Tabiiy vakolat - Natural competence - Wikipedia
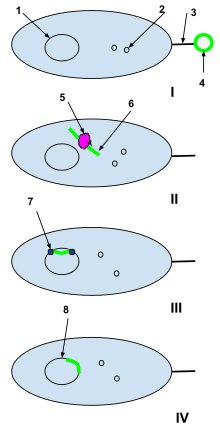
1-bakterial hujayralar DNKsi
2-bakterial hujayralar plazmidlari
3-jinsiy pili
4-o'lik hujayradan yot DNK plazmidasi
5-bakteriyalar hujayralarining restriksion fermenti
6-Yaralanmagan xorijiy plazmid
7-DNK ligazasi
Men: O'lik hujayradan yot DNK plazmidini tabiiy ravishda vakolatli bakterial hujayraning jinsiy pili ushlaydi.
II: Chet ellik plazmid jinsiy pili orqali bakteriyalar hujayrasiga o'tkaziladi va u erda bakteriyalar hujayralarining cheklash fermentlari tomonidan qayta ishlanadi. Restrikt fermentlari begona plazmidni bakterial DNKga qo'shilishi mumkin bo'lgan nukleotidlar zanjiriga aylantiradi.
III: DNK ligazasi begona nukleotidlarni bakteriyalar hujayrasi DNKga birlashtiradi.
IV: Rekombinatsiya tugadi va begona DNK asl bakterial hujayraning DNKsiga qo'shildi va keyinchalik bakteriya hujayrasi takrorlanganda uning bir qismi bo'lib qoladi.
Yilda mikrobiologiya, genetika, hujayra biologiyasi va molekulyar biologiya, vakolat a qobiliyatidir hujayra hujayradan tashqari ("yalang'och") genetikani o'zgartirish DNK deb nomlangan jarayonda uning muhitidan transformatsiya. Qobiliyat o'rtasida farqlanishi mumkin tabiiy vakolat, ning genetik jihatdan ko'rsatilgan qobiliyati bakteriyalar laboratoriyada bo'lgani kabi tabiiy sharoitda ham sodir bo'ladi deb o'ylashadi va induktsiya qilingan yoki sun'iy vakolatlaboratoriya madaniyatidagi hujayralarni DNKga o'tkazuvchanligini ta'minlash uchun ularni davolashda paydo bo'ladi. Qobiliyat hujayraning tez moslashishi va DNKni tiklashga imkon beradi. Ushbu maqolada, birinchi navbatda, bakteriyalarning tabiiy vakolatlari haqida so'z yuritiladi, ammo sun'iy vakolatlar to'g'risida ham ma'lumot berilgan.
Tarix
Tabiiy vakolatlar tomonidan kashf etilgan Frederik Griffit 1928 yilda u patogen bakteriyaning o'ldirilgan hujayralari tarkibida patogen bo'lmagan hujayralarni patogen turga aylantiradigan narsa borligini ko'rsatganda. 1944 yilda Osvald Avery, Kolin MacLeod va Maklin Makkarti ushbu "o'zgaruvchan omil" toza ekanligini namoyish etdi DNK[1] . Bu DNK hujayraning genetik ma'lumotlarini olib borishini tasdiqlovchi birinchi dalil edi.
O'shandan beri tabiiy kompetentsiya turli xil bakteriyalarda, xususan o'rganilgan Bacillus subtilis, Streptokokk pnevmoniyasi (Griffitning "pnevmokokklari"), Neisseria gonorrhoeae, Gemofilus grippi va a'zolari Acinetobacter tur. Faol tadqiqot yo'nalishlari DNKni tashish mexanizmlari, turli bakteriyalarda kompetensiyani tartibga solish va kompetensiyaning evolyutsion funktsiyasini o'z ichiga oladi.
