Sefalopod - Cephalopod
| Sefalopod | |
|---|---|
 | |
| Bigfin rif kalmar (Sepioteuthis Lessiana) | |
| Ilmiy tasnif | |
| Qirollik: | Animalia |
| Filum: | Molluska |
| Sinf: | Tsefalopoda Kuvier, 1797 |
| Subklasslar | |
A sefalopod /ˈsɛfələpɒd/ ning har qanday a'zosi mollyuska sinf Tsefalopoda /sɛfəˈlɒpədə/ (Yunoncha ko'plik aλόπoz, kefalopodlar; "bosh-oyoq")[2] kabi a Kalmar, sakkizoyoq, muzqaymoq, yoki nautilus. Bu faqat dengiz hayvonlari bilan tavsiflanadi ikki tomonlama tana simmetriyasi, taniqli bosh va to'plam qo'llar yoki chodirlar (mushak gidrostatlari ) ibtidoiy mollyuskaning oyoqidan o'zgartirilgan. Baliqchilar ba'zan sefalopodlarni chaqirishadi "siyoh baliq, "ularning chayqalishning umumiy qobiliyatiga ishora qilmoqda siyoh. Sefalopodlarni o'rganish - bu filial malakologiya sifatida tanilgan teutologiya.
Davrida sefalopodlar dominant bo'lib qoldi Ordovik ibtidoiy bilan ifodalangan davr nautiloidlar. Endi sinf ikkita, faqat bir-biriga yaqin bo'lgan, mavjud kichik sinflar: Coleoidea o'z ichiga oladi ahtapot, Kalmar va muzqaymoq; va Nautiloidea tomonidan ifodalangan Nautilus va Allonautilus. Coleoidea-da mollyuskan qobig'i ichki holatga keltirilgan yoki yo'q, Nautiloidea-da esa tashqi qobiq qoladi. 800 ga yaqin yashash turlari sefalopodlar aniqlandi. Ikki muhim yo'q bo'lib ketgan taksonlar ular Ammonoida (ammonitlar) va Belemnoidea (belemnitlar).
Tarqatish


To'g'ri: Ahtapot (Benthoctopus sp.) bo'yicha Devidson Seamount 2,422 m chuqurlikda
800 dan ortiq mavjud sefalopod turlari,[3] garchi yangi turlarni ta'riflashni davom ettirsa. Taxminan 11000 yo'q bo'lib ketgan taksonlar tasvirlangan, garchi sefalopodlarning yumshoq tanasi ularni osongina toshbo'ron qilinmasligini anglatadi.[4]
Sefalopodlar Yerning barcha okeanlarida uchraydi. Ularning hech biri toqat qila olmaydi chuchuk suv, lekin qisqa kalmar, Lolliguncula brevis, topilgan Chesapeake Bay, toqat qilayotgani bilan ajralib turadigan qisman istisno sho'r suv.[5] Tsefalopodlar ko'plab biokimyoviy cheklovlar tufayli chuchuk suvda yashay olmaydilar va ularning 400+ million yillik hayotlarida hech qachon chuchuk suvlarning yashash joylariga kirishmagan.[6]
Sefalopodlar okean tubining katta qismini egallaydi tubsiz tekislik dengiz sathiga. Ularning xilma-xilligi ekvator yaqinida eng katta (xilma-xillikni o'rganish natijasida 11 ° N da to'rlarda olingan ~ 40 tur) va qutblarga qarab kamayadi (60 ° N da qo'lga kiritilgan ~ 5 tur).[7]
Biologiya
Asab tizimi va o'zini tutishi


To'g'ri: Gavayi bobtail kalmar, Evropima skoloplar, o'zini qumga ko'mib, faqat ko'zlar ochiq qoladi
Sefalopodlar keng tarqalgan bo'lib, eng aqlli hisoblanadi umurtqasizlar va yaxshi rivojlangan hissiyotlarga ega va katta miyalar (ulardan kattaroq gastropodlar ).[8] The asab tizimi sefalopodlar umurtqasizlarning eng murakkabidir[9][10] va ularning miya-tana-massa nisbati ular orasida bo'ladi endotermik va ektotermik umurtqali hayvonlar.[7]:14 Asirga tushgan sefalopodlar o'zlarining akvaryumlaridan chiqib, laboratoriya qavatining masofasini manevr qilib, boshqa akvariumga kirib, qisqichbaqalar bilan oziqlanish va o'z akvariumlariga qaytishlari bilan ham tanilgan.[11]
Miya a da himoyalangan xaftaga oid bosh suyagi. Gigant asab sefalopodning tolalari mantiya ko'p yillar davomida eksperimental material sifatida keng qo'llanilib kelinmoqda neyrofiziologiya; ularning katta diametri (etishmasligi tufayli miyelinatsiya ) ularni boshqa hayvonlar bilan taqqoslaganda o'rganishni nisbatan osonlashtiradi.[12]
Ko'p sefalopodlar ijtimoiy jonzotlardir; o'z turidan ajratilganda ba'zi turlari kuzatilgan shoaling baliq bilan.[13]
Ba'zi sefalopodlar havodan 50 m gacha masofani bosib o'tishga qodir. Sefalopodlar ayniqsa aerodinamik bo'lmasa-da, ushbu ta'sirchan diapazonlarga reaktiv harakatlanish orqali erishadilar; organizm havoda bo'lganida, huni ichidan suv chiqarilishi davom etmoqda.[14] Hayvonlar qanotlarini hosil qilish uchun suyaklari va chodirlarini yoyib, tana holati bilan ko'tarish kuchini faol ravishda boshqaradilar.[15] Bitta tur, Todarodes pacificus, individual tentacles o'rtasida shilimshiq plyonka bilan tekis fanat shaklida tentaklarning tarqalishi kuzatilgan[15][16] boshqasi esa, Sepioteuthis sepioidea, tentaklarni aylana shaklida joylashtirilganligi kuzatilgan.[17]
Sezgilar
Sefalopodlar rivojlangan ko'rish qobiliyatiga ega, ular bilan tortishish kuchini aniqlay olishadi statotsistlar va turli xil kimyoviy sezgi organlariga ega.[7]:34 Ahtapot atrof-muhitni o'rganish uchun qo'llarini ishlatadi va ularni chuqur idrok etish uchun ishlatishi mumkin.[7]
Vizyon


Aksariyat sefalopodlar yirtqichlar va o'ljalarni aniqlash va bir-biri bilan aloqa qilish uchun ko'rishga tayanadi.[18] Binobarin, sefalopodni ko'rish keskin: o'quv tajribalari shuni ko'rsatdiki oddiy ahtapot narsalarning yorqinligini, o'lchamini, shakli va gorizontal yoki vertikal yo'nalishini ajrata oladi. Morfologik konstruktsiya sefalopod ko'zlarga akula kabi bir xil ko'rsatkich beradi '; ammo, ularning tuzilishi bir-biridan farq qiladi, chunki sefalopodlarda shox parda yo'q va doimiy retinaga ega.[18] Sefalopodlarning ko'zlari ham tekisligiga sezgir qutblanish nur.[19] Ko'p boshqa sefalopodlardan farqli o'laroq, nautiluslar yaxshi ko'rishga ega emas; ularning ko'z tuzilishi juda rivojlangan, ammo qattiq moddasi yo'q ob'ektiv. Ular oddiy "teshik "Ko'z orqali suv o'tishi mumkin. Vizyon o'rniga hayvon ishlatadi deb o'ylashadi olfaktsiya uchun asosiy ma'no sifatida em-xashak, shuningdek, potentsial juftlarni topish yoki aniqlash.

Ajablanarlisi shundaki, ularning rangini o'zgartirish qobiliyatini hisobga olgan holda, barcha oktopodlar[20] va ko'pchilik sefalopodlar[21][22] deb hisoblanadi rangli ko'r. Koleoid sefalopodlar (sakkizoyoq, kalmar, mushukcha baliqlari) bitta fotoreseptor turiga ega va aniqlangan foton intensivligini bir nechta spektral kanallar bilan taqqoslash orqali rangni aniqlash qobiliyatiga ega emas. Qachon kamuflyaj o'zlari, ular o'zlarining xromatoforlarini yorqinlik va naqshni ko'rgan fonga qarab o'zgartirish uchun ishlatishadi, lekin ularning fonning o'ziga xos rangiga mos kelish qobiliyati, masalan, hujayralardan kelib chiqishi mumkin. iridoforlar va leykoforlar atrofdan yorug'likni aks ettiradi.[23] Shuningdek, ular tanada vizual pigmentlarni ishlab chiqaradi va yorug'lik darajasini bevosita tanasidan sezishi mumkin.[24] Dalillar rangni ko'rish da topilgan ko'pikli kalamar (Watasenia sintillanlari),[21][25] uchta aniqlik yordamida rang ko'rishga erishadi setchatka molekulalar: A1, yashil-ko'k rangga eng sezgir (484 nm); A2, ko'k-yashil ranggacha (500 nm) va A4, ko'kga (470 nm) bog'lab turadi opsin.[26]
2015 yilda sefalopodlarda spektral kamsitishning yangi mexanizmi tasvirlangan. Bu ekspluatatsiyaga asoslanadi xromatik aberratsiya (fokus uzunligiga to'lqin uzunligiga bog'liqligi). Raqamli modellashtirish shuni ko'rsatadiki, xromatik aberratsiya tasvirning aniqligini akkomodatsiyaga bog'liqligi orqali foydali xromatik ma'lumot berishi mumkin. Bosh qafas oyoqlarida o'qdan tashqari tirqish va halqasimon shakllar bu qobiliyatni oshiradi.[27]
Fotorezeptsiya
2015 yilda sefalopod xromatoforlari fotosensitiv ekanligini ko'rsatadigan molekulyar dalillar nashr etildi; teskari transkripsiya polimeraza zanjiri reaktsiyalari (RT-PCR) aniqlandi stenogrammalar kodlash rodopsin va retinoxrom retinalar va terining ichida longfin qirg'oqbo'yi (Doryteuthis pealeii), va oddiy kotletfish (Sepia officinalis) va keng kulfat baliqlari (Sepiya latimanus ). Mualliflarning ta'kidlashicha, bu sefalopod dermal to'qimalari nurga javob berish uchun molekulalarning kerakli birikmasiga ega bo'lishining birinchi dalilidir.[28]
Eshitish
Ba'zi kalmarlarning ovozini ularning yordamida aniqlaydiganligi ko'rsatilgan statotsistlar.[29]
Nurdan foydalanish

Ko'p sefalopodlar yorug'lik bilan ta'sir o'tkazadigan terining tarkibiy qismlariga ega. Bunga o'z ichiga olishi mumkin iridoforlar, leykoforlar, xromatoforlar va (ba'zi turlarda) fotoforlar. Xromatoforlar rangli pigment hujayralari bo'lib, ular rang va naqsh hosil qilish uchun mos ravishda kengayadi va qisqaradi, ular hayratlanarli modalarda foydalanishlari mumkin.[7][28] Kamuflyajni o'zlarining fonlari bilan ta'minlash bilan bir qatorda ba'zi sefalopodlar bioluminesce, soyalarini yashirish uchun pastga nur sochib turadi quyida yashirinishi mumkin bo'lgan har qanday yirtqichlardan.[7] The biolyuminesans bakterial simbiontlar tomonidan ishlab chiqariladi; mezbon sefalopod bu organizmlar tomonidan ishlab chiqarilgan yorug'likni aniqlashga qodir.[30] Biyolüminesans o'ljani jalb qilish uchun ham ishlatilishi mumkin va ba'zi turlar juftlarni hayratda qoldirish, hayratda qoldiradigan yirtqichlar yoki hatto bir-birlari bilan muloqot qilish uchun rangli displeylardan foydalanadilar.[7]
Rang berish
Sefalopodlar ranglarini va naqshlarini millisekundalarda o'zgartirishi mumkin signal berish (ikkalasi ham tur ichida va uchun ogohlantirish ) yoki faol kamuflyaj,[7] ularning xromatoforlari kengaygan yoki qisqarganligi sababli.[31] Garchi rang o'zgarishi, asosan, ko'rish qobiliyatiga bog'liq bo'lsa-da, teri hujayralari, xususan, dalillar mavjud xromatoforlar, yorug'likni aniqlay oladi va yorug'lik sharoitlariga ko'zdan mustaqil ravishda moslasha oladi.[32] Sefalopodlar mushaklar singari xromatoforlardan foydalanishlari mumkin, shuning uchun ham ular terining tusini xuddi ular kabi tez o'zgartirishi mumkin. Rang o'zgarishi odatda qirg'oqqa yaqin turlarda ochiq okeanda yashovchilarga qaraganda kuchliroq bo'lib, ularning funktsiyalari cheklangan. buzadigan kamuflyaj.[7]:2 Ahtapotlarning aksariyati o'zlarining fonida tanlangan tuzilmalarni taqlid qilishadi, aksincha ularning to'liq fonining tarkibiy rangiga aylanishadi.[33]
Dastlabki davrda boshlangan sefalopod qoldiqlarida asl rangning dalillari aniqlangan Siluriya; bu ortokonik shaxslar kamuflyaj vazifasini o'tagan deb hisoblangan konsentrik chiziqlarni tug'dirgan.[34] Devoniyalik sefalopodlar noma'lum funktsiyaga ega bo'lgan yanada murakkab rang naqshlariga ega.[35]
Siyoh
Bundan mustasno Nautilidae va turlari sakkizoyoq ga tegishli suborder Sirna,[36] ma'lum bo'lgan barcha sefalopodlarda siyoh xaltasi bor, ular yordamida qorong'i siyoh bulutini chalkashtirib yuborish mumkin yirtqichlar.[20] Ushbu sumka orqa mushakning kengaytmasi sifatida paydo bo'lgan mushak sumkasidir. U ichak ostida yotadi va anusga ochiladi, uning tarkibiga deyarli toza bo'ladi melanin - chayqalishi mumkin; uning voronka tagiga yaqinligi, siyohni chiqarilgan suv bilan taqsimlashni anglatadi, chunki sefalopod uning reaktiv harakatlanishidan foydalanadi.[20] Chiqarilgan melanin buluti, odatda, chiqarib yuborilganda aralashtiriladi mukus mantiyaning boshqa joylarida ishlab chiqarilgan va shuning uchun qalin bulutni hosil qiladi, natijada yirtqich hayvonning ingl. (va ehtimol xemosensor) buzilishi tutunli ekran. Shu bilan birga, yanada murakkab xatti-harakatlar kuzatilgan bo'lib, unda sefalopod bulutni chiqarib yuboradi, shilimshiq miqdori ko'proq bo'lib, uni chiqarib yuborgan sefalopodga o'xshaydi (bu aldanish "deb ataladi Psevdomorf ). Ushbu strategiya tez-tez yirtqichning tezda ketayotgan o'ljasiga emas, balki psevdomorfga hujum qilishiga olib keladi.[20] Qo'shimcha ma'lumot olish uchun qarang Ink xatti-harakatlari.
Sefalopodlarning siyoh xaltasi "siyoh baliqlari" ning umumiy nomini keltirib chiqardi,[37] ilgari qalam bilan siyoh baliqlari.[38]
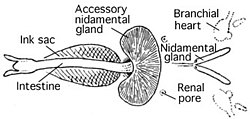

