O'simliklar morfologiyasi - Plant morphology
Fitomorfologiya ning o'rganilishi jismoniy shakli va tashqi tuzilishi ning o'simliklar.[1] Bu odatda alohida deb hisoblanadi o'simlik anatomiyasi,[1] bu ichki narsani o'rganishdir tuzilishi o'simliklar, ayniqsa mikroskopik darajada.[2] O'simliklar morfologiyasi o'simliklarni vizual identifikatsiyalashda foydalidir. Yaqinda o'tkazilgan tadqiqotlar molekulyar biologiya o'simlik morfologiyalarining saqlanishi va xilma-xilligini aniqlash bilan bog'liq bo'lgan molekulyar jarayonlarni o'rganishni boshladi. Ushbu tadqiqotlarda transkriptom tabiatni muhofaza qilishning hal qiluvchi ahamiyatga ega ekanligi aniqlandi ontogenetik davomida o'tish o'simlik turli xillikni cheklaydigan evolyutsion cheklashlarga olib kelishi mumkin bo'lgan hayot tsikli[3].

Qo'llash sohasi


O'simliklar morfologiyasi "o'simliklarning rivojlanishi, shakli va tuzilishini o'rganishni anglatadi va demak, ularni reja va kelib chiqish o'xshashligi asosida izohlashga urinishni anglatadi".[4] O'simliklar morfologiyasining to'rtta asosiy tadqiqot yo'nalishlari mavjud va ularning har biri boshqa sohalar bilan bir-biriga to'g'ri keladi biologiya fanlari.
Avvalo, morfologiya qiyosiy, ya'ni morfolog bir xil yoki har xil turdagi ko'plab turli o'simliklarning tuzilmalarini tekshiradi, so'ngra taqqoslashlar o'tkazadi va o'xshashlik haqida g'oyalarni shakllantiradi. Turli xil turlardagi tuzilmalar umumiy va merosxo'rlik natijasida mavjud bo'lib rivojlanadi deb hisoblaganda genetik yo'llar, bu tuzilmalar deb nomlanadi gomologik. Masalan, barglar ning qarag'ay, eman va karam barchasi bir-biridan juda farq qiladi, lekin ba'zi bir asosiy tuzilmalar va qismlarning joylashuvi bilan o'rtoqlashadi. Barglarning gomologiyasi - bu oson xulosa qilishdir. O'simlik morfologi oldinga boradi va ekanligini aniqlaydi tikanlar ning kaktus shuningdek, boshqa o'simliklarda barglar bilan bir xil asosiy tuzilish va rivojlanishni baham ko'radi va shuning uchun kaktus tikanlar ham barglar uchun gomologik hisoblanadi. O'simliklar morfologiyasining bu jihati o'simliklarni o'rganish bilan bir-biriga to'g'ri keladi evolyutsiya va paleobotanika.
Ikkinchidan, o'simlik morfologiyasi ikkalasini ham kuzatadi vegetativ (badandagi) o'simliklarning tuzilmalari, shuningdek reproduktiv tuzilmalar. Ning vegetativ tuzilmalari qon tomir o'simliklar tarkibiga kiritilgan tortishish tizimini o'rganishni o'z ichiga oladi borib taqaladi va barglar, shuningdek ildiz tizim. Reproduktiv tuzilmalar xilma-xil bo'lib, odatda ma'lum bir o'simlik guruhiga xosdir, masalan gullar va urug'lar, fern sori va mox kapsulalar. O'simliklardagi reproduktiv tuzilmalarni batafsil o'rganish natijasida kashfiyot olib borildi avlodlar almashinuvi barcha o'simliklarda va ko'pchiligida mavjud suv o'tlari. O'simliklar morfologiyasining ushbu sohasi biologik xilma-xillik va o'simliklar sistematikasi.
Uchinchidan, o'simliklar morfologiyasi o'simliklarning tuzilishini bir qator miqyosda o'rganadi. Eng kichik tarozida ultrastruktura, hujayralar faqat an yordamida ko'rinadigan umumiy tuzilish xususiyatlari elektron mikroskop va sitologiya, yordamida hujayralarni o'rganish optik mikroskopiya. Ushbu miqyosda o'simliklar morfologiyasi bir-biriga to'g'ri keladi o'simlik anatomiyasi o'rganish sohasi sifatida. Eng katta miqyosda o'simliklarni o'rganish o'sish odati, o'simlikning umumiy arxitekturasi. A-dagi dallanish shakli daraxt har xil turga qarab o'zgaradi, shuningdek o'simlikning tashqi ko'rinishi a daraxt, o't, yoki o't.
To'rtinchidan, o'simliklar morfologiyasi naqshini o'rganadi rivojlanish, o'simliklarning o'sishi bilan tuzilmalarning kelib chiqishi va pishishi jarayoni. Hayvonlar umrining boshidanoq tanadagi barcha qismlarni ishlab chiqarar ekan, o'simliklar butun umri davomida doimo yangi to'qima va tuzilmalarni ishlab chiqaradi. Tirik o'simlik har doim embrion to'qimalariga ega. Yangi tuzilmalar ishlab chiqarilishida ularning pishib etish uslubiga, ular rivojlana boshlagan davrda o'simlik hayotining nuqtasi, shuningdek, tuzilmalar ta'sir qiladigan muhit ta'sir qilishi mumkin. Morfolog bu jarayonni, sabablarini va natijasini o'rganadi. O'simliklar morfologiyasining bu sohasi bir-biriga to'g'ri keladi o'simliklar fiziologiyasi va ekologiya.
