Sirkadiyalik ritm - Circadian rhythm
| Sirkadiyalik ritm | |
|---|---|
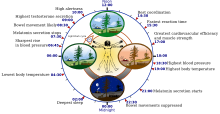 Odam sirkadiyanining (24 soatlik) biologik soatining ba'zi xususiyatlari | |
| Talaffuz | |
| Chastotani | Har 24 soatda bir marta |
A sirkadiyalik ritm uyqudan uyg'onish tsiklini tartibga soluvchi va Yerning har bir aylanishida taxminan har 24 soatda takrorlanadigan tabiiy, ichki jarayondir.[1] An ko'rsatadigan har qanday biologik jarayonga murojaat qilishi mumkin endogen, qiziquvchan tebranish taxminan 24 soat. Ushbu 24 soatlik ritmlarni a boshqaradi sirkadiyalik soat va ular keng kuzatilgan o'simliklar, hayvonlar, qo'ziqorinlar va siyanobakteriyalar.[2]
Atama sirkadiyalik dan keladi Lotin taxminan, "atrofida" (yoki "taxminan") ma'nosini anglatadi va diēm, "kun" ma'nosini anglatadi. Biologik vaqtinchalik ritmlarni, masalan kundalik, to'lqin, haftalik, mavsumiy va yillik ritmlar deyiladi xronobiologiya. 24 soatlik tebranishlar bo'lgan jarayonlar odatda ko'proq chaqiriladi kunlik ritmlar; qat'iyan aytganda, endogen tabiati tasdiqlanmasa, ularni sirkadiyalik ritm deb atash kerak emas.[3]
Sirkadiyalik ritmlar endogen ("ichki", o'z-o'zini qo'llab-quvvatlaydigan) bo'lsa-da, ular tashqi muhit yordamida mahalliy muhitga o'rnatiladi (qo'shiladi) zeitgebers (nemis tilidan "vaqt beruvchi"), bu yorug'lik, harorat va oksidlanish-qaytarilish tsikllar. Tibbiyot fanida odamlarda g'ayritabiiy sirkadiyalik ritm ma'lum sirkadiyalik ritm buzilishi.[4]
2017 yilda Fiziologiya yoki tibbiyot bo'yicha Nobel mukofoti bilan taqdirlandi Jeffri C. Xoll, Maykl Rosbash va Maykl V. Yang "sirkadiyalik ritmni boshqaruvchi molekulyar mexanizmlarning kashfiyotlari uchun" mevali chivinlar.[5]
Tarix
Sharqiy va tub amerikalik madaniyatlarda "tabiiy tana aylanishi" haqida bir necha bor eslatib o'tilgan bo'lsa-da, sirkadiy jarayonining dastlabki G'arbdagi qaydlari miloddan avvalgi IV asrga to'g'ri keladi. Androsthenes, ostida xizmat ko'rsatadigan kema kapitani Buyuk Aleksandr tasvirlangan kunduzgi ning barg harakatlari tamarind daraxt.[6] Odamlarda sirkadiy yoki kunduzgi jarayonning kuzatilishi eslatib o'tilgan Xitoy tibbiy matnlari taxminan 13-asrga tegishli, shu jumladan Tush va yarim tunda qo'llanma va Kunduzgi tsikl, oy kuni va yil fasli bo'yicha aku nuqtalarini tanlashda yordam beradigan mnemonik qofiya..[7]
1729 yilda frantsuz olimi Jan-Jak d'Ortuz de Mairan endogen soatni kundalik ogohlantirishlarga ta'siridan ajratish uchun mo'ljallangan birinchi tajribani o'tkazdi. Uning ta'kidlashicha, o'simlik barglari harakatida 24 soatlik naqshlar Mimoza pudica o'simliklar doimiy zulmatda saqlanganda ham davom etdi.[8][9]
1896 yilda Patrik va Gilbert uzoq vaqt davomida uyqusiz qolish vaqtida uyquchanlik ko'payib, taxminan 24 soat davom etishi bilan kamayishini kuzatdilar.[10] 1918 yilda, J.S. Szimanski yorug'lik va harorat o'zgarishi kabi tashqi belgilar bo'lmasa, hayvonlar 24 soatlik faoliyat tartibini saqlab turishga qodir ekanligini ko'rsatdi.[11]
20-asrning boshlarida asalarilarning ritmik ovqatlanish davrida sirkadiyalik ritmlar kuzatilgan. Ogyust Forel, Ingeborg Beling, va Oskar Vahl ushbu ritmning ichki soatga bog'liqligini aniqlash uchun ko'plab tajribalar o'tkazdi.[12] Sirkadiyalik ritmning mavjudligi mustaqil ravishda kashf etilgan mevali chivin 1935 yilda ikkita nemis zoologlari tomonidan, Xans Kalmus va Ervin Bünning.[13][14]
1954 yilda muhim tajriba tomonidan xabar berilgan Kolin Pittendrigh buni namoyish etdi portlash (pupaning kattalarga aylanish jarayoni) yilda Drosophila pseudoobscura sirkadiyalik xatti-harakatlar edi. Uning ta'kidlashicha, ekloslanish ritmida harorat o'ta muhim rol o'ynagan bo'lsa-da, tutilish davri kechiktirilgan, ammo harorat pasayganda to'xtamagan.[15][14]
Atama sirkadiyalik tomonidan yaratilgan Frants Xelberg 1959 yilda.[16] Halbergning asl ta'rifiga ko'ra:
"Sirkadiyan" atamasi kelib chiqqan taxminan (haqida) va o'ladi (kun); bu ma'lum fiziologik davrlar, agar aynan shu uzunlik bo'lmasa, 24 soatga yaqin ekanligini anglatishi mumkin. Bu erda "tsirkadian" barcha "24 soatlik" ritmlarga, ularning davrlari alohida yoki o'rtacha bo'ladimi-yo'qligiga qaramay, 24 soatdan, uzoqroq yoki qisqaroq, bir necha daqiqa yoki soatga farq qilishi mumkin.[17][18]
1977 yilda Xalqaro nomenklatura qo'mitasi Xalqaro xronobiologiya jamiyati rasmiy ravishda ta'rifni qabul qildi:
Sirkadiyan: 24 ± 4 soat ichida 1 tsikl chastotali biologik o'zgarishlarga yoki ritmlarga taalluqli; taxminan (taxminan, taxminan) va o'ladi (kun yoki 24 soat). Izoh: atama 24 soatlik tsiklning ritmini tavsiflaydi, ular chastotada sinxronlashtiriladimi (qabul qilinadi) yoki mahalliy vaqt shkalasi bilan sinxronlashtiriladimi yoki erkin harakat qiladimi, davri 24 soatdan bir oz farqli o'laroq.[19]
Ron Konopka va Seymur shunga o'xshash soat mutatsiyasini aniqladi Drosophila 1971 yilda gen nomini berish "davr " (per) gen, xulq-atvor ritmikligining birinchi kashf etilgan genetik determinanti.[20] per gen 1984 yilda ikkita tadqiqotchilar guruhi tomonidan ajratilgan. Konopka, Jeffri Xoll, Maykl Roshbash va ularning jamoasi buni ko'rsatdi per lokus sirkadiyalik ritmning markazidir va bu yo'qotish per sirkadiyalik faoliyatni to'xtatadi.[21][22] Shu bilan birga, Maykl V. Yangning jamoasi shunga o'xshash ta'sirlarni qayd etdi perva gen X xromosomasidagi 7,1 kilobaza (kb) oralig'ini qamrab oladi va 4,5 kbli poli (A) + RNKni kodlaydi.[23][24] Ular asosiy genlarni va neyronlarni kashf etishga kirishdilar Drosophila sirkadiy tizim, buning uchun Xoll, Rosbash va Young olganlar Fiziologiya yoki tibbiyot bo'yicha Nobel mukofoti 2017 yil.[5]
Jozef Takaxashi birinchi sutemizuvchilar sirkadiyalik soat mutatsiyasini topdi (soat Δ19) 1994 yilda sichqonlar yordamida.[25][26] Biroq, so'nggi tadqiqotlar shuni ko'rsatadiki, soat xulq-atvor fenotipiga olib kelmaydi (hayvonlar hanuzgacha oddiy tsirkadiyalik ritmlarga ega), bu uning ritm hosil qilishdagi ahamiyatini shubha ostiga qo'yadi.[27][28]
Birinchi inson soati mutatsiyasini Yuta shtatining kengaytirilgan oilasida Kris Jons aniqlagan va genetik jihatdan Ying-Xuy Fu va Lui Ptakeklar tomonidan tavsiflangan. Ta'sirga uchragan shaxslar 4 soatlik yuqori uyqu va boshqa ritmlarga ega bo'lgan o'ta "ertalab laklar" dir. Familial Advanced uyqu fazasining ushbu shakli insonning PER2 oqsilidagi S662➔G aminokislota o'zgarishi natijasida yuzaga keladi.[29][30]
Mezon
Sirkadiy deb atash uchun biologik ritm quyidagi uchta umumiy mezonga javob berishi kerak:[31]
- Ritm endogen erkin harakatlanish davriga ega, u taxminan 24 soat davom etadi. Ritm doimiy sharoitda (ya'ni doimiy zulmatda) davom etadi va taxminan 24 soat davom etadi. Ritmning doimiy sharoitdagi davri erkin yugurish davri deb ataladi va yunoncha τ (tau) harfi bilan belgilanadi. Ushbu mezonning asoslari sirkadiyalik ritmlarni oddiy javoblardan kundalik tashqi signallarga ajratishdir. Ritmni aytish mumkin emas endogen agar u sinovdan o'tkazilmasa va tashqi davriy kiritilmasdan davom etsa. Kunduzgi hayvonlarda (kunduzgi soatlarda faol) umuman τ 24 soatdan biroz kattaroq, tungi hayvonlarda (tunda faol) umuman 24 24 soatdan qisqa.