Qon aylanish tizimi
Sefalopodlar - qon aylanish sistemasi yopiq bo'lgan yagona mollyuskalar. Koleoidlar ikkita gill bor qalblar (shuningdek, nomi bilan tanilgan filial yuraklari ) ning kapillyarlari orqali qonni harakatga keltiruvchi gilzalar. Keyin bitta tizimli yurak butun kislorod bilan qonni butun tanasi orqali pompalaydi.[39]
Ko'pgina mollyuskalar singari, sefalopodlar ham foydalanadilar gemosiyanin, o'rniga mis tarkibidagi oqsil gemoglobin, kislorodni tashish uchun. Natijada, ularning qoni kislorodsizlanganda rangsiz va havo ta'sirida ko'k rangga aylanadi.[40]
Nafas olish
Sefalopodlar organizmning tomiga yopishib olgan gilzalari orqali suvni majburlab, dengiz suvi bilan gazlarni almashadilar.[41]:488[42] Gillalarning tashqi qismida mantiya bo'shlig'iga suv kiradi va mantiya bo'shlig'ining kirish joyi yopiladi. Mantiya qisqarganda, mantiya bo'shlig'i va huni o'rtasida joylashgan gilzalar orqali suv majburan o'tkaziladi. Suvni huni orqali chiqarib yuborish reaktiv harakatlanishni kuchaytirish uchun ishlatilishi mumkin. Boshqa mollyuskalarga qaraganda ancha samarali bo'lgan gilzalar mantiya bo'shlig'ining ventral yuzasiga biriktirilgan.[42]Turmush tarzi bilan bog'liq gil o'lchamlari bilan savdo-sotiq mavjud. Tez tezlikka erishish uchun gilzalar kichik bo'lishi kerak - energiya zarur bo'lganda suv ular orqali tezda o'tadi va ularning kichik o'lchamlarini qoplaydi. Biroq, ko'p vaqtini tubi bo'ylab sekin harakatlanishga sarflaydigan organizmlar, tabiiy ravishda, harakatlanish uchun bo'shliqlaridan ko'p suv o'tkazmaydilar; Shunday qilib, ular organizm harakatsiz bo'lsa ham, ularning gillalari orqali suvning doimiy yuvilishini ta'minlash uchun murakkab tizimlar bilan bir qatorda katta gillalarga ega.[41] Suv oqimi radiusli va dumaloq mantiya bo'shlig'i mushaklarining qisqarishi bilan boshqariladi.[43]
Sefalopodlarning gillalari mustahkam tolali oqsillar skeletlari bilan ta'minlanadi; mukopolisakkaridlarning etishmasligi bu matritsani xaftaga ajratib turadi.[44][45] Gilllar NH bilan birga ekskretsiyada ishtirok etadi deb o'ylashadi4+ K bilan almashtirildi+ dengiz suvidan.[42]
Harakatlanish va suzish qobiliyati

Aksariyat sefalopodlar harakatlanishi mumkin samolyot qo'zg'alish, bu baliqlar foydalanadigan quyruq qo'zg'alishi bilan taqqoslaganda juda ko'p energiya sarflaydigan usuldir.[46] A samaradorligi pervanel - haydovchi suv oqimi (ya'ni Froude samaradorligi ) a dan katta raketa.[47] Ning nisbiy samaradorligi reaktiv harakatlanish hayvonlarning kattalashishi bilan yanada kamayadi; paralarva voyaga etmaganlar va kattalar shaxslariga qaraganda ancha samarali.[48] Beri Paleozoy davri, bilan raqobat sifatida baliq samarali harakat omon qolish uchun juda muhim bo'lgan muhitni yaratdi, reaktiv harakatlanish orqa rol o'ynadi qanotlari va chodirlar tezlikni barqaror ushlab turish uchun ishlatiladi.[4]Reaktiv harakatlanish hech qachon harakatlanishning yagona usuli bo'lmasa ham,[4]:208 reaktivlar tomonidan ta'minlangan to'xtash-qo'zg'alish harakati yuqori tezlikda portlashni ta'minlash uchun foydali bo'lib qolmoqda - bu qo'lga kiritishda ham kam emas o'lja yoki qochish yirtqichlar.[4] Darhaqiqat, u sefalopodlarni eng tez dengiz umurtqasizlari qiladi,[7]:Muqaddimava ular ko'pchilik baliqlarni tezlashtirishi mumkin.[41]Jet fin harakati bilan to'ldiriladi; kalamarda har bir reaktiv uchqun chiqqanda qanotlari qoqilib, itarishni kuchaytiradi; keyinchalik ular samolyotlar o'rtasida uzaytiriladi (ehtimol cho'kib ketmaslik uchun).[48]Kislorodli suv ichkariga olinadi mantiya bo'shlig'i uchun gilzalar va bu bo'shliqning mushak qisqarishi orqali sarflangan suv giponoma, mantiyada katlama hosil qilgan. Ushbu organning orqa va oldingi uchlari orasidagi kattalik farqi organizm ishlab chiqarishi mumkin bo'lgan reaktiv tezlikni boshqaradi.[49] Hayvonning ma'lum bir massasi va morfologiyasi uchun organizmning tezligini aniq taxmin qilish mumkin.[50] Sefalopodlarning harakati odatda orqaga qarab ketadi, chunki suv old tomondan giponoma orqali tashqariga chiqarib yuboriladi, ammo uni turli yo'nalishlarga yo'naltirish orqali yo'nalishni biroz boshqarish mumkin.[51] Ba'zi sefalopodlar suvni chiqarib yuborishni o'q otish ovoziga o'xshash shovqin bilan birga olib borishadi, chunki ular potentsial yirtqichlarni qo'rqitish uchun ishlaydi.[52]
Sefalopodlar o'zlari topgan suvning dinamikasini o'zgartirganda (kattalashganda) kattalashishiga qaramay, shunga o'xshash harakatlanish usulidan foydalanadilar. Shunday qilib, ularning paralarva parraklaridan keng foydalanmaydi (past darajada unchalik samarasiz) Reynolds raqamlari ) va birinchi navbatda samolyotlarini o'zlarini yuqoriga ko'tarish uchun ishlatishadi, katta kattalar sefalopodlar esa kamroq samarali va qanotlariga suyanib suzishga moyil.[48]

Dastlabki sefalopodlar tanasini qobig'iga tortib, samolyotlar ishlab chiqargan deb o'ylashadi Nautilus bugun qiladi.[53] Nautilus shuningdek, uning huni to'lqinlari bilan reaktiv yaratishga qodir; suvning sekinroq oqishi suvdan kislorod olish uchun ko'proq mos keladi.[53] Jetning tezligi Nautilus ga qaraganda ancha sekinroq koleoidlar, ammo uni ishlab chiqarishda kamroq mushak va energiya ishtirok etadi.[54] Sefalopodlardagi reaktiv tortish asosan huni teshigining maksimal diametri (yoki, ehtimol, huni o'rtacha diametri) bilan boshqariladi.[55]:440 va mantiya bo'shlig'ining diametri.[56] Teshik kattaligining o'zgarishi ko'pincha oraliq tezlikda qo'llaniladi.[55] Amalga oshirilgan mutlaq tezlik sefalopodning haydab chiqarish uchun nafas olish talablari bilan cheklanadi; bu qabul qilish maksimal tezlikni sekundiga sakkizta tana uzunligiga qadar cheklaydi, bu ko'pchilik sefalopodlar ikkita voronka zarbasidan keyin erisha oladigan tezlik.[55] Suv nafaqat teshiklar orqali, balki huni orqali ham kirib, bo'shliqni to'ldiradi.[55] Squid bitta reaktiv harakat bilan bo'shliq ichidagi suyuqlikning 94% gacha chiqarishi mumkin.[47] Suv olish va chiqarib yuborishdagi tez o'zgarishlarni qondirish uchun teshiklar juda moslashuvchan va ularning o'lchamlarini yigirma marta o'zgartirishi mumkin; huni radiusi, aksincha, atigi 1,5 marta o'zgaradi.[55]
Ahtapotning ayrim turlari dengiz tubi bo'ylab yurishga ham qodir. Qisqichbaqalar va mushuk baliqlari qanotlarini silkitib, har qanday yo'nalishda qisqa masofani bosib o'tishlari mumkin muskul mantiya atrofida.
Aksariyat sefalopodlar suzadi (ya'ni neytral suzuvchi yoki deyarli shunday; aslida ko'pchilik sefalopodlar dengiz suvidan taxminan 2-3% zichroqdir[13]), ular bunga har xil yo'llar bilan erishadilar.[46]Ba'zilar, masalan Nautilus, gazning mantiya va qobiq orasidagi bo'shliqqa tarqalishiga imkon bering; boshqalar buyraklaridan toza suv oqishiga yo'l qo'yib, tana bo'shlig'idan zichroq sho'r suvni chiqarib yuborishadi;[46] boshqalari, ba'zi baliqlar singari, jigarda yog'larni to'playdi;[46] va ba'zi sakkizoyoqlarning jelatinli tanasi engilroq xlor ionlari almashtirish sulfat tanadagi kimyo.[46]
The Makrotritopus defilippiyoki qumda yashovchi ahtapot, qumda yashovchilarning rangini ham, suzish harakatlarini ham taqlid qilgani ko'rinib turibdi. Bothus lunatus yirtqichlardan saqlanish uchun. Sakkizoyoqlar tanalarini tekislab, qo'llarini orqaga qaytarib, pastroqlar bilan bir xil ko'rinishga ega bo'lishdi, shuningdek bir xil tezlik va harakatlar bilan harakat qilishdi.[57]
Ikki turdagi urg'ochilar - Ocythoe tuberculata va Haliphron atlanticus, haqiqiy rivojlandi suzish pufagi.[58]
Qobiq


Nautiluslar haqiqiy tashqi qobig'i bo'lgan yagona mavjud sefalopodlar. Biroq, barcha mollyuskali chig'anoqlar ektoderm (embrionning tashqi qatlami); yilda muzqaymoq (Sepiya spp.), masalan, embrion davrida ektodermaning invazinatsiyasi hosil bo'lib, natijada qobiq paydo bo'ladi (kotletbone ) bu kattalardagi ichki narsadir.[59] Xuddi shu narsa xitinozga tegishli gladius kalmar[59] va sakkizoyoqlar.[60] Cirrat sakkizoyoqlilarga ega kamar shaklidagi xaftaga oid suyanchiqlar,[61] ba'zan ularni "qobiq qoldig'i" yoki "gladius" deb atashadi.[62] The Incirrina bir juft novda shaklida bo'ladi uslublar yoki ichki qobiq qoldiqlari yo'q,[63] va ba'zi bir kalmarlarda gladius etishmaydi.[64] Qobiqlangan koleoidlar qoplama yoki hatto parafiletik guruh hosil qilmaydi.[65] The Spirula qobiq organik tuzilish sifatida boshlanadi va keyinchalik juda tez minerallashadi.[66] "Yo'qolgan" chig'anoqlar kaltsiy karbonat komponentining rezorbsiyasi natijasida yo'qolishi mumkin.[67]
Ahtapotning urg'ochi ayollari Argonauta ular yashaydigan ixtisoslashgan qog'ozga o'xshash ingichka tuxum sumkasini chiqaring va bu hayvon tanasiga yopishmagan va alohida evolyutsion kelib chiqishiga qaramay, bu "qobiq" deb qaraladi.
Qobiqlangan sefalopodlarning eng katta guruhi ammonitlar, yo'q bo'lib ketgan, ammo ularning qobiqlari juda keng tarqalgan fotoalbomlar.
Karbonatning cho'kishi mineralizatsiyalangan qobiqqa olib keladi, organik qobiq matritsasining kislotaligi bilan bog'liq ko'rinadi (qarang Mollyusk qobig'i ); qobiq hosil qiluvchi sefalopodlar kislotali matritsaga ega, kalamar gladiusi esa asosiy matritsaga ega.[68] Sefalopod tashqi devorining asosiy joylashuvi: tashqi (sferulitik) prizmatik qatlam, laminar (nakreus) va ichki prizmatik qatlam. Har bir qatlamning qalinligi taksonlarga bog'liq.[69] Zamonaviy sefalopodlarda Ca karbonat aragonit hisoblanadi. Boshqa mollyuskalar qobig'i yoki mercan skeletlariga kelsak, eng kichik ko'rinadigan birliklar tartibsiz yumaloq donachalardir.[70]