Qiyosiy fan
O'simlik morfologi bir xil yoki har xil turdagi ko'plab turli xil o'simliklar tarkibidagi tuzilmalarni taqqoslashni amalga oshiradi. Turli o'simliklardagi o'xshash tuzilmalar o'rtasida bunday taqqoslashlar savolga javob beradi nima uchun tuzilmalari o'xshash. Ehtimol, genetika, fiziologiya yoki atrof-muhitga ta'sir ko'rsatishning o'xshash sabablari tashqi ko'rinishdagi o'xshashlikka olib keldi. Ushbu sabablarni ilmiy tekshirish natijalari asosiy biologiya haqidagi ikkita tushunchadan biriga olib kelishi mumkin:
- Gomologiya - umumiy nasab va umumiy genetika tufayli tuzilish ikki tur o'rtasida o'xshashdir.
- Yaqinlashish - umumiy atrof-muhit bosimiga mustaqil moslashish tufayli tuzilish ikki tur o'rtasida o'xshashdir.
Qaysi xususiyatlar va tuzilmalarning har bir turga tegishli ekanligini tushunish o'simlikni tushunishning muhim qismidir evolyutsiya. Evolyutsion biolog tuzilmalarni talqin qilishda o'simlik morfologiga ishonadi va o'z navbatida ta'minlaydi filogeniyalar yangi morfologik tushunchalarga olib kelishi mumkin bo'lgan o'simlik munosabatlarining.
Gomologiya
Turli xil turlardagi tuzilmalar umumiy va meros bo'lib o'tgan genetik yo'llar natijasida mavjud bo'lib rivojlanadi deb hisoblasa, bu tuzilmalar gomologik. Masalan, qarag'ay, eman va karamning barglari bir-biridan juda farq qiladi, ammo ba'zi bir asosiy tuzilmalar va qismlarning joylashishi bilan bo'lishadi. Barglarning gomologiyasi - bu oson xulosa qilishdir. O'simliklar morfologi yanada rivojlanib, kaktusning tikanlari ham boshqa o'simliklarda barglar bilan bir xil asosiy tuzilishga va rivojlanishga ega ekanligini va shuning uchun kaktus tikanlar ham barglar uchun gomologik ekanligini aniqlaydi.
Yaqinlashish
Turli xil turlardagi tuzilmalar atrof-muhit bosimiga umumiy moslashuvchan reaktsiyalar natijasida mavjud va rivojlanadi deb hisoblasa, bu tuzilmalar yaqinlashuvchi. Masalan, Bryopsis plumosa va jarohatlaydi Qushqo'nmas setaceus ikkalasi ham bir xil tukli tarvaqaylab ko'rinishga ega, garchi biri alg bo'lsa, biri gulli o'simlik. Umumiy tuzilishdagi o'xshashlik yaqinlashish natijasida mustaqil ravishda yuzaga keladi. Ko'pchilikning o'sish shakli kaktuslar va turlari Eforhoriya juda o'xshash, garchi ular juda uzoq oilalarga tegishli bo'lsa ham. O'xshashlik issiq va quruq muhitda omon qolish muammosining umumiy echimlaridan kelib chiqadi.
Vegetativ va reproduktiv xususiyatlar
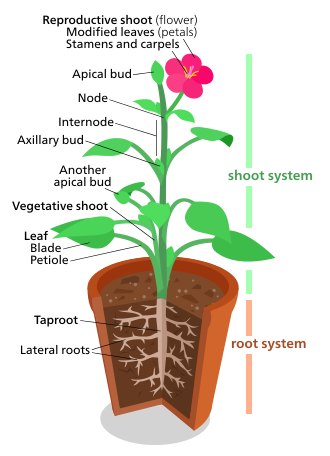
O'simliklar morfologiyasi o'simliklarning vegetativ tuzilishini ham, jinsiy tizimini ham davolaydi.
The vegetativ (badandagi) tuzilmalari qon tomir o'simliklar ikkita asosiy organ tizimini o'z ichiga oladi: (1) a tortishish tizimi, tarkib topgan borib taqaladi va barglar va (2) a ildiz tizimi. Ushbu ikkita tizim deyarli barcha qon tomir o'simliklar uchun umumiydir va o'simliklar morfologiyasini o'rganish uchun birlashtiruvchi mavzuni beradi.
Aksincha, reproduktiv tuzilmalari har xil va odatda o'simliklarning ma'lum bir guruhiga xosdir. Kabi tuzilmalar gullar va mevalar faqat angiospermlar; sori faqat topilgan ferns; va urug 'konuslari faqat topilgan ignabargli daraxtlar va boshqa gimnospermlar. Shuning uchun reproduktiv belgilar vegetativ belgilarga qaraganda o'simliklarni tasniflash uchun ko'proq foydali hisoblanadi.
Identifikatsiya qilishda foydalaning
O'simliklar biologlari o'simliklarning taksonlaridagi farqlarni yoki o'xshashliklarni baholash uchun ularni taqqoslash, o'lchash, hisoblash va tavsiflash mumkin bo'lgan o'simliklarning morfologik belgilaridan foydalanadilar va bu belgilarni o'simliklarni identifikatsiyalash, tasniflash va tavsiflash uchun ishlatadilar.