- Ritmlar o'ziga jalb qiladi. Ritmni tashqi ta'sir (masalan, yorug'lik va issiqlik) ta'sirida tiklash mumkin, bu jarayon deyiladi qiziqish. Ritmni jalb qilish uchun ishlatiladigan tashqi stimulga deyiladi Zeitgeber yoki "vaqt beruvchi". Sayohat bo'ylab vaqt zonalari inson biologik soatining mahalliy vaqtga moslash qobiliyatini aks ettiradi; bir kishi odatda boshdan kechiradi vaqt mintaqasi o'zgarishi xastaligi oldin ularning sirkadiyalik soatlari mahalliy vaqt bilan sinxronlashtirildi.
- Ritmlar haroratni qoplashni namoyish etadi. Boshqacha qilib aytganda, ular fiziologik harorat oralig'ida sirkad davriyligini saqlab turadilar. Ko'pgina organizmlar keng haroratda yashaydilar va issiqlik energiyasidagi farqlar ta'sir qiladi kinetika ularning hujayralaridagi barcha molekulyar jarayonlarning. Vaqtni hisobga olish uchun organizmning sirkadiy soati harorat o'zgarishi deb nomlanuvchi kinetikaga qaramay, taxminan 24 soatlik davriylikni saqlab turishi kerak. The Q10 harorat koeffitsienti bu kompensatsiya ta'sirining o'lchovidir. Agar harorat oshishi bilan Q10 koeffitsienti taxminan 1 bo'lib qolsa, ritm harorat bilan qoplanadi deb hisoblanadi.
Kelib chiqishi
Sirkadiyalik ritmlar organizmlarga atrof-muhitning aniq va muntazam o'zgarishini kutish va tayyorlashga imkon beradi. Shunday qilib, ular organizmlarga atrof-muhit resurslaridan (masalan, yorug'lik va oziq-ovqat) bu kabi mavjudlikni taxmin qila olmaydigan narsalardan yaxshiroq foydalanishga imkon beradi. Shuning uchun sirkadiyalik ritmlar evolyutsiya nuqtai nazaridan organizmlarni tanlab afzalliklarga olib keladi degan fikrlar mavjud. Biroq, ritmiklik tartibga solish va muvofiqlashtirishda muhim ahamiyatga ega ko'rinadi ichki metabolik jarayonlar, bilan muvofiqlashtirganidek atrof-muhit.[32] Bu doimiy laboratoriya sharoitida bir necha yuz avloddan keyin mevali chivinlarda sirkadiyalik ritmlarning saqlanib turishi (merosxo'rligi) tomonidan taklif qilingan,[33] yovvoyi tabiatda doimiy zulmatda bo'lgan jonzotlarda va fiziologik emas, balki xulq-atvorni eksperimental tarzda yo'q qilish orqali bedana.[34][35]
Sirkadiyalik ritmlarni rivojlanishiga undagan narsa sirli savol edi. Oldingi gipotezalarda ta'kidlanishicha, fotosensitiv oqsillar va tsirkadiyan ritmlari dastlabki hujayralarda paydo bo'lishi mumkin, bu esa replikatsiya qilinadigan DNKni yuqori darajadagi zararli moddalardan himoya qilishdir. ultrabinafsha kunduzi nurlanish. Natijada, replikatsiya zulmatga tushdi. Biroq, buning dalillari kam, chunki tsirkad ritmiga ega bo'lgan eng sodda organizmlar - siyanobakteriyalar buning aksini qiladi - ular kunduzi ko'proq bo'linadi.[36] So'nggi tadqiqotlar o'rniga oksidlanish-qaytarilish oqsillarining sirkadiyali osilatorlar bilan birgalikda evolyutsiyasi hayotning uchta sohasidagi quyidagi muhim ahamiyatga ega. Ajoyib Oksidlanish hodisasi taxminan 2,3 milliard yil oldin.[2][4] Hozirgi ko'rinish shundan iboratki, atrof-muhitdagi kislorod darajasidagi va ishlab chiqarishdagi sirkadiyalik o'zgarishlar reaktiv kislorod turlari (ROS) kunduzgi yorug'lik mavjud bo'lganda, ehtimol kundalik ravishda oksidlanish-qaytarilish reaktsiyalarini oldini olish uchun sirkadiyalik ritmlarni rivojlantirish zarurati tug'dirgan.
Eng sodda ma'lum sirkadiyalik soatlar bor bakterial sirkadiyalik ritmlar, prokaryot tomonidan misol keltirilgan siyanobakteriyalar. Yaqinda o'tkazilgan tadqiqotlar shuni ko'rsatdiki, sirkadiyalik soat Synechococcus elongatus qayta tiklanishi mumkin in vitro faqat uchta oqsil bilan (KaiA, KaiB, KaiC )[37] ularning markaziy osilatorining Ushbu soat qo'shilganidan keyin bir necha kun davomida 22 soatlik ritmni ushlab turishi ko'rsatilgan ATP. Ning oldingi izohlari prokaryotik sirkadiyalik timekeeper DNKning transkripsiyasi / tarjimasi bilan qayta aloqa mexanizmiga bog'liq edi.[iqtibos kerak ]
Inson gomologidagi nuqson Drosophila "davr "FASPS uyqu buzilishining sababi sifatida gen aniqlandi (Oilaviy rivojlangan uyqu fazasi sindromi ), evolyutsiya orqali molekulyar sirkadiyalik soatning saqlanib qolgan xususiyatini ta'kidlaydi. Hozirgi kunda biologik soatning ko'plab boshqa genetik tarkibiy qismlari ma'lum. Ularning o'zaro ta'siri natijasida gen hujayralarining bir-biriga bog'langan teskari aloqasi mavjud bo'lib, natijada tana hujayralari kunning ma'lum bir vaqti sifatida talqin qiladi.[38]
Hozir ma'lumki, molekulyar sirkadiyalik soat bitta hujayra ichida ishlashi mumkin; ya'ni hujayra-avtonomdir.[39] Bu tomonidan ko'rsatilgan Gen bloki ajratilgan mollyusk bazal retinal neyronlarda (BRN).[40] Shu bilan birga, turli xil hujayralar bir-biri bilan aloqa qilishlari mumkin, natijada elektr signalizatsiyasi sinxronlashtiriladi. Ular bilan interfeys bo'lishi mumkin ichki sekretsiya bezlari gormonlarning vaqti-vaqti bilan chiqarilishiga olib keladigan miyaning. Ushbu gormonlar retseptorlari tanadan uzoqroq masofada joylashgan bo'lishi va turli organlarning periferik soatlarini sinxronlashtirishi mumkin. Shunday qilib, kunning vaqti haqidagi ma'lumot ko'zlar miyadagi soatga boradi va shu orqali tananing qolgan qismidagi soatlar sinxronlashtirilishi mumkin. Masalan, uxlash / uyg'onish, tana harorati, chanqoqlik va ishtaha vaqti biologik soat tomonidan muvofiqlashtirilib boshqariladi.[41][42]
Hayvonlarda ahamiyati
