Ikki marta urug'lantirish - Double fertilization


Ikki marta urug'lantirish kompleks urug'lantirish gullarni o'simliklarning mexanizmi (angiospermlar ). Ushbu jarayon ayolning qo'shilishini o'z ichiga oladi gametofit (megagametofit, shuningdek embrion sumkasi ) ikki erkak bilan jinsiy hujayralar (sperma). Bu qachon boshlanadi polen donasi ning isnodiga rioya qiladi karpel, gulning ayol jinsiy tizimi. Keyin polen donasi namlikni oladi va boshlanadi nihol, shakllantirish a polen naychasi tomon pastga cho'zilgan tuxumdon uslub orqali. Keyin polen naychasining uchi tuxumdonga kirib, orqali kirib boradi mikropil ovulda ochilish. Polen naychasi megagametofitdagi ikkita spermani bo'shatishga davom etadi.
Urug'lanmagan tuxumdon hujayralari soni 8 tadan bo'lib, 3 + 2 + 3 (yuqoridan pastgacha) shaklida joylashtirilgan, ya'ni 3 antipodal hujayralar, 2 qutbli markaziy hujayralar, 2 ta sinergidlar va 1 tuxum hujayralari. Bir sperma tuxum hujayrasini urug'lantiradi, ikkinchisi esa ikkisi bilan birlashadi qutb yadrolari katta markaziy hujayra megagametofit. Gaploid sperma va gaploid tuxum birlashib, a hosil qiladi diploid zigota, jarayon chaqirilmoqda sinxamiya, ikkinchisi esa sperma va ikkalasi gaploid megagametofitning yirik markaziy hujayrasining qutb yadrolari a hosil qiladi triploid yadro (uch marta termoyadroviy). Ba'zi o'simliklar shakllanishi mumkin poliploid yadrolar. Gametofitning katta hujayrasi keyinchalik rivojlanib boradi endosperm, rivojlanayotgan embrionni oziqlantiruvchi ozuqaviy moddalarga boy to'qima. Tuxumdonni o'rab turgan tuxumdon mevaga aylanadi, u urug'larni himoya qiladi va ularni tarqatish uchun ishlashi mumkin.[1]
Endospermga hissa qo'shadigan ikkita markaziy hujayraning onalik yadrolari (qutb yadrolari) bir xil mitozdan kelib chiqadi. meiotik tuxumni keltirib chiqaradigan mahsulot. Triploid endospermning genetik konstitutsiyasiga onalik hissasi embrionning hissasi bilan ikki baravar ko'pdir.
Zavodning 2008 yilda o'tkazilgan tadqiqotida Arabidopsis talianasi, ayol yadrolari bilan birlashganda, ayol jinsiy hujayralardagi erkak yadrolarining migratsiyasi birinchi marta yordamida hujjatlashtirildi. jonli ravishda tasvirlash. Migratsiya va termoyadroviy jarayonda ishtirok etgan ba'zi genlar ham aniqlandi.[2]
Ikki marta urug'lanishning dalili Gnetales, gul bo'lmagan urug 'o'simliklari bo'lganligi haqida xabar berilgan.[3]
Qisqa tarix
Ikki marta urug'lantirish bir asrdan ko'proq oldin kashf etilgan Sergey Navaschin va Grignard Kiev,[4] Rossiya imperiyasi, va Leon Gignard Frantsiya. Ularning har biri kashfiyotni boshqasidan mustaqil ravishda amalga oshirdi.[5] Lilium martagon va Fritillariya tenella klassik yordamida amalga oshirilgan er-xotin urug'lantirishning birinchi kuzatuvlarida ishlatilgan yorug'lik mikroskopi. Yorug'lik mikroskopining cheklanganligi sababli, er-xotin urug'lantirish jarayoni bilan bog'liq ko'plab javobsiz savollar mavjud edi. Biroq, rivojlanishi bilan elektron mikroskop, ko'plab savollarga javoblar berildi. Eng muhimi, V. Jensen guruhi tomonidan olib borilgan kuzatuvlar shuni ko'rsatdiki, erkak jinsiy hujayralarida yo'q hujayra devorlari va bu plazma membranasi jinsiy hujayralar polen donasi ichida ularni o'rab turgan hujayraning plazma membranasiga yaqin.[6]
Gimnospermlarda er-xotin urug'lantirish