Belgilar tavsiflashda yoki identifikatsiya qilishda ishlatilganda ular chaqiriladi diagnostik yoki asosiy belgilar bu ham sifatli, ham miqdoriy bo'lishi mumkin.
- Miqdoriy belgilar - bu hisoblash yoki o'lchash mumkin bo'lgan morfologik xususiyatlar, masalan, o'simlik turlari 10-12 mm kenglikdagi gul barglariga ega.
- Sifatli belgilar - bu barg shakli, gul rangi yoki o'sishi kabi morfologik xususiyatlar.
Ikkala turdagi belgilar o'simliklarni aniqlash uchun juda foydali bo'lishi mumkin.
Avlodlarning o'zgarishi
O'simliklardagi reproduktiv tuzilmalarni batafsil o'rganish natijasida kashfiyot olib borildi avlodlar almashinuvi, barcha o'simliklarda va ko'pchiligida mavjud suv o'tlari, nemis botanik tomonidan Wilhelm Hofmeister. Ushbu kashfiyot barcha o'simliklar morfologiyasida yaratilgan eng muhimlardan biridir, chunki u barcha o'simliklarning hayot aylanish jarayonini tushunish uchun umumiy asos yaratadi.
O'simliklardagi pigmentatsiya
O'simliklardagi pigmentlarning asosiy vazifasi fotosintez, bu yashil pigmentdan foydalanadi xlorofill iloji boricha ko'proq yorug'lik energiyasini olishga yordam beradigan bir nechta qizil va sariq pigmentlar bilan birga. Pigmentlar ham jalb qilishning muhim omilidir hasharotlar dalda berish uchun gullarga changlanish.
O'simlik pigmentlari tarkibiga turli xil molekulalar kiradi porfirinlar, karotenoidlar, antosiyaninlar va betaleynlar. Barcha biologik pigmentlar ma'lum to'lqin uzunliklarini tanlab oladi yorug'lik esa aks ettiradi boshqalar. Yutilgan yorug'lik o'simlik tomonidan kimyoviy reaktsiyalarni kuchaytirish uchun ishlatilishi mumkin, yorug'likning aks ettirilgan to'lqin uzunliklari pigment ko'zga ko'rinadigan rangni aniqlaydi.
Rivojlanishdagi morfologiya
O'simlik rivojlanish o'simliklarning o'sishi bilan tuzilishlarning kelib chiqishi va pishib etish jarayoni. Bu mavzu bo'yicha tadqiqotlar o'simlik anatomiyasi va o'simliklar fiziologiyasi shuningdek, o'simliklar morfologiyasi.
O'simliklardagi rivojlanish jarayoni ko'rinib turganidan tubdan farq qiladi umurtqali hayvonlar hayvonlar. Qachonki hayvon embrion rivojlana boshlaydi, u hayotida mavjud bo'lgan barcha tana qismlarini juda erta ishlab chiqaradi. Hayvon tug'ilganda (yoki tuxumidan chiqqanda), uning barcha tana qismlari bor va shu vaqtdan boshlab faqat kattalashib, etuk bo'ladi. Aksincha, o'simliklar butun hayoti davomida doimiy ravishda yangi to'qima va tuzilmalarni ishlab chiqaradi meristemalar[5] organlarning uchida yoki etuk to'qimalar o'rtasida joylashgan. Shunday qilib, tirik o'simlik har doim embrion to'qimalariga ega.
O'simlikda ko'rinadigan tashkilot xususiyatlari paydo bo'ladigan xususiyatlar bu alohida qismlarning yig'indisidan ko'proq. "Ushbu to'qimalar va funktsiyalarni birlashtirilgan ko'p hujayrali organizmga biriktirish nafaqat alohida qismlar va jarayonlarning xususiyatlarini, balki alohida qismlarni tekshirish asosida oldindan taxmin qilinmaydigan yangi xususiyatlar to'plamini ham beradi."[6] Boshqacha qilib aytganda, o'simlikdagi molekulalar haqida hamma narsani bilish hujayralarning xususiyatlarini taxmin qilish uchun etarli emas; va hujayralarning barcha xususiyatlarini bilish o'simlik tuzilishining barcha xususiyatlarini oldindan aytib bo'lmaydi.
O'sish
A qon tomir o'simlik bitta hujayradan boshlanadi zigota tomonidan tashkil etilgan urug'lantirish Sperma hujayrasi tomonidan tuxum hujayrasining. Shu paytdan boshlab, u o'simlik hosil qilish uchun bo'linishni boshlaydi embrion jarayoni orqali embriogenez. Bu sodir bo'lganda, hosil bo'lgan hujayralar bir uchi birinchi ildizga aylanishi uchun, boshqa uchi esa o'qning uchini tashkil qilishi uchun tartibga solinadi. Yilda urug ' o'simliklar, embrion bir yoki bir nechta "urug 'barglari" ni rivojlantiradi (kotletonlar ). Embriogenezning oxirida yosh o'simlik hayotida boshlash uchun zarur bo'lgan barcha qismlarga ega bo'ladi.