Sirkadiyalik ritmiklik hayvonlarning, shu jumladan odamlarning uxlash va ovqatlanish tartiblarida mavjud. Tana haroratining aniq namunalari ham mavjud, miya to'lqini faoliyat, gormon ishlab chiqarish, hujayralarni qayta tiklash va boshqa biologik faoliyat. Bunga qo'chimcha, fotoperiodizm, organizmlarning fiziologik reaktsiyasi kun yoki tun davomiyligi, o'simliklar va hayvonlar uchun juda muhimdir va sirkadiya tizimi kun uzunligini o'lchash va talqin qilishda muhim rol o'ynaydi. Ob-havo sharoitining mavsumiy davrlarini, oziq-ovqat mavjudligini yoki yirtqich hayvonlarning faolligini o'z vaqtida bashorat qilish ko'plab turlarning yashashi uchun juda muhimdir. Faqatgina parametr bo'lmasa-da, fotoperiodning o'zgaruvchan uzunligi ("kun bo'yi") fiziologiya va xulq-atvorning mavsumiy vaqti uchun, eng muhimi, migratsiya, qish uyqusi va ko'payish vaqtlari uchun eng taxminiy ekologik belgidir.[43]
Sirkadiy buzilishining ta'siri
Sichqonlardagi mutatsiyalar yoki soat genining yo'q qilinishi uyali / metabolik hodisalarning to'g'ri vaqtini ta'minlash uchun tana soatlarining muhimligini ko'rsatdi; mutant sichqonlar giperfagik va semirib ketgan va glyukoza metabolizmini o'zgartirgan.[44] Sichqonlarda Rev-ErbA alfa soat geni dietadan kelib chiqqan semirishni osonlashtiradi va diabetga moyil bo'lgan glyukoza va lipidlardan foydalanish o'rtasidagi muvozanatni o'zgartiradi.[45] Biroq, odamlarda soat geni polimorfizmlari va metabolik sindromni rivojlanishiga moyilligi o'rtasida kuchli bog'liqlik mavjudmi yoki yo'qligi aniq emas.[46][47]
Yorug'lik-qorong'i tsiklning ta'siri
Ritm qorong'ulik va tsikl tsikli bilan bog'liq. To'liq zulmatda bo'lgan hayvonlar, shu jumladan odamlar, uzoq vaqt davomida a bilan ishlaydi erkin ishlaydigan ritm. Ularning uyqu tsikli har bir "kun" oldinga yoki oldinga suriladi, bu ularning "kuni" ga, ularning endogen 24 soatdan qisqa yoki uzoqroq. Har kuni ritmlarni qayta tiklaydigan atrof-muhit signallari zeitgeberlar deb nomlanadi (nemis tilidan "vaqt ajratuvchilar").[48] Umuman ko'r er osti sutemizuvchilar, masalan. ko'r mol kalamush Spalaks sp., tashqi stimul yo'qligida o'zlarining endogen soatlarini saqlab turishga qodir. Ularda tasvir hosil qiluvchi ko'zlar etishmasa ham, ularning fotoreseptorlari (yorug'likni aniqlaydigan) hanuzgacha ishlaydi; ular vaqti-vaqti bilan ham sirt qilishadi.[sahifa kerak ][49]
Odatda, bir yoki ikkita konsolidatsiyalangan uyqu epizodiga ega bo'lgan erkin harakatlanuvchi organizmlar tashqi signallardan himoyalangan muhitda bo'lishadi, ammo ritm tabiatdagi 24 soatlik qorong'ulik tsikliga bog'liq emas. Bunday holda, uyquni uyg'otish ritmi boshqa sirkadiyaliklar bilan fazadan chiqib ketishi mumkin ultradian metabolik, gormonal, CNS elektr yoki neyrotransmitter ritmlari kabi ritmlar.[50]
So'nggi tadqiqotlar dizayniga ta'sir ko'rsatdi kosmik kemalar atrof-muhit, chunki yorug'lik-qorong'i tsiklni taqlid qiluvchi tizimlar astronavtlar uchun juda foydali ekanligi aniqlandi.[51]
Arktika hayvonlari
Norvegiyalik tadqiqotchilar Tromsø universiteti ba'zi ekanligini ko'rsatdi Arktika hayvonlari (ptarmigan, kiyik ) sirkadiyalik ritmlarni faqat kunning quyoshi va quyosh botishi bo'lgan qismlarida ko'rsating. Kiyiklarning bir tadqiqotida hayvonlar 70 daraja shimol kuzda, qishda va bahorda sirkadiyalik ritmlarni namoyish etdi, ammo yozda emas. Bug'u yoqilgan Svalbard da Shimoliy 78 daraja bunday ritmlarni faqat kuzda va bahorda namoyish etdi. Tadqiqotchilar boshqa Arktika hayvonlari ham yozning doimiy yorug'ida va qishda doimiy qorong'ida sirkadiyalik ritmlarni namoyish etmasligi mumkin deb taxmin qilishmoqda.[52]
Shimoliy Alyaskada 2006 yilda o'tkazilgan tadqiqotlar shuni aniqladiki, kunlik hayot tuproqli sincaplar va tungi kirpiklar 82 kun va tun davomida quyosh nurlari bilan sirkadiyalik ritmlarni qat'iyan saqlab turing. Tadqiqotchilarning fikriga ko'ra, bu ikkita kemiruvchi quyosh va ufq orasidagi aniq masofa kuniga bir marta eng qisqa va shuning uchun ularni jalb qilish (sozlash) uchun etarli signalga ega.[53]
Kelebek va kuya
Ning kuzgi migratsiyasi navigatsiyasi Sharqiy Shimoliy Amerika monarxi kapalagi (Danaus pleksippus) Meksikaning markazida o'zlarining qishlash joylarida antennalaridagi sirkadiyalik soatga bog'liq bo'lgan vaqtni qoplaydigan quyosh kompasidan foydalanadilar.[54][55] Shuningdek, sirkadiyalik ritm ma'lum bir kuya turlarida juftlashish xatti-harakatlarini boshqarishi ham ma'lum Spodoptera littoralis, bu erda urg'ochilar aniq ishlab chiqaradilar feromon tunda juftlashishni boshlash uchun erkak sirkadiyalik ritmni o'ziga jalb qiladi va tiklaydi.[56]
O'simliklarda

O'simliklarning tsirkadiyan ritmlari o'simlikka qaysi fasl va qachon gullashini changlatuvchilarni jalb qilishning eng yaxshi imkoniyati to'g'risida aytib beradi. Ritmlarni ko'rsatadigan xatti-harakatlarga barglar harakati, o'sishi, unib chiqishi, stomatal / gaz almashinuvi, fermentlar faolligi, fotosintez faolligi va parfyum emissiyasi va boshqalar kiradi.[57] Sirkadiyalik ritmlar o'simlik atrofdagi muhitning yorug'lik aylanishi bilan sinxronlash uchun majburlashi natijasida paydo bo'ladi. Ushbu ritmlar ichki hosil bo'ladi va o'zini o'zi ta'minlaydi va atrof-muhit harorati bo'yicha nisbatan doimiydir. Muhim xususiyatlarga ikkita o'zaro ta'sir qiluvchi transkripsiya-tarjima teskari aloqa davri kiradi: protein-oqsillarning o'zaro ta'sirini osonlashtiradigan PAS domenlarini o'z ichiga olgan oqsillar; va soatni har xil yorug'lik sharoitlariga moslashtiradigan bir nechta fotoreseptorlar. Atrof muhitdagi o'zgarishlarni kutish o'simlikning fiziologik holatini mos ravishda o'zgartirishga, adaptiv ustunlik berishga imkon beradi.[58] O'simliklarning tsirkadiyan ritmlarini yaxshiroq anglash qishloq xo'jaligida, masalan, dehqonlarga hosilni yig'ib-terib olishda yordam berish va ob-havo tufayli katta yo'qotishlarga qarshi kurashishda yordam beradi.