Ikki baravar o'g'itlashning ancha ibtidoiy shakli, odatda Gnetales deb nomlanuvchi gimnospermlar tartibining jinsiy ko'payishida yuzaga keladi.[3] Xususan, ushbu tadbir ikkalasida ham hujjatlashtirilgan Efedra va Gnetum, ning pastki qismi Gnetofitlar.[7] Yilda Ephedra nevadensis, bitta binukleat sperma hujayrasi tuxum hujayrasiga yotqizilgan. Dastlabki urug'lanish hodisasidan so'ng, ikkinchi siterma yadrosi tuxum sitoplazmasida topilgan qo'shimcha tuxum yadrosini urug'lantirish uchun yo'naltiriladi. Ko'pgina boshqa urug'li o'simliklarda ushbu ikkinchi "ventral kanal yadrosi" odatda funktsional jihatdan foydasiz deb topilgan.[8] Yilda Gnetum gnemon, ayolning sitoplazmasida ko'plab bo'sh tuxum yadrolari mavjud gametofit. Etuk ayol gametofitining penetratsiyasini polen naychasi, ayol sitoplazmasi va erkin yadrolari polen naychasini o'rab olish uchun harakat qiladi. Ikki urug 'yadrosi binukleat sperma hujayrasidan ajralib chiqadi, so'ngra erkin tuxum yadrolari bilan qo'shilib, hayotga yaroqli zigotalar hosil qiladi, bu oilalar orasidagi homologik xususiyatdir. Efedra va Gnetum.[9] Ikkala oilada ham urug'lantirishning ikkinchi hodisasi qo'shimcha diploid hosil qiladi embrion. Ushbu ortiqcha sonli embrion keyinchalik uzilib, faqat bitta etuk embrionning sinteziga olib keladi.[10] Qo'shimcha urug'lantirish mahsuloti Efedra birlamchi embrionni oziqlantirmaydi, chunki ayol gametofit ozuqa bilan ta'minlash uchun javobgardir.[9] Ikki marta urug'lantirishning ancha ibtidoiy jarayoni gimnospermlar natijada bir xil tuxum hujayrasida joylashgan ikkita diploid yadro paydo bo'ladi. Bu farq qiladi angiosperm holati, buning natijasida tuxum hujayrasi ajralib chiqadi va endosperm.[11] Genomiga nisbatan qiyosiy molekulyar tadqiqotlar G. gnemon gnetofitlar bilan chambarchas bog'liqligini aniqladi ignabargli daraxtlar angiospermlarga qaraganda.[12][13][14] Gnetales va angiospermlarni qardosh taksonlar deb belgilaydigan antofit gipotezasining rad etilishi, er-xotin urug'lanish jarayoni hosil bo'lgan degan taxminlarga olib keladi. konvergent evolyutsiyasi va gnetofitlar va angiospermlar orasida mustaqil ravishda paydo bo'lgan.[15]
In vitro o'g'itlash
In vitro er-xotin urug'lantirish ko'pincha gulli o'simliklardagi gametalar sintezining boshqa jihatlari bilan bir qatorda molekulyar o'zaro ta'sirini o'rganish uchun ishlatiladi. Erkak va urg'ochi jinsiy hujayralar o'rtasida in vitro qo'shaloq urug'lantirishni rivojlanishidagi asosiy to'siqlardan biri bu sperma polen naychasida va tuxumning embrion xaltachasida saqlanishi. Tuxum va spermatozoidlarning boshqariladigan sinteziga allaqachon erishilgan haşhaş o'simliklar.[16] Polen unib chiqishi, polen naychasiga kirish va er-xotin urug'lantirish jarayonlari normal davom etishi kuzatilgan. Darhaqiqat, ushbu usul allaqachon turli xil gulli o'simliklarda urug'larni olish uchun ishlatilgan va "sinov naychasining urug'lantirilishi" deb nomlangan.[17]
Bilan bog'liq tuzilmalar va funktsiyalar
Megagametofit
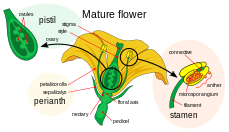
Ikki marta urug'lanishda ishtirok etadigan ayol gametofit, megagametofit angiospermlar gaploid bo'lgan embrion xaltachasi deyiladi. Bu an ichida rivojlanadi tuxumdon, a asosidagi tuxumdon bilan o'ralgan karpel. Megagametofit atrofida (bitta yoki) ikkitasi bor tamsayılar, deb nomlangan ochilishni hosil qiladi mikropil. Odatda megagametofit gaploid, (odatda) dan kelib chiqadi diploid ) megaspora ona hujayrasi, deb ham ataladi megasporotsit. Hodisalarning navbatdagi ketma-ketligi, ma'lum bir turga qarab o'zgarib turadi, lekin aksariyat turlarda quyidagi hodisalar sodir bo'ladi. Megasporotsit to'rtta gaploid megasporalar hosil qilib, mayoz hujayralar bo'linishidan o'tadi. Natijada paydo bo'lgan to'rtta megasporadan faqat bittasi omon qoladi. Ushbu megaspora uch tur mitoz bo'linishidan o'tadi, natijada sakkizta gaploid yadroli etti hujayradan iborat (markaziy hujayrada qutb yadrolari deb ataladigan ikkita yadro mavjud). Embrion xaltachasining pastki uchi boshqa ikkita gaploid hujayralar o'rtasida joylashgan gaploid tuxum hujayrasidan iborat bo'lib, ular sinergidlar. Sinergidlar polen naychasini mikropil orqali megagametofitga tortish va yo'naltirishda ishlaydi. Megagametofitning yuqori uchida antipodal hujayralar joylashgan.
