Bir-birini qoplaydigan gen - Overlapping gene
An bir-birini qoplaydigan gen (yoki OLG)[1] a gen kimning tushunarli nukleotidlar ketma-ketligi qisman boshqa genning ifodalanadigan nukleotidlar ketma-ketligi bilan qoplanadi.[2] Shu tarzda, nukleotidlar ketma-ketligi bir yoki bir nechtasining ishlashiga hissa qo'shishi mumkin gen mahsulotlari. Bosib chiqarish bitta genning ketma-ketligi to'liq yoki qisman muqobil ravishda o'qiladigan takrorlanish turini bildiradi o'qish doirasi bir vaqtning o'zida boshqa gendan lokus. Overprinting mexanizmi sifatida faraz qilingan de novo yangi genlarning paydo bo'lishi eski genlardan yoki ilgari mavjud bo'lgan ketma-ketliklardan kodlamaslik genomning mintaqalari.[3] Haddan tashqari bosib chiqarilgan genlar ayniqsa genomik viruslarni tashkil qilish, ehtimol virusli genetik ma'lumotlarning kichik to'plamidan potentsial aniqlanadigan genlar sonini ko'paytiradi.
Tasnifi
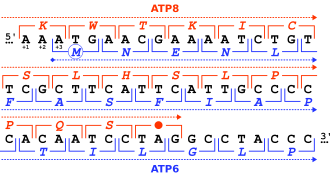
Genlar turli yo'llar bilan o'zaro to'qnashishi mumkin va bir-biriga nisbatan pozitsiyalari bo'yicha tasniflanishi mumkin.[2][5][6][7][8]
- Bir tomonlama yoki tandem qoplash: 3' bitta genning uchi bilan 5' xuddi shu yo'nalishdagi boshqa genning oxiri. Ushbu tartib → → yozuvi bilan ramziylashtirilishi mumkin, bu erda o'qlar boshidan oxirigacha o'qish doirasini bildiradi.
- Konvergent yoki oxirigacha qoplash: 3' ikkala genning uchlari bir-biriga qarama-qarshi iplar ustiga yopishgan. Buni → ← deb yozish mumkin.
- Turli xil yoki dumaloq qoplash: 5' ikkala genning uchlari bir-biriga qarama-qarshi iplar ustiga yopishgan. Buni ← → shaklida yozish mumkin.
Bir-birini qoplaydigan genlarni ham tasniflash mumkin fazalar, ularning qarindoshlarini tavsiflovchi o'qish ramkalari:[2][5][6][7][8]
- Faza ichidagi bir-birining ustiga chiqish umumiy ketma-ketliklar bir xil o'qish doirasidan foydalanganda paydo bo'ladi. Bu "0 bosqich" deb ham nomlanadi. 0 faza ustma-ust keladigan bir yo'nalishli genlar alohida gen deb qaralmaydi, aksincha muqobil boshlash saytlari bir xil gen.
- Fazadan tashqari bir-biriga to'g'ri keladi umumiy ketma-ketliklar turli xil o'qish ramkalarini ishlatganda paydo bo'ladi. Bu o'qish ramkalari 1 yoki 2 nukleotid bilan almashtirilganligiga qarab, "1-bosqich" yoki "2-bosqich" da sodir bo'lishi mumkin. Chunki a kodon uchta nukleotidga teng, uchta nukleotidning ofseti fazali, faza 0 kadridir.
Evolyutsiya
Bir-birining ustiga tushadigan genlar, ayniqsa tez rivojlanayotgan genomlarda, masalan, tez-tez uchraydi viruslar, bakteriyalar va mitoxondriya. Ular uchta yo'l bilan paydo bo'lishi mumkin:[9]
- Mavjudni kengaytirish orqali ochiq o'qish doirasi (ORF) a oqimining yo'qolishi tufayli quyi oqimda tutashgan genga aylanadi kodonni to'xtatish;
- An yo'qotilishi tufayli mavjud bo'lgan ORF oqimining yuqori oqimiga qo'shilib genga kengayishi bilan boshlash kodoni;
- Mavjud bo'lgan yangi ORFning yaratilishi bo'yicha a nuqta mutatsiyasi.