DNKni qabul qilish mexanizmlari
Laboratoriyada DNK tadqiqotchi tomonidan taqdim etiladi, ko'pincha genetik jihatdan yaratilgan qism yoki plazmid. Qabul qilish paytida DNK orqali uzatiladi hujayra membranasi, va hujayra devori agar u mavjud bo'lsa. DNK hujayraning ichiga kirgandan so'ng uni parchalanishi mumkin nukleotidlar uchun qayta ishlatilgan DNKning replikatsiyasi va boshqalar metabolik funktsiyalari. Shu bilan bir qatorda bo'lishi mumkin birlashtirilgan hujayraning ichiga genom uning tomonidan DNKni tiklash fermentlar. Agar bu rekombinatsiya hujayrani o'zgartirsa genotip hujayra o'zgartirilgan deb aytiladi. Sun'iy vakolat va transformatsiya ko'plab organizmlarda tadqiqot vositalari sifatida ishlatiladi (qarang Transformatsiya (genetika) ).[2]
Deyarli barcha tabiiy vakolatli bakteriyalarda hujayradan tashqaridagi filamentlarning tarkibiy qismlari deyiladi IV pili (turi fimbriya ) hujayra tashqari ikki qavatli DNKni bog'laydi. Keyin DNK membrana bo'ylab translokatsiya qilinadi (yoki uchun membranalar) gramm salbiy bakteriyalar) DNKning bir zanjirining parchalanishi natijasida hosil bo'lgan ko'p komponentli oqsil komplekslari orqali. Hujayradagi bitta zanjirli DNK DNKni yuklaydigan yaxshi saqlangan DprA oqsillari bilan bog'lanadi RecA, bu klassik orqali gomologik rekombinatsiyani vositachilik qiladi DNKni tiklash yo'l.[3]
Vakolatni tartibga solish
Laboratoriya madaniyatlarida tabiiy kompetentsiya odatda qat'iy tartibga solinadi va ko'pincha ozuqaviy etishmovchilik yoki noqulay sharoitlar tufayli yuzaga keladi. Shu bilan birga, o'ziga xos induktsiya signallari va tartibga soluvchi mexanizmlar qabul qilish mexanizmlariga qaraganda ancha o'zgaruvchan va bu bakteriyalarning tabiiy muhitida vakolatlarni tartibga solish haqida kam narsa ma'lum.[4] Transkripsiya omillari vakolatni tartibga soluvchi kashf etilgan; Masalan, sxy (tfoX nomi bilan ham tanilgan) a tomonidan tartibga solinishi aniqlandi 5 'kodlamaydigan RNK elementi.[5] Shakllanish qobiliyatiga ega bakteriyalarda sporlar, sporulyatsiyani keltirib chiqaradigan holatlar ko'pincha vakolatli vakolatlar bilan mos keladi. Shunday qilib, sporulyatsion hujayralarni o'z ichiga olgan kulturalar yoki koloniyalar ko'pincha vakolatli hujayralarni o'z ichiga oladi. Syulning so'nggi tadqiqotlari va boshq. vakolatga kirish va chiqishni tushuntirib beradigan genlarning qo'zg'atuvchi asosiy modulini aniqladi uyali shovqin hisobga olinadi.[6]
Aksariyat vakolatli bakteriyalar samaradorligi taxminan teng bo'lgan barcha DNK molekulalarini oladi, ammo oilalardagi bakteriyalar Neisseriaceae va Pasterellalar o'z ichiga olgan DNK parchalarini olish afzalroq qisqa DNK sekanslari, Neisseriaceae-da DNKni qabul qilish ketma-ketligi (DUS) va o'zlarining genomlarida juda tez-tez uchraydigan Pasterellarda (SUS) signal signallari ketma-ketligi. Neisserial genomlari GCCGTCTGAA, va Pasterellean genomlarda AAGTGCGGT yoki ACAAGCGGT mavjud.[2][7]
Kompetensiyaning evolyutsion funktsiyalari va natijalari
Ko'pchilik takliflar boshlang'ich bosqichida o'tkazildi evolyutsion tabiiy bakteriyalar transformatsiyasining bir qismi sifatida tabiiy kompetensiyaning vazifasi uch toifaga bo'linadi: (1) genetik xilma-xillikning selektiv afzalligi; (2) nukleotidlar manbai sifatida DNKni qabul qilish ("oziq-ovqat" sifatida DNK); va (3) zararlangan DNKning gomologik rekombinatsion tiklanishiga ko'maklashish uchun DNKning yangi zanjirining selektiv afzalligi. Ikkilamchi taklif ham berildi, bu vaqti-vaqti bilan afzalligini ta'kidladi gorizontal genlarning uzatilishi.