To'g'ri: Tentakular klubi haqida batafsil ma'lumot Abraliopsis morisi
Bosh qo'shimchalar
Sefalopodlar, nomidan ko'rinib turibdiki, mushaklarning qo'shimchalari boshlaridan va og'zini o'rab olgan. Ular oziqlantirishda, harakatchanlikda va hatto ko'payishda ishlatiladi. Yilda koleoidlar ularning soni sakkiz yoki o'nga teng. Kekterfish va kalmar kabi dekapodlarning besh jufti bor. Ikkisi uzunroq chodirlar, yirtqichni ushlashda faol ishtirok etmoqda;[1]:225 ular tez uzaytirishi mumkin (15 millisekunddan ozroq vaqt ichida)[1]:225). Yilda ulkan kalmar ularning uzunligi 8 metrga etishi mumkin. Ular kengaytirilgan, so'rg'ich bilan qoplangan klubda tugashi mumkin.[1]:225 Qisqa to'rt juftlik deb nomlanadi qo'llar, va qo'lga olingan organizmni ushlab turish va manipulyatsiya qilishda ishtirok etadi.[1]:225 Ularning ham og'ziga eng yaqin tomonida so'rg'ichlar bor; bular o'ljani ushlab turishga yordam beradi.[1]:226 Sakkizoyoqlilar atigi to'rtta juft so'rg'ich bilan qoplangan qo'llarga ega, garchi rivojlanish anormalliklari ifodalangan qo'llar sonini o'zgartirishi mumkin.[71]
Tentacle qalin markaziy asab simidan iborat (u har bir so'rg'ichni mustaqil ravishda boshqarishi uchun qalin bo'lishi kerak).[72] dumaloq va radial mushaklar bilan o'ralgan. Tentakning hajmi doimiy bo'lib qolganligi sababli, dumaloq mushaklarning qisqarishi radiusni pasaytiradi va uzunlikning tez o'sishiga imkon beradi. Odatda 70 foizga uzayishga kenglikni 23 foizga kamaytirish orqali erishiladi.[1]:227 Qisqa qo'llarda bunday imkoniyat yo'q.
Tentakning kattaligi bukkal kavitaning kattaligi bilan bog'liq; kattaroq, kuchliroq tentakllar o'lja tutishi mumkin, chunki undan kichik chaqishlar olinadi; ko'proq sonli, kichikroq chodirlar bilan o'lja butunlay yutiladi, shuning uchun og'iz bo'shlig'i kattaroq bo'lishi kerak.[73]
Tashqi qobiq nautilidlar (Nautilus va Allonautilus ) tartibida 90 barmoqqa o'xshash qo'shimchalar mavjud chodirlar, so'rg'ichlar etishmayotgan, ammo ularning o'rniga yopishqoq va qisman tortib olinadigan.
Oziqlantirish

Barcha tirik sefalopodlar ikki qismdan iborat tumshuq;[7]:7 ko'pchiligida a radula, aksariyat ahtapotda kamaygan va umuman yo'q bo'lsa ham Spirula.[7]:7[74]:110 Ular o'zlarining tentaklari bilan o'ljani ushlab, og'ziga tortib, undan tishlash orqali ovqatlanadilar.[20] Ularda zaharli ovqat hazm qilish sharbatlari aralashmasi bor, ularning ba'zilari simbiyotik suv o'tlari tomonidan ishlab chiqariladi, ular tuprik bezlaridan og'ziga tutilgan o'ljasiga chiqarib tashlaydi. Ushbu sharbatlar o'ljasining go'shtini suyak yoki qobiqdan ajratib turadi.[20] Tuprik bezining uchida kichik tish bor, uni ichkaridan hazm qilish uchun organizmga solib qo'yish mumkin.[20]
Ovqat hazm qilish bezining o'zi juda qisqa.[20] U to'rt elementdan iborat bo'lib, oziq-ovqat mahsuloti ichakka kirmasdan oldin hosil, oshqozon va ko'r ichak orqali o'tadi. Ovqat hazm qilishning ko'p qismi, shuningdek, ozuqaviy moddalarni singdirishi ovqat hazm qilish bezida, ba'zida jigar deb ataladi. Ichak va ovqat hazm qilish bezi o'rtasida ozuqa moddalari va chiqindilarni almashinuvi bezni oshqozon va ko'r ichakning tutashgan joyiga bog'laydigan juftlik birikmasi orqali amalga oshiriladi.[20] Ovqat hazm qilish bezidagi hujayralar to'g'ridan-to'g'ri ichak lümenine pigmentli ajralib chiqadigan kimyoviy moddalarni chiqarib yuboradi, so'ngra ular anus orqali uzun qorong'u torlar singari shilimshiq bilan bog'lanib, huni ichidan chiqarilgan suv yordamida chiqariladi.[20] Sefalopodlar tanadagi to'qimalarda yutilgan og'ir metallarni to'plashga moyil.[75]
Radula

Sefalopod radulasi to'qqiz taga qadar bo'lgan bir nechta nosimmetrik qatorlardan iborat[76] - fotoalbomlarda o'n uchta.[77] Organ ahtapotning ayrim turlarida kamaygan yoki hatto vestigial bo'lib, ularda yo'q Spirula.[77] Tishlar homodont (ya'ni bir qatorga o'xshash), heterodont (aks holda) yoki ctenodont (taroqsimon) bo'lishi mumkin.[77] Ularning balandligi, kengligi va sonlari turlar orasida o'zgaruvchan.[77] Tishlarning naqshlari takrorlanadi, lekin har bir satr oxirgisi bilan bir xil bo'lmasligi mumkin; masalan, ahtapotda ketma-ketlik har besh qatorda takrorlanadi.[77]:79
Tsefalopod radulalari Ordovik davridan qolgan qazilma konlardan ma'lum.[78] Ular odatda sefalopodning tanasi xonasida, odatda mandibular bilan birgalikda saqlanadi; ammo bu har doim ham shunday bo'lmasligi kerak;[79] ko'plab radulalar Mason Creek-da bir qator sozlamalarda saqlanib qolgan.[80]Radulalarni, odatda, ular qoldiqlarda saqlanib qolgan taqdirda ham, aniqlash qiyin, chunki tosh ularni ochish uchun ob-havo sharoitida va aniq tarzda yorilishi kerak; Masalan, radulalar 43 ta ammonit naslining to'qqiztasida topilgan,[81][tushuntirish kerak ] va ular hali ham ammonoid bo'lmagan shakllarda kam uchraydi: faqat mezozoydan oldingi uchta tur bittasiga ega.[78]
Ekskretator tizimi
Aksariyat sefalopodlar bitta juft juftga ega nefridiya. Filtrlangan azotli chiqindilar da ishlab chiqariladi perikardial bo'shlig'i filial yuraklari, ularning har biri nefridiyaga tor kanal orqali bog'langan. Kanal chiqarishni siydik pufagiga o'xshash buyrak xaltachasiga etkazib beradi, shuningdek filtratdan ortiqcha suvni qayta emiradi. Yon tomondan bir nechta o'sish vena kava buyrak xaltachasiga kirib, doimiy ravishda shishiradi va dumaloq yurak urishi bilan ajralib turadi. Ushbu harakat salgılanan chiqindilarni xaltachalarga quyish, teshik orqali mantiya bo'shlig'iga chiqarilishiga yordam beradi.[82]
Nautilus, odatdagidan tashqari to'rtta nefridiyaga ega, ularning hech biri perikardial bo'shliqlar bilan bog'lanmagan.
Ning qo'shilishi ammiak quruqlikdagi mollyuskalar va boshqa mollyuskan bo'lmagan nasllarda qobiq hosil bo'lishi uchun muhimdir.[83] Chunki oqsil (ya'ni go'sht) sefalopod dietasining asosiy tarkibiy qismidir, ko'p miqdorda ammoniy ionlari chiqindilar sifatida ishlab chiqariladi. Ushbu ortiqcha ammoniyni chiqarish bilan bog'liq bo'lgan asosiy organlar gilzalardir.[84] Chiqib ketish darajasi qobiqli sefalopodlarda eng past ko'rsatkichdir Nautilus va Sepiya ulardan foydalanish natijasida azot suzishni kuchaytirish uchun qobiqlarini gaz bilan to'ldirish.[84] Boshqa sefalopodlar ammoniydan xuddi shunday foydalanib, ionlari (kabi ammoniy xlorid ) ularning umumiy zichligini kamaytirish va suzuvchanlikni oshirish.[84]
Ko'paytirish va hayot aylanishi