Bir marta embrion unib chiqadi uning urug'idan yoki ota-ona o'simlikidan, bu jarayon orqali qo'shimcha organlar (barglar, poyalar va ildizlar) hosil qila boshlaydi organogenez. Yangi ildizlar ildizdan o'sib chiqadi meristemalar ildizning uchida joylashgan bo'lib, yangi novdalar va barglar o'simtadan o'sadi meristemalar otishni o'rganish uchida joylashgan.[7] Dallanish meristemada qoldirilgan va hali o'tmagan hujayralarning kichik to'plamlari bo'lganda paydo bo'ladi uyali farqlash ixtisoslashgan to'qima hosil qilish uchun yangi ildizning uchi yoki o'simtasi sifatida o'sishni boshlaydi. Ildiz yoki kurtak uchidagi har qanday bunday meristemadan o'sish deb ataladi birlamchi o'sish va natijada bu ildiz yoki o'qning uzayishiga olib keladi. Ikkilamchi o'sish ildizlarning kengayishiga yoki a hujayralar bo'linmalaridan otishga olib keladi kambiy.[8]
Tomonidan o'sishdan tashqari hujayra bo'linish, o'simlik o'sishi mumkin hujayralarni cho'zish. Bu alohida hujayralar yoki hujayralar guruhlari uzoqroq o'sganda sodir bo'ladi. Hamma o'simlik hujayralari bir xil uzunlikda o'smaydi. Ildizning bir tomonidagi hujayralar boshqa tarafdagi hujayralarga qaraganda uzunroq va tezroq o'sganda, natijada ildiz sekin o'sayotgan hujayralar tomoniga egilib qoladi. Ushbu yo'naltirilgan o'sish o'simlikning ma'lum bir stimulga, masalan, nurga bo'lgan munosabati orqali sodir bo'lishi mumkin (fototropizm ), tortishish kuchi (gravitropizm ), suv, (gidrotropizm ) va jismoniy aloqa (thigmotropism ).
O'simliklarning o'sishi va rivojlanishi o'ziga xoslik bilan amalga oshiriladi o'simlik gormonlari va o'simliklarning o'sishini tartibga soluvchilar (PGRs) (Ross va boshq. 1983).[9] Endogen gormonlar darajasiga o'simlik yoshi, sovuqqa chidamlilik, uyqusizlik va boshqa metabolizm sharoitlari ta'sir qiladi; fotoperiod, qurg'oqchilik, harorat va boshqa tashqi muhit sharoitlari; va PGRlarning ekzogen manbalari, masalan, tashqi qo'llaniladigan va rizosfera kelib chiqishi.
Morfologik xilma-xillik
O'simliklar shakli va tuzilishida tabiiy o'zgarishni namoyish etadi. Barcha organizmlar har bir kishidan farq qilsa, o'simliklar qo'shimcha o'zgarish turini namoyish etadi. Bitta shaxs ichida shakli va tuzilishi jihatidan boshqa o'xshash qismlardan farq qilishi mumkin bo'lgan qismlar takrorlanadi. Ushbu o'zgarish o'simlik barglarida osonlikcha ko'rinadi, ammo boshqa organlar, masalan, poyalar va gullar shu kabi o'zgarishni ko'rsatishi mumkin. Ushbu o'zgarishning uchta asosiy sababi bor: pozitsion ta'sir, atrof muhitga ta'sir qilish va balog'atga etmaganlik.
O'simliklar morfologiyasining evolyutsiyasi
Transkripsiya omillari va transkripsiyaviy tartibga solish tarmoqlari o'simliklar morfogenezida va ularning evolyutsiyasida asosiy rol o'ynaydi. O'simliklar qo'nish paytida ko'plab yangi transkripsiya omillari oilalari paydo bo'ldi va imtiyozli ravishda ko'p hujayrali rivojlanish, ko'payish va organlarning rivojlanish tarmoqlariga ulanib, er o'simliklarining murakkab morfogeneziga hissa qo'shdi.[10]
Pozitsion effektlar

O'simliklar hayot davomida bir xil organning ko'p nusxalarini ishlab chiqarganiga qaramay, ma'lum bir organning barcha nusxalari bir xil bo'lmaydi. Yetuk o'simlik qismlari orasida organ ishlab chiqariladigan nisbiy holatdan kelib chiqadigan xilma-xillik mavjud. Masalan, yangi novda bo'ylab barglar novdalar bo'ylab izchil ravishda o'zgarishi mumkin. Filial poydevori yonida hosil bo'lgan barglarning shakli o'simlik uchida hosil bo'lgan barglardan farq qiladi va bu farq ma'lum bir o'simlikda va ma'lum bir turda shoxdan shoxga mos keladi. Ushbu farq novdaning ikkala uchidagi barglar pishib bo'lgandan keyin ham saqlanib qoladi va ba'zi barglarning boshqalaridan yosh bo'lishining natijasi emas.
Atrof muhitga ta'siri
Yangi tuzilmalar ishlab chiqarilishida ularning etuklashish uslubiga, ular rivojlana boshlagan davrda o'simliklar hayotidagi nuqta va shuningdek, tuzilmalar ta'sir qiladigan muhit ta'sir qilishi mumkin. Buni ko'rish mumkin suv o'simliklari va paydo bo'lgan o'simliklar.
Harorat
Harorat o'simliklarga turli xil omillarga, shu jumladan o'simlikning kattaligi va holatiga va ta'sir qilish harorati va davomiyligiga bog'liq ravishda ko'p ta'sir ko'rsatadi. Kichikroq va ko'proq o'simlik suvli, juda yuqori yoki juda past haroratlarda shikastlanish yoki o'limga nisbatan sezgirlik. Harorat biokimyoviy va fiziologik jarayonlarning tezligiga ta'sir qiladi, harorat odatda harorat oshib borishi (chegaralar ichida). Shu bilan birga, monomolekulyar reaktsiyalar uchun Van't Hoff munosabati (bu reaktsiyaning tezligi 10 ° S harorat ko'tarilishi bilan ikki yoki uch marta ko'payishini bildiradi) biologik jarayonlar uchun, ayniqsa past va yuqori haroratlarda qat'iy amal qilmaydi.