Genetik xilma-xillik gipotezasi
Genetika xilma-xilligini qo'llab-quvvatlash uchun argumentlarni jinsiy aloqaning asosiy evolyutsiyasi funktsiyasi (shu jumladan bakterial transformatsiya) Barton va Charlvort tomonidan keltirilgan.[8] va Otto va Gershteyn tomonidan.[9] Biroq, bilan bog'liq nazariy qiyinchiliklar jinsiy aloqa evolyutsiyasi genetik xilma-xillik uchun jinsiy aloqa muammoli ekanligini taxmin qilish. Xususan, bakterial transformatsiyaga kelsak, kompetensiya global oqsil sintezi kaliti uchun yuqori narxni talab qiladi, masalan, 16 dan ortiq genlar faqat vakolati davrida yoqiladi. Streptokokk pnevmoniyasi.[10] Ammo, bakteriyalar klonlarda o'sishga moyil bo'lganligi sababli, transformatsiya uchun mavjud bo'lgan DNK odatda qabul qiluvchi hujayralar bilan bir xil genotipga ega bo'ladi. Shunday qilib, umuman olganda, xilma-xillikni ko'paytirmasdan, oqsil ekspresyonida har doim ham yuqori narx mavjud. Barkamollik va jinsiy munosabatlar o'rtasidagi boshqa farqlar kompetensiyani keltirib chiqaradigan genlar evolyutsiyasi modellarida ko'rib chiqilgan; Ushbu modellar kompetensiyaning postulyatsiya qilingan rekombinatsion foydalari jinsiy aloqaga qaraganda ancha qiyinligini aniqladi.[11]
Oziq-ovqat sifatida DNK gipotezasi
Ikkinchi gipoteza, oziq-ovqat sifatida DNK, DNKni qabul qiladigan hujayralarning muqarrar ravishda DNK tarkibidagi nukleotidlarni egallashiga va, chunki nukleotidlar DNK va RNK sintez va sintez qilish qimmat bo'lganligi sababli, ular hujayraning energiya byudjetiga katta hissa qo'shishi mumkin.[12] Ba'zi tabiiy vakolatli bakteriyalar atrofdagi nukleazalarni ham ajratib turadi va barcha bakteriyalar ushbu nukleazalar atrof-muhit DNKidan hosil bo'lgan erkin nukleotidlarni egallashi mumkin.[13] DNKni yutish energetikasi har qanday tizimda tushunilmaydi, shuning uchun nukleaz sekretsiyasining samaradorligini DNKni qabul qilish va ichki parchalanish bilan taqqoslash qiyin. Printsipial jihatdan nukleaz ishlab chiqarish xarajatlari va nukleotidni tiklash noaniqligi qabul qilish mexanizmini sintez qilish va DNKni tortib olish uchun zarur bo'lgan energiyaga nisbatan muvozanatli bo'lishi kerak. Boshqa muhim omillar - nukleazalar va vakolatli hujayralar DNK molekulalariga duch kelish ehtimoli, nisbiy samarasizlik atrof-muhitdan va periplazmadan nukleotidlarni o'zlashtirishi (bu erda bitta ip vakolatli hujayralar tomonidan parchalanadi) va sitoplazmadagi boshqa ipdan foydalanishga tayyor nukleotid monofosfatlar ishlab chiqarishning afzalligi. Yana bir qiyinlashtiruvchi omil - bu oiladagi turlarning DNKni qabul qilish tizimlarining o'zboshimchalikidir Pasterellalar va tur NayseriyaBu rekombinatsiya yoki mexanik jihatdan samarali qabul qilish uchun tanlovni aks ettirishi mumkin.[14][15]
DNK zararini tiklash gipotezasi
