiqib ketadi. Ikkala kichik operatorlarning ortiqcha tabiati shundan dalolat beradiki, bu o'ziga xos halqalangan kompleks emas. Bitta fikr shundan iboratki, tizim bog'lash orqali ishlaydi; agar bog'langan repressor O dan chiqsa1 bir lahzada, kichik operator uchun majburiylik uni tezda qayta tiklanishi uchun uni yaqin joyda saqlaydi. Bu repressorning O ga yaqinligini oshiradi1.
Induksiya mexanizmi

Repressor - bu allosterik oqsil, ya'ni u bir-birlari bilan muvozanat holatida bo'lgan bir-biridan biroz farq qiladigan ikkita shakldan birini qabul qilishi mumkin. Bir shaklda repressor yuqori aniqlik bilan operator DNK bilan bog'lanadi, boshqa shaklda esa u o'ziga xosligini yo'qotadi. Klassik induksiya modeliga ko'ra induktorning yoki allolakaktoza yoki IPTGni repressor bilan bog'lashi repressorning ikki shakl o'rtasida taqsimlanishiga ta'sir qiladi. Shunday qilib, induktor bog'langan repressor DNK bilan bog'lanmaydigan konformatsiyada barqarorlashadi. Biroq, bu oddiy model butun voqea bo'lishi mumkin emas, chunki repressor DNK bilan barqaror ravishda bog'langan, ammo induktor qo'shilishi bilan tezda ajralib chiqadi. Shuning uchun, agar induktor repressor bilan DNKga bog'langan bo'lsa, u ham repressor bilan bog'lanishi aniq ko'rinadi. Bog'lanishning aniq mexanizmi nima ekanligi hali ham to'liq ma'lum emas.[9]
Maxsus bo'lmagan majburiylikning roli
Repressorning DNK bilan o'ziga xos bo'lmagan birikishi Lak-operonning repressiyasi va induktsiyasida hal qiluvchi rol o'ynaydi. Lak-repressor oqsilining o'ziga xos bog'lanish joyi operator hisoblanadi. Xususiy bo'lmagan o'zaro ta'sir asosan zaryad-zaryadlarning o'zaro ta'siri orqali amalga oshiriladi, operator bilan bog'lanish esa hidrofobik o'zaro ta'sirlar bilan mustahkamlanadi. Bundan tashqari, repressor bog'lanishi mumkin bo'lgan o'ziga xos bo'lmagan DNK sekanslari ko'pligi. Aslida, operator bo'lmagan har qanday ketma-ketlik o'ziga xos bo'lmagan hisoblanadi. Tadqiqotlar shuni ko'rsatdiki, o'ziga xos bo'lmagan bog'lanish mavjud bo'lmaganda, Lak-operonning induksiyasi (yoki repressiyasi) to'yingan induktor darajasida ham sodir bo'lishi mumkin emas. Maxsus bog'lamasdan induksiyaning bazal darajasi odatdagidan o'n ming marta kichik ekanligi isbotlangan. Buning sababi shundaki, o'ziga xos bo'lmagan DNK repressor oqsillari uchun "cho'kma" vazifasini bajaradi va ularni operatordan chalg'itadi. Maxsus bo'lmagan ketma-ketliklar hujayradagi mavjud repressor miqdorini kamaytiradi. Bu o'z navbatida tizimni qayta ochish uchun zarur bo'lgan induktor miqdorini kamaytiradi.[10]
Laktoza analoglari

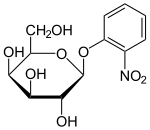
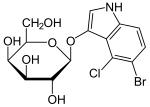

Lakozli operon bilan ishlash uchun foydali bo'lgan bir qator laktoza hosilalari yoki analoglari tasvirlangan. Ushbu birikmalar asosan o'rnini bosuvchi galaktozidlar bo'lib, bu erda laktozaning glyukoza qismi boshqa kimyoviy guruh bilan almashtiriladi.