Nur - bu o'simliklar o'zlarining ichki soatlarini atrof-muhit bilan sinxronlashtiradigan va turli xil fotoreseptorlar tomonidan seziladigan signaldir. Qizil va ko'k yorug'lik bir nechta fitoxromlar orqali va kriptoxromlar. Bitta fitoxrom, phyA, qorong'ida o'sgan ko'chatlarning asosiy fitoxromidir, ammo nurda tezda parchalanib, Cry1 hosil qiladi. B-E fitoxromlari nurda o'stirilgan ko'chatlarda asosiy fitoxrom bo'lgan fit bilan barqarorroq bo'ladi. Kriptoxrom (qichqiriq) geni ham sirkadiyalik soatning nurga sezgir tarkibiy qismidir va u fotoreseptor sifatida ham, soatning endogen yurak stimulyatori mexanizmining bir qismi sifatida ham ishtirok etadi deb o'ylashadi. 1-2 kriptoxromlari (ko'k-UVA bilan bog'liq) butun yorug'lik sharoitida soat uzunligini saqlab turishga yordam beradi.[57][58]
Markaziy osilator o'zini o'zi ushlab turuvchi ritmni hosil qiladi va uni kunning turli vaqtlarida faol bo'lgan ikkita o'zaro ta'sir qaytaruvchi ko'chadan boshqaradi. Ertalabki tsikl CCA1 (Circadian va Clock-Associated 1) va LHY (kech cho'zilgan gipokotil) dan iborat bo'lib, ular atrofdagi sirkadiyalik ritmlarni tartibga soluvchi MYB transkripsiyasi omillarini kodlaydi. Arabidopsis, shuningdek PRR 7 va 9 (Pseudo-Response Regulators.) Kechki tsikl GI (Gigantea) va ELF4 dan iborat bo'lib, ikkalasi ham gullash vaqtining genlarini boshqarishda ishtirok etadi.[59][60] CCA1 va LHY haddan tashqari ta'sirlanganda (doimiy yorug'lik yoki qorong'i sharoitda) o'simliklar aritmik holga keladi va mRNK signallari kamayadi va salbiy teskari aloqa aylanishiga yordam beradi. CCA1 va LHY ning gen ekspresiyasi tebranadi va erta tongda eng yuqori darajaga etadi TOC1 geni erta kechqurun ifoda tebranadi va eng yuqori nuqtaga etadi. Ilgari ushbu uchta gen salbiy eksklyuziv tsiklni modellashtiradi, unda haddan tashqari ifoda etilgan CCA1 va LHY TOC1ni repressiya qiladi va haddan tashqari ifoda etilgan TOC1 CCA1 va LHY ning ijobiy regulyatori hisoblanadi.[58] 2012 yilda Endryu Millar va boshqalar tomonidan TOC1 aslida CCA1, LHY va PRR7 va 9 ning nafaqat ertalabki tsiklda, balki GI va ELF4 da ham kechqurun repressor bo'lib xizmat qilishi ko'rsatilgan. Ushbu topilma va keyingi hisoblash modellashtirish TOC1 geni funktsiyalar va o'zaro ta'sirlar o'simlik tsirkadiy soatining uch baravar salbiy komponent sifatida qayta tuzilishini taklif qiladi repressilator sutemizuvchilarda soatni tavsiflovchi ijobiy / manfiy elementlarning teskari aloqasi o'rniga model.[61]
2018 yilda tadqiqotchilar PRR5 va TOC1 hnRNA tug'ma transkriptlarining ekspressioni A.thaliana-da ritmik ravishda qayta ishlangan mRNA transkriptlari bilan bir xil tebranish sxemasiga mos kelishini aniqladilar. LNKs PRR5 va TOC1 ning 5 mintaqasi bilan bog'lanib, RNAP II va boshqa transkripsiya omillari bilan o'zaro ta'sir qiladi. . Bundan tashqari, RVE8-LNKlarning o'zaro ta'siri, ruxsat etilgan histon-metilasyon naqshini (H3K4me3) o'zgartirishga imkon beradi va giston modifikatsiyasining o'zi soat geni ekspresiyasining tebranishiga parallel.[62]
Ilgari o'simlikning tsirkadiy ritmini tashqi muhitning qorong'i va qorong'u davrlariga mos kelishi o'simlikka ijobiy ta'sir ko'rsatishi mumkinligi aniqlangan edi.[63] Tadqiqotchilar uch xil nav bo'yicha tajribalar o'tkazib, shunday xulosaga kelishdi Arabidopsis talianasi. Ushbu navlardan biri odatiy 24 soatlik tsirkadiyalik tsiklga ega edi.[63] Qolgan ikkita nav mutatsiyaga uchragan, ulardan biri sirkadiyalik tsikl 27 soatdan ortiq, bittasi oddiy sirkadiyalik tsikl tsikli 20 soatdan iborat.[63]
24 soatlik tsirkad tsikli bilan ishlaydigan Arabidopsis uch xil muhitda o'stirildi.[63] Ushbu muhitlarning birida 20 soatlik yorug'lik va qorong'i tsikl (10 soat yorug'lik va 10 soat qorong'i), ikkinchisida 24 soat yorug'lik va qorong'i tsikl (12 soat yorug'lik va 12 soat qorong'i) va yakuniy muhit 28 soatlik qorong'i va qorong'i tsiklga ega edi (14 soat yorug'lik va 14 soat qorong'i).[63] Ikkita mutatsiyaga uchragan o'simliklar 20 soatlik yorug'lik va qorong'u tsiklga ega bo'lgan muhitda ham, 28 soatlik yorug'lik va qorong'i tsiklga ega bo'lgan muhitda o'stirildi.[63] 24 soatlik yorug'lik va qorong'i tsiklga ega bo'lgan muhitda 24 soatlik tsirkadiy ritm tsikli bilan arabidopsisning xilma-xilligi eng yaxshi o'sganligi aniqlandi.[63] Umuman olganda, barcha navlari aniqlandi Arabidopsis talianasi ko'proq xlorofill darajalariga ega va yorug'lik va qorong'u tsikllar o'zlarining tsirkadiy ritmiga mos keladigan muhitda o'sishni oshirgan.[63]
Tadqiqotchilar buning sababi Arabidopsisning sirkadiyalik ritmini uning atrof-muhitiga mos kelishi o'simlikning tong va shom tushishiga yaxshiroq tayyorlanishiga va shu bilan uning jarayonlarini yaxshiroq sinxronlashtirishga imkon berishi mumkin deb taxmin qilishdi.[63] Ushbu tadqiqotda xlorofillni boshqarishda yordam beradigan genlar tong otgandan bir necha soat o'tgach eng yuqori darajaga ko'tarilganligi aniqlandi.[63] Bu metabolik shafaq deb nomlanuvchi taklif qilingan hodisaga mos keladi.[64]
Metabolik tong gipotezasiga ko'ra, fotosintez natijasida hosil bo'lgan qandlar tsirkad ritmini va ba'zi fotosintez va metabolizm yo'llarini boshqarishda yordam beradi.[64][65] Quyosh ko'tarilishi bilan ko'proq yorug'lik paydo bo'ladi, bu odatda ko'proq fotosintez sodir bo'lishiga imkon beradi.[64] Fotosintez natijasida hosil bo'lgan shakar PRR7ni siqib chiqaradi.[66] Ushbu PRR7 repressiyasi keyinchalik CCA1 ekspressionining kuchayishiga olib keladi.[66] Boshqa tomondan, fotosentetik shakar darajasining pasayishi PRR7 ekspressionini oshiradi va CCA1 ekspressionini pasaytiradi.[64] CCA1 va PRR7 o'rtasidagi ushbu teskari aloqa metabolik tongni keltirib chiqaradigan narsa.[64][67]
Yilda Drosophila
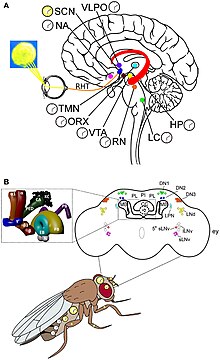
Sirkadiyalik ritm va yorug'likni sezishning molekulyar mexanizmi eng yaxshi tushuniladi Drosophila. Soat genlari topilgan Drosophilava ular soat neyronlari bilan birgalikda harakat qilishadi. Ikkita noyob ritm mavjud, ulardan biri lyuklash jarayonida (deyiladi) portlash ) kukladan, ikkinchisi esa juftlash paytida.[68] Soat neyronlari markaziy miyada alohida klasterlarda joylashgan. Eng yaxshi tushunilgan soat neyronlari bu katta va kichik lateral ventral neyronlar (l-LNvs va s-LNvs). optik lob. Ushbu neyronlar pigmentlarning tarqaladigan omilini (PDF) ishlab chiqaradi, bu neyropeptid bo'lib, u har xil soat neyronlari o'rtasida sirkadiyalik neyromodulyator vazifasini bajaradi.[69]

Drosophila sirkadiyalik ritm transkripsiya-tarjima teskari aloqasi orqali amalga oshiriladi. Asosiy soat mexanizmi o'zaro bog'liq ikkita teskari aloqa tsiklidan iborat, ya'ni PER / TIM tsikli va CLK / CYC tsikli.[70] CLK / CYC tsikli kun davomida sodir bo'ladi va transkripsiyasini boshlaydi per va tim genlar. Ammo ularning oqsillari shom tushguncha past bo'lib qoladi, chunki kunduzi ham faollashadi ikkilamchi vaqt (dbt) gen. DBT oqsili monomerik PER oqsillarining fosforlanishiga va aylanishiga olib keladi.[71][72] TIM, shuningdek, quyosh botguncha shaggy tomonidan fosforillanadi. Quyosh botganidan keyin DBT yo'qoladi, shuning uchun PER molekulalari TIM bilan barqaror bog'lanadi. PER / TIM dimer tunda bir necha marta yadroga kiradi va CLK / CYC dimerlari bilan bog'lanadi. Bound PER CLK va CYC ning transkripsiya faolligini to'liq to'xtatadi.[73]
Erta tongda yorug'lik yig'lamoq geni va uning oqsili CRY TIM ning parchalanishiga olib keladi. Shunday qilib PER / TIM dimer ajralib chiqadi va bog'lanmagan PER beqaror bo'ladi. PER progressiv fosforillanish va oxir-oqibat degradatsiyaga uchraydi. PER va TIM yo'qligi faollashtirishga imkon beradi clk va tsikl genlar. Shunday qilib, keyingi sirkadiyalik tsiklni boshlash uchun soat tiklanadi.[74]
PER-TIM modeli