Sefalopodlar turlarning xilma-xil guruhidir, lekin umumiy hayot tarixi xususiyatlariga ega, masalan, ular tez o'sish tezligiga va qisqa umrga ega.[85] Sterns (1992) tirik naslni ko'paytirish uchun yumurtlama hodisalari organizmning ekologik atrof-muhit omillariga bog'liq deb taxmin qildi. Sefalopodlarning aksariyati o'z avlodlariga ota-ona qaramog'ini bermaydilar, masalan, ahtapot, bu organizmga o'z avlodlarining omon qolish darajasini oshirishga yordam beradi.[85] Dengiz turlarining hayot aylanish jarayonlariga turli xil atrof-muhit sharoitlari ta'sir qiladi.[86] Sefalopod embrionining rivojlanishiga harorat, kislorod bilan to'yinganlik, ifloslanish, yorug'lik intensivligi va sho'rlanish katta ta'sir ko'rsatishi mumkin.[85] Ushbu omillar embrional rivojlanish tezligi va embrionlarning lyuklash muvaffaqiyatida muhim ahamiyatga ega. Sefalopodlarning reproduktiv tsiklida oziq-ovqat mavjudligi ham muhim rol o'ynaydi. Oziq-ovqat mahsulotlarining cheklanishi yumurtlamanın vaqtiga va ularning ishlashiga va o'sishiga ta'sir qiladi.[86] Dengiz turlari orasida yumurtlama vaqti va yumurtlama farq qiladi; Bu harorat bilan bog'liq, ammo sayoz suvdagi sefalopodlar sovuq oylarda urug'laydi, shunda avlod iliqroq haroratda chiqadi. Naslchilik bir necha kundan bir oygacha davom etishi mumkin.[85]
Jinsiy etuklik
Jinsiy jihatdan etuk va kattalar kattalikdagi sefalopodlar yumurtlamoq va ko'payishni boshlaydi. Genetik materialni keyingi avlodga o'tkazgandan so'ng, kattalar sefalopodlar o'ladi.[85] Erkak va ayol sefalopodlarda jinsiy etilish jinsiy bezlar va qo'shimcha bezlarning kattalashishi bilan ichki kuzatilishi mumkin.[87] Juftlik ayollarda jinsiy etilishning yomon ko'rsatkichi bo'ladi; ular reproduktiv yetishmovchilikda sperma olishlari va tuxumni urug'lantirishga tayyor bo'lguncha saqlashlari mumkin.[86] Erkaklar jinsiy etuk ayol uchun raqobatlashgandan ko'ra, etuk bo'lmagan urg'ochilar ishtirokida, juftlik oldidagi musobaqada ko'proq tajovuzkor bo'lishadi.[88] Ko'pgina sefalopod erkaklarida spermatozoidlarni ayol mantalining bo'shlig'iga o'tkazishga qodir bo'lgan qo'l uchi bo'lgan gektokotil rivojlanadi. Garchi barcha turlar gektokotildan foydalanmasa ham; masalan, kattalar nautilusi spadiksni chiqaradi.[89] Erkaklarni jalb qilish uchun brakiyal fotoforlarning rivojlanishi ayollarning jinsiy etukligining ko'rsatkichidir.[90]
Urug'lantirish
Sefalopodlar emas translyatsiya spawners. Urug'lantirish jarayonida urg'ochilar erkak tomonidan taqdim etilgan spermatozoidlardan foydalanadilar tashqi urug'lantirish. Ichki urug'lantirish faqat oktopodlarda uchraydi.[87] Kopulyatsiyaning boshlanishi, erkak turga qarab, urg'ochi ayolni ushlaganida va qo'lini "erkakdan ayolga bo'yin" holatida yoki og'izdan og'izga qarab o'ralganidan boshlanadi. Keyin erkaklar spermatozoidlarni bo'shatish uchun mantiyasini bir necha marta qisqartirib, urug'lantirish jarayonini boshlashadi.[91] Sefalopodlar ko'pincha bir necha marta juftlashadi, bu esa erkaklar ilgari urg'ochi urg'ochilar bilan uzoqroq juftlanishiga ta'sir qiladi va mantiyaning qisqarish sonini deyarli uch baravar oshiradi.[91] Tuxumlarning urug'lanishini ta'minlash uchun urg'ochi sefalopodlar spermatozoidani yo'naltirish uchun tuxumning jelatinli qatlamlari orqali spermatozoidlarni jalb qiluvchi peptidni chiqaradi. Ayol sefalopodlar debriyajda tuxum qo'yadilar; har bir tuxum suv ustuniga chiqarilganda rivojlanayotgan embrionning xavfsizligini ta'minlash uchun himoya po'stidan iborat. Reproduktiv strategiyalar sefalopod turlari orasida farq qiladi. Gigant Tinch okean ahtapotida katta tuxumlar uyaga yotqizilgan; ularning barchasini yotqizish uchun ko'pincha bir necha kun kerak bo'ladi.[87] Tuxumlar bo'shatilgandan keyin va himoyalangan substratga biriktirilgach, urg'ochilar o'ladi,[87] ularni tayyorlash yarim semiz. Tsefalopodlarning ayrim turlarida tuxum kavramalari shilimshiq yopishtiruvchi moddadan substratlarga bog'langan. Ushbu tuxumlar perivitellin suyuqligi (PVF) bilan shishiradi, bu gipertonik suyuqlik, erta chiqishni oldini oladi.[92] Urug'lantirilgan tuxum klasterlari yotqizilgan chuqurlikka qarab neytral suzuvchan bo'ladi, ammo qum, marjon matritsasi yoki dengiz o'tlari kabi substratlarda ham bo'lishi mumkin.[86] Ushbu turlar o'z avlodlariga ota-onalarning g'amxo'rligini ta'minlamaganligi sababli, embrionlarni yirtqichlardan kamuflyaj qilish uchun urg'ochi tomonidan tuxum kapsulalarini siyoh bilan kiritish mumkin.[86]
Erkaklar va erkaklar o'rtasidagi musobaqa
Aksariyat sefalopodlar tajovuzkor jinsiy aloqa bilan shug'ullanishadi: erkak kapsulasi qobig'idagi oqsil bu xatti-harakatni rag'batlantiradi. Ular, shuningdek, erkak-erkak tajovuzkorligi bilan shug'ullanishadi, bu erda katta erkaklar o'zaro aloqada g'alaba qozonishadi.[85] Urg'ochi yaqinida bo'lsa, erkaklar bir-birini doimiy ravishda zaryad qiladi va qo'llarini silkitadi. Agar biron bir erkak orqaga chekinmasa, qo'llar orqa tomonga cho'zilib, og'zini ochib, so'ngra qo'l uchlarini tishlaydi.[93] Juftlik raqobati paytida erkaklar ham yuvish usulida qatnashadilar. Ushbu uslub ayol bilan juftlashmoqchi bo'lgan ikkinchi erkak tomonidan qo'llaniladi. Yuvish, bu erdagi birinchi juftlik tomonidan joylashtirilgan bukkali bo'shliqdagi spermatoforlarni bo'shliqqa suv kiritish orqali olib tashlaydi.[85] Erkaklar bilan shug'ullanadigan yana bir xatti-harakatlar - bu krossovkalar bilan juftlashish yoki taqlid qilish - kichikroq erkaklar tajovuzkorlikni kamaytirish uchun o'zlarining xatti-harakatlarini ayolga moslashtiradilar. Ushbu texnikadan foydalanib, ular tuxumni urug'lantirishga qodir, kattaroq erkak esa boshqa erkak tomonidan chalg'itiladi.[93] Ushbu jarayonda krossovkalar erkaklar urug 'idishiga tomchi singari spermani tezda kiritadilar.[94]
Turmush o'rtog'ini tanlash
Juftlik tanlovi mushuk baliqlarining turlarida uchraydi, bu erda urg'ochilar ba'zi erkaklarni boshqalarga qaraganda afzal ko'rishadi, lekin afzal qilingan erkaklarning xususiyatlari noma'lum.[85] Gipotezada ta'kidlanishicha, urg'ochi erkaklar ingl.[85] Bir nechta sefalopod turlari DNK barmoq izlari bilan aniqlangan bir nechta erkak spermatoforlarni qabul qiladi va saqlaydi.[91] Urg'ochilar endi tuxumlarini qo'llarida ushlab turganda juftlashishga urinishlarni qabul qilmaydilar. Urg'ochilar spermani ikki joyda saqlashi mumkin (1) yaqinda juftlashgan erkaklar spermatoforlarini joylashtiradigan bukkal bo'shliqda va (2) oldingi erkaklardan olingan sperma to'plamlari saqlanadigan ichki sperma saqlanadigan idishlarda.[85] Spermatoforni saqlash natijasida sperma raqobati paydo bo'ladi; bu ayolning jufti qaysi tuxumni urug'lantiradi deb ta'kidlaydi. Ushbu turdagi raqobatni kamaytirish uchun erkaklar erni himoya qilish va qizarish kabi agonistik xatti-harakatlarni rivojlantiradilar.[85] The Hapalochlaena lunulatayoki ko'k halqali sakkizoyoq erkak va urg'ochi bilan osonlikcha juftlashadi.[95]
Jinsiy dimorfizm
Turli xil dengiz organizmlarida urg'ochilar ba'zi bir-biriga yaqin turlardagi erkaklarnikiga qaraganda kattaroq ekanligi ko'rinib turibdi. Kabi ba'zi nasllarda adyol ahtapot, erkaklar strukturasi jihatidan kichrayib, atamaga o'xshash bo'lib, "mitti" mitti erkaklar odatda past zichlikda uchraydi.[96] Ahtapot adyol erkak jinsiy-evolyutsion mitti misolidir; urg'ochilar erkaklarnikidan 10 000 dan 40 000 martagacha kattalashadi va erkaklar va urg'ochilar o'rtasidagi jins nisbati tuxum chiqqandan keyin darhol ajralib turishi mumkin.[96]

Embriologiya
Sefalopod tuxumlari diametri 1 dan 30 mm gacha bo'lgan turli xil o'lchamlarga ega.[97] Urug'langan tuxumdon dastlab bo'linib, bir qutbda germinal hujayralar diskini hosil qiladi sarig'i qarama-qarshi qutbda qolgan. Germinal disk o'sib chiqadi va oxir-oqibat sarig'ni so'rib oladi va embrionni hosil qiladi. Tentak va qo'llar avval tananing orqa qismida paydo bo'ladi, u erda oyoq boshqa mollyuskalarda bo'ladi va keyinchalik faqat bosh tomon siljiydi.[82][98]
Sefalopodlarning huni ularning bosh qismida rivojlanadi, og'iz esa qarama-qarshi yuzada rivojlanadi.[99]:86 Dastlabki embriologik bosqichlar ajdodlarni eslatadi gastropodlar va mavjud Monoplakofora.[98]
Chig'anoqlar ektodermadan organik asos sifatida rivojlanib, keyinchalik minerallashadi.[59] Yilda Sepiyaichki qobig'iga ega bo'lgan ektoderma bu organik ramka yotqizilishidan oldin teshiklari yopilgan invazinatsiyani hosil qiladi.[59]
Rivojlanish


To'g'ri: Yetuk Chiroteuthis veranyi. This species has some of the longest tentacles in proportion to its size of any known cephalopod.
The length of time before hatching is highly variable; smaller eggs in warmer waters are the fastest to hatch, and newborns can emerge after as little as a few days. Larger eggs in colder waters can develop for over a year before hatching.[97]
The process from spawning to hatching follows a similar trajectory in all species, the main variable being the amount of yolk available to the young and when it is absorbed by the embryo.[97]
Unlike most other mollusks, cephalopods do not have a morphologically distinct lichinka bosqich. Instead, the juveniles are known as paralarvae. They quickly learn how to hunt, using encounters with prey to refine their strategies.[97]
Growth in juveniles is usually allometrik, whilst adult growth is izometrik.[100]
Evolyutsiya
The traditional view of cephalopod evolution holds that they evolved in the Late Cambrian from a monoplakoforan - ajdod kabi[101] with a curved, tapering shell,[102] which was closely related to the gastropodlar (shilliq qurtlar).[103] Dastlabki qobiqli sefalopodning o'xshashligi Plectronoceras to some gastropods was used in support of this view. A rivojlanishi sifon bu dastlabki shakllarning chig'anoqlari, ularni qo'llab-quvvatlash va chig'anoqlarni vertikal holatda ushlab turish uchun, ularni to'ldirish uchun gaz bilan to'ldirishga imkon bergan bo'lar edi; va hayvonlar pol bo'ylab yurayotganda va haqiqiy sefalopodlarni taxmin qiluvchi ajdodlardan ajratdilar. Naytokonus sifonga ega bo'lmagan.[103] Neutral or positive buoyancy (i.e. the ability to float) would have come later, followed by swimming in the Plectronocerida va oxir-oqibat ko'proq olingan sefalopodlarda reaktiv harakatlanish.[104]
However, some morphological evidence is difficult to reconcile with this view, and the redescription of Nektokaris ptereks, which did not have a shell and appeared to possess jet propulsion in the manner of "derived" cephalopods, complicated the question of the order in which cephalopod features developed – provided Nektokaris is a cephalopod at all.[105]
Early cephalopods were likely predators near the top of the food chain.[20] Keyin late Cambrian extinction led to the disappearance of many Anomalokarididlar, predatory niches became available for other animals.[106] During the Ordovician period the primitive cephalopods underwent pulses of diversification[107] ichida xilma-xil va hukmron bo'lish Paleozoy va Mezozoy dengizlar.[108]
In the Early Palaeozoic, their range was far more restricted than today; they were mainly constrained to sublittoral regions of shallow shelves of the low latitudes, and usually occurred in association with trombolitlar.[109] A more pelagic habit was gradually adopted as the Ordovician progressed.[109] Chuqur suvli sefalopodlar, kamdan-kam hollarda, Quyi Ordovikda topilgan, ammo faqat yuqori kenglikdagi suvlarda.[109]The mid-Ordovician saw the first cephalopods with septa strong enough to cope with the pressures associated with deeper water, and could inhabit depths greater than 100–200 m.[107] The direction of shell coiling would prove to be crucial to the future success of the lineages; endogastric coiling would only permit large size to be attained with a straight shell, whereas exogastric coiling – initially rather rare – permitted the spirals familiar from the fossil record to develop, with their corresponding large size and diversity.[110] (Endogastric mean the shell is curved so as the ventral or lower side is longitudinally concave (belly in); exogastric means the shell is curved so as the ventral side is longitudinally convex (belly out) allowing the funnel to be pointed backward beneath the shell.)[110]

The ancestors of coleoids (including most modern cephalopods) and the ancestors of the modern nautilus, had diverged by the Floian Age of the Early Ordovician Period, over 470 million years ago.[109][111] The Bactritida, a Silurian–Triassic group of orthocones, are widely held to be paraphyletic to the coleoids and ammonoids, that is, the latter groups arose from within the Bactritida.[112]:393 An increase in the diversity of the coleoids and ammonoids is observed around the start of the Devonian period and corresponds with a profound increase in fish diversity. Bu ikkita olingan guruhning kelib chiqishini anglatishi mumkin.[112]
Ko'pgina zamonaviy sefalopodlardan farqli o'laroq, qadimgi navlarning aksariyati himoya qobig'iga ega edi. Ushbu chig'anoqlar dastlab konus shaklida bo'lgan, ammo keyinchalik zamonaviy ko'rinishda kavisli nautiloid shakllariga aylangan nautilus species.Competitive pressure from fish is thought to have forced the shelled forms into deeper water, which provided an evolutionary pressure towards shell loss and gave rise to the modern coleoids, a change which led to greater metabolic costs associated with the loss of buoyancy, but which allowed them to recolonize shallow waters.[103]:36 However, some of the straight-shelled nautiloidlar ga aylandi belemnitlar, out of which some evolved into Kalmar va muzqaymoq.[tekshirish kerak ] The loss of the shell may also have resulted from evolutionary pressure to increase maneuverability, resulting in a more fish-like habit.[1]:289
There has been debate on the embryological origin of cephalopod appendages.[113] Until the mid-twentieth century, the "Arms as Head" hypothesis was widely recognized. In this theory, the arms and tentacles of cephalopods look similar to the head appendages of gastropods, suggesting that they might be gomologik tuzilmalar. Cephalopod appendages surround the mouth, so logically they could be derived from embryonic head tissues.[114] However, the "Arms as Foot" hypothesis, proposed by Adolf Naf in 1928, has increasingly been favoured;[113] masalan, fate mapping of limb buds in the kamerali nautilus indicates that limb buds originate from "foot" embryonic tissues.[115]
Genetika
The sequencing of a full Cephalopod genome has remained challenging to researchers due to the length and repetition of their DNA.[116] The characteristics of Cephalopod genomes were initially hypothesized to be the result of entire genomning takrorlanishi. Following the full sequencing of a Kaliforniya ikki dog'li ahtapot, the genome showed similar patterns to other marine invertebrates with significant additions to the genome assumed to be unique to Cephalopods. No evidence of full genome duplication was found.[117]
Within the California two-spot octopus genome there are substantial replications of two gene families. Significantly, the expanded gene families were only previously known to exhibit replicative behaviour within vertebrates.[117] The first gene family was identified as the Protocadherins which are attributed to neuron development. Protocadherins function as cell adhesion molecules, essential for synaptic specificity. The mechanism for Protocadherin gene family replication in vertebrates is attributed to complex splicing, or cutting and pasting, from a locus. Following the sequencing of the California two-spot octopus, researchers found that the Prorocadherin gene family in Cephalopods has expanded in the genome due to tandem gene duplication. The different replication mechanisms for Protocadherin genes indicate an independent evolution of Protocadherin gene expansion in vertebrates and invertebrates.[117] Analysis of individual Cephalopod Protocadherin genes indicate independent evolution between species of Cephalopod. A species of shore squid Doryteuthis pealeii with expanded Protocadherin gene families differ significantly from those of the California two-spot octopus suggesting gene expansion did not occur before spetsifikatsiya within Cephalopods. Despite different mechanisms for gene expansion, the two-spot octopus Protocadherin genes were more similar to vertebrates than squid, suggesting a konvergent evolyutsiya mexanizm. The second gene family known as C2H2 are small proteins that function as zinc transcription factors. C2H2 are understood to moderate DNA, RNA and protein functions within the cell.[116]
The sequenced California two spot octopus genome also showed a significant presence of bir marta ishlatiladigan elementlar as well as transposon expression. Although the role of transposable elements in marine vertebrates is still relatively unknown, significant expression of transposons in nervous system tissues have been observed.[118] In a study conducted on vertebrates, the expression of transposons during development in Drosophila melanogaster activated genomic diversity between neurons.[119] This diversity has been linked to increased memory and learning in mammals. The connection between transposons and increased neuron capability may provide insight into the observed intelligence, memory and function of Cephalopods.[118]
Filogeniya
The approximate consensus of extant cephalopod phylogeny, after Strugnell va boshq. 2007, is shown in the kladogramma.[65] Mineralized taxa are in qalin. The attachment of the clade including Sepiya va Spirula tushunarsiz; either of the points marked with an asterisk may represent the root of this clade.
| Tsefalopoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||