- Izopropil-b-D-tiogalaktopiranozid (IPTG) ning induktori sifatida tez-tez ishlatiladi lak fiziologik ish uchun operon.[1] IPTG repressor bilan bog'lanadi va uni inaktiv qiladi, ammo b-galaktozidaza uchun substrat emas. IPTG-ning bitta afzalligi jonli ravishda tadqiqotlar shundan iboratki, uni metabolizm qilish mumkin emas E. coli uning kontsentratsiyasi doimiy bo'lib qoladi va ifoda tezligi lac p / o- boshqariladigan genlar, tajribada o'zgaruvchan emas. IPTGni qabul qilish laktoza permeaza ta'siriga bog'liq P. flüoresanlar, lekin emas E. coli.[11]
- Fenil-b-D-galaktoza (fenil-Gal) b-galaktozidaza uchun substrat, ammo repressorni inaktiv qilmaydi va shuning uchun induktor ham emas. Yovvoyi tipdagi hujayralar b-galaktozidazani juda kam hosil qilganligi sababli, ular fenil-Galda uglerod va energiya manbai sifatida o'sishi mumkin emas. Repressorga ega bo'lmagan mutantlar fenil-Galda o'sishga qodir. Shunday qilib, uglerod va energiya manbai sifatida faqat fenil-Gal bo'lgan minimal vosita repressor mutantlari va operator mutantlari uchun tanlab olinadi. Agar 10 bo'lsa8 yovvoyi turdagi shtamm hujayralari fenil-Gal o'z ichiga olgan agar plastinkalarida qoplanadi, o'sib chiqadigan noyob koloniyalar asosan repressorga ta'sir qiluvchi spontan mutantlardir. Repressor va operator mutantlarining nisbiy taqsimlanishiga maqsad hajmi ta'sir qiladi. Beri lacI geni kodlash repressori operatordan taxminan 50 baravar katta, tanlovda repressor mutantlari ustunlik qiladi.
- Tiometil galaktozidaza [TMG] boshqa laktoza analogidir. Ular lacI repressorini inhibe qiladi. Past induktor kontsentratsiyasida ham TMG, ham IPTG hujayralarga laktoza permeaza orqali kirib borishi mumkin. Ammo induktorning yuqori konsentratsiyasida ikkala analog ham hujayraga mustaqil ravishda kirishi mumkin. TMG hujayradan tashqari yuqori konsentrasiyalarda o'sish sur'atlarini pasaytirishi mumkin.[12]
- Boshqa birikmalar b-galaktozidaza faolligining rangli ko'rsatkichlari bo'lib xizmat qiladi.
- ONPG qattiq sariq birikma hosil qilish uchun ajratilgan, ortonitrofenol va galaktoza va odatda b-galaktozidazani tahlil qilish uchun substrat sifatida ishlatiladi in vitro.[13]
- B-galaktozidaza ishlab chiqaradigan koloniyalar ko'k rangga aylanadi X-gal (5-bromo-4-xloro-3-indolil-b-D-galaktozid), bu B-galaktozidaza uchun sun'iy substrat bo'lib, uning parchalanishi natijasida galaktoza va 4-Cl, 3-Br indigo hosil bo'lib, quyuq ko'k rang hosil qiladi.[14]
- Alloaktoza bu izomer laktoza va lak operonining induktoridir. Laktoza - galaktoza- (-1-> 4) -glyukoza, allolakaktoza esa galaktoza- (-1-> 6) -glyukoza. Laktoza gidrolitikka alternativ reaktsiyada b-galaktozidaza bilan allolaktozaga aylanadi. LacZ ning "haqiqiy" induktorni ishlab chiqarishdagi rolini ko'rsatadigan fiziologik tajriba E. coli hujayralar - bu nol mutant bo'lgan kuzatuv lacZ allolaktozaning gidrolizlanmaydigan analogi bo'lgan IPTG bilan o'stirilganda LacY permeaza hosil qilishi mumkin, ammo laktoza bilan o'stirilmasa. Tushuntirish shundan iboratki, hujayra ichidagi induktorni hosil qilish uchun laktozani allolaktozaga (b-galaktozidaza tomonidan katalizlangan) qayta ishlash kerak.