Bakteriyalarda DNKning shikastlanishi muammosi stress yoki ochlik sharoitida yuzaga keladigan stress, ayniqsa oksidlovchi stress paytida eng ko'p uchraydi. Bunday sharoitda ko'pincha bitta xromosoma mavjud. Ba'zi bakteriyalar bunday stress sharoitida kompetentsiyani keltirib chiqaradi, degan xulosa, DNKni tiklashga imkon beradigan transformatsiya mavjud bo'lgan uchinchi farazni qo'llab-quvvatlaydi. Eksperimental sinovlarda, ularning DNKlariga zarar etkazadigan vositalarga duchor bo'lgan va keyinchalik transformatsiyaga uchragan bakterial hujayralar DNKning shikastlanishiga duchor bo'lmagan hujayralarga qaraganda yaxshiroq omon qolishdi (Hoelzer va Michod, 1991).[16] Bundan tashqari, transformatsiyani boshdan kechirish vakolati ko'pincha ma'lum DNKga zarar etkazuvchi vositalar tomonidan qo'zg'atiladi (Mikod tomonidan ko'rib chiqilgan) va boshq., 2008 va Bernshteyn va boshq., 2012).[17][18] Shunday qilib, tabiiy vakolat va transformatsiya uchun kuchli qisqa muddatli selektiv ustunlik uning stress sharoitida DNKning gomologik rekombinatsion tiklanishiga ko'maklashish qobiliyatidir. Bunday stress holatlari sezgir xujayraning bakterial infektsiyasi paytida yuzaga kelishi mumkin. Ushbu fikrga muvofiq, Li va boshq.[19] turli xil yuqori darajada o'zgaruvchanligi haqida xabar berdi S. pnevmoniya ajratadi, nazal kolonizatsiyaning yaroqliligi va virulentligi (o'pkaning yuqumli kasalligi) buzilmagan kompetentsiya tizimiga bog'liq.
1993 yilda Redfildning xromosoma DNKsiga bir qatorli va ikki qatorli zarar etkazilishi kompetensiyani yoki transformatsiyani keltirib chiqarmagan yoki kuchaytirmaganligini aniqlagan qarama-qarshi dalillar keltirildi. B. subtilis yoki H. grippi, ta'mirlash uchun tanlov ushbu turlarning vakolatlari evolyutsiyasida kam yoki umuman rol o'ynamaganligini ko'rsatmoqda[20]
Ammo yaqinda keltirilgan dalillar shuni ko'rsatadiki, transformatsiya qobiliyatlari, albatta, DNKning zarar etkazadigan sharoitlari bilan bog'liq. Masalan, Claverys va boshq. 2006 yilda[21] DNKga zarar etkazuvchi vositalar mitomitsin C (DNKni o'zaro bog'laydigan vosita) va ftorxinolon (er-xotin uzilishlarni keltirib chiqaradigan topoizomeraza inhibitori) Streptokokk pnevmoniyasi. Bundan tashqari, Engelmoer va Rozen[22] 2011 yilda buni namoyish etdi S. pnevmoniya transformatsiya mitomitsin S ning bakteritsid ta'siridan himoya qiladi. Kanomitsin va streptomitsin antibiotiklaridan himoyalangan kompetensiya induksiyasi.[21][22] Ushbu aminoglikozid antibiotiklari ilgari DNKga zarar etkazuvchi emas deb hisoblangan bo'lsa-da, 2012 yilda Fotining so'nggi tadqiqotlari va boshq.[23] ularning bakteritsid faolligining katta qismi ajralib chiqishi natijasida kelib chiqishini ko'rsatdi gidroksil radikal va DNK ziyonini induktsiya qilish, shu jumladan ikki zanjirli tanaffuslar.