Ko'pgina genlarni kodlash uchun bir xil nukleotidlar ketma-ketligidan foydalanish ta'minlanishi mumkin evolyutsion kamayishi tufayli afzallik genom hajmi va uchun imkoniyat tufayli transkripsiyaviy va tarjima birgalikda tartibga solish bir-birini qoplaydigan genlarning.[6][10][11][12] Genlarning ustma-ust tushishi bir-birini qoplagan mintaqalar ketma-ketligida yangi evolyutsion cheklovlarni keltirib chiqaradi.[8][13]
Yangi genlarning kelib chiqishi

1977 yilda, Per-Pol Grasse juftlikdagi genlardan biri kelib chiqishi mumkin deb taklif qildi de novo o'zgaruvchan o'qish ramkalarida yangi ORFlarni joriy qilish uchun mutatsiyalar orqali; u mexanizmni quyidagicha tasvirlab berdi ortiqcha bosib chiqarish.[15]:231 Keyinchalik bu tomonidan tasdiqlangan Susumu Ohno, ushbu mexanizmda paydo bo'lishi mumkin bo'lgan nomzod genini aniqlagan.[16] Shu tarzda kelib chiqqan ba'zi de novo genlari bir-birini takrorlamasligi mumkin, ammo subfunktsionalizatsiya quyidagi genlarning takrorlanishi[3], tarqalishiga hissa qo'shadi yetim genlar. Qatnashgan genlar juftligining qaysi a'zosi yoshroq ekanligini aniqlash mumkin bioinformatik jihatdan yoki yanada cheklangan filogenetik tarqatish yoki kamroq optimallashtirilgan kodondan foydalanish.[14][17][18] Juftlikning yosh a'zolari yuqoriroq bo'lishadi ichki tuzilish buzilishi keksa a'zolarga qaraganda, lekin keksa a'zolar boshqa oqsillarga qaraganda tartibsizroqdir, ehtimol bu bir-birining ustiga chiqib ketish evolyutsiyasi cheklovlarini yumshatish usuli sifatida.[17] Qatlamlar allaqachon yuqori tartibsizlikka ega bo'lgan oqsillardan kelib chiqishi ehtimoli yuqori.[17]
Taksonomik taqsimot

Bir-birining ustiga chiqadigan genlar umuman paydo bo'ladi hayot sohalari, ammo turli xil chastotalar bilan. Ular, ayniqsa, keng tarqalgan virusli genomlar.
Viruslar

Bir-birining ustiga chiqadigan genlarning mavjudligi birinchi marta viruslarda aniqlangan; hozirgi kungacha ketma-ket birinchi DNK genomi bakteriyofag ΦX174, bir nechta misollarni o'z ichiga olgan.[19] OLG-ning yana bir misoli - 2020 yil oktyabr oyida ORF3d gen topilgan Covid19 virusi genom bu omil bo'lishi mumkin SARS-CoV-2 pandemiyasi.[1][21] Bir-birining ustiga tushadigan genlar ayniqsa keng tarqalgan virusli genomlar.[14] Ba'zi tadkikotlar bu kuzatuvni sabab deb biladi selektiv bosim a ichida genomni qadoqlashning jismoniy cheklovlari vositasida kichik genom o'lchamlari tomon virusli kapsid, ayniqsa biri ikosahedral geometriya.[22] Shu bilan birga, boshqa tadqiqotlar ushbu xulosaga qarshi chiqadi va virus genomlaridagi bir-birining ustiga chiqib ketish tarqalishining ko'payishi viruslar genlarining evolyutsion kelib chiqishi sifatida ortiqcha bosishni aks ettirish ehtimoli ko'proq ekanligini ta'kidlaydi.[23] Overprinting - bu keng tarqalgan manba de novo viruslardagi genlar.[18]
Bosib chiqarilgan virusli genlarni o'rganish shuni ko'rsatadiki, ularning oqsil mahsulotlari aksariyat aksessuar oqsillari bo'lib qoladi muhim viruslarning ko'payishiga, lekin hissa qo'shadi patogenlik. Bosib chiqarilgan oqsillar odatda g'ayrioddiy narsalarga ega aminokislota ichki darajadagi taqsimotlar va yuqori darajalar tartibsizlik.[24] Ba'zi hollarda ortiqcha bosilgan oqsillar aniq belgilangan, ammo yangi, uch o'lchovli tuzilishga ega;[25] bitta misol RNK susaytiruvchi supressor p19 ichida topilgan Tombusviruslar Ikkala romani ham bor oqsil qatlami va tanib olishda yangi majburiy rejim siRNAlar.[18][20][26]