The internal phylogeny of the cephalopods is difficult to constrain; many molecular techniques have been adopted, but the results produced are conflicting.[65][120] Nautilus tends to be considered an outgroup, with Vampiroteutis forming an outgroup to other squid; however in one analysis the nautiloids, octopus and teuthids plot as a polotomiya.[65] Some molecular phylogenies do not recover the mineralized coleoids (Spirula, Sepiyava Metasepia) as a clade; however, others do recover this more parsimonious-seeming clade, with Spirula as a sister group to Sepiya va Metasepia in a clade that had probably diverged before the end of the Triassic.[121][122]
Molecular estimates for clade divergence vary. One 'statistically robust' estimate has Nautilus diverging from Sakkizoyoq da 415 ± 24 million years ago.[123]
Taksonomiya





The classification presented here, for recent cephalopods, follows largely from So'nggi sefalopodaning hozirgi tasnifi (May 2001), for fossil cephalopods takes from Arkell et al. 1957, Teichert and Moore 1964, Teichert 1988, and others. The three subclasses are traditional, corresponding to the three orders of cephalopods recognized by Bather.[124]
Sefalopoda sinfi († indicates yo'q bo'lib ketgan guruhlar)
- Subklass Nautiloidea: Fundamental ectocochliate cephalopods that provided the source for the Ammonoidea and Coleoidea.
- Order † Plectronocerida: the ancestral cephalopods from the Kembriy Davr
- Order † Ellesmerocerida (500 to 470 Ma)
- Order † Endocerida (485 to 430 Ma)
- Order † Aktinotserida (480 to 312 Ma)
- Order † Discosorida (482 to 392 Ma)
- Order † Pseudorthocerida (432 to 272 Ma)
- Order † Tarfitserida (485 to 386 Ma)
- Order † Oncocerida (478.5 to 324 Ma)
- Buyurtma Nautilida (extant; 410.5 Ma to present)
- Order † Orthocerida (482.5 to 211.5 Ma)
- Order † Ascocerida (478 to 412 Ma)
- Order † Bactritida (418.1 to 260.5 Ma)
- Subclass † Ammonoida: Ammonites (479 to 66 Ma)
- Order † Goniatitida (388.5 to 252 Ma)
- Order † Ceratitida (254 to 200 Ma)
- Order † Ammonitida (215 to 66 Ma)
- Subklass Coleoidea (410.0 Ma-Rec)
- Cohort † Belemnoidea: Belemnites and kin
- Jins † Jeletzkya
- Order † Aulacocerida (265 to 183 Ma)
- Order † Phragmoteuthida (189.6 to 183 Ma)
- Order † Hematitida (339.4 to 318.1 Ma)
- Order † Belemnitida (339.4 to 66 Ma)
- Jins † Belemnoteuthis (189.6 to 183 Ma)
- Kohort Neokoleoidea
- Superorder Dekapodiformes (also known as Decabrachia or Decembranchiata)
- Superorder Octopodiformes (also known as Vampyropoda)
- Oila † Trachyteuthididae
- Buyurtma Vampyromorphida: Vampire squid
- Buyurtma Octopoda: octopus
- Superorder † Paleoteutomorfa
- Order † Boletskida
- Cohort † Belemnoidea: Belemnites and kin
Other classifications differ, primarily in how the various dekapod orders are related, and whether they should be orders or families.
Suprafamilial classification of the Treatise
This is the older classification that combines those found in parts K and L of the Umurtqasiz hayvonlar paleontologiyasi haqida risola, which forms the basis for and is retained in large part by classifications that have come later.
Nautiloids in general (Teichert and Moore, 1964) sequence as given.
- Subclass † Endoceratoidea. Not used by Flower, e.g. Flower and Kummel 1950, interjocerids included in the Endocerida.
- Order † Endocerida
- Order † Intejocerida
- Subclass † Actinoceratoidea Not used by Flower, ibid
- Order † Aktinotserida
- Subklass Nautiloidea Nautiloidea in the restricted sense.
- Order † Ellesmerocerida Plectronocerida subsequently split off as separate order.
- Order † Orthocerida Includes orthocerids and pseudorthocerids
- Order † Ascocerida
- Order † Oncocerida
- Order † Discosorida
- Order † Tarfitserida
- Order † Barrandeocerida A polyphyletic group now included in the Tarphycerida
- Buyurtma Nautilida
- Subclass † Bactritoidea
- Order † Bactritida
Paleozoic Ammonoidea (Miller, Furnish and Schindewolf, 1957)
- Suborder † Anarcestina
- Suborder † Clymeniina
- Suborder † Goniatitina
- Suborder † Prolecanitina
Mesozoic Ammonoidea (Arkel et al., 1957)
- Suborder † Ceratitina
- Suborder † Filloceratina
- Suborder † Lytoceratina
- Suborder † Ammonitina
Subsequent revisions include the establishment of three Upper Cambrian orders, the Plectronocerida, Protactinocerida, and Yanhecerida; separation of the pseudorthocerids as the Pseudorthocerida, and elevating orthoceratid as the Subclass Orthoceratoidea.
Shevyrev classification
Shevyrev (2005) suggested a division into eight subclasses, mostly comprising the more diverse and numerous fossil forms,[125][126] although this classification has been criticized as arbitrary.[127]




Sefalopoda sinfi
- Subclass † Ellesmeroceratoidea
- Order † Plectronocerida (501 to 490 Ma)
- Order † Protactinocerida
- Order † Yanhecerida
- Order † Ellesmerocerida (500 to 470 Ma)
- Subclass † Endoceratoidea (485 to 430 Ma)
- Order † Endocerida (485 to 430 Ma)
- Order † Intejocerida (485 to 480 Ma)
- Subclass † Actinoceratoidea
- Order † Aktinotserida (480 to 312 Ma)
- Subklass Nautiloidea (490.0 Ma- Rec)
- Order † Basslerocerida (490 to 480 Ma)
- Order † Tarfitserida (485 to 386 Ma)
- Order † Lituitida (485 to 480 Ma)
- Order † Discosorida (482 to 392 Ma)
- Order † Oncocerida (478.5 to 324 Ma)
- Buyurtma Nautilida (410.5 Ma-Rec)
- Subclass † Orthoceratoidea (482.5 to 211.5 Ma)
- Order † Orthocerida (482.5 to 211.5 Ma)
- Order † Ascocerida (478 to 412 Ma)
- Order † Dissidocerida (479 to 457.5 Ma)
- Order † Bajkalocerida
- Subclass † Bactritoidea (422 to 252 Ma)
- Subclass † Ammonoida (410 to 66 Ma)
- Subklass Coleoidea (410.0 Ma-rec)[128]
Cladistic classification

Another recent system divides all cephalopods into two qoplamalar. One includes nautilus and most fossil nautiloids. The other clade (Neosefalopoda or Angusteradulata) is closer to modern coleoids, and includes belemnoids, ammonoids, and many orteritsid oilalar. Shuningdek, bor ildiz guruhi cephalopods of the traditional Ellesmerocerida that belong to neither clade.[129][130]
The coleoids, despite some doubts,[1]:289 appear from molecular data to be monophyletic.[131]
Madaniyatda

Ancient seafaring people were aware of cephalopods, as evidenced by artworks such as a stone carving found in the archaeological recovery from Bronze Age Minoan Kriti da Knossos (1900 – 1100 BC) has a depiction of a fisherman carrying an octopus.[132] The terrifyingly powerful Gorgon ning Yunon mifologiyasi may have been inspired by the octopus or squid, the octopus's body representing the severed head of Meduza, the beak as the protruding tongue and fangs, and its tentacles as the snakes.[133]