Dorer va boshq.,[24] 2010 yilda DNK giraza bilan o'zaro ta'sir qiluvchi va ikki qatorli tanaffuslar hosil bo'lishiga olib keladigan siprofloksatsin genlarning vakolatlanishini keltirib chiqaradi. Helicobacter pylori, transformatsiyani kuchayishiga olib keladi. 2011 yilda o'tkazilgan tadqiqotlar Legionella pneumophila, Charpentier va boshq.[25] 64 ta toksik molekulani sinovdan o'tkazib, qaysi biri malakaga ega ekanligini aniqlash uchun Ushbu molekulalarning atigi oltitasi, barcha DNKga zarar etkazuvchi vositalar, vakolatni kuchaytirgan. Ushbu molekulalar edi norfloksatsin, ofloksatsin va nalidiksik kislota (ingibitorlari DNK-giraza ikki qatorli uzilishlarni keltirib chiqaradi[26]), mitomitsin C (bu tarmoqlararo o'zaro bog'lanishlarni ishlab chiqaradi), bisiklomitsin (bitta va ikki qatorli tanaffuslarni keltirib chiqaradi[27]) va gidroksiüre (DNK asoslarining oksidlanishiga sabab bo'ladi[28]). Charpentier va boshq.[25] buni ham ko'rsatdi Ultrabinafsha nurlanish vakolatni keltirib chiqaradi L. pneumophila va bundan tashqari, transformatsiya qobiliyatlari DNKning zararlanishiga javob sifatida rivojlangan deb taxmin qildi.
Genlarni gorizontal ravishda uzatish
Vaqti-vaqti bilan uzoq muddatli afzalliklarga ba'zida berilishi mumkin gorizontal genlarning uzatilishi ham chaqirdi genlarni lateral uzatishi, (natijada bo'lishi mumkin homolog bo'lmagan rekombinatsiya antibiotiklarga qarshilik yoki boshqa afzalliklarni ta'minlashi mumkin bo'lgan vakolat paydo bo'lgandan keyin).
Qobiliyatni tanlash xususiyatidan qat'i nazar, bakteriyalar genomlarining kompozitsion tabiati, kompetensiya natijasida kelib chiqqan gorizontal genlar almashinuvi evolyutsiyani amalga oshiradigan genetik xilma-xillikka hissa qo'shganligi to'g'risida ko'plab dalillarni keltiradi.
Shuningdek qarang
Adabiyotlar
- ^ Avery OT, Makleod CM, Makkarti M (1944). "Pnevmokokk turlarini o'zgartiradigan moddaning kimyoviy tabiati bo'yicha tadqiqotlar". J. Exp. Med. 79 (2): 137–58. doi:10.1084 / jem.79.2.137. PMC 2135445. PMID 19871359.
- ^ a b Chen I, Dubnau D (2004). "Bakterial transformatsiya paytida DNKni qabul qilish". Nat. Vahiy Mikrobiol. 2 (3): 241–9. doi:10.1038 / nrmicro844. PMID 15083159. S2CID 205499369.
- ^ Johnston C, Martin B, Fichant G, Polard P, Claverys J (2014). "Bakterial transformatsiya: tarqalish, umumiy mexanizmlar va divergent nazorat". Nat. Vahiy Mikrobiol. 12 (3): 181–96. doi:10.1038 / nrmicro3199. PMID 24509783. S2CID 23559881.
- ^ Solomon JM, Grossman AD (1996). "Kim va qachon vakolatli: bakteriyalarda tabiiy genetik kompetensiyani tartibga solish". Trends Genet. 12 (4): 150–5. doi:10.1016/0168-9525(96)10014-7. PMID 8901420.
- ^ Redfield RJ (sentyabr 1991). "sxy-1, gemofilus grippi mutatsiyasi, spontan kompetensiyani kuchaytiradi". J. Bakteriol. 173 (18): 5612–8. doi:10.1128 / jb.173.18.5612-5618.1991. PMC 208288. PMID 1653215.