Suv o'simliklarda muzlaganida, o'simlik uchun oqibatlar muzlashning hujayralararo (hujayralar ichida) yoki hujayralararo (hujayradan tashqari) bo'shliqlarda tashqi hujayralar sodir bo'lishiga juda bog'liq.[11] Hujayra ichidagi muzlash, odatda o'simlik va uning to'qimalarining chidamliligidan qat'i nazar hujayrani o'ldiradi.[12] Hujayra ichidagi muzlash tabiatda kamdan-kam uchraydi, ammo haroratning o'rtacha pasayishi, masalan, 1 ° C dan 6 ° C / soatgacha hujayralararo muz hosil bo'lishiga olib keladi va bu "organik bo'lmagan muz"[13] to'qimalarning qattiqligiga qarab o'limga olib kelishi mumkin yoki bo'lmasligi mumkin.
Sovuq haroratda, o'simliklar to'qimalarining hujayralararo bo'shliqlaridagi suv avval muzlaydi, ammo harorat 7 ° C dan pastga tushguncha suv muzlatilmasligi mumkin.[11] Muz hujayralararo shakllanishidan so'ng hujayralar qisqaradi, chunki suv ajratilgan muzga yo'qoladi. Hujayralar muzlashdan quriydi, suvsizlanish shikastlanishning asosiy sababi hisoblanadi.
Sovutish tezligi to'qimalarning sovuqqa chidamliligiga ta'sir qiladi,[14] ammo haqiqiy muzlash tezligi nafaqat sovutish tezligiga, balki super sovutish darajasiga va to'qimalarning xususiyatlariga ham bog'liq bo'ladi.[15] Sakai (1979a)[14] Alaskan oq va qora qoraqarag'ay qoraqarag'aylarini asta-sekin 30 ° C dan -40 ° C gacha sovutganda primerdiada muzning ajratilishini namoyish etdi. Bu muzlatilgan suvsiz kurtaklar suvga cho'mgandan omon qoldi suyuq azot sekin qayta isitilganda. Floral primordia xuddi shunday javob berdi. Primordiyadagi ekstraorganik muzlash, havo harorati -50 ° C yoki undan pastroq bo'lgan mintaqalarda boreal ignabargli daraxtlarning qishda eng omon qolish qobiliyatini hisobga oladi.[13] Bundaylarning qish kurtaklarining chidamliligi ignabargli daraxtlar kurtaklarning kichikligi, suvning tezroq ko'chishi evolyutsiyasi va muzlashning intensiv dehidratsiyasiga toqat qilish qobiliyati bilan kuchayadi. Boreal turlarida Picea va Pinus, 1 yoshli ko'chatlarning sovuqqa chidamliligi etuk o'simliklar bilan tenglashadi,[16] uyqudagi o'xshash holatlarni hisobga olgan holda.
Voyaga etmaganlik

Kabi yosh o'simlik tomonidan ishlab chiqarilgan organlar va to'qimalar, masalan ko'chat, ko'pincha yoshi ulg'ayganida bir xil o'simlik tomonidan ishlab chiqariladiganlardan farq qiladi. Ushbu hodisa sifatida tanilgan voyaga etmaganlik yoki heteroblastika. Masalan, yosh daraxtlar to'liq o'sib chiqqan daraxtga qaraganda uzunroq, ingichka novdalar hosil qiladi. Bundan tashqari, erta o'sishda hosil bo'lgan barglar kattalar o'simlikidagi barglarga qaraganda kattaroq, ingichka va tartibsiz bo'ladi. Voyaga etmagan o'simliklarning namunalari bir xil turdagi kattalar o'simliklaridan shunchalik farq qilishi mumkinki, tuxum qo'yadigan hasharotlar o'simlikni o'z bolalari uchun oziq-ovqat sifatida tanimaydilar. Turli xilliklar ildiz otish va gullashda ko'rinadi va ularni bir xil etuk daraxtda ko'rish mumkin. Daraxt tubidan olingan o'spirin so'qmoqlari ildizlarni tojning o'rtasidan yuqori tojigacha hosil bo'lishiga qaraganda ancha oson hosil qiladi. Daraxt poydevoriga yaqin gullash yuqori shoxlarda gullashdan ko'ra yo'q yoki kamroq bo'ladi, ayniqsa yosh daraxt birinchi marta gullash yoshiga etganida.[17]
O'sishning erta shakllaridan kech shakllariga o'tish "deb nomlanadivegetativ faza o'zgarishi ', ammo terminologiya bo'yicha ba'zi kelishmovchiliklar mavjud.[18]
Zamonaviy o'simlik morfologiyasi