The Kraken are legendary sea monsters of giant proportions said to dwell off the coasts of Norway and Greenland, usually portrayed in art as giant cephalopods attacking ships. Linney included it in the first edition of his 1735 Systema Naturae.[134][135] Gavayi yaratish afsonasi says that the present cosmos is the last of a series which arose in stages from the ruins of the previous universe. In this account, the octopus is the lone survivor of the previous, alien universe.[136] The Akkorokamui is a gigantic tentacled hayvon dan Aynu folklor.[137]
A battle with an octopus plays a significant role in Viktor Gyugo kitobi Travailleurs de la mer (Dengiz mehnatkashlari ), relating to his time in exile on Gernsi.[138] Yan Fleming 's 1966 short story collection Sakkizoyoq va tirik kunduzgi yorug'lik va 1983 yil Jeyms Bond film were partly inspired by Hugo's book.[139]
Japanese erotic art, shunga o'xshash, o'z ichiga oladi ukiyo-e woodblock prints such as Katsushika Xokusai 's 1814 print Tako to ama (Baliqchining xotinining orzusi ), unda an dalgıç is sexually intertwined with a large and a small octopus.[140][141] The print is a forerunner of tentacle erotika.[142] Biolog P. Z. Myers noted in his science blog, Faringula, that octopuses appear in "extraordinary" graphic illustrations involving women, tentacles, and bare breasts.[143][144]
Since it has numerous arms emanating from a common center, the octopus is often used as a symbol for a powerful and manipulative organization, usually negatively.[145]
Shuningdek qarang
- Sefalopod hajmi
- Tsefalopod ko'z
- Cephalopod intelligence
- Sefalopodlarda og'riq
- Kraken
- Nautiloidlar ro'yxati
- List of ammonites
Adabiyotlar
- ^ a b v d e f g h men Uilbur, Karl M.; Trueman, ER; Klark, M.R., nashr. (1985), Molluska, 11. Formasi va funktsiyasi, Nyu-York: Academic Press, ISBN 0-12-728702-7
- ^ Queiroz, K.; Cantino, P.D.; Gauthier, J.A. (2020). Filonimlar: Filokodning sherigi. CRC Press. p. 1843 yil. ISBN 978-1-138-33293-5.
- ^ "CephBase-ga xush kelibsiz". CephBase. Olingan 29 yanvar 2016.
- ^ a b v d Uilbur, Karl M.; Klark, M.R .; Trueman, ER, eds. (1985), Molluska, 12. Sefalopodlarning paleontologiyasi va neontologiyasi, Nyu-York: Academic Press, ISBN 0-12-728702-7
- ^ Bartol, I. K.; Mann, R .; Vecchione, M. (2002). "Distribution of the euryhaline squid Lolliguncula brevis in Chesapeake Bay: effects of selected abiotic factors". Dengiz ekologiyasi taraqqiyoti seriyasi. 226: 235–247. Bibcode:2002MEPS..226..235B. doi:10.3354/meps226235.
- ^ "Chuchuk suvli sefalopodlar bormi?". ABC Science. 2013 yil 16-yanvar.
- ^ a b v d e f g h men j k l Nikson, Marion; Young, J. Z. (2003). The Brains and Lives of Cephalopods. Nyu-York: Oksford universiteti matbuoti. ISBN 978-0-19-852761-9.
- ^ Tricarico, E.; Amodio, P.; Ponte, G.; Fiorito, G. (2014). "Cognition and recognition in the cephalopod mollusc Octopus vulgaris: coordinating interaction with environment and conspecifics". In Witzany, G. (ed.). Biocommunication of Animals. Springer. 337-349 betlar. ISBN 978-94-007-7413-1.
- ^ Budelmann, B. U. (1995). "The cephalopod nervous system: What evolution has made of the molluscan design". In Breidbach, O.; Kutsch, W. (eds.). The nervous systems of invertebrates: An evolutionary and comparative approach. ISBN 978-3-7643-5076-5.
- ^ Chung, Wen-Sung; Kurniawan, Nyoman D.; Marshall, N. Justin (2020). "Toward an MRI-Based Mesoscale Connectome of the Squid Brain". iScience. 23 (1): 100816. Bibcode:2020iSci...23j0816C. doi:10.1016/j.isci.2019.100816. PMC 6974791. PMID 31972515.
- ^ Raven, Peter et al. (2003). Biologiya, p. 669. McGraw-Hill Education, New York. ISBN 9780073383071.
- ^ Tasaki, I.; Takenaka, T. (1963). "Resting and action potential of squid giant axons intracellularly perfused with sodium-rich solutions". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 50 (4): 619–626. Bibcode:1963PNAS...50..619T. doi:10.1073/pnas.50.4.619. PMC 221236. PMID 14077488.
- ^ a b Packard, A. (1972). "Cephalopods and fish: the limits of convergence". Biologik sharhlar. 47 (2): 241–307. doi:10.1111/j.1469-185X.1972.tb00975.x. S2CID 85088231.
- ^ Macia, Silvia; Robinson, Michael P.; Craze, Paul; Dalton, Robert; Thomas, James D. (2004). "New observations on airborne jet propulsion (flight) in squid, with a review of previous reports". Molluskan tadqiqotlari jurnali. 70 (3): 297–299. doi:10.1093/mollus/70.3.297.
- ^ a b Muramatsu, K.; Yamamoto, J.; Abe, T .; Sekiguchi, K .; Hoshi, N.; Sakurai, Y. (2013). "Oceanic squid do fly". Dengiz biologiyasi. 160 (5): 1171–1175. doi:10.1007/s00227-013-2169-9. S2CID 84388744.
- ^ "Scientists Unravel Mystery of Flying Squid". Ocean Views. National Geographic. 2013 yil 20-fevral.
- ^ Jabr, Ferris (2 August 2010). "Fakt yoki uydirma: kalmar suvdan ucha oladimi?". Ilmiy Amerika.
- ^ a b Serb, J. M.; Eernisse, D. J. (2008). "Charting Evolution's Trajectory: Using Molluscan Eye Diversity to Understand Parallel and Convergent Evolution". Evolyutsiya: Ta'lim va targ'ibot. 1 (4): 439–447. doi:10.1007/s12052-008-0084-1. S2CID 2881223.
- ^ Wells, Martin J. (2011). "Part M, Chapter 4: Physiology of Coleoids". Treatise Online. Lawrence, Kansas, USA. doi:10.17161/to.v0i0.4226. Arxivlandi asl nusxasi 2016-08-22. Olingan 2013-05-10.(obuna kerak)
- ^ a b v d e f g h men j k Boyl, Piter; Rodxaus, Pol (2004). Sefalopodlar: ekologiya va baliqchilik. Blekvell. doi:10.1002 / 9780470995310.ch2. ISBN 978-0-632-06048-1.
- ^ a b Messenger, John B.; Hanlon, Roger T. (1998). Cephalopod Behaviour. Kembrij: Kembrij universiteti matbuoti. 17-21 betlar. ISBN 978-0-521-64583-6.
- ^ Chung, Wen-Sung; Marshall, N. Justin (2016-09-14). "Comparative visual ecology of cephalopods from different habitats". Qirollik jamiyati materiallari B: Biologiya fanlari. 283 (1838): 20161346. doi:10.1098/rspb.2016.1346. ISSN 0962-8452. PMC 5031660. PMID 27629028.
- ^ Hanlon and Messenger, 68.
- ^ Mäthger, L.; Roberts, S .; Hanlon, R. (2010). "Evidence for distributed light sensing in the skin of cuttlefish, Sepia officinalis". Biologiya xatlari. 6 (5): 600–603. doi:10.1098/rsbl.2010.0223. PMC 2936158. PMID 20392722.
- ^ Michinomae, M.; Masuda, H.; Seidou, M.; Kito, Y. (1994). "Structural basis for wavelength discrimination in the banked retina of the firefly squid Watasenia sintillanlari". Eksperimental biologiya jurnali. 193 (1): 1–12. PMID 9317205.
- ^ Seidou, M.; Sugahara, M.; Uchiyama, H.; Hiraki, K.; Hamanaka, T.; Michinomae, M.; Yoshihara, K.; Kito, Y. (1990). "On the three visual pigments in the retina of the firefly squid, Watasenia sintillanlari". Qiyosiy fiziologiya jurnali A. 166 (6). doi:10.1007/BF00187321. S2CID 25707481.
- ^ Stubbs, A. L.; Stubbs, C. W. (2015). "A novel mechanism for color vision: Pupil shape and chromatic aberration can provide spectral discrimination for 'color blind' organisms". bioRxiv 10.1101/017756.
- ^ a b Kingston, A. C.; Kuzirian, A. M.; Hanlon, R. T.; Cronin, T. W. (2015). "Visual phototransduction components in cephalopod chromatophores suggest dermal photoreception". Eksperimental biologiya jurnali. 218 (10): 1596–1602. doi:10.1242/jeb.117945. PMID 25994635. S2CID 25431963.
- ^ "The cephalopods can hear you". BBC yangiliklari. 2009-06-15. Olingan 2010-04-28.
- ^ Tong, D.; Rozas, S.; Oakley, H.; Mitchell, J .; Koli, J .; Mcfall-Ngai, J. (Jun 2009). "Evidence for light perception in a bioluminescent organ". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 106 (24): 9836–9841. Bibcode:2009PNAS..106.9836T. doi:10.1073/pnas.0904571106. ISSN 0027-8424. PMC 2700988. PMID 19509343.
- ^ "integument (mollusks)."Encyclopædia Britannica. 2009 yil. Entsiklopediya Britannica 2006 Ultimate Reference Suite DVD
- ^ Ramirez, M. D.; Oakley, T. H (2015). "Eye-independent, light-activated chromatophore expansion (LACE) and expression of phototransduction genes in the skin of Ahtapot bimakuloidlar" (PDF). Eksperimental biologiya jurnali. 218 (10): 1513–1520. doi:10.1242/jeb.110908. PMC 4448664. PMID 25994633.
- ^ Josef, Noam; Amodio, Piero; Fiorito, Graziano; Shashar, Nadav (2012-05-23). "Camouflaging in a Complex Environment—Octopuses Use Specific Features of Their Surroundings for Background Matching". PLOS ONE. 7 (5): e37579. Bibcode:2012PLoSO...737579J. doi:10.1371/journal.pone.0037579. ISSN 1932-6203. PMC 3359305. PMID 22649542.
- ^ Manda, Shtepan; Turek, Vojtěch (2009). "Minute Silurian oncocerid nautiloids with unusual color patterns". Acta Palaeontologica Polonica. 54 (3): 503–512. doi:10.4202/app.2008.0062. S2CID 54043278.
- ^ Turek, Vojtěch (2009). "Colour patterns in Early Devonian cephalopods from the Barrandian Area: Taphonomy and taxonomy". Acta Palaeontologica Polonica. 54 (3): 491–502. doi:10.4202/app.2007.0064. S2CID 55851070.
- ^ Hanlon, Rojer T.; Messenger, John B. (1999). Cephalopod Behaviour. Kembrij universiteti matbuoti. p. 2018-04-02 121 2. ISBN 978-0-521-64583-6.
- ^ "inkfish". Merriam-Vebster. Olingan 1 fevral 2018.
- ^ Bickerdyke, John (1895). Dengiz baliq ovi. London: Longmans, Green va Co. p. 114.
the common squid or calamary (Loligo vulgaris). It is sometimes called the pen-and-ink fish, on account of its ink bag, and the delicate elongated shell which is found within it.
- ^ Wells, M.J. (1 April 1980). "Nervous control of the heartbeat in octopus". Eksperimental biologiya jurnali. 85 (1): 111–28. PMID 7373208.
- ^ Ghiretti-Magaldi, A. (October 1992). "The Pre-history of Hemocyanin. The Discovery of Copper in the Blood of Molluscs". Uyali va molekulyar hayot haqidagi fanlar. 48 (10): 971–972. doi:10.1007/BF01919143. S2CID 33290596.
- ^ a b v Gilbert, Daniel L.; Adelman, William J.; Arnold, John M. (1990). Squid as Experimental Animals (tasvirlangan tahrir). Springer. ISBN 978-0-306-43513-3.
- ^ a b v Schipp, Rudolf; Mollenhauer, Stephan; Boletzky, Sigurd (1979). "Electron Microscopical and Histochemical Studies of Differentiation and Function of the Cephalopod Gill (Sepia Officinalis L.) ". Zoomorfologiya. 93 (3): 193–207. doi:10.1007/BF00993999. S2CID 20214206.
- ^ Suyak, Q .; Brown, E. R.; Travers, G. (1994). "On the respiratory flow in the cuttlefish Sepia Officinalis" (PDF). Eksperimental biologiya jurnali. 194 (1): 153–165. PMID 9317534.
- ^ Cole, A.; Hall, B. (2009). "Cartilage differentiation in cephalopod molluscs". Zoologiya. 112 (1): 2–15. doi:10.1016/j.zool.2008.01.003. PMID 18722759.
- ^ Shuningdek qarang http://tolweb.org/articles/?article_id=4200
- ^ a b v d e Uilbur, Karl M.; Klark, M.R .; Trueman, ER, eds. (1985), "11: Evolution of Buoyancy and Locomotion in recent cephalopods", Molluska, 12. Sefalopodlarning paleontologiyasi va neontologiyasi, Nyu-York: Academic Press, ISBN 0-12-728702-7
- ^ a b Anderson, E .; Demont, M. (2000). "The mechanics of locomotion in the squid Loligo pealei: Locomotory function and unsteady hydrodynamics of the jet and intramantle pressure". Eksperimental biologiya jurnali. 203 (18): 2851–2863. PMID 10952883.
- ^ a b v Bartol, I. K.; Krueger, P. S.; Thompson, J. T.; Stewart, W. J. (2008). "Swimming dynamics and propulsive efficiency of squids throughout ontogeny". Integrativ va qiyosiy biologiya. 48 (6): 720–733. doi:10.1093/icb/icn043. PMID 21669828.