Mikrogametofit
Ikki marta urug'lanishga qatnashadigan erkak gametofitlar yoki mikrogametofitlar tarkibiga kiradi polen donalar. Ular mikrosporangiya ichida rivojlanadi yoki polen xaltachalari stamensdagi anteriyalardan. Har bir mikrosporangiyada diploid mavjud mikrospora ona hujayralari yoki mikrosporotsitlar. Har bir mikrosporotsit mayozga uchraydi va to'rtta gaploid mikrosporani hosil qiladi, ularning har biri oxir-oqibat polen donasiga aylanishi mumkin. Mikrosporaga uchraydi mitoz va sitokinez ikkita alohida hujayrani hosil qilish uchun, generativ hujayra va kolba hujayrasi. Bu ikki hujayra, spora devoridan tashqari, etuk bo'lmagan polen donasini tashkil qiladi. Erkak gametofitning pishishi bilan generativ hujayra naycha hujayrasiga o'tadi va generativ hujayra mitozga uchraydi va ikkita sperma hujayrasini hosil qiladi. Polen donasi pishib bo'lgach, anterlar changni bo'shatib, oching. Toz changlari pistil shamol yoki hayvonlar changlatuvchi vositalar yordamida va dog 'ustiga qo'yilgan boshqa gul. Polen donasi unib chiqqach, naycha xujayrasi polp naychasini ishlab chiqaradi, u cho'zilib, karpelning uzun uslubi bo'ylab va tuxumdonga cho'zilib, uning sperma hujayralari megagametofitda ajralib chiqadi. Ikki marta urug'lantirish bu erdan keladi.[18]
Shuningdek qarang
Adabiyotlar
- ^ Berger, F. (2008 yil yanvar). "Ikki marta urug'lantirish, afsonalardan haqiqatga". Jinsiy o'simliklarning ko'payishi. 21 (1): 3–5. doi:10.1007 / s00497-007-0066-4. S2CID 8928640.
- ^ Berger, F.; Hamamura, Y. va Ingouff, M. va Xigashiyama, T. (Avgust 2008). "Ikki marta urug'lantirish - amalda qo'lga kiritildi". O'simlikshunoslik tendentsiyalari. 13 (8): 437–443. doi:10.1016 / j.tplants.2008.05.011. PMID 18650119.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ a b V. Raghavan (2003 yil sentyabr). "Ikki marta urug'lanishga oid ba'zi mulohazalar, uning kashf etilishidan to hozirgi kungacha". Yangi fitolog. 159 (3): 565–583. doi:10.1046 / j.1469-8137.2003.00846.x.
- ^ Kordium EL (2008). "[Gulli o'simliklarda er-xotin urug'lantirish: 1898-2008]". Tsitol. Genet. (rus tilida). 42 (3): 12–26. PMID 18822860.
- ^ Jensen, W. A. (1998 yil fevral). "Ikki marta urug'lantirish: shaxsiy ko'rinish". Jinsiy o'simliklarning ko'payishi. 11 (1): 1–5. doi:10.1007 / s004970050113. S2CID 33416360.
- ^ Dumas, C. & Rogowsky, P. (avgust 2008). "Urug'lantirish va erta urug'larning shakllanishi". Comptes Rendus Biologies. 331 (10): 715–725. doi:10.1016 / j.crvi.2008.07.013. PMID 18926485.
- ^ Karmikel, J. S .; Fridman, V. E. (1995-12-01). "Gnetum gnemonida er-xotin urug'lantirish: hujayra tsikli va jinsiy ko'payish o'rtasidagi bog'liqlik". O'simlik hujayrasi. 7 (12): 1975–1988. doi:10.1105 / tpc.7.12.1975. ISSN 1040-4651. PMC 161055. PMID 12242365.
- ^ Fridman, Uilyam E. (1990). "Ephedra nevadensis (Ephedraceae) da jinsiy ko'payish: gulsiz urug 'o'simlikida er-xotin urug'lanishning keyingi dalillari". Amerika botanika jurnali. 77 (12): 1582–1598. doi:10.1002 / j.1537-2197.1990.tb11399.x. JSTOR 2444491.