Klassik modelni ishlab chiqish
Eksperimental mikroorganizm tomonidan ishlatilgan Fransua Yakob va Jak Monod umumiy laboratoriya bakteriyasi edi, E. coli, lekin Yoqub va Monod tomonidan kashf etilgan ko'plab asosiy tartibga soluvchi tushunchalar uchun muhimdir uyali barcha organizmlarda tartibga solish.[15] Asosiy g'oya shundaki, oqsillar kerak bo'lmaganda sintez qilinmaydi.E. coli Laktozani metabolizmga ehtiyoj qolmaganda, masalan, boshqa shakarlarda bo'lgani kabi, uchta Lak oqsilini hosil qilmasdan hujayra resurslari va energiyasini tejaydi. glyukoza mavjud. Quyidagi bo'limda qanday qilib muhokama qilinadi E. coli metabolik ehtiyojlarga javoban ba'zi genlarni boshqaradi.
Davomida Ikkinchi jahon urushi, Monod shakarlarning kombinatsiyasi ta'sirini ozuqa manbalari sifatida sinab ko'rdi E. coli va B. subtilis. Monod boshqa olimlar tomonidan bakteriya va xamirturush bilan o'tkazilgan shunga o'xshash tadqiqotlarni kuzatib bordi. U ikki xil shakar bilan o'sgan bakteriyalar ko'pincha o'sishning ikki bosqichini ko'rsatishini aniqladi. Masalan, glyukoza va laktoza ikkalasi ham ta'minlangan bo'lsa, avval glyukoza metabolizmga uchragan (o'sish bosqichi I, 2-rasmga qarang), so'ngra laktoza (o'sish bosqichi II). Dioksik o'sish egri chizig'ining birinchi qismida laktoza metabolizmga uchramadi, chunki muhitda glyukoza va laktoza mavjud bo'lganda b-galaktozidaza hosil bo'lmadi. Monod ushbu hodisani nomladi diauxie.[16]

Keyin Monod diqqatini laktoza madaniy muhitda yagona shakar bo'lganida yuzaga kelgan b-galaktozidaza hosil bo'lishining induktsiyasiga qaratdi.[17]
Regulyatsion mutantlarning tasnifi
Yoqub va Monodning kontseptual yutug'i[18] tartibga soluvchi moddalar va ular gen ekspressionini o'zgartirish uchun harakat qiladigan joylar o'rtasidagi farqni tan olish edi. Sobiq askar Yoqub maxsus radioeshittirish yoki signal olgandan keyin halokatli yukini bo'shatib yuboradigan bombardimonchining o'xshashidan foydalangan. Ishlaydigan tizim samolyotda erga uzatuvchi va qabul qiluvchiga ham ehtiyoj sezadi. Endi odatdagi transmitter buzilgan deb taxmin qiling. Ushbu tizimni ikkinchi, ishlaydigan transmitterni ishga tushirish orqali amalga oshirish mumkin. Aksincha, dedi u, qabul qiluvchisi nuqsoni bo'lgan bombardimonchini ko'rib chiqing. Ning xatti-harakati bu bombardimonchi samolyotni ikkinchi marta ishlab chiqarish bilan o'zgartirish mumkin emas.
Ning regulyatsion mutantlarini tahlil qilish lak operon, Jeykob tizimning ikkinchi nusxasini yaratgan tizim yaratdi lak genlar (lacI uning promouteri bilan va lacZYA promouter va operator bilan) bitta katakka kiritilishi mumkin. Uchun diploid bo'lgan bunday bakteriyalar madaniyati lak genlar, ammo boshqacha normal bo'lsa, keyinchalik tartibga soluvchi fenotip uchun sinovdan o'tkaziladi. Xususan, LacZ va LacY IPTG bo'lmagan taqdirda ham ishlab chiqarilganligi aniqlanadi (tufayli laktoza repressori mutant geni tomonidan ishlab chiqilgan). Genlar yoki gen klasterlari juft sinovdan o'tkaziladigan ushbu tajriba a deb nomlanadi komplementatsiya testi.

Ushbu test rasmda ko'rsatilgan (lacA soddaligi uchun chiqarib tashlangan). Birinchidan, ba'zi gaploid holatlar ko'rsatilgan (ya'ni hujayra faqat bitta nusxasini olib yuradi lak genlar). Panel (a) repressiyani, (b) IPTG tomonidan induksiyani va (c) va (d) mutatsiyaning ta'sirini ko'rsatadi lacI navbati bilan gen yoki operatorga. Panelning (e) qismida repressor uchun komplementatsiya testi ko'rsatilgan. Agar bitta nusxasi bo'lsa lak genlar mutatsiyaga ega lacI, lekin ikkinchi nusxasi yovvoyi turi uchun lacI, hosil bo'lgan fenotip normaldir, ammo lacZ induktor IPTG ta'sirida namoyon bo'ladi. Repressorga ta'sir qiluvchi mutatsiyalar deyiladi retsessiv yovvoyi turga (va bu yovvoyi tur dominant) va bu repressor hujayrada tarqalishi mumkin bo'lgan kichik oqsil ekanligi bilan izohlanadi. Nusxasi lak nuqsonga qo'shni operon lacI gen ikkinchi nusxada ishlab chiqarilgan oqsil bilan samarali ravishda o'chiriladi lacI.