- ^ Shea, E.; Vecchione, M. (2002). "Quantification of ontogenetic discontinuities in three species of oegopsid squids using model II piecewise linear regression". Dengiz biologiyasi. 140 (5): 971–979. doi:10.1007/s00227-001-0772-7. S2CID 84822175.
- ^ Jonson, V.; Soden, P. D.; Trueman, E. R. (February 1972). "A study in jet propulsion: an analysis of the motion of the squid, Loligo vulgaris". Eksperimental biologiya jurnali. 56 (1972): 155–165.
- ^ Campbell, Reece & Mitchell (1999), p. 612.
- ^ Guerra, A.; Martinell, X.; González, A. F.; Vecchione, M.; Gracia, J.; Martinell, J. (2007). "A new noise detected in the ocean". Buyuk Britaniyaning dengiz biologik assotsiatsiyasi jurnali. 87 (5): 1255–1256. doi:10.1017/S0025315407058225. hdl:10261/27009.
- ^ a b Wells, Martin J.; O'Dor, R. K. (July 1991). "Reaktiv harakatlanish va sefalopodlarning evolyutsiyasi". Dengizchilik fanlari byulleteni. 49 (1): 419–432(14).
- ^ Chamberlain, J., Jr. (1993). "Locomotion in ancient seas: Constraint and opportunity in cephalopod adaptive design". Geobios. 26 (Suppl. 1): 49–61. doi:10.1016/S0016-6995(06)80360-8.
- ^ a b v d e O'Dor, R. K. (1988). "The forces acting on swimming squid". Eksperimental biologiya jurnali. 137: 421–442.
- ^ O'Dor, R. K.; Hoar, J. A. (2000). "Does geometry limit squid growth?". ICES Marine Science jurnali. 57: 8–14. doi:10.1006/jmsc.1999.0502.
- ^ Hanlon, Rojer T.; Watson, Anya C.; Barbosa, Alexandra (2010-02-01). "Atlantika okeanidagi" mimik ahtapot ": tekis baliqlar mimikasi va kamuflyaj Macrotritopus defilippi". Biologik byulleten. 218 (1): 15–24. doi:10.1086/BBLv218n1p15. hdl:1912/4811. ISSN 0006-3185. PMID 20203250.
- ^ "The argonaut shell: Gas-mediated buoyancy control in a pelagic octopus".
- ^ a b v d Baratte, S.; Andouche, A.; Bonnaud, L. (2007). "Engrailed in cephalopods: a key gene related to the emergence of morphological novelties". Development Genes and Evolution. 217 (5): 353–362. doi:10.1007/s00427-007-0147-2. PMID 17394016. S2CID 22241391.
- ^ von Boletzky, S. (2004). "'Ammonoïdes nus': un défi pour la phylogénie des céphalopodes ?" ['Nude ammonoids': a challenge to cephalopod phylogeny?]. Geobios. 37: 117–118. doi:10.1016/j.geobios.2003.01.009.
- ^ Gibson, R. N.; Atkinson, R. J. A .; Gordon, J. D. M., eds. (2006). Okeanografiya va dengiz biologiyasi: yillik sharh. CRC Press. p. 288. ISBN 978-1420006391.
- ^ Aldred, R. G.; Nixon, M.; Young, J. Z. (1983). "Cirrothauma murrayi Chun, a finned octopod". Qirollik jamiyatining falsafiy operatsiyalari B: Biologiya fanlari. 301 (1103): 1–54. Bibcode:1983RSPTB.301....1A. doi:10.1098/rstb.1983.0021.
- ^ Fuks, D .; Ifrim, C .; Stinnesbek, V. (2008). "Yangi Paleoctopus (Cephalopoda: Coleoidea) from the Late Cretaceous of Vallecillo, north-eastern Mexico, and implications for the evolution of Octopoda". Paleontologiya. 51 (5): 1129–1139. doi:10.1111 / j.1475-4983.2008.00797.x.
- ^ von Boletzky, Sigurd (July 1991). "The terminal spine of sepiolid hatchlings: its development and functional morphology (Mollusca, Cephalopoda)". Dengizchilik fanlari byulleteni. 49: 107–112.
- ^ a b v d Strugnell, J.; Nishiguchi, M. K. (2007). "Molecular phylogeny of coleoid cephalopods (Mollusca: Cephalopoda) inferred from three mitochondrial and six nuclear loci: a comparison of alignment, implied alignment and analysis methods". Molluskan tadqiqotlari jurnali. 73 (4): 399–410. doi:10.1093/mollus/eym038.
- ^ Warnke, K .; Keupp, H. (2005). "Spirula – a window to the embryonic development of ammonoids? Morphological and molecular indications for a palaeontological hypothesis". Yuzlar. 51 (1–4): 60–65. doi:10.1007 / s10347-005-0054-9. S2CID 85026080.
- ^ Furuhashi, T.; Schwarzinger, C.; Miksik, I.; Smrz, M.; Beran, A. (2009). "Molluscan shell evolution with review of shell calcification hypothesis". Qiyosiy biokimyo va fiziologiya B. 154 (3): 351–371. doi:10.1016/j.cbpb.2009.07.011. PMID 19665573.
- ^ Dauphin, Y. (1996). "The organic matrix of coleoid cephalopod shells: molecular weights and isoelectric properties of the soluble matrix in relation to biomineralization processes". Dengiz biologiyasi. 125 (3): 525–529. doi:10.1007/BF00353265 (harakatsiz 2020-10-09).CS1 maint: DOI 2020 yil oktyabr holatiga ko'ra faol emas (havola)
- ^ Dauphin, Y. (1983). Les subdivisions majeures de la classe des céphalopodes : bases de la systématique actuelle : apport de l'analyse microstructurale. These Doct. Etat, Université Paris Sud. OCLC 972899981.
- ^ Dauphin, Y. (2001). "Nanostructures de la nacre des tests de céphalopodes actuels". Paläontologische Zeitschrift. 75 (1): 113–122. doi:10.1007/bf03022601. ISSN 0031-0220. S2CID 126900936.
- ^ Toll, R. B.; Binger, L. C. (1991). "Arm anomalies: Cases of supernumerary development and bilateral agenesis of arm pairs in Octopoda (Mollusca, Cephalopoda)". Zoomorfologiya. 110 (6): 313–316. doi:10.1007 / BF01668021. S2CID 34858474.
- ^ Anatomy of the Common Squid. 1912.
- ^ Nixon 1988 in Wippich, M. G. E.; Lehmann, J. (2004). "Allocrioceras from the Cenomanian (mid-Cretaceous) of the Lebanon and its bearing on the palaeobiological interpretation of heteromorphic ammonites". Paleontologiya. 47 (5): 1093–1107. doi:10.1111/j.0031-0239.2004.00408.x.
- ^ Uilbur, Karl M.; Klark, M.R .; Trueman, ER, eds. (1985), "5", Molluska, 12. Sefalopodlarning paleontologiyasi va neontologiyasi, Nyu-York: Academic Press, ISBN 0-12-728702-7
- ^ Maykl Xogan. 2011 yil. Kelt dengizi. eds. P. Kir yuvish va C. Klivlend. Yer entsiklopediyasi. Fan va atrof-muhit bo'yicha milliy kengash. Vashington shahar.
- ^ "Cephalopod radula". Tree of Life web project.
- ^ a b v d e Nixon, M. (1995). "A nomenclature for the radula of the Cephalopoda (Mollusca) – living and fossil". Zoologiya jurnali. 236: 73–81. doi:10.1111/j.1469-7998.1995.tb01785.x.
- ^ a b Gabbott, S. E. (1999). "Janubiy Afrikaning marhum Ordovikist Soom Shale Lagerstatte-dan ortokonik sefalopodlar va ular bilan bog'liq bo'lgan hayvonot dunyosi". Paleontologiya. 42: 123–148. doi:10.1111/1475-4983.00065.
- ^ Landman, Nil X.; Davis, Richard Arnold; Mapes, Royal H., eds. (2007). Cephalopods present and past: new insights and fresh perspectives. Springer. ISBN 978-1-4020-6461-6.
- ^ Richardson & ... (1977). Fossils of the Mason Creek.
- ^ Kruta, I.; Landman, N.; Rouget, I.; Cecca, F.; Tafforeau, P. (2011). "The role of ammonites in the Mesozoic marine food web revealed by jaw preservation". Ilm-fan. 331 (6013): 70–72. Bibcode:2011Sci...331...70K. doi:10.1126/science.1198793. PMID 21212354. S2CID 206530342.
- ^ a b Barns, Robert D. (1982). Umurtqasizlar zoologiyasi. Filadelfiya, Pensilvaniya: Xolt-Sonders Xalqaro. pp. 450–460. ISBN 978-0-03-056747-6.
- ^ Loest, R. A. (1979). "Ammiakning volatilizatsiyasi va quruqlikdagi gastropodlar tomonidan so'rilishi - qobiq va qobiqsiz turlarni taqqoslash". Fiziologik zoologiya. 52 (4): 461–469. doi:10.1086/physzool.52.4.30155937. JSTOR 30155937.
- ^ a b v Boucher-Rodoni, R.; Mangold, K. (1994). "Ammonia production in cephalopods, physiological and evolutionary aspects". Dengiz va chuchuk suvlarning o'zini tutishi va fiziologiyasi. 25 (1–3): 53–60. doi:10.1080/10236249409378907.
- ^ a b v d e f g h men j k Vidal, Erica A. G. Advances in Cephalopod Science: Biology, Ecology, Cultivation and Fisheries.
- ^ a b v d e Rodriges, M.; Gerra; Troncoso (2010). "The embryonic phase and its implication in the hatchling size and condition of Atlantic bobtail squid Sepiola Atlantica". Helgoland dengiz tadqiqotlari. 65 (2): 211–216. Bibcode:2011HMR....65..211R. doi:10.1007/s10152-010-0217-0. S2CID 41577834.
- ^ a b v d Arkhipkin, A. I. (1992). "Reproductive system structure, development and function in cephalopods with a new general scale for maturity stages". Journal of Northwest Atlantic Fishery Science. 12: 63–74. doi:10.2960/j.v12.a7.
- ^ Mohanty, Sobhi; Ojanguren, Alfredo F.; Fuiman, Lee A. (2014-07-01). "Aggressive male mating behavior depends on female maturity in Ahtapot bimakuloidlar". Dengiz biologiyasi. 161 (7): 1521–1530. doi:10.1007/s00227-014-2437-3. ISSN 0025-3162. S2CID 85256742.
- ^ Saunders, W. B; Spinosa, C. (1978). "Sexual dimorphism in Nautilus from Palau". Paleobiologiya. 4 (3): 349–358. doi:10.1017/S0094837300006047.
- ^ Young, R. B. (1975). "A Systematic Approach to Planning Occupational Programs". Community College Review. 3 (2): 19–25. doi:10.1177/009155217500300204. S2CID 145374345.
- ^ a b v Squires, Z. E; Norman, M. D; Stuart-Fox, D. (2013). "Mating behaviour and general spawning patterns of the southern dumpling squid Euprymna tasmanica". Molluskan tadqiqotlari jurnali. 79 (3): 263–269. doi:10.1093/mollus/eyt025.
- ^ Marthy, H. J.; Xauzer, R; Scholl, A. (1976). "Natural tranquilizer in cephalopod eggs". Tabiat. 261 (5560): 496–7. Bibcode:1976Natur.261..496M. doi:10.1038/261496a0. PMID 945466. S2CID 8693207.
- ^ a b Norman, M. D.; Lu, C. C. (1997). "Redescription of the southern dumpling squid Euprymna tasmanica va naslni qayta ko'rib chiqish Evropima (Cephalopoda: Sepiolidae)". Buyuk Britaniyaning dengiz biologik assotsiatsiyasi jurnali. 77 (4): 1109–1137. doi:10.1017/s0025315400038662.
- ^ Ivata, Y .; Ito, K .; Sakurai, Y. (2008). "Effect of low temperature on mating behavior of squid Loligo bleekeri". Baliqchilik fani. 74 (6): 1345–1347. doi:10.1111/j.1444-2906.2008.01664.x. S2CID 43094931.
- ^ Cheng, Meri V.; Kolduell, Roy L. (2000 yil iyul). "Moviy halqali ahtapotda jinsni aniqlash va juftlash, Hapalochlaena lunulata". Hayvonlar harakati. 60 (1): 27–33. doi:10.1006 / anbe.2000.1447. ISSN 0003-3472. PMID 10924200. S2CID 32899443.
- ^ a b Fairbairn, D. (2013). "Blanket Octopus: Drifting urg'ochilari va mitti erkaklar". Toq juftliklar: Hayvonot dunyosidagi jinslar o'rtasidagi g'ayrioddiy farqlar. Prinston universiteti matbuoti. 104-115 betlar.
- ^ a b v d Von Boletski, S. (2003). Sefalopod mollyuskalarida erta hayot bosqichlari biologiyasi. Dengiz biologiyasining yutuqlari. 44. 143-203 betlar. doi:10.1016 / S0065-2881 (03) 44003-0. ISBN 978-0-12-026144-4. PMID 12846042.
- ^ a b Shigeno, S .; Sasaki, T .; Moritaki, T .; Kasugay, T .; Vecchione, M.; Agata, K. (Yanvar 2008). "Ko'p boshli mollyuskaning tana qismlarini yig'ish yo'li bilan bosh suyagi bosh kompleksi evolyutsiyasi Nautilus embrional rivojlanish ". Morfologiya jurnali. 269 (1): 1–17. doi:10.1002 / jmor.10564. PMID 17654542. S2CID 13109195.
- ^ Gilbert, Daniel L.; Adelman, Uilyam J.; Arnold, Jon M. (1990). Kalamar eksperimental hayvonlar sifatida. Nyu-York: Plenum matbuoti. ISBN 978-0-306-43513-3.
- ^ Moltschaniwskyj, Natalie A. (2004). "Sefalopodlarning o'sish jarayonini tushunish". Dengiz va chuchuk suv tadqiqotlari. 55 (4): 379–386. doi:10.1071 / MF03147.
- ^ Lemche, H.; Wingstrand, K. G. (1959). "Anatomiyasi Neopilina galatheae Lemche, 1957 (Molluska, Tryblidiacea) " (Bepul to'liq matn + lavhalarga havola). Galateya hisoboti. 3: 9–73.