- ^ Syel GM, Garcia-Ojalvo J, Liberman LM, Elowitz MB (2006). "Qo'zg'aluvchan genlarni tartibga solish davri vaqtinchalik uyali differentsiatsiyani keltirib chiqaradi" (PDF). Tabiat. 440 (7083): 545–50. doi:10.1038 / nature04588. PMID 16554821. S2CID 4327745.
- ^ Findlay, VA; Redfild, RJ (2009). "DNKni qabul qilish ketma-ketligi va bakterial proteomlarning koevolyutsiyasi". Genom biologiyasi va evolyutsiyasi. 1: 45–55. doi:10.1093 / gbe / evp005. PMC 2817400. PMID 20333176.
- ^ Barton NH, Charlvort B (1998). "Nima uchun jinsiy aloqa va rekombinatsiya?". Ilm-fan. 281 (5385): 1986–1990. doi:10.1126 / science.281.5385.1986. PMID 9748151.
- ^ Otto SP, Gershteyn AC (avgust 2006). "Nima uchun jinsiy aloqa qilish kerak? Jinsiy populyatsiya genetikasi va rekombinatsiya". Biochem Soc Trans. 34 (Pt 4): 519-522. doi:10.1042 / BST0340519. PMID 16856849.
- ^ Peterson S, Cline RT, Tettelin H, Sharov V, Morrison DA (Nov 2000). "Streptococcus pneumoniae kompetensiya regulyatorlarining DNK mikro-nurlari yordamida gen ekspression tahlili". J. Bakteriol. 182 (21): 6192–6202. doi:10.1128 / JB.182.21.6192-6202.2000. PMC 94756. PMID 11029442.
- ^ Redfield R (1988). "O'lik hujayralar bilan jinsiy aloqa hech qachon jinsiy aloqadan yaxshiroq bo'ladimi?". Genetika. 119 (1): 213–21. PMC 1203342. PMID 3396864.
- ^ Redfield RJ (2001). "Bakteriyalar jinsiy aloqada bo'ladimi?". Nat. Rev. Genet. 2 (8): 634–9. doi:10.1038/35084593. PMID 11483988. S2CID 5465846.
- ^ Dubnau D (1999). "Bakteriyalarda DNKning emirilishi". Annu Rev Microbiol. 53 (1): 217–44. doi:10.1146 / annurev.micro.53.1.217. PMID 10547691.
- ^ Maughan H (2010). "DNKni bakteriyalarga singdirish ketma-ketligi faqat molekulyar qo'zg'alish orqali to'planishi mumkin". Genetika. 186 (2): 613–27. doi:10.1534 / genetika.110.119438. PMC 2954483. PMID 20628039.
- ^ Redfield R, Schrag M, Dead A (1997). "Bakterial transformatsiya evolyutsiyasi: jinsiy munosabatlar yomon munosabatlar". Genetika. 146 (1): 27–38. PMC 1207942. PMID 9135998.
- ^ Hoelzer MA, Michod RE (1991). "DNKning tiklanishi va uning o'zgarishi evolyutsiyasi Bacillus subtilis. III. DNK bilan zararlangan jinsiy aloqa ". Genetika. 128 (2): 215–23. PMC 1204460. PMID 1906416.