Prokaryotlar
Genlarning taxminiy taxminlari bakterial genomlar odatda bakteriyalar genlarining uchdan bir qismi ustma-ust tushganligini aniqlaydilar, garchi odatda faqat bir necha tayanch juftlari.[6][27][28] Bakteriyalar genomlaridagi bir-birining ustiga o'ralganligi bo'yicha olib borilgan ko'plab tadqiqotlar, o'zaro faoliyat funktsiyani bajaradigan dalillarni topadi genlarni tartibga solish, bir-birining ustiga tushgan genlarga ruxsat berish transkripsiyaviy ravishda va tarjimaviy birgalikda tartibga solinadi.[6][12] Prokaryotik genomlarda bir yo'nalishda bir-biri bilan qoplanish eng ko'p uchraydi, ehtimol qo'shni prokaryotik genlarning yo'nalishni birgalikda bo'lish tendentsiyasi tufayli.[6][8][5] Bir yo'nalishli takrorlanishlar orasida o'qish doirasidagi bir nukleotidli ofset bilan (masalan, 1-faza) uzoq qoplamalar ko'proq o'qiladi va 2-bosqichda qisqa tutashishlar ko'proq o'qiladi.[28][29] 60 dan katta uzunlikdagi ustma-ust tushishlar tayanch juftliklari konvergent genlar uchun ko'proq tarqalgan; ammo, taxminiy uzunlikdagi qoplamalar juda yuqori ko'rsatkichlarga ega noto'g'ri izoh.[30] Bakterial genomlarda uzoq vaqt bir-birining ustiga chiqib ketishining ishonchli tasdiqlangan misollari kam uchraydi; yaxshi o'rganilgan model organizm Escherichia coli, faqat to'rtta gen juftligi uzun, ortiqcha bosilgan bir-biriga o'xshashligi sababli yaxshi tasdiqlangan.[31]
Eukaryotlar
Prokaryotik genomlar bilan taqqoslaganda, eukaryotik genomlar ko'pincha izohlanmagan va shu bilan haqiqiy bir-biriga o'xshashligini aniqlash nisbatan qiyin.[18] Shu bilan birga, tasdiqlangan genlarning bir-biriga o'xshashligi misollari turli xil ökaryotik organizmlarda, jumladan, sichqonlar va odamlar kabi sutemizuvchilarda qayd etilgan.[32][33][34][35] Eukariotlar prokaryotlardan bir-birining ustiga chiqib ketish turlarining tarqalishi bilan farq qiladi: bir tomonlama (ya'ni bir xil zanjirli) qoplamalar prokaryotlarda, qarama-qarshi yoki antiparallel-zanjirli qoplamalar esa ko'proq eukaryotlarda uchraydi. Qarama-qarshi chiziqlar orasida konvergent yo'nalish eng keng tarqalgan.[33] Eukaryotik genlarning bir-birini qoplashi bo'yicha olib borilgan ko'pgina tadqiqotlar shuni ko'rsatdiki, bir-birining ustiga yopishgan genlar genomik qayta tashkil etilishga yaqin turlarda ham keng tarqalgan bo'lib, shuning uchun ham har doim ham yaxshi saqlanib qolinmaydi.[34][36] Eski yoki kamroq taksonomik cheklangan genlar bilan qoplanish ham kelib chiqishi mumkin bo'lgan genlarning umumiy xususiyatidir de novo ma'lum bir ökaryotik nasabda.[34][37][38]
Adabiyotlar
- ^ a b Nelson, Chayz V; va boshq. (1 oktyabr 2020). "SARS-CoV-2 pandemiyasining omili sifatida dinamik ravishda rivojlanayotgan romanning bir-birini qoplaydigan geni". eLife. 9. doi:10.7554 / eLife.59633. PMC 7655111. PMID 33001029. Olingan 11 noyabr 2020.
- ^ a b v Y. Fukuda, M. Tomita va T. Vashio (1999). "Genomlaridagi ustma-ust keladigan genlarni qiyosiy o'rganish Mikoplazma genitalium va Mikoplazma pnevmoniyasi". Nuklein kislotalari rez. 27 (8): 1847–1853. doi:10.1093 / nar / 27.8.1847. PMC 148392. PMID 10101192.
- ^ a b Kiz, PK; Gibbs, A (1992 yil 15 oktyabr). "Genlarning kelib chiqishi:" katta portlash "yoki uzluksiz ijodmi?". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 89 (20): 9489–93. Bibcode:1992 yil PNAS ... 89.9489K. doi:10.1073 / pnas.89.20.9489. PMC 50157. PMID 1329098.