- ^ Wingstrand, K. G. (1985). "So'nggi monoplakoforaning anatomiyasi va aloqalari to'g'risida". Galateya hisoboti. 16: 7-94. Arxivlandi asl nusxasi (Bepul to'liq matn + lavhalarga havola) 2016-03-03 da. Olingan 2009-03-23.
- ^ a b v Boyl, P .; Rodhouse, P. (2005). "Kelib chiqishi va evolyutsiyasi". Sefalopodlar. p. 36. doi:10.1002 / 9780470995310.ch3. ISBN 9780470995310.
- ^ Kröger, B. R. (2007). "Qadimgi sefalopod buyrug'ining ba'zi kam ma'lum bo'lgan xususiyatlari Ellesmerocerida (Nautiloidea, Cephalopoda)". Paleontologiya. 50 (3): 565–572. doi:10.1111 / j.1475-4983.2007.00644.x.
- ^ Smit, Martin R.; Karon, Jan-Bernard (2010). "Kembriyalik ibtidoiy yumshoq tanali sefalopodlar". Tabiat. 465 (7297): 427–428. Bibcode:2010 yil natur.465..427B. doi:10.1038 / 465427a. PMID 20505713. S2CID 205055896.
- ^ Jain, Sreepat (2016). Umurtqasiz hayvonlar paleontologiyasi asoslari: Makrofosil. Springer. p. 73. ISBN 978-81-322-3658-0.
- ^ a b Kröger, B .; Yun-bai, Y. B. (2009). "Ordovik davrida impulsli sefalopod diversifikatsiyasi". Paleogeografiya, paleoklimatologiya, paleoekologiya. 273 (1–2): 174–201. Bibcode:2009PPP ... 273..174K. doi:10.1016 / j.palaeo.2008.12.015.
- ^ Dzik, J. (1981). "Sefalopodaning kelib chiqishi" (PDF). Acta Palaeontologica Polonica. 26 (2): 161–191.
- ^ a b v d Kröger, B. R .; Servayz, T .; Chjan, Y .; Kosnik, M. (2009). "Ordovikada pelagik sefalopodlarning kelib chiqishi va dastlabki ko'tarilishi". PLOS ONE. 4 (9): e7262. Bibcode:2009PLoSO ... 4.7262K. doi:10.1371 / journal.pone.0007262. PMC 2749442. PMID 19789709.
- ^ a b Holland, C. H. (1987). "Nautiloid sefalopodlar: g'alati muvaffaqiyat: Prezidentning yubiley 1986 yilgi murojaatnomasi". Geologiya jamiyati jurnali. 144 (1): 1–15. Bibcode:1987JGSoc.144 .... 1H. doi:10.1144 / gsjgs.144.1.0001. S2CID 128629737.
- ^ Kryger, Byörn (2006). "Baltoskandiyaning Darrivillian (O'rta Ordovik) ning ortoseridan sefalopodlarining erta o'sish bosqichlari va tasnifi". Leteya. 39 (2): 129–139. doi:10.1080/00241160600623749.
- ^ a b Yosh, R. E .; Vecchione, M.; Donovan, D. T. (1998). "Koleoid sefalopodlarning rivojlanishi va ularning hozirgi biologik xilma-xilligi va ekologiyasi". Janubiy Afrika dengizshunoslik jurnali. 20 (1): 393–420. doi:10.2989/025776198784126287.
- ^ a b Tanabe, K. (2008). Sefalopodlar - hozirgi va o'tmish. Tokio: Tokai universiteti matbuoti.[sahifa kerak ]
- ^ Basil, Jennifer; Bahktinova, Irina; Kuroyva, Kristin; Li, Nandi; Mims, Desirei; Preis, Maykl; Soucier, Christian (2005-09-01). "Nautilus pompilius L. (Cephalopoda, Nautiloidea) ning hidoforiya va tentaklarning hidga yo'naltirilganligi". Dengiz va chuchuk suvlarning o'zini tutishi va fiziologiyasi. 38 (3): 209–221. doi:10.1080/10236240500310096. S2CID 33835096.
- ^ Shigeno, Shuichi; Sasaki, Takenori; Moritaki, Takeya; Kasugay, Takashi; Vecchione, Maykl; Agata, Kiyokazu (2008 yil yanvar). "Ko'p boshli mollyuskaning tana qismlarini yig'ish yo'li bilan bosh suyagi boshi kompleksining evolyutsiyasi: Nautilus embrional rivojlanishidan dalillar". Morfologiya jurnali. 269 (1): 1–17. doi:10.1002 / jmor.10564. PMID 17654542. S2CID 13109195.
- ^ a b O'Brayen, Keytlin E.; Roumbedakis, Katina; Vinkelmann, Inger E. (2018-06-06). "Sefalopod ilmining hozirgi holati va uchta dastlabki martaba tadqiqotchilarining oldidagi eng muhim vazifalar istiqbollari". Fiziologiyadagi chegara. 9: 700. doi:10.3389 / fphys.2018.00700. ISSN 1664-042X. PMC 6014164. PMID 29962956.
- ^ a b v Albertin, Kerolin B.; Simakov, Oleg; Mitros, Tereza; Vang, Z. Yan; Pungor, Judit R.; Edsinger-Gonsales, Erik; Brenner, Sidney; Ragdseyl, Klifton V.; Roxsar, Daniel S. (avgust 2015). "Ahtapot genomi va bosh miya oyoqlari nerv-morfologik yangiliklari evolyutsiyasi". Tabiat. 524 (7564): 220–224. Bibcode:2015 Noyabr 524 ... 220A. doi:10.1038 / tabiat14668. ISSN 0028-0836. PMC 4795812. PMID 26268193.
- ^ a b Gehring, Meri A. (2013-02-04), O'simliklar transpozonlari va evolyutsiyadagi genom dinamikasi, Uili-Blekvell, 117–142 betlar, doi:10.1002 / 9781118500156.ch7, ISBN 978-1-118-50015-6 Yo'qolgan yoki bo'sh
sarlavha =(Yordam bering);| bob =mensimagan (Yordam bering) - ^ Ervin, Jennifer A.; Marchetto, Mariya S.; Gage, Fred H. (2014 yil avgust). "Miyada xilma-xillik va murakkablikni yaratishda mobil DNK elementlari". Neuroscience-ning tabiat sharhlari. 15 (8): 497–506. doi:10.1038 / nrn3730. ISSN 1471-003X. PMC 4443810. PMID 25005482.
- ^ Strugnell, J .; Norman, M .; Jekson, J .; Drummond, A .; Kuper, A. (2005). "Multigenli yondashuv yordamida koleoid sefalopodlarning molekulyar filogeniyasi (Mollusca: Cephalopoda); ma'lumotlar taqsimotining Bayesiya doirasidagi filogeniyalarni hal qilishga ta'siri". Molekulyar filogenetik va evolyutsiyasi. 37 (2): 426–441. doi:10.1016 / j.ympev.2005.03.020. PMID 15935706.
- ^ Strugnell, J .; Jekson, J .; Drummond, A. J .; Kuper, A. (2006). "Asosiy sefalopod guruhlari uchun divergentsiya vaqtini taxmin qilish: ko'plab genlarning dalillari". Kladistika. 22: 89–96. doi:10.1111 / j.1096-0031.2006.00086.x. S2CID 84743000.
- ^ Karlini, D. B.; Reece, K. S .; Graves, J. E. (2000). "Aktin genlari oilasi evolyutsiyasi va koleoid sefalopodlarning filogeniyasi (Mollusca: Cephalopoda)". Molekulyar biologiya va evolyutsiya. 17 (9): 1353–1370. doi:10.1093 / oxfordjournals.molbev.a026419. PMID 10958852.
- ^ Bergmann, S .; Lib, B .; Rut, P.; Markl, J. (2006). "Tirik qoldiqlardan olingan gemosiyanin, sefalopod Nautilus pompilius: oqsil tuzilishi, genlar tashkiloti va evolyutsiyasi ". Molekulyar evolyutsiya jurnali. 62 (3): 362–374. Bibcode:2006JMolE..62..362B. doi:10.1007 / s00239-005-0160-x. PMID 16501879. S2CID 4389953.
- ^ Bather, F.A. (1888b). "Professor Bleyk va Sefalopodadagi Shell-Growth". Tabiiy tarix yilnomalari va jurnali. 6. 1 (6): 421–426. doi:10.1080/00222938809460761.
- ^ Shevyrev, A. A. (2005). "Sefalopod makrosistemasi: tarixiy sharh, bilimning hozirgi holati va hal qilinmagan muammolar: 1. Sefalopod mollyuskalarining asosiy xususiyatlari va umumiy tasnifi". Paleontologik jurnal. 39 (6): 606–614. Dan tarjima qilingan Paleontologicheskii Jurnal № 6, 2005, 33-42.
- ^ Shevyrev, A. A. (2006). "Sefalopod makrosistemasi; tarixiy sharh, bilimlarning hozirgi holati va hal qilinmagan muammolar; 2, nautiloid sefalopodlarning tasnifi". Paleontologik jurnal. 40 (1): 46–54. doi:10.1134 / S0031030106010059. S2CID 84616115.
- ^ Kroger, B. "Rossiyaning" Paleontologik jurnalida ekspertlar fikri'". Arxivlandi asl nusxasi 2009-08-31.
- ^ Bather, F.A. (1888a). "Sefalopodadagi qobiq o'sishi (Siphonopoda)". Tabiiy tarix yilnomalari va jurnali. 6. 1 (4): 298–310. doi:10.1080/00222938809460727.
- ^ Bertold, Tomas; Engeser, Teo (1987). "Cefalopoda (Mollusca) ning filogenetik tahlili va tizimlashtirilishi". Gamburgdagi Verhandlungen Naturwissenschaftlichen Vereins. 29: 187–220.
- ^ Engeser, Teo (1997). "Nautiloidea fotoalbomlari sahifasi". Arxivlandi asl nusxasi 2006-09-25.
- ^ Lindgren, A. R .; Giribet, G .; Nishiguchi, M. K. (2004). "Sefalopoda (Molluska) filogeniyasiga birlashgan usul". Kladistika. 20 (5): 454–486. CiteSeerX 10.1.1.693.2026. doi:10.1111 / j.1096-0031.2004.00032.x. S2CID 85975284.
- ^ Xogan, S Maykl (2007 yil 22-dekabr). "Knossos yozuvlari". Zamonaviy antikvar.
- ^ Uilk, Stiven R. (2000). Meduza: Gorgon sirini echish. Oksford universiteti matbuoti. ISBN 978-0-19-988773-6.
- ^ "Caroli Linnaei Systema naturae sistens regna tria naturae". google.com.
- ^ Smedli, Edvard; Rouz, Xyu Jeyms; Rose, Genri Jon (1845). Entsiklopediya Metropolitana, yoki universal bilimlar lug'ati: falsafiy va alifbo tartibida ikki tomonlama afzalliklarni tegishli gravyuralar bilan shakllantirish. B. Felles. 255– betlar.
- ^ Dikson, Roland Burrage (1916). Okean. Barcha irqlarning mifologiyasi. 9. Marshall Jons kompaniyasi. 2–2 betlar.
- ^ Batchelor, John (1901). Aynu va ularning folklorlari. London: Diniy yo'llar jamiyati.
- ^ Chisholm, Xyu, nashr. (1911). . Britannica entsiklopediyasi (11-nashr). Kembrij universiteti matbuoti.
- ^ Koen-Vrignaud, Jerar (2012). "Sakkizoyoqlarda yoki ayol kuchi anatomiyasida". Farqi. 23 (2): 32–61. doi:10.1215/10407391-1533520.
- ^ Fritze, Sointu; Suojoki, Saara (2000). Taqiqlangan rasmlar: Yaponiyaning Edo davridagi erotik san'at (fin tilida). Helsingin kaupungin taidemuseo. 23-28 betlar. ISBN 978-951-8965-54-4.
- ^ Uhlenbek, Kris; Margarita Vinkel; Ellis Tinios; Amy Reigle Newland (2005). Yapon shahvoniy fantaziyalari: Edo davridagi jinsiy tasvir. Hotei. p. 161. ISBN 978-90-74822-66-4.
- ^ Briel, Xolger (2010). Berninger, Mark; Ekke, Xoxen; Xaberkorn, Gideon (tahrir). Roving Eye sayohat qiluvchi rasmlarni uchratadi: Vizyon sohasi va kattalar uchun manga global o'sishi. Komikslar madaniyatlarning aloqasi sifatida: ommaviy axborot vositalari, intizomlarning o'zaro aloqalari haqida insholar. McFarland. p. 203. ISBN 978-0-7864-3987-4.
- ^ Myers, P. Z. (2017 yil 17-may). "Favqulodda ahtapot rasmlari". Faringula. Olingan 18 mart 2017.
- ^ Myers, P. Z. (2006 yil 29 oktyabr). "Albatta, ish uchun xavfsiz emas". Faringula. Olingan 18 mart 2017.
- ^ Smit, S. (26 fevral, 2010 yil). "Nima uchun Mark Tsukerberg Ahtapot multfilmi" fashistlarning tashviqotini "uyg'otmoqda, nemis qog'ozi kechirim so'raydi". iMediaEthics. Olingan 31 may 2017.
Qo'shimcha o'qish
- Barskov, I. S .; Boiko, M. S .; Konovalova, V. A .; Leonova, T. B.; Nikolaeva, S. V. (2008). "Paleozoyning dengiz ekotizimidagi sefalopodlar". Paleontologik jurnal. 42 (11): 1167–1284. doi:10.1134 / S0031030108110014. S2CID 83608661. Paleozoy sefalopodlarining keng qamrovli ko'rinishi.
- Kempbell, Nil A.; Reece, Jeyn B.; Mitchell, Lourens G. (1999). Biologiya, beshinchi nashr. Menlo Park, Kaliforniya: Addison Wesley Longman, Inc. ISBN 978-0-8053-6566-5.
- Felli, J., Vekchione, M., Roper, C. F. E., Suini, M. va Kristensen, T., 2001-2003: So'nggi sefalopodaning hozirgi tasnifi. Milliy tabiiy tarix muzeyi: Tizimli biologiya bo'limi: Umurtqasizlar zoologiyasi: Sefalopodlar
- N. Joan Abbott, Roddi Uilyamson, Linda Meddok. Sefalopod nevrologiyasi. Oksford universiteti matbuoti, 1995 yil. ISBN 0-19-854790-0
- Marion Nikson va Jon Z. Young. Sefalopodlarning miyasi va hayoti. Oksford universiteti matbuoti, 2003 yil. ISBN 0-19-852761-6
- Hanlon, Rojer T. va Jon B. Messenger. Sefalopod harakati. Kembrij universiteti matbuoti, 1996 y. ISBN 0-521-42083-0
- Martin Stivens va Sami Merilayta. Hayvonlarning kamuflyaji: mexanizmlari va funktsiyasi. Kembrij universiteti matbuoti, 2011 yil. ISBN 0-521-19911-5
- Rodxaus, P. G.; Nigmatullin, Ch. M. (1996). "Iste'molchining roli". Qirollik jamiyatining falsafiy operatsiyalari B: Biologiya fanlari. 351 (1343): 1003–1022. doi:10.1098 / rstb.1996.0090.
- Zamonaviy sefalopodlarning tasnifi kaliti: ftp://ftp.fao.org/docrep/fao/009/a0150e/a0150e03.pdf[doimiy o'lik havola ]
Tashqi havolalar
- Baliq va sefalopodlarga qarshi
- TONMO.COM - "Octopus News Magazine Online" - sefalopod maqolalari va muhokamasi
- Ilmiy amerikalik: kalmar suvdan uchib chiqa oladimi?
- Rojer Xanlonning seminari: "Sefalopodlarda tez moslashuvchan kamuflyaj va signalizatsiya"
- Chuqur dengizda yashovchi tukli qurtlar
| Vikimedia Commons-ga tegishli ommaviy axborot vositalari mavjud Tsefalopoda. |