- ^ Michod RE, Bernstein H, Nedelcu AM (2008). "Mikrobial patogenlarda jinsiy aloqaning adaptiv qiymati". Genet Evol-ni yuqtirish. 8 (3): 267–85. doi:10.1016 / j.meegid.2008.01.002. PMID 18295550. http://www.hummingbirds.arizona.edu/Faculty/Michod/Downloads/IGE%20review%20sex.pdf
- ^ Bernshteyn, Xarris; Kerol Bernshteyn; Richard E. Mixod (2012). "1-bob - DNKni tiklash bakteriyalar va evkaryotlarda jinsiy aloqaning asosiy adaptiv funktsiyasi sifatida". DNKni tiklash: yangi tadqiqotlar. NOVA nashriyotlari. 1-50 betlar. ISBN 978-1-62100-756-2. Arxivlandi asl nusxasi 2013-10-29 kunlari. Olingan 2012-04-13. https://www.novapublishers.com/catalog/product_info.php?products_id=31918
- ^ Li G, Liang Z, Vang X, Yang Y, Shao Z, Li M, Ma Y, Qu F, Morrison DA, Chjan JR (2016). "In Vivo Fitness va Virulentlik uchun Gipertransformatsiyalangan Pnevmokokk izolyatorlarning tabiiy o'zgarishga qo'shilishi". Yuqtirish. Immun. 84 (6): 1887–901. doi:10.1128 / IAI.00097-16. PMC 4907133. PMID 27068094.
- ^ Redfield R (1993). "Tabiiy o'zgarish evolyutsiyasi: DNKni tiklash gipotezasini sinab ko'rish Bacillus subtilis va Gemofilus grippi". Genetika. 133 (4): 755–61. PMC 1205397. PMID 8462839.
- ^ a b Claverys, JP; Prudxom, M; Martin, B (2006). "Stressga umumiy javob sifatida kompetentsiya regulyatorlarini kiritish grammusbat bakteriyalar ". Annu Rev Microbiol. 60 (1): 451–475. doi:10.1146 / annurev.micro.60.080805.142139. PMID 16771651.
- ^ a b Engelmoer, D J; Rozen, D E (2011). "Quvvatlilik stress paytida hayotni oshiradi Streptokokk pnevmoniyasi". Evolyutsiya. 65 (12): 3475–3485. doi:10.1111 / j.1558-5646.2011.01402.x. PMID 22133219.
- ^ Foti, JJ; Devadoss, B; Vinkler, JA; Kollinz, JJ; Walker, GC (2012). "Guanin nukleotid havzasining oksidlanishi bakteritsidli antibiotiklar ta'sirida hujayralar o'limiga asos bo'ladi". Ilm-fan. 336 (6079): 315–319. doi:10.1126 / science.1219192. PMC 3357493. PMID 22517853.
- ^ Dorer, MS; Fero, J; Salama, NR (2010). "DNKning shikastlanishi genetik almashinuvni keltirib chiqaradi Helicobacter pylori". PLOS Pathog. 6 (7): e1001026. doi:10.1371 / journal.ppat.1001026. PMC 2912397. PMID 20686662.
- ^ a b Charpentier, X; Kay, E; Shnayder, D; Shuman, XA (2011). "Antibiotiklar va ultrabinafsha nurlanish tabiiy o'zgarishlarning malakasini keltirib chiqaradi Legionella pneumophila". J bakteriol. 193 (5): 1114–1121. doi:10.1128 / JB.01146-10. PMC 3067580. PMID 21169481.
- ^ Albertini, S; Chetelat, A A; Miller, B; Muster, Vt; Pujadas, E; Strobel, R; Gocke, E (1995). "Prokaryotik va eukaryotik sinov tizimlarida 17 giraza- va to'rtta sutemizuvchi topoizomeraza II-zaharlarining genotoksikligi". Mutagenez. 10 (4): 343–351. doi:10.1093 / mutage / 10.4.343. PMID 7476271.
- ^ Washburn, R S; Gottesman, M E (2011). "Transkripsiya tugashi xromosoma yaxlitligini saqlaydi". Proc Natl Acad Sci U S A. 108 (2): 792–7. doi:10.1073 / pnas.1009564108. PMC 3021005. PMID 21183718.
- ^ Sakano, K; Oikava, S; Xasegava, K; Kawanishi, S (2001). "Gidroksiüre vodorod peroksid va azot oksidi hosil bo'lishi bilan saytga xos DNKning shikastlanishiga olib keladi". Jpn J saraton kasalligi. 92 (11): 1166–1174. doi:10.1111 / j.1349-7006.2001.tb02136.x. PMC 5926660. PMID 11714440.