Agar xuddi shu tajriba operator mutatsiyasi yordamida o'tkazilsa, boshqacha natija olinadi (panel (f)). Bitta mutant va bitta yovvoyi turdagi operator uchastkasini olib yuradigan hujayraning fenotipi shundaki, LacZ va LacY IPTG induktori bo'lmagan taqdirda ham ishlab chiqariladi; chunki shikastlangan operator uchastkasi strukturali genlarning transkripsiyasini inhibe qilish uchun repressorni bog'lashiga yo'l qo'ymaydi. Operator mutatsiyasi ustunlik qiladi. Repressor bog'lanishi kerak bo'lgan operator maydonchasi mutatsiyaga uchraganida, o'sha katakchada ikkinchi funktsional uchastkaning mavjudligi mutant uchastkasi tomonidan boshqariladigan genlarning ekspressioni uchun hech qanday farq qilmaydi.
Ushbu tajribaning yanada murakkab versiyasidan foydalaniladi belgilangan ning ikki nusxasini ajratish uchun operonlar lak genlar va tartibga solinmagan tizimli gen (lar) mutant operator (panel (g)) yonidagi (lar) ekanligini ko'rsatadi. Masalan, bitta nusxa mutatsiya bilan belgilanadi lacZ shuning uchun u faqat LacY oqsilini ishlab chiqarishi mumkin, ikkinchi nusxasi esa mutatsiyaga ta'sir qiladi lacY va faqat LacZ ishlab chiqarishi mumkin. Ushbu versiyada faqat lak mutant operatoriga qo'shni bo'lgan operon IPTG holda ifodalanadi. Biz operator mutatsiyasini shunday deymiz cis-dominant, u yovvoyi turda dominant bo'lib, faqat unga yaqin bo'lgan operon nusxasiga ta'sir qiladi.
Ushbu tushuntirish muhim ma'noda chalg'itadi, chunki u eksperiment tavsifidan kelib chiqadi va natijada model nuqtai nazaridan tushuntiriladi. Ammo aslida, ko'pincha model birinchi o'rinda turishi haqiqatdir va eksperiment modelni sinash uchun maxsus ishlab chiqilgan. Yoqub va Monod dastlab a bo'lishi kerakligini tasavvur qilishdi sayt operatorning xususiyatlari bilan DNKda, keyin esa buni ko'rsatish uchun ularning komplementatsiya testlarini ishlab chiqdi.
Operator mutantlarining ustunligi ularni maxsus tanlash tartibini ham taklif qiladi. Agar tartibga soluvchi mutantlar, yuqorida tavsiflanganidek, fenil-Gal yordamida yovvoyi turdagi madaniyatdan tanlansa, operator mutatsiyalari repressor mutantlariga nisbatan kam uchraydi, chunki nishon hajmi juda kichikdir. Ammo buning o'rniga biz butun nusxaning ikki nusxasini ko'taradigan zo'riqishdan boshlaymiz lak mintaqa (ya'ni diploid) lak), repressor mutatsiyalari (hali ham ro'y beradi) tiklanmaydi, chunki ikkinchi, yovvoyi tur bilan to'ldiriladi lacI gen yovvoyi turdagi fenotipni beradi. Aksincha, operatorning bitta nusxasining mutatsiyasi mutant fenotipni keltirib chiqaradi, chunki u ikkinchi, yovvoyi turdagi nusxada dominant hisoblanadi.