Rolf Sattler tushunchasi kabi qiyosiy morfologiyaning fundamental tushunchalarini qayta ko'rib chiqdi homologiya. U gomologiya qisman gomologiya va miqdoriy gomologiyani ham o'z ichiga olishi kerakligini ta'kidladi.[19][20] Bu ildiz, kurtak, poya (kaulom), barg (filom) va soch (trichome) morfologik toifalari o'rtasida davomiylikni namoyish etadigan doimiy morfologiyaga olib keladi. Kategoriyalar orasidagi qidiruv vositalar qanday eng yaxshi tavsiflanganligi haqida Bryus K. Kirxof va boshq.[21] Stalk instituti tomonidan o'tkazilgan yaqinda o'tkazilgan tadqiqotda har bir o'simlik bazasiga va 3D fazosidagi barglarga mos keladigan koordinatalar chiqarildi. Grafadagi o'simliklar ularning ozuqaviy harakatlanish masofalariga va shoxlarning umumiy uzunliklariga qarab joylashtirilganda, o'simliklar Pareto egri chizig'iga deyarli mukammal tushdi. "Bu o'simliklarning o'z me'morchiligini o'stirish uslubi, shuningdek, juda keng tarqalgan tarmoq dizayni o'zgarishini optimallashtiradi degan ma'noni anglatadi. Atrof-muhit va turlarga asoslanib, o'simlik ushbu atrof-muhit sharoitlari uchun savdo qilishning turli usullarini tanlamoqda." [22]
Agnes Arberni hurmat qilish, muallifi bargning qisman otish nazariyasi, Rutishauser va Isler doimiylikni yondashuvni loyqa Arberian morfologiyasi (FAM) deb atashdi. "Loyqa" degan ma'noni anglatadi loyqa mantiq, "Arberian" ga Agnes Arber. Rutishauzer va Isler ushbu yondashuvni nafaqat ko'plab morfologik ma'lumotlar, balki molekulyar genetika dalillari ham qo'llab-quvvatlashini ta'kidladilar.[23] Molekulyar genetikaning so'nggi dalillari doimiy morfologiyani qo'llab-quvvatlaydi. Jeyms (2009) "endi keng tarqalgan bo'lib qabul qilingan ... radiallik [ko'pchilik poyalarga xos] va dorsiventrallik [barglarga xos] uzluksiz spektrning haddan tashqari tomonlari. Aslida bu shunchaki KNOX genining vaqti ifoda !. "[24] Ekkardt va Baum (2010) xulosa qilishicha, "hozirda aralash barglar barg va otish xususiyatlarini ifoda etishi umuman qabul qilingan".[25]
Jarayon morfologiyasi o'simlik shaklining dinamik davomiyligini tavsiflaydi va tahlil qiladi. Ushbu yondashuvga ko'ra, tuzilmalar buni qilmaydi bor jarayon (lar), ular bor jarayon (lar).[26][27][28] Shunday qilib, tuzilish / jarayon dixotomiyasi bizning "tuzilish" tushunchamizni kengayishi bilan bartaraf etiladi, shunda tirik organizmda shunchaki fazoviy tuzilish haqida emas, balki "faoliyat" ga qarshi yoki qarshi bo'lgan narsa haqida gap boradi. u, lekin aniq organizm bu a bo'shliqvaqtinchalik tuzilishi va bu makon-vaqtinchalik tuzilish faoliyatning o'zi ".[29]
Jeune, Barabé va Lacroix uchun klassik morfologiya (ya'ni bir-birini istisno qiladigan toifalarni nazarda tutadigan sifatli homologiya kontseptsiyasiga asoslangan asosiy morfologiya) va doimiy morfologiya ko'proq qamrab oluvchi jarayon morfologiyasining (dinamik morfologiya) kichik sinflari hisoblanadi.[30]
Klassik morfologiya, doimiy morfologiya va jarayon morfologiyasi o'simliklar evolyutsiyasi, ayniqsa o'simliklar morfologiyasi va o'simlik molekulyar genetikasini birlashtirishga harakat qiladigan o'simliklar evolyutsiyasi biologiyasi (o'simlik evo-devo) uchun juda muhimdir.[31] G'ayrioddiy morfologiyalar bo'yicha batafsil amaliy ishda Rutishauser (2016) o'simlik evo-devosining turli xil mavzularini tasvirlab berdi va muhokama qildi, masalan, morfologik tushunchalarning xiralashganligi (davomiyligi), strukturaviy toifalar va gen ekspresiyasi o'rtasida bir-biriga mos kelmaslik. , morfosfera tushunchasi, veranda ludenlariga nisbatan bouplan xususiyatlarining moslashuvchan qiymati, fiziologik moslashuvlar, umidvor hayvonlar va sho'r evolyutsiyasi, rivojlanish mustahkamligining ahamiyati va chegaralari va boshqalar.[32]
Biz xohlaymizmi yoki yo'qmi, morfologik tadqiqotlarga falsafiy taxminlar yoki / yoki mantiq, loyqa mantiq, tuzilish / jarayon dualizmi yoki uning transsendentsiyasi kabi ta'sir qiladi. Va empirik topilmalar falsafiy taxminlarga ta'sir qilishi mumkin. Shunday qilib, falsafa va empirik topilmalar o'rtasida o'zaro ta'sir mavjud. Ushbu o'zaro ta'sirlar o'simliklar morfologiyasi falsafasi deb nomlangan narsaning mavzusidir.[33]
Shuningdek qarang
- O'simliklar morfologiyasining lug'ati
- O'simliklar anatomiyasi
- O'simliklarni identifikatsiyalash
- O'simliklar fiziologiyasi
- O'simliklar evolyutsion rivojlanish biologiyasi
- Taksonomiya
Adabiyotlar
- ^ a b Raven, P. H., R. F. Evert va S. E. Eichhorn. O'simliklar biologiyasi, 7-nashr, 9-bet (Nyu-York: W. H. Freeman, 2005). ISBN 0-7167-1007-2.