- ^ Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJ, Staden R, Young IG (aprel 1981). "Inson mitoxondriyal genomining ketma-ketligi va tashkil etilishi". Tabiat. 290 (5806): 457–465. Bibcode:1981 yil Noyabr.290..457A. doi:10.1038 / 290457a0. PMID 7219534. S2CID 4355527.
- ^ a b v Fukuda, Yoko; Nakayama, Yoichi; Tomita, Masaru (2003 yil dekabr). "Bakterial genomlarda bir-birining ustiga chiqadigan genlarning dinamikasi to'g'risida". Gen. 323: 181–187. doi:10.1016 / j.gene.2003.09.021. PMID 14659892.
- ^ a b v d e f Jonson Z, Chisholm S (2004). "Qatnashgan genlarning xususiyatlari mikrobial genomlarda saqlanib qoladi". Genom Res. 14 (11): 2268–72. doi:10.1101 / gr.2433104. PMC 525685. PMID 15520290.
- ^ a b Normark S .; Bergstrom S .; Edlund T.; Grundstrom T .; Jaurin B.; Lindberg F.P.; Olsson O. (1983). "Bir-birini qoplaydigan genlar". Genetika fanining yillik sharhi. 17: 499–525. doi:10.1146 / annurev.ge.17.120183.002435. PMID 6198955.
- ^ a b v d Rogozin, Igor B.; Spiridonov, Aleksey N.; Sorokin, Aleksandr V.; Bo'ri, Yuriy I.; Iordaniya, I.King; Tatusov, Roman L.; Koonin, Eugene V. (2002 yil may). "Prokaryotik genlarning ustma-ust tushishida tozalovchi va yo'naltirilgan tanlov". Genetika tendentsiyalari. 18 (5): 228–232. doi:10.1016 / S0168-9525 (02) 02649-5. PMID 12047938.
- ^ Krakauer, Devid C. (iyun 2000). "Qatnashgan genlarning barqarorligi va evolyutsiyasi". Evolyutsiya. 54 (3): 731–739. doi:10.1111 / j.0014-3820.2000.tb00075.x. PMID 10937248. S2CID 8818055.
- ^ Delaye, Luis; DeLuna, Aleksandr; Lazkano, Antonio; Bekerra, Arturo (2008). "Escherichia coli-da ortiqcha bosib chiqarish orqali yangi genning kelib chiqishi". BMC evolyutsion biologiyasi. 8 (1): 31. doi:10.1186/1471-2148-8-31. PMC 2268670. PMID 18226237.
- ^ Saxa, Deeya; Yem, Soumita; Panda, Arup; Ghosh, Tapash Chandra (2016 yil may). "Bir-birini qoplaydigan genlar: prokaryotik o'sish sur'atlarining sezilarli genomik korrelyatsiyasi". Gen. 582 (2): 143–147. doi:10.1016 / j.gene.2016.02.002. PMID 26853049.
- ^ a b Luo, Yingqin; Battistuzzi, Fabiya; Lin, Kui; Gibas, Sintiya (2013 yil 29-noyabr). "Salmonellalardagi bir-birining ustiga chiqib ketgan genlarning evolyutsion dinamikasi". PLOS ONE. 8 (11): e81016. doi:10.1371 / journal.pone.0081016. PMC 3843671. PMID 24312259.
- ^ Vey X.; Zhang, J. (2014 yil 31-dekabr). "Bir-birini qoplaydigan genlar bo'yicha tabiiy selektsiya kuchini baholashning oddiy usuli". Genom biologiyasi va evolyutsiyasi. 7 (1): 381–390. doi:10.1093 / gbe / evu294. PMC 4316641. PMID 25552532.
- ^ a b v Pavesi, Anjelo; Magiorkinis, Gkikas; Karlin, Devid G.; Wilke, Claus O. (2013 yil 15-avgust). "De Novo-dan ortiqcha bosmadan kelib chiqqan virusli oqsillarni kodon yordamida aniqlash mumkin: Deltaretroviruslarning" Gen pitomnikiga "murojaat qilish". PLOS hisoblash biologiyasi. 9 (8): e1003162. doi:10.1371 / journal.pcbi.1003162. PMC 3744397. PMID 23966842.
- ^ Grass, Per-Pol (1977). Tirik organizmlar evolyutsiyasi: transformatsiyaning yangi nazariyasining dalili. Akademik matbuot. ISBN 9781483274096.