Siklik AMP bilan tartibga solish[19]
Diauxie-ni tushuntirish, ta'sir qiluvchi qo'shimcha mutatsiyalarning tavsifiga bog'liq edi lak klassik model bilan izohlanganlardan boshqa genlar. Boshqa ikkita gen, cya va crp, keyinchalik xaritada joylashganligi aniqlandi lakva mutatsiyaga uchraganda, ning ifoda darajasi pasayishiga olib keladi mavjudlik IPTG va hatto repressor yoki operatorga ega bo'lmagan bakteriya shtammlarida. Kashfiyoti lager yilda E. coli mutantlarning nuqsonli ekanligini namoyish etishga olib keldi cya gen, lekin emas crp muhitga CAMP qo'shilishi bilan gen to'liq faollikda tiklanishi mumkin.
The cya gen adenilat siklazni kodlaydi, bu esa CAMP hosil qiladi. A cya mutant, cAMP yo'qligi ifodasini beradi lacZYA genlar odatdagidan o'n baravar past. CAMP qo'shilishi past Lac ifodasini tuzatadi cya mutantlar. Ikkinchi gen, crp, deb nomlangan oqsilni kodlaydi katabolit faollashtiruvchi oqsil (CAP) yoki cAMP retseptorlari oqsili (CRP).[20]
Ammo laktoza metabolizmi fermentlari oz miqdordagi glyukoza va laktoza (ba'zida sızıntılı ifoda deb nomlanadi) ishtirokida amalga oshiriladi, chunki LacI represörü DNK bilan qattiq bog'lanish o'rniga tez bog'lanib, ajralib chiqadi, bu vaqtni berishi mumkin. RNAP ning mRNKlarini bog'lashi va transkripsiyasi uchun lacZYA. Noqonuniy ekspluatatsiya glyukoza manbai sarflangandan so'ng, lekin oldinroq ba'zi laktoza metabolizmini ta'minlash uchun zarurdir lak ifoda to'liq faollashtirilgan.
Qisqa bayoni; yakunida:
- Laktoza yo'q bo'lganda, Lak fermenti ishlab chiqarilishi juda kam (operator unga Lac repressorini bog'lab qo'ygan).
- Laktoza mavjud bo'lganda, lekin afzal qilingan uglerod manbai (masalan, glyukoza) bo'lsa, unda oz miqdordagi ferment hosil bo'ladi (Lak repressori operator bilan bog'lanmagan).
- Glyukoza yo'q bo'lganda, CAP-cAMP promotorning yuqori qismida ma'lum bir DNK uchastkasi bilan bog'lanadi va RNAP bilan to'g'ridan-to'g'ri protein-oqsil ta'sirini o'tkazadi, bu esa RNAPning promotor bilan bog'lanishini osonlashtiradi.
O'sish fazalari orasidagi kechikish etarli miqdordagi laktoza almashinadigan fermentlarni ishlab chiqarish uchun zarur bo'lgan vaqtni aks ettiradi. Birinchidan, CAP me'yoriy oqsili birlashtirilishi kerak lak promouter, natijada ishlab chiqarish ko'payadi lak mRNA. Qo'shimcha nusxalari lak mRNK ishlab chiqarishga olib keladi (qarang) tarjima ) LacZ (b-galaktosidaza, laktoza almashinuvi uchun) va LacY (laktoza hujayraga laktoza tashish uchun permeaza) nusxalarining sezilarli darajada ko'pligi. Laktoza almashinadigan fermentlar darajasini oshirish uchun zarur bo'lgan kechikishdan keyin bakteriyalar yangi tez bosqichga o'tadilar. hujayralar o'sishi.