- ^ Evert, Rey Franklin va Esov, Ketrin (2006) Esov o'simlik anatomiyasi: o'simlik tanasining meristemalari, hujayralari va to'qimalari - ularning tuzilishi, faoliyati va rivojlanishi Uili, Xoboken, Nyu-Jersi, sahifa xv, ISBN 0-471-73843-3
- ^ Drost, Xaj-Georg; Bellstaedt, Yuliya; Ó'Maoiléidigh, Diarmuid S.; Silva, Anderson T.; Gabel, Aleksandr; Vaynxoldt, Klaus; Rayan, Patrik T.; Dekkers, Bas J.W.; Bentsink, Leoni; Xilxorst, Xenk VM.; Ligterink, Wilco; Vellmer, Frank; Grosse, Ivo; Kvint, Marsel (2016-02-23). "Embriondan keyingi soat soati naqshlari o'simliklarning rivojlanishidagi ontogenetik o'tishni belgilaydi". Molekulyar biologiya va evolyutsiya. 33 (5): 1158–1163. doi:10.1093 / molbev / msw039. PMID 26912813.
- ^ Garold C. Bold, C. J. Aleksopulos va T. Delevoryas. O'simliklar va qo'ziqorinlarning morfologiyasi, 5-nashr, 3. bet (Nyu-York: Harper-Kollinz, 1987). ISBN 0-06-040839-1.
- ^ Byorle, men; Laux, T (2003). "Apikal meristemalar: o'simlikning yoshlar favvorasi". BioEssays. 25 (10): 961–70. doi:10.1002 / bies.10341. PMID 14505363. Ko'rib chiqish.
- ^ Leopold, A. S O'simliklarning o'sishi va rivojlanishi, 183-bet (Nyu-York: McGraw-Hill, 1964).
- ^ Tovar belgisi, U; Xobe, M; Simon, R (2001). "O'simliklar zarbasi meristemalaridagi funktsional domenlar". BioEssays. 23 (2): 134–41. doi:10.1002 / 1521-1878 (200102) 23: 2 <134 :: AID-BIES1020> 3.0.CO; 2-3. PMID 11169586. Ko'rib chiqish.
- ^ Barlow, P (2005). "O'simliklar to'qimasida naqshli hujayralarni aniqlash: daraxtlarning ikkilamchi floemasi". BioEssays. 27 (5): 533–41. doi:10.1002 / bies.20214. PMID 15832381.
- ^ Ross, S.D .; Faris, R.P.; Binder, WD 1983. O'sishni tartibga soluvchilar va ignabargli daraxtlar: ularning fiziologiyasi va o'rmon xo'jaligida potentsial foydalanish. p. 35-78 yilda Nikel, L.G. (Ed.), O'simliklar o'sishini tartibga soluvchi kimyoviy moddalar. Vol. 2, CRC Press, Boca Raton FL.
- ^ Jin JP; va boshq. (2015 yil iyul). "Arabidopsis transkripsiyaviy tartibga solish xaritasi yangi transkripsiya omillarining o'ziga xos funktsional va evolyutsion xususiyatlarini ochib beradi". Molekulyar biologiya va evolyutsiya. 32 (7): 1767–1773. doi:10.1093 / molbev / msv058. PMC 4476157. PMID 25750178.
- ^ a b Glerum, C. 1985. Ignabargli ko'chatlarning sovuqqa chidamliligi: printsiplari va qo'llanilishi. p. 107–123 yilda Duryea, M.L. (Ed.). Ish yuritish: Ko'chatlarning sifatini baholash: asosiy testlarning printsiplari, protseduralari va bashorat qilish qobiliyatlari. Seminar, oktyabr 1984 yil, Oregon shtati universiteti., Uchun. Res. Laboratoriya., Corvallis OR.
- ^ Lyons, JM .; Raison, J.K .; Steponkus, P.L. 1979. Past haroratga javoban o'simlik membranasi: umumiy ko'rinish. p. 1–24 yilda Lyons, JM .; Grem, D.; Raison, J.K. (Eds.). O'simlik o'simliklarida past harorat stresi. Academic Press, Nyu-York, Nyu-York.
- ^ a b Sakay, A .; Larcher, V. (Eds.) 1987. O'simliklarning Frost Survival. Springer-Verlag.
- ^ a b Sakai, A. 1979a. Ignabargli kurtaklarning ibtidoiy kurtaklarining muzlashdan saqlanish mexanizmi. O'simlik hujayralari fizioli. 20: 1381-1390.
- ^ Levitt, J. 1980. O'simliklarning ekologik stresslarga javoblari. Jild 1. Sovutish, muzlash va yuqori harorat stresslari, 2-nashr. Academic Press, Nyu-York, Nyu-York. 497 p.
- ^ Sakay, A .; Okada, S. 1971. Ignalilarning muzlashga chidamliligi. Silva Genet. 20 (3): 91-97.
- ^ Maykl A Dirr; Charlz V Xuzer, kichik (2006). "2". Yog'ochli o'simliklarni ko'paytirish bo'yicha qo'llanma (Ikkinchi nashr). Varsity Press Inc. 26, 28, 29 betlar. ISBN 0942375092.