- ^ a b Karmayl, Jeffri S.; Fridman, Uilyam E. (1996). "Gnetum gnemonida (Gnetaceae) qo'shaloq urug'lantirish: uning Gnetales va antofitlar pardasida jinsiy ko'payish evolyutsiyasi". Amerika botanika jurnali. 83 (6): 767–780. doi:10.1002 / j.1537-2197.1996.tb12766.x. JSTOR 2445854.
- ^ Fridman, V. E. (1995-04-25). "Organik takrorlanish, inklyuziv fitness nazariyasi va altruizm: endosperm va angiosperm reproduktiv sindrom evolyutsiyasini tushunish". Milliy fanlar akademiyasi materiallari. 92 (9): 3913–3917. Bibcode:1995 yil PNAS ... 92.3913F. doi:10.1073 / pnas.92.9.3913. ISSN 0027-8424. PMC 42072. PMID 11607532.
- ^ Fridman, Uilyam E. (1994). "Urug'li o'simliklarda embriogeniya evolyutsiyasi va endospermaning rivojlanish tarixi va dastlabki tarixi". Amerika botanika jurnali. 81 (11): 1468–1486. doi:10.1002 / j.1537-2197.1994.tb15633.x. JSTOR 2445320.
- ^ Bou, L. Mishel; Palto, Gvineya; dePamphilis, Klod V. (2000-04-11). "Uchala genomik bo'linmalarga asoslangan urug 'o'simliklarining filogeniyasi: ekstremal gimnospermlar monofil, Gnetalesning eng yaqin qarindoshlari esa ignabargli daraxtlardir". Milliy fanlar akademiyasi materiallari. 97 (8): 4092–4097. Bibcode:2000PNAS ... 97.4092B. doi:10.1073 / pnas.97.8.4092. ISSN 0027-8424. PMC 18159. PMID 10760278.
- ^ Qish, Kay-Uve; Beker, Annet; Myunster, Tomas; Kim, Yan T.; Saydler, Xaynts; Tissen, Gyunter (1999-06-22). "MADS-box genlari gnetofitlar gulli o'simliklarga qaraganda ignabargli daraxtlar bilan ko'proq bog'liqligini aniqlaydi". Milliy fanlar akademiyasi materiallari. 96 (13): 7342–7347. Bibcode:1999 PNAS ... 96.7342W. doi:10.1073 / pnas.96.13.7342. ISSN 0027-8424. PMC 22087. PMID 10377416.
- ^ Magallon, S .; Sanderson, J. J. (2002-12-01). "Yuqori darajada saqlanib qolgan genlardan kelib chiqadigan urug 'o'simliklari o'rtasidagi munosabatlar: qadimiy nasablar orasida qarama-qarshi filogenetik signallarni saralash". Amerika botanika jurnali. 89 (12): 1991–2006. doi:10.3732 / ajb.89.12.1991. ISSN 1537-2197. PMID 21665628.
- ^ Chau, Shu-Miyav; Parkinson, Kristofer L.; Cheng, Yuchang; Vinsent, Tomas M.; Palmer, Jeffri D. (2000-04-11). "Uchala o'simlik genomidan kelib chiqadigan urug'lik o'simlik filogeniyasi: mavjud gimnospermlarning monofili va Gnetalesning ignabargli daraxtlardan kelib chiqishi". Milliy fanlar akademiyasi materiallari. 97 (8): 4086–4091. Bibcode:2000PNAS ... 97.4086C. doi:10.1073 / pnas.97.8.4086. ISSN 0027-8424. PMC 18157. PMID 10760277.
- ^ Zenkteler, M. (1990). "In vitro urug'lantirish va yuqori o'simliklarda keng duragaylash". Crit Rev Plant Sci. 9 (3): 267–279. doi:10.1080/07352689009382290.
- ^ Raghavan, V. (2005). Ikki marta urug'lantirish: gulli o'simliklarda embrion va endosperm rivojlanishi (tasvirlangan tahrir). Birxauzer. 17-19 betlar. ISBN 978-3-540-27791-0.
- ^ Kempbell NA; Reece J.B (2005). Biologiya (7 nashr). San-Fransisko, Kaliforniya: Pearson Education, Inc., 774–777-betlar. ISBN 978-0-8053-7171-0.
| Subdiziplar | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| O'simlik guruhlar | |||||||||||
| |||||||||||
| |||||||||||
| O'simliklarning o'sishi va odati | |||||||||||
| Ko'paytirish | |||||||||||
| O'simliklar taksonomiyasi | |||||||||||
| Amaliyot | |||||||||||
| |||||||||||
| |||||||||||