Ikki jumboq katabolit repressiyasi CAMP darajalari glyukoza borligi bilan qanday bog'liqligi, ikkinchidan, hujayralar nima uchun bezovtalanishi kerakligi bilan bog'liq. Laktoza bo'linib bo'lgach, u aslida glyukoza va galaktozani hosil qiladi (osonlik bilan glyukozaga aylanadi). Metabolik jihatdan laktoza xuddi shunday yaxshi glyukoza sifatida uglerod va energiya manbai. CAMP darajasi hujayra ichidagi glyukoza konsentratsiyasi bilan emas, balki glyukoza tashish tezligi bilan bog'liq bo'lib, bu adenilat siklaza faolligiga ta'sir qiladi. (Bundan tashqari, glyukoza tashish ham laktoza permeazasini bevosita inhibisyoniga olib keladi.) Nima uchun E. coli shu tarzda ishlaydi, faqat taxmin qilish mumkin. Barcha ichak bakteriyalari glyukozani fermentatsiyalaydi, bu esa ular bilan tez-tez uchrashishini ko'rsatadi. Ehtimol, glyukoza v. Laktozani tashish yoki metabolizm samaradorligining ozgina farqi hujayralarni regulyatsiya qilish uchun foydalidir. lak shu tarzda operon.[21]
Molekulyar biologiyada foydalaning
The lak gen va uning hosilalari a sifatida ishlatilishi mumkin muxbir gen kabi bir qator bakteriyalarga asoslangan tanlov usullarida ikkita gibrid tahlil qilish, unda muvaffaqiyatli bog'lanish transkripsiya faollashtiruvchisi ma'lum bir promouterlik ketma-ketligi aniqlanishi kerak.[14] Yilda FUNT o'z ichiga olgan plitalar X-gal, rangning oq rangdagi koloniyalardan ko'k ranggacha o'zgarishi taxminan 20-100 g-galaktozidaza birliklariga to'g'ri keladi, tetrazolium laktoza va MacConkey laktoza tashuvchisi 100-1000 birlik oralig'iga ega bo'lib, ushbu diapazonning yuqori va past qismlarida eng sezgir.[14] MacConkey laktoza va tetrazolium laktoza vositalari ikkalasi ham laktoza parchalanish mahsulotlariga ishonganligi sababli, ikkalasi ham mavjud bo'lishini talab qiladi lacZ va lacY genlar. Ko'pchilik lak faqat o'z ichiga olgan termoyadroviy texnikasi lacZ gen X-gal plitalariga mos keladi[14] yoki ONPG suyuq bulyonlar.[22]
Shuningdek qarang
Adabiyotlar
- ^ a b Griffits, Entoni JF.; Vessler, Syuzan R.; Kerol, Shon B.; Doebley, Jon (2015). Genetik tahlilga kirish (11 nashr). Freeman, W.H. & Kompaniyasi. 400-412 betlar. ISBN 9781464109485.
- ^ MakKlin, Fillip (1997). "Prokaryotik gen ekspressioni". ndsu.edu. Olingan 19 may 2017.
- ^ "Prokaryotik gen ekspressioni". ndsu.edu. Olingan 19 may 2017.
- ^ Basbi S., Ebrayt RH. (2001). "Katabolit faollashtiruvchi oqsil (CAP) bilan transkripsiyani faollashtirish". J. Mol. Biol. 293 (2): 199–213. doi:10.1006 / jmbi.1999.3161. PMID 10550204.
- ^ Kennell, Devid; Rizman, Xovard (1977 yil iyul). "Escherichia coli lac operonining transkripsiyasi va tarjimani boshlash chastotalari". Molekulyar biologiya jurnali. 114 (1): 1–21. doi:10.1016/0022-2836(77)90279-0. PMID 409848.
- ^ Malan, T. Filipp; Kolb, Enni; Buk, Anri; Makklur, Uilyam (1984 yil dekabr). "CR1-cAMP operatsiyasining transkripsiyasini boshlash operatsiyasini faollashtirish mexanizmi P1 promouterini faollashtirish". J. Mol. Biol. 180 (4): 881–909. doi:10.1016/0022-2836(84)90262-6. PMID 6098691.
- ^ Görke B, Stulke J (Avgust 2008). "Bakteriyalarda uglerod katabolitining repressiyasi: foydali moddalardan maksimal darajada foydalanishning ko'plab usullari". Tabiat sharhlari. Mikrobiologiya. 6 (8): 613–24. doi:10.1038 / nrmicro1932. PMID 18628769. S2CID 8782171.
- ^ Oler, S .; Eismann, E. R.; Krämer, H .; Myuller-Xill, B. (1990). "Operator operatsiyasining uchta operatori repressiyada hamkorlik qiladi". EMBO jurnali. 9 (4): 973–979. doi:10.1002 / j.1460-2075.1990.tb08199.x. PMC 551766. PMID 2182324.
- ^ Griffits, Entoni JF; Gelbart, Uilyam M.; Miller, Jefri X.; Lewontin, Richard C. (1999). "Laktoza tizimini tartibga solish". Zamonaviy genetik tahlil. Nyu-York: W. H. Freeman. ISBN 0-7167-3118-5.