Ushbu oqsil modeli Drozofiladagi PER va TIM oqsillarining tebranishlari asosida ishlab chiqilgan.[75] Uning asosi PER modeliga asoslangan bo'lib, u erda per gen va uning oqsili biologik soatga qanday ta'sir qilishi tushuntirildi.[76] Model, per va tim genlarining transkripsiyasiga (salbiy teskari aloqani ta'minlash orqali) va ushbu ikki oqsilning ko'p fosforlanishiga ta'sir qiluvchi yadro PER-TIM kompleksini shakllantirishni o'z ichiga oladi. Ushbu ikkita oqsilning sirkadiyali tebranishlari, agar ular unga bog'liq bo'lmasa ham, qorong'u qorong'i tsikl bilan sinxronlashadi.[77][75] Ikkala PER va TIM oqsillari ham fosforillanadi va ular PER-TIM yadro kompleksini hosil qilgandan so'ng ular per va tim mRNA ekspressionini to'xtatish uchun yadro ichiga qaytadilar. Ushbu inhibisyon oqsilgacha davom etadi yoki mRNK parchalanmaydi.[75] Bu sodir bo'lganda, kompleks inhibisyonni chiqaradi. Bu erda TIM oqsilining parchalanishi yorug'lik bilan tezlashishi haqida ham aytib o'tish mumkin.[77]
Sutemizuvchilarda
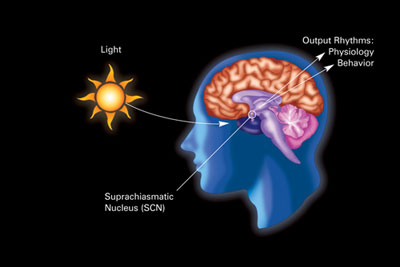
Birlamchi sirkadiyalik soat yilda sutemizuvchilar da joylashgan supraxiyazmatik yadro (yoki yadrolari) (SCN), alohida guruhlarning juftligi hujayralar joylashgan gipotalamus. SCNni yo'q qilish muntazam ravishda uyqudan uyg'onish ritmining yo'qligiga olib keladi. SCN yorug'lik haqida ma'lumotni ko'z orqali oladi. The retina ko'zda "klassik" mavjud fotoreseptorlar ("tayoqchalar "va"konuslar "), odatiy ko'rish uchun ishlatiladi. Ammo retinada ixtisoslashgan ham mavjud ganglion hujayralari to'g'ridan-to'g'ri fotosensitiv va to'g'ridan-to'g'ri SCN-ga loyihalashadi, bu erda ular ushbu asosiy sirkadiyalik soatni jalb qilishda (sinxronlashtirishda) yordam beradi.[78]
Ushbu hujayralar fotopigmentni o'z ichiga oladi melanopsin va ularning signallari the deb nomlangan yo'ldan boradi retinohipotalamik trakt, SCN ga olib boradi. Agar SCN dan hujayralar olib tashlansa va o'stirilsa, tashqi belgilar bo'lmasa, ular o'zlarining ritmini saqlab turadilar.[79]
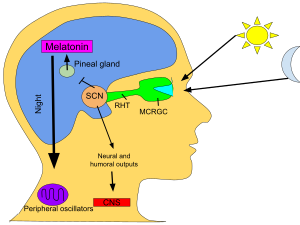
SCN kecha va kunduz davomiyligi haqidagi ma'lumotlarni retinadan oladi, uni sharhlaydi va epifiz bezi, a kabi shakllangan mayda inshoot qarag'ay konusi va joylashgan epitalamus. Bunga javoban epifal gormonni chiqaradi melatonin.[80] Kechasi melatonin sekretsiyasi va kunduzi havo pasayadi va uning mavjudligi tun bo'yi haqida ma'lumot beradi.
Bir necha tadkikotlar shuni ko'rsatdiki, epifel melatonin sirkadiyalik faoliyat turlarini va boshqa jarayonlarni modulyatsiya qilish uchun SCN ritmliligi bilan oziqlanadi. Biroq, ushbu mulohazaning mohiyati va tizim darajasidagi ahamiyati noma'lum.[81]
Odamlarning sirkadiyalik ritmlari Yerning 24 soatiga nisbatan bir oz qisqaroq va uzoqroq muddatlarga ega bo'lishi mumkin. Garvard tadqiqotchilari inson sub'ektlarini hech bo'lmaganda 23,5 soatlik tsiklga va 24,65 soatlik tsiklga (ikkinchisi sayyoradagi tabiiy quyosh kecha-kunduz tsikliga) jalb qilish mumkinligini ko'rsatdilar. Mars ).[82]
Odamlar
Sirkadiyalik ritmlarni o'rganish bo'yicha dastlabki tadqiqotlar shuni ko'rsatdiki, ko'pchilik odamlar kunduzgi yorug'lik va vaqtni saqlash kabi tashqi ta'sirlardan ajratilganda 25 soatga yaqin kunni afzal ko'rishadi. Biroq, ushbu tadqiqot noto'g'ri edi, chunki u ishtirokchilarni sun'iy nurdan himoya qila olmadi. Mavzular (soat kabi) va kunduzgi yorug'likdan himoyalangan bo'lsa-da, tadqiqotchilar ichki elektr chiroqlarning fazani kechiktiradigan ta'siridan xabardor emas edilar.[83][shubhali ] Mavzularga uyg'onganda yorug'likni yoqishga va uxlashni xohlaganda uni o'chirishga ruxsat berildi. Kechqurun elektr nuri ularning sirkadiy fazasini kechiktirdi.[84] 1999 yilda o'tkazilgan yanada qat'iy tadqiqot Garvard universiteti insonning tabiiy ritmini 24 soat 11 daqiqaga yaqinroq deb taxmin qildi: ga juda yaqin quyosh kuni.[85] Ushbu tadqiqotga muvofiq, 2010 yildagi so'nggi tadqiqotlar bo'lib, unda erkaklarnikiga qaraganda (24.09 soat) ayollar uchun bir oz qisqaroq (24.09 soat) bo'lgan tsirkad davri bilan jinsiy farqlar aniqlandi.[86] Ushbu tadqiqotda ayollar erkaklarnikidan ko'ra erta uyg'onishga moyil edilar va erkaklarnikiga qaraganda ertalabki mashg'ulotlarga ko'proq ustunlik berishadi, ammo bu farqlarning asosiy biologik mexanizmlari noma'lum.[86]
Biologik markerlar va effektlar
Sutemizuvchilarning sirkadiyan ritmining vaqtini o'lchash uchun klassik faz belgilari:
- melatonin tomonidan sekretsiya epifiz bezi,[87]
- tana harorati eng kam,[87] va
- plazma darajasi kortizol.[88]
Haroratni o'rganish uchun mavzular hushyor turishi kerak, ammo rektumdagi harorat doimiy ravishda olinib turganda, yaqin qorong'ida tinch va yarim egiluvchan bo'lishi kerak. O'zgarishlar odatdagidek juda yaxshi xronotiplar, kattalardagi odamlarning o'rtacha harorati odatdagi uyg'onish vaqtidan taxminan ikki soat oldin taxminan soat 5:00 da minimal darajaga etadi. Baehr va boshq.[89] yosh kattalarda kunlik tana harorati ertalabki turlar uchun soat 04:00 da (soat 4 da), ammo kechqurun turlari uchun soat 06:00 da (soat 6 da) sodir bo'lganligi aniqlandi. Ushbu minimal ko'rsatkich ertalabki turlar uchun taxminan sakkiz soatlik uyqu davrining o'rtalarida sodir bo'lgan, ammo kechqurun uyg'onishga yaqinroq bo'lgan.
Melatonin tizimda yo'q yoki kunduzi aniqlanmaydi. Uning xira nurda boshlanishi, xira nurli melatonin boshlanishi (DLMO), taxminan soat 21:00 da (21:00) qon yoki tupurik bilan o'lchash mumkin. Uning asosiy qismi metabolit shuningdek, ertalab siydik bilan o'lchanishi mumkin. Ikkala DLMO va qonda yoki tupurikda gormon mavjudligining o'rta nuqtasi (o'z vaqtida) sirkadiyalik belgilar sifatida ishlatilgan. Ammo, yangi tadqiqotlar shuni ko'rsatadiki, melatonin ofset ishonchli marker bo'lishi mumkin. Benlousif va boshq.[87] melatonin fazasi markerlari eng past haroratga qaraganda ancha barqaror va uxlash vaqti bilan ko'proq bog'liqligini aniqladi. Ularning fikriga ko'ra, uyquni qoplash va melatonin bilan qoplash uyqu boshlanishidan ko'ra faza markerlari bilan ko'proq bog'liqdir. Bundan tashqari, melatonin darajasining pasayish bosqichi melatonin sintezining tugashiga qaraganda ancha ishonchli va barqarordir.
Sirkadiyalik ritmga ko'ra sodir bo'ladigan boshqa fiziologik o'zgarishlar kiradi yurak urish tezligi va ko'plab uyali jarayonlar ", shu jumladan oksidlovchi stress, hujayra metabolizmi immunitet va yallig'lanish reaktsiyalari, epigenetik o'zgartirish, gipoksiya /giperoksiya javob berish yo'llari, endoplazmatik retikulyar stress, avtofagiya va tartibga solish ildiz hujayrasi atrof-muhit. "[90] Yigitlarni o'rganish paytida, yurak urish tezligi uxlash vaqtida o'rtacha eng past ko'rsatkichga va uyg'onganidan ko'p o'tmay eng yuqori ko'rsatkichga erishishi aniqlandi.[91]
Oldingi tadqiqotlarga zid ravishda, tana haroratining psixologik testlarda ishlashga ta'siri yo'qligi aniqlandi. Bu, avvalgi tadqiqotlarda ko'rib chiqilgan funktsiyalarning boshqa sohalariga nisbatan yuqori bilim funktsiyasi uchun evolyutsion bosim tufayli yuzaga kelishi mumkin.[92]
"Asosiy soat" dan tashqarida
Oz-ozdan mustaqil sirkadiyalik ritmlar suprachiasmatik yadrolardan (SCN) tashqaridagi tanadagi ko'plab organlar va hujayralarda uchraydi, "master soat". Darhaqiqat, nevrolog olim Jozef Takaxashi va uning hamkasblari 2013 yilgi maqolasida "tanadagi deyarli har bir hujayrada sirkadiyalik soat bor" deb ta'kidlagan edi.[93] Masalan, periferik osilator deb ataladigan bu soatlar buyrak usti bezidan topilgan, qizilo'ngach, o'pka, jigar, oshqozon osti bezi, taloq, timus va teri.[94][95][96] Xushbo'y lampochkaning mavjudligiga oid ba'zi dalillar mavjud[97] va prostata[98] hech bo'lmaganda madaniylashtirilganda tebranishlarga duch kelishi mumkin.