- ^ Jons, Sintiya S. (1999-11-01). "Urug'lik o'simliklarida voyaga etmaganlik, o'zgarishlar o'zgarishi va heteroblastika to'g'risida insho". Xalqaro o'simlik fanlari jurnali. 160 (S6): –105 – S111. doi:10.1086/314215. ISSN 1058-5893. PMID 10572025.
- ^ Sattler, R. (1984). "Gomologiya - davom etayotgan muammo". Tizimli botanika. 9 (4): 382–394. doi:10.2307/2418787. JSTOR 2418787.
- ^ Sattler, R., 1994, Gomologiya, gomeoz va o'simliklardagi morfologiya. In: B.K. Zal (tahrir) Gomologiya: qiyosiy morfologiyaning ierarxik asoslari. Nyu-York: Academic Press, 423-475 betlar.
- ^ Kirxof, B K; Pfeifer, E; Rutishauser, R (2008). "O'simliklar tuzilishi ontologiyasi: shubhali yoki aralash identifikatsiyaga ega bo'lgan o'simlik tuzilmalarini qanday belgilashimiz kerak?". Zootaxa. 1950: 103–122. doi:10.11646 / zootaxa.1950.1.10.
- ^ Conn, Adam; Pedmale, Ullas; Chori, Joan (2017). "Yuqori aniqlikdagi lazer yordamida skanerlash universal tarmoq dizayni tamoyillarini aks ettiruvchi o'simlik me'morchiligini ochib beradi". Hujayra tizimlari. 5 (1): 103–122. doi:10.1016 / j.cels.2017.06.017. PMID 28750198.
- ^ Rutishauser, R .; Isler, B. (2001). "Gulli o'simliklarning rivojlanish genetikasi va morfologik evolyutsiyasi, ayniqsa siydik pufagi (Utrikulariya): loyqa Arberian morfologiyasi klassik morfologiyani to'ldiradi" (PDF). Botanika yilnomalari. 88 (6): 1173–1202. doi:10.1006 / anbo.2001.1498.
- ^ Jeyms, P. J. (2009). "'Daraxt va barg ': Boshqa burchak ". Linnean. 25: 13–19.
- ^ Ekkardt, NA; Baum, D (2010). "Podostemad jumboq: Podostemaceae-da odatiy bo'lmagan morfologiya evolyutsiyasi". O'simlik hujayrasi. 22 (7): 2131–2140. doi:10.1105 / tpc.110.220711. PMC 2929115. PMID 20647343.
- ^ Sattler, R. (1992). "Jarayon morfologiyasi: rivojlanish va evolyutsiyadagi strukturaviy dinamikasi". Kanada Botanika jurnali. 70 (4): 708–714. doi:10.1139 / b92-091.
- ^ Vergara-Silva, F. (2003). "O'simliklar va evolyutsion rivojlanish biologiyasining kontseptual artikulyatsiyasi". Biologiya va falsafa. 18 (2): 261–264. doi:10.1023 / A: 1023936102602. S2CID 81013686.
- ^ Sattler, R. 2019. O'simliklar shaklining rivojlanishi va evolyutsiyasiga tizimli va dinamik yondashuvlar. In: Fusco, G. (ed) Evolyutsion va rivojlanish biologiyasining istiqbollari. Alessandro Minelli uchun insholar. 6-bob, 57-70-betlar [1]
- ^ Woodger, J.H. 1967 yil. Biologik tamoyillar. London: Routledge & Kegoan Paul (yangi Kirish bilan qayta nashr etilgan).
- ^ Jeune, B; Barabe, D; Lakroix, S (2006). "Klassik va dinamik morfologiya: shakllar maydoni orqali sintezga borishga". Acta Biotheoretica. 54 (4): 277–293. doi:10.1007 / s10441-007-9007-8. PMID 17486414. S2CID 25928998.
- ^ Minelli, A. 2018. O'simliklar evolyutsion rivojlanish biologiyasi. Fenotipning evolyutsiyasi. Nyu-York: Kembrij universiteti matbuoti.
- ^ Rutishauser, R. (2016). "Lentibulariaceae (siydik pufagi va ittifoqdoshlari) va Podostemonaceae (daryo-begona o'tlar) da odatiy bo'lmagan morfologiyalar evolyutsiyasi: rivojlanish biologiyasi va morfologik diversifikatsiya interfeysida tasviriy ma'ruza". Botanika yilnomalari. 117 (5): 811–832. doi:10.1093 / aob / mcv172. PMC 4845801. PMID 26589968.
- ^ Sattler, R. (2018). "O'simliklar morfologiyasi falsafasi". Elemente der Naturwissenschaft. 108: 55–79.(ushbu maqolaning kengaytirilgan versiyasi uchun qarang [2] )
Tashqi havolalar
| Maydonlar |  | |
|---|---|---|
| Bakteriyalar | ||
| Himoyachilar |
| |
| O'simliklar | ||
| Umurtqasiz hayvonlar | ||
| Sutemizuvchilar | ||
| Boshqa umurtqali hayvonlar | ||
| Boshqa mavzular | ||
| ||
| Subdiziplar | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| O'simlik guruhlar | |||||||||||
| |||||||||||
| |||||||||||
| O'simliklarning o'sishi va odati | |||||||||||
| Ko'paytirish | |||||||||||
| O'simliklar taksonomiyasi | |||||||||||
| Amaliyot | |||||||||||
| |||||||||||
| |||||||||||