- ^ Ohno, S (1984 yil aprel). "Oldindan, ichki takrorlanadigan kodlash ketma-ketligining muqobil o'qish doirasidan noyob ferment tug'ilishi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 81 (8): 2421–5. Bibcode:1984PNAS ... 81.2421O. doi:10.1073 / pnas.81.8.2421. PMC 345072. PMID 6585807.
- ^ a b v Uillis, Sara; Masel, Joanna (2018 yil 19-iyul). "Gen tug'ilishi, bir-birining ustiga chiqadigan genlar tomonidan kodlangan tuzilish buzilishiga hissa qo'shadi". Genetika. 210 (1): 303–313. doi:10.1534 / genetika.118.301249. PMC 6116962. PMID 30026186.
- ^ a b v d Sabat, N .; Vagner, A .; Karlin, D. (2012 yil 19-iyul). "Virusli oqsillarning evolyutsiyasi De Novo tomonidan ortiqcha bosmadan kelib chiqqan". Molekulyar biologiya va evolyutsiya. 29 (12): 3767–3780. doi:10.1093 / molbev / mss179. PMC 3494269. PMID 22821011.
- ^ a b Sanger, F.; Air, G. M .; Barrel, B. G.; Braun, N. L .; Kulson, A. R .; Fiddes, J. C .; Xetchison, C. A .; Slocombe, P. M.; Smit, M. (1977). "Bakteriyofagning nukleotidlar ketma-ketligi ΦX174 DNK". Tabiat. 265 (5596): 687–95. Bibcode:1977 yil natur.265..687S. doi:10.1038 / 265687a0. PMID 870828. S2CID 4206886.
- ^ a b Ye, Keqiong; Malinina, Lyusi; Patel, Dinshaw J. (2003 yil 3-dekabr). "Kichik interferentsiyali RNKni virusli RNK susaytirishning supressori tomonidan tan olinishi". Tabiat. 426 (6968): 874–878. Bibcode:2003 yil natur.426..874Y. doi:10.1038 / tabiat02213. PMC 4694583. PMID 14661029.
- ^ Dockrill, Peter (11 noyabr 2020). "Olimlar shunchaki SARS-CoV-2-da sirli ravishda yashiringan" gen ichida gen "topdilar". ScienceAlert. Olingan 11 noyabr 2020.
- ^ Chiriko, N .; Vianelli, A .; Belshaw, R. (2010 yil 7-iyul). "Nima uchun genlar viruslarda bir-biriga to'g'ri keladi". Qirollik jamiyati materiallari B: Biologiya fanlari. 277 (1701): 3809–3817. doi:10.1098 / rspb.2010.1052. PMC 2992710. PMID 20610432.
- ^ Brendlar, Nadav; Linial, Mixal (2016 yil 21-may). "Virus dunyosida genlarning bir-birining ustiga chiqishi va cheklovlari". Biologiya to'g'ridan-to'g'ri. 11 (1): 26. doi:10.1186 / s13062-016-0128-3. PMC 4875738. PMID 27209091.
- ^ Rancurel, C .; Xosravi, M.; Dunker, A. K .; Romero, P. R .; Karlin, D. (2009 yil 29-iyul). "Bir-birining ustiga tushgan genlar g'ayrioddiy ketma-ketlik xususiyatlariga ega oqsillarni ishlab chiqaradi va De Novo oqsillarini yaratish to'g'risida tushuncha beradi". Virusologiya jurnali. 83 (20): 10719–10736. doi:10.1128 / JVI.00595-09. PMC 2753099. PMID 19640978.
- ^ Abroi, Aare (2015 yil 1-dekabr). "Virosfera va mezbon munosabatlarining oqsil domeniga asoslangan ko'rinishi". Biochimie. 119: 231–243. doi:10.1016 / j.biochi.2015.08.008. PMID 26296474.
- ^ Vargason, Jeffri M; Szittya, Dyurji; Burgyan, Jozef; Hall, Traci M.Tanaka (2003 yil dekabr). "RNK susaytiruvchi supressor tomonidan siRNK ni tanlab tanib olish". Hujayra. 115 (7): 799–811. doi:10.1016 / S0092-8674 (03) 00984-X. PMID 14697199. S2CID 12993441.
- ^ Xuvet, Maksim; Stumpf, Maykl PH (2014 yil 1-yanvar). "Bir-birini takrorlaydigan genlar: genlarning evolyutsiyasi oynasi". BMC Genomics. 15 (1): 721. doi:10.1186/1471-2164-15-721. ISSN 1471-2164. PMC 4161906. PMID 25159814.