Teridagi osilatorlar nurga ta'sir etsa-da, tizimli ta'sir isbotlanmagan.[99] Bundan tashqari, masalan, jigar hujayralari kabi ko'plab osilatorlar yorug'likdan tashqari, masalan, oziqlantirish kabi kirishga javob berishi aniqlangan.[100]
Nur va biologik soat
Yorug'lik biologik soatni qayta tiklaydi fazali javob egri chizig'i (XXR). Vaqtga qarab, yorug'lik sirkadiyalik ritmni oshirishi yoki kechiktirishi mumkin. Ham XXR, ham talab qilinadi yorug'lik turlardan turlarga farq qiladi va soatlarni qayta tiklash uchun yorug'likning past darajalari talab qilinadi tungi kemiruvchilar odamlarga qaraganda.[101]
Uzunroq yoki qisqaroq tsikllar
Odamlar ustida olib borilgan turli xil tadqiqotlar majburiy uyqu / uyg'onish davrlarini 24 soatdan farq qiladi, masalan Nataniel Kleitman 1938 yilda (28 soat) va Derk-Yan Dijk va Charlz Chezler 1990-yillarda (20 soat). Because normal people cannot entrain to such abnormal day/night rhythms,[102] this is referred to as a forced desynchrony protocol. Under such a protocol, sleep and wake episodes are uncoupled from the body's endogenous circadian period, which allows researchers to assess the effects of circadian phase (i.e. the relative timing of the circadian cycle) on aspects of sleep and wakefulness including uyqu kechikishi and other functions - both physiological, behavioral, and cognitive.[103][104][105][106][107]
Inson salomatligi

Timing of medical treatment in coordination with the body clock, chronotherapeutics, may significantly increase efficacy and reduce drug toxicity or adverse reactions.[108]
A number of studies have concluded that a short period of sleep during the day, a power-nap, does not have any measurable effect on normal circadian rhythms but can decrease stress and improve productivity.[109][110][111]
Health problems can result from a disturbance to the circadian rhythm.[112] Circadian rhythms also play a part in the retikulyar faollashtiruvchi tizim, which is crucial for maintaining a state of consciousness. A reversal[tushuntirish kerak ] in the sleep–wake cycle may be a sign or complication of uremiya,[113] azotemiya yoki buyrakning o'tkir shikastlanishi.[114][115]
Studies have also shown that light has a to'g'ridan-to'g'ri ta'sir on human health because of the way it influences the circadian rhythms.[116]
Indoor lighting
Lighting requirements for circadian regulation are not simply the same as those for vision; planning of indoor lighting in offices and institutions is beginning to take this into account.[117] Animal studies on the effects of light in laboratory conditions have until recently considered light intensity (nurlanish ) but not color, which can be shown to "act as an essential regulator of biological timing in more natural settings".[118]
Obesity and diabetes
Semirib ketish va diabet are associated with lifestyle and genetic factors. Among those factors, disruption of the circadian clockwork and/or misalignment of the circadian timing system with the external environment (e.g., light–dark cycle) might play a role in the development of metabolic disorders.[112]
Smenali ish or chronic vaqt mintaqasi o'zgarishi xastaligi have profound consequences for circadian and metabolic events in the body. Animals that are forced to eat during their resting period show increased body mass and altered expression of clock and metabolic genes.[119][tibbiy ma'lumotnoma kerak ] In humans, shift work that favors irregular eating times is associated with altered insulin sensitivity and higher body mass. Shift work also leads to increased metabolic risks for cardio-metabolic syndrome, hypertension, and inflammation.[120]
Airline pilots and cabin crew
Due to the work nature of airline pilots, who often cross several time zones and regions of sunlight and darkness in one day, and spend many hours awake both day and night, they are often unable to maintain sleep patterns that correspond to the natural human circadian rhythm; this situation can easily lead to charchoq. The NTSB cites this as contributing to many accidents[121] and has conducted several research studies in order to find methods of combating fatigue in pilots.[122]
Buzilish
Disruption to rhythms usually has a negative effect. Many travelers have experienced the condition known as vaqt mintaqasi o'zgarishi xastaligi, with its associated symptoms of charchoq, disorientation, and uyqusizlik.[123]
A number of other disorders, for example bipolyar buzilish va ba'zilari uyqu buzilishi kabi kechiktirilgan uyqu fazasi buzilishi (DSPD), are associated with irregular or pathological functioning of circadian rhythms.[124]
Disruption to rhythms in the longer term is believed to have significant adverse health consequences for peripheral organs outside the brain, in particular in the development or exacerbation of cardiovascular disease.[112][125] Blue LED lighting suppresses melatonin production five times more than the orange-yellow high-pressure sodium (HPS) light; a metal halide lamp, which is white light, suppresses melatonin at a rate more than three times greater than HPS.[126] Depression symptoms from long term nighttime light exposure can be undone by returning to a normal cycle.[127]
Effect of drugs
Studies conducted on both animals and humans show major bidirectional relationships between the circadian system and abusive drugs. It is indicated that these abusive drugs affect the central circadian pacemaker. Individuals suffering from substance abuse display disrupted rhythms. These disrupted rhythms can increase the risk for substance abuse and relapse. It is possible that genetic and/or environmental disturbances to the normal sleep and wake cycle can increase the susceptibility to addiction.[128]
It is difficult to determine if a disturbance in the circadian rhythm is at fault for an increase in prevalence for substance abuse or if other environmental factors such as stress are to blame.Changes to the circadian rhythm and sleep occur once an individual begins abusing drugs and alcohol. Once an individual chooses to stop using drugs and alcohol, the circadian rhythm continues to be disrupted.[128]
The stabilization of sleep and the circadian rhythm might possibly help to reduce the vulnerability to addiction and reduce the chances of relapse.[128]
Circadian rhythms and clock genes expressed in brain regions outside the supraxiyazmatik yadro may significantly influence the effects produced by drugs such as kokain.[iqtibos kerak ] Moreover, genetic manipulations of clock genes profoundly affect cocaine's actions.[129]
Jamiyat va madaniyat
2017 yilda, Jeffri C. Xoll, Maykl V. Yang va Maykl Rosbash taqdirlandi Fiziologiya yoki tibbiyot bo'yicha Nobel mukofoti "for their discoveries of molecular mechanisms controlling the circadian rhythm".[130][131]
Shuningdek qarang
- Aktigrafiya (also known as actimetry)
- ARNTL
- ARNTL2
- Bakterial sirkadiyalik ritmlar
- Sirkadiyalik ritmning uyqusizliklari, kabi
- Xronobiologiya
- SAAT
- Circasemidian rhythm
- Sirkaseptan, 7 kunlik biologik tsikl
- Kriptoxrom
- YIG'LASH1 va CRY2: the cryptochrome family genes
- Kundalik tsikl
- Sirkadiyalik ritmga yorug'lik effektlari
- Light in school buildings
- PER1, PER2 va PER3: the period family genes
- Fotosensitiv ganglion hujayrasi: part of the eye which is involved in regulating circadian rhythm.
- Polifazik uyqu
- Rev-ErbA alfa
- Segmentli uyqu
- Kutish me'morchiligi (sleep in humans)
- Sleep in non-human animals
- Stefaniya Follini
Adabiyotlar
- ^ "What makes us sleep?". NICHD - Eunice Kennedy Shriver National Institute of Child Health and Human Development. Olingan 6 may 2019.
- ^ a b Edgar RS, Green EW, Zhao Y, van Ooijen G, Olmedo M, Qin X, et al. (2012 yil may). "Peroxiredoxins are conserved markers of circadian rhythms". Tabiat. 485 (7399): 459–64. Bibcode:2012Natur.485..459E. doi:10.1038/nature11088. PMC 3398137. PMID 22622569.
- ^ Vitaterna MH, Takahashi JS, Turek FW (2001). "Overview of circadian rhythms". Alcohol Research & Health. 25 (2): 85–93. PMC 6707128. PMID 11584554.
- ^ a b Bass J (November 2012). "Circadian topology of metabolism". Tabiat. 491 (7424): 348–56. Bibcode:2012Natur.491..348B. doi:10.1038/nature11704. PMID 23151577. S2CID 27778254.
- ^ a b "The Nobel Prize in Physiology or Medicine 2017". www.nobelprize.org. Olingan 2017-10-06.