- ^ fon Xippel, PH .; Revzin, A .; Gross, C.A .; Vang, A.C. (1974 yil dekabr). "Biologik nazorat mexanizmi sifatida genomni boshqaruvchi oqsillarni o'ziga xos bo'lmagan DNK bilan bog'lash: I. Lak operon: muvozanat aspektlari". PNAS. 71 (12): 4808–12. doi:10.1073 / pnas.71.12.4808. PMC 433986. PMID 4612528.
- ^ Hansen LH, Knudsen S, Sørensen SJ (iyun 1998). "O'rganilgan lacY genining IPTG induktsiyali promotorlari induksiyasiga ta'siri Escherichia coli va Pseudomonas floresanlari". Curr. Mikrobiol. 36 (6): 341–7. doi:10.1007 / s002849900320. PMID 9608745. S2CID 22257399. Arxivlandi asl nusxasi 2000 yil 18 oktyabrda.
- ^ Marbax A, Bettenbrok K (2012 yil yanvar). "Escherichia coli-da lak operon induktsiyasi: IPTG va TMG induksiyasini va transatsetilaza LacA ta'sirini tizimli ravishda taqqoslash". Biotexnologiya jurnali. 157 (1): 82–88. doi:10.1016 / j.jbiotec.2011.10.009. PMID 22079752.
- ^ "ONPG (β-Galaktosidaza) sinovi". Sentyabr 2000. Arxivlangan asl nusxasi 2007 yil 3-noyabrda. Olingan 25 oktyabr 2007.
- ^ a b v d Joung J, Ramm E, Pabo S (2000). "Oqsil-DNK va oqsil-oqsilning o'zaro ta'sirini o'rganish uchun bakterial ikki-gibrid selektsiya tizimi". Proc Natl Acad Sci AQSh. 97 (13): 7382–7. doi:10.1073 / pnas.110149297. PMC 16554. PMID 10852947.
- ^ "Milestone 2 - Vizyoner juftlik: Genlarni ifodalashdagi tabiatning muhim bosqichlari". www.nature.com. Olingan 27 dekabr 2015.
- ^ Myuller-Xill, Benno (1996). Lac Operon, Genetik Paradigmaning Qisqa Tarixi. Berlin: Valter de Gruyter. 7-10 betlar. ISBN 3-11-014830-7.
- ^ McKnight, Steven L. (1992). Transkripsiyani tartibga solish. Cold Spring Harbor, NY: Cold Spring Harbor laboratoriyasining matbuoti. pp.3–24. ISBN 0-87969-410-6.
- ^ Jeykob F.; Monod J (1961 yil iyun). "Oqsillarni sintez qilishda genetik tartibga solish mexanizmlari". J Mol Biol. 3 (3): 318–56. doi:10.1016 / S0022-2836 (61) 80072-7. PMID 13718526.
- ^ Montminy, M. (1997). "Tsiklik AMP bilan transkripsiyani tartibga solish". Biokimyo fanining yillik sharhi. 66: 807–822. doi:10.1146 / annurev.biochem.66.1.807. ISSN 0066-4154. PMID 9242925.
- ^ Botsford, J L; Harman, J G (1992 yil mart). "Prokaryotlarda tsiklik AMP". Mikrobiologik sharhlar. 56 (1): 100–122. doi:10.1128 / MMBR.56.1.100-122.1992. ISSN 0146-0749. PMC 372856. PMID 1315922.
- ^ Vazquez A, Beg QK, Demenezes MA va boshq. (2008). "Erituvchi qobiliyatini cheklashning E. coli metabolizmiga ta'siri". BMC Syst Biol. 2: 7. doi:10.1186/1752-0509-2-7. PMC 2270259. PMID 18215292.
- ^ "Lak operonini E. coli-ga induksiya qilish" (PDF). SAPS. Olingan 29 iyun 2016.
Tashqi havolalar
- Lac + Operon AQSh Milliy tibbiyot kutubxonasida Tibbiy mavzu sarlavhalari (MeSH)
- lak operon NCBI Bookshelf-da [2]
- Virtual hujayra animatsiyasi to'plami Kirish: Lac Operon
- Lac Operon: Bozeman Science
- B-Galaktosidaza (lacZ) faolligi uchun butun sichqonchani embrionlarini bo'yash