- ^ a b Cock, Peter J. A.; Uitvort, Devid E. (2007 yil 19 mart). "Genlarning bir-birining ustiga chiqishi evolyutsiyasi: Prokaryotik ikki komponentli tizim genlaridagi o'qish doirasining nisbiy tomoni". Molekulyar evolyutsiya jurnali. 64 (4): 457–462. doi:10.1007 / s00239-006-0180-1. PMID 17479344. S2CID 21612308.
- ^ Fonseka, M. M.; Xarris, D. J .; Posada, D. (2013 yil 5-noyabr). "Bir tomonlama prokaryotik ustma-ust keladigan genlarning kelib chiqishi va uzunligini taqsimlash". G3: Genlar, Genomlar, Genetika. 4 (1): 19–27. doi:10.1534 / g3.113.005652. PMC 3887535. PMID 24192837.
- ^ Palleja, Albert; Harrington, Eoghan D; Bork, tengdosh (2008). "Prokaryotik genomlarda katta genlar bir-biriga to'g'ri keladi: funktsional cheklovlarmi yoki noto'g'ri taxminlarmi?". BMC Genomics. 9 (1): 335. doi:10.1186/1471-2164-9-335. PMC 2478687. PMID 18627618.
- ^ Fellner, Lea; Simon, Svenja; Sherling, nasroniy; Witting, Maykl; Shober, Sffen; Polte, Kristin; Shmitt-Kopplin, Filippe; Keim, Daniel A.; Sherer, Zigfrid; Noyhaus, Klaus (2015 yil 18-dekabr). "Evolyutsion ortiqcha bosim bilan bakterial oqsil kodlovchi, ustma-ust yetim genning yaqinda paydo bo'lganligi to'g'risida dalillar". BMC evolyutsion biologiyasi. 15 (1): 283. doi:10.1186 / s12862-015-0558-z. PMC 4683798. PMID 26677845.
- ^ Maklisagt, Aoife; Gerzoni, Daniele (2015 yil 31-avgust). "Kodlamaydigan ketma-ketlikdagi yangi genlar: evrokotik evolyutsion innovatsiyada de novo oqsil kodlovchi genlarning roli". Qirollik jamiyatining falsafiy operatsiyalari B: Biologiya fanlari. 370 (1678): 20140332. doi:10.1098 / rstb.2014.0332. PMC 4571571. PMID 26323763.
- ^ a b C. Sanna, V. Li va L. Zhang (2008). "Odam va sichqon genomidagi bir-birining ustiga chiqadigan genlar". BMC Genomics. 9 (169): 169. doi:10.1186/1471-2164-9-169. PMC 2335118. PMID 18410680.
- ^ a b v Makalowska, Izabela; Lin, Chiao-Feng; Ernandes, Krisitina (2007). "Genning tug'ilishi va o'limi umurtqali hayvonlarda bir-biriga to'g'ri keladi". BMC evolyutsion biologiyasi. 7 (1): 193. doi:10.1186/1471-2148-7-193. PMC 2151771. PMID 17939861.
- ^ Veeramachaneni, V. (2004 yil 1-fevral). "Sutemizuvchilarning ustma-ust tushgan genlari: qiyosiy istiqbol". Genom tadqiqotlari. 14 (2): 280–286. doi:10.1101 / gr. 1590904. PMC 327103. PMID 14762064.
- ^ Bexura, Susanta K; Severson, Devid V (2013). "Aedes aegypti genlarining ustma-ust tushishi: Anopheles gambiae va boshqa hasharotlar ortologlari bilan taqqoslash evolyutsiyasi". BMC evolyutsion biologiyasi. 13 (1): 124. doi:10.1186/1471-2148-13-124. PMC 3689595. PMID 23777277.
- ^ Merfi, Daniel N.; Maklisagt, Aoife; Karmel, Liran (2012 yil 21-noyabr). "Murin kemiruvchilarida oqsillarni kodlovchi genlarning De Novo kelib chiqishi". PLOS ONE. 7 (11): e48650. Bibcode:2012PLoSO ... 748650M. doi:10.1371 / journal.pone.0048650. PMC 3504067. PMID 23185269.
- ^ Nouuls, D. G .; McLysaght, A. (2 sentyabr 2009). "Odamning oqsillarni kodlovchi genlarining so'nggi de novo kelib chiqishi". Genom tadqiqotlari. 19 (10): 1752–1759. doi:10.1101 / gr.095026.109. PMC 2765279. PMID 19726446.