- ^ Bretzl H (1903). Botanische Forschungen des Alexanderzuges. Leypsig: Teubner.[sahifa kerak ]
- ^ Lu G (25 October 2002). Celestial Lancets. Psixologiya matbuoti. 137-140 betlar. ISBN 978-0-7007-1458-2.
- ^ de Mairan JJ (1729). "Observation Botanique". Histoire de l'Académie Royale des Sciences: 35–36.
- ^ Gardner MJ, Hubbard KE, Hotta CT, Dodd AN, Webb AA (July 2006). "How plants tell the time". Biokimyoviy jurnal. 397 (1): 15–24. doi:10.1042/BJ20060484. PMC 1479754. PMID 16761955.
- ^ Dijk DJ, von Schantz M (August 2005). "Timing and consolidation of human sleep, wakefulness, and performance by a symphony of oscillators". Biologik ritmlar jurnali. 20 (4): 279–90. doi:10.1177/0748730405278292. PMID 16077148. S2CID 13538323.
- ^ Danchin A. "Important dates 1900–1919". HKU-Pasteur Research Centre. Arxivlandi asl nusxasi 2003-10-20. Olingan 2008-01-12.
- ^ Antle MC, Silver R (November 2009). "Neural basis of timing and anticipatory behaviors". Evropa nevrologiya jurnali. 30 (9): 1643–9. doi:10.1111/j.1460-9568.2009.06959.x. PMC 2929840. PMID 19878281.
- ^ Bruce VG, Pittendrigh CS (1957). "Endogenous Rhythms in Insects and Microorganisms". Amerikalik tabiatshunos. 91 (858): 179–195. doi:10.1086/281977. S2CID 83886607.
- ^ a b Pittendrigh CS (1993). "Temporal organization: reflections of a Darwinian clock-watcher". Fiziologiyaning yillik sharhi. 55 (1): 16–54. doi:10.1146/annurev.ph.55.030193.000313. PMID 8466172. S2CID 45054898.
- ^ Pittendrigh CS (October 1954). "On Temperature Independence in the Clock System Controlling Emergence Time in Drosophila". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 40 (10): 1018–29. Bibcode:1954PNAS...40.1018P. doi:10.1073/pnas.40.10.1018. PMC 534216. PMID 16589583.
- ^ Halberg F, Cornélissen G, Katinas G, Syutkina EV, Sothern RB, Zaslavskaya R, et al. (2003 yil oktyabr). "Transdisciplinary unifying implications of circadian findings in the 1950s". Journal of Circadian Rhythms. 1 (1): 2. doi:10.1186/1740-3391-1-2. PMC 317388. PMID 14728726.
Eventually I reverted, for the same reason, to "circadian" ...
- ^ Halberg F (1959). "[Physiologic 24-hour periodicity; general and procedural considerations with reference to the adrenal cycle]". Internationale Zeitschrift für Vitaminforschung. Beiheft. 10: 225–96. PMID 14398945.
- ^ Koukkari WL, Sothern RB (2006). Introducing Biological Rhythms: A Primer on the Temporal Organization of Life, with Implications for Health, Society, Reproduction, and the Natural Environment. Nyu-York: Springer. p. 23. ISBN 978-1-4020-3691-0.
- ^ Halberg F, Carandente F, Cornelissen G, Katinas GS (1977). "[Glossary of chronobiology (author's transl)]". Xronobiologiya. 4 Suppl 1: 1–189. PMID 352650.
- ^ Konopka RJ, Benzer S (September 1971). "Clock mutants of Drosophila melanogaster". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 68 (9): 2112–6. Bibcode:1971PNAS...68.2112K. doi:10.1073/pnas.68.9.2112. PMC 389363. PMID 5002428.
- ^ Reddy P, Zehring WA, Wheeler DA, Pirrotta V, Hadfield C, Hall JC, Rosbash M (October 1984). "Molecular analysis of the period locus in Drosophila melanogaster and identification of a transcript involved in biological rhythms". Hujayra. 38 (3): 701–10. doi:10.1016/0092-8674(84)90265-4. PMID 6435882. S2CID 316424.
- ^ Zehring WA, Wheeler DA, Reddy P, Konopka RJ, Kyriacou CP, Rosbash M, Hall JC (December 1984). "P-element transformation with period locus DNA restores rhythmicity to mutant, arrhythmic Drosophila melanogaster". Hujayra. 39 (2 Pt 1): 369–76. doi:10.1016/0092-8674(84)90015-1. PMID 6094014.
- ^ Bargiello TA, Jackson FR, Young MW (1984). "Restoration of circadian behavioural rhythms by gene transfer in Drosophila". Tabiat. 312 (5996): 752–4. Bibcode:1984Natur.312..752B. doi:10.1038/312752a0. PMID 6440029. S2CID 4259316.
- ^ Bargiello TA, Young MW (April 1984). "Molecular genetics of a biological clock in Drosophila". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 81 (7): 2142–6. Bibcode:1984Natur.312..752B. doi:10.1038/312752a0. PMC 345453. PMID 16593450.
- ^ [ishonchsiz tibbiy manbami? ] "Gene Discovered in Mice that Regulates Biological Clock". Chicago Tribune. 1994 yil 29 aprel.
- ^ [birlamchi bo'lmagan manba kerak ] Vitaterna MH, King DP, Chang AM, Kornhauser JM, Lowrey PL, McDonald JD, et al. (1994 yil aprel). "Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior". Ilm-fan. 264 (5159): 719–25. Bibcode:1994Sci...264..719H. doi:10.1126/science.8171325. PMC 3839659. PMID 8171325.
- ^ Debruyne JP, Noton E, Lambert CM, Maywood ES, Weaver DR, Reppert SM (May 2006). "A clock shock: mouse CLOCK is not required for circadian oscillator function". Neyron. 50 (3): 465–77. doi:10.1016/j.neuron.2006.03.041. PMID 16675400. S2CID 19028601.
- ^ Collins B, Blau J (May 2006). "Keeping time without a clock". Neyron. 50 (3): 348–50. doi:10.1016/j.neuron.2006.04.022. PMID 16675389.
- ^ Toh KL, Jones CR, He Y, Eide EJ, Hinz WA, Virshup DM, et al. (2001 yil fevral). "An hPer2 phosphorylation site mutation in familial advanced sleep phase syndrome". Ilm-fan. 291 (5506): 1040–3. Bibcode:2001 yil ... 291.1040T. doi:10.1126 / science.1057499. PMID 11232563. S2CID 1848310.
- ^ Jones CR, Campbell SS, Zone SE, Cooper F, DeSano A, Murphy PJ, et al. (1999 yil sentyabr). "Familial advanced sleep-phase syndrome: A short-period circadian rhythm variant in humans". Tabiat tibbiyoti. 5 (9): 1062–5. doi:10.1038/12502. PMID 10470086. S2CID 14809619.
- ^ Johnson C (2004). Chronobiology: Biological Timekeeping. Sunderland, Massachusetts, USA: Sinauer Associates, Inc. pp. 67–105.
- ^ Sharma VK (November 2003). "Adaptive significance of circadian clocks". Xalqaro xronobiologiya. 20 (6): 901–19. doi:10.1081/CBI-120026099. PMID 14680135. S2CID 10899279.
- ^ [birlamchi bo'lmagan manba kerak ] Sheeba V, Sharma VK, Chandrashekaran MK, Joshi A (September 1999). "Persistence of eclosion rhythm in Drosophila melanogaster after 600 generations in an aperiodic environment". Naturwissenschaften vafot etdi. 86 (9): 448–9. Bibcode:1999NW.....86..448S. doi:10.1007/s001140050651. PMID 10501695. S2CID 13401297.
- ^ [birlamchi bo'lmagan manba kerak ] Guyomarc'h C, Lumineau S, Richard JP (May 1998). "Circadian rhythm of activity in Japanese quail in constant darkness: variability of clarity and possibility of selection". Xalqaro xronobiologiya. 15 (3): 219–30. doi:10.3109/07420529808998685. PMID 9653576.
- ^ [birlamchi bo'lmagan manba kerak ] Zivkovic BD, Underwood H, Steele CT, Edmonds K (October 1999). "Formal properties of the circadian and photoperiodic systems of Japanese quail: phase response curve and effects of T-cycles". Biologik ritmlar jurnali. 14 (5): 378–90. doi:10.1177/074873099129000786. PMID 10511005. S2CID 13390422.
- ^ Mori T, Johnson CH (April 2001). "Independence of circadian timing from cell division in cyanobacteria". Bakteriologiya jurnali. 183 (8): 2439–44. doi:10.1128/JB.183.8.2439-2444.2001. PMC 95159. PMID 11274102.
- ^ Hut RA, Beersma DG (July 2011). "Evolution of time-keeping mechanisms: early emergence and adaptation to photoperiod". London Qirollik Jamiyatining falsafiy operatsiyalari. B seriyasi, Biologiya fanlari. 366 (1574): 2141–54. doi:10.1098/rstb.2010.0409. PMC 3130368. PMID 21690131.
- ^ Dubowy C, Sehgal A (April 2017). "Drosophila melanogaster". Genetika. 205 (4): 1373–1397. doi:10.1534/genetics.115.185157. PMC 5378101. PMID 28360128.
- ^ [ishonchsiz tibbiy manbami? ] Nagoshi E, Saini C, Bauer C, Laroche T, Naef F, Schibler U (November 2004). "Circadian gene expression in individual fibroblasts: cell-autonomous and self-sustained oscillators pass time to daughter cells". Hujayra. 119 (5): 693–705. doi:10.1016/j.cell.2004.11.015. PMID 15550250. S2CID 15633902.
- ^ [birlamchi bo'lmagan manba kerak ] Michel S, Geusz ME, Zaritsky JJ, Block GD (January 1993). "Circadian rhythm in membrane conductance expressed in isolated neurons". Ilm-fan. 259 (5092): 239–41. Bibcode:1993Sci...259..239M. doi:10.1126/science.8421785. PMID 8421785.
- ^ Refinetti R (January 2010). "The circadian rhythm of body temperature". Bioscience-dagi chegara. 15 (3): 564–94. doi:10.2741/3634. PMID 20036834. S2CID 36170900.
- ^ Scheer FA, Morris CJ, Shea SA (March 2013). "The internal circadian clock increases hunger and appetite in the evening independent of food intake and other behaviors". Semirib ketish. 21 (3): 421–3. doi:10.1002/oby.20351. PMC 3655529. PMID 23456944.
- ^ [ishonchsiz tibbiy manbami? ] Zivkovic BC (2007-07-25). "Clock Tutorial #16: Photoperiodism – Models and Experimental Approaches (original work from 2005-08-13)". Soat atrofida blog. ScienceBlogs. Arxivlandi asl nusxasi 2008-01-01 kuni. Olingan 2007-12-09.
- ^ [birlamchi bo'lmagan manba kerak ] Turek FW, Joshu C, Kohsaka A, Lin E, Ivanova G, McDearmon E, et al. (2005 yil may). "Obesity and metabolic syndrome in circadian Clock mutant mice". Ilm-fan. 308 (5724): 1043–5. Bibcode:2005Sci...308.1043T. doi:10.1126/science.1108750. PMC 3764501. PMID 15845877.
- ^ Delezie J, Dumont S, Dardente H, Oudart H, Gréchez-Cassiau A, Klosen P, et al. (Avgust 2012). "The nuclear receptor REV-ERBα is required for the daily balance of carbohydrate and lipid metabolism". FASEB jurnali. 26 (8): 3321–35. doi:10.1096/fj.12-208751. PMID 22562834. S2CID 31204290.
- ^ [birlamchi bo'lmagan manba kerak ] Delezie J, Dumont S, Dardente H, Oudart H, Gréchez-Cassiau A, Klosen P, et al. (Avgust 2012). "The nuclear receptor REV-ERBα is required for the daily balance of carbohydrate and lipid metabolism". FASEB jurnali. 26 (8): 3321–35. doi:10.1096/fj.12-208751. PMID 22562834. S2CID 31204290.
- ^ [birlamchi bo'lmagan manba kerak ] Scott EM, Carter AM, Grant PJ (April 2008). "Association between polymorphisms in the Clock gene, obesity and the metabolic syndrome in man". Xalqaro semirish jurnali. 32 (4): 658–62. doi:10.1038/sj.ijo.0803778. PMID 18071340.
- ^ [ishonchsiz tibbiy manbami? ] Shneerson JM, Ohayon MM, Carskadon MA (2007). "Circadian rhythms". Rapid eye movement (REM) sleep. Armaniston tibbiyot tarmog'i. Olingan 2007-09-19.
- ^ "The Rhythms of Life: The Biological Clocks That Control the Daily Lives of Every Living Thing" Russell Foster & Leon Kreitzman, Publisher: Profile Books Ltd.
- ^ [ishonchsiz tibbiy manbami? ] Regestein QR, Pavlova M (September 1995). "Treatment of delayed sleep phase syndrome". Umumiy kasalxona psixiatriyasi. 17 (5): 335–45. doi:10.1016/0163-8343(95)00062-V. PMID 8522148.
- ^ [ishonchsiz tibbiy manbami? ] Howell E (14 December 2012). "Space Station to Get New Insomnia-Fighting Light Bulbs". Olingan 2012-12-17.
- ^ [birlamchi bo'lmagan manba kerak ] Spilde I (December 2005). "Reinsdyr uten døgnrytme" (Norvegiya Bokmal tilida). forskning.no. Arxivlandi asl nusxasi 2007-12-03 kunlari. Olingan 2007-11-24.
...så det ikke ut til at reinen hadde noen døgnrytme om sommeren. Svalbardreinen hadde det heller ikke om vinteren.
- ^ Folk GE, Thrift DL, Zimmerman MB, Reimann P (2006-12-01). "Mammalian activity – rest rhythms in Arctic continuous daylight". Biological Rhythm Research. 37 (6): 455–469. doi:10.1080/09291010600738551. S2CID 84625255.
Would local animals maintained under natural continuous daylight demonstrate the Aschoff effect described in previously published laboratory experiments using continuous light, in which rats' circadian activity patterns changed systematically to a longer period, expressing a 26-hour day of activity and rest?
- ^ [birlamchi bo'lmagan manba kerak ] Merlin C, Gegear RJ, Reppert SM (September 2009). "Antennal circadian clocks coordinate sun compass orientation in migratory monarch butterflies". Ilm-fan. 325 (5948): 1700–4. Bibcode:2009Sci...325.1700M. doi:10.1126/science.1176221. PMC 2754321. PMID 19779201.
- ^ [birlamchi bo'lmagan manba kerak ] Kyriacou CP (September 2009). "Physiology. Unraveling traveling". Ilm-fan. 325 (5948): 1629–30. doi:10.1126/science.1178935. PMID 19779177. S2CID 206522416.
- ^ Silvegren G, Löfstedt C, Qi Rosén W (March 2005). "Circadian mating activity and effect of pheromone pre-exposure on pheromone response rhythms in the moth Spodoptera littoralis". Hasharotlar fiziologiyasi jurnali. 51 (3): 277–86. doi:10.1016/j.jinsphys.2004.11.013. PMID 15749110.
- ^ a b Webb AA (June 2003). "The physiology of circadian rhythms in plants". Yangi fitolog. 160 (2): 281–303. doi:10.1046/j.1469-8137.2003.00895.x. JSTOR 1514280. S2CID 15688409.
- ^ a b v McClung CR (April 2006). "Plant circadian rhythms". O'simlik hujayrasi. 18 (4): 792–803. doi:10.1105/tpc.106.040980. PMC 1425852. PMID 16595397.
- ^ Mizoguchi T, Wright L, Fujiwara S, Cremer F, Lee K, Onouchi H, et al. (2005 yil avgust). "Distinct roles of GIGANTEA in promoting flowering and regulating circadian rhythms in Arabidopsis". O'simlik hujayrasi. 17 (8): 2255–70. doi:10.1105/tpc.105.033464. PMC 1182487. PMID 16006578.
- ^ Kolmos E, Davis SJ (September 2007). "ELF4 as a Central Gene in the Circadian Clock". O'simlik signalizatsiyasi va o'zini tutishi. 2 (5): 370–2. doi:10.4161/psb.2.5.4463. PMC 2634215. PMID 19704602.
- ^ Pokhilko A, Fernández AP, Edwards KD, Southern MM, Halliday KJ, Millar AJ (March 2012). "The clock gene circuit in Arabidopsis includes a repressilator with additional feedback loops". Molekulyar tizimlar biologiyasi. 8: 574. doi:10.1038/msb.2012.6. PMC 3321525. PMID 22395476.
- ^ Ma Y, Gil S, Grasser KD, Mas P (April 2018). "Targeted Recruitment of the Basal Transcriptional Machinery by LNK Clock Components Controls the Circadian Rhythms of Nascent RNAs in Arabidopsis". O'simlik hujayrasi. 30 (4): 907–924. doi:10.1105/tpc.18.00052. PMC 5973845. PMID 29618629.
- ^ a b v d e f g h men j Dodd AN, Salathia N, Hall A, Kévei E, Tóth R, Nagy F, Webb AA (2005). "Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage". Ilm-fan. 309 (5734): 630–633. Bibcode:2005Sci...309..630D. doi:10.1126 / science.1115581. PMID 16040710. S2CID 25739247.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ a b v d e Dodd AN, Belbin FE, Frank A, Webb AA (2015). "Interactions between circadian clocks and photosynthesis for the temporal and spatial coordination of metabolism". O'simlikshunoslik chegaralari. 6: 245. doi:10.3389/fpls.2015.00245. PMC 4391236. PMID 25914715.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Webb AA, Seki M, Satake A, Caldana C (2019). "Continuous dynamic adjustment of the plant circadian oscillator". Tabiat aloqalari. 10 (1): 550. Bibcode:2019NatCo..10..550W. doi:10.1038/s41467-019-08398-5. PMC 6358598. PMID 30710080.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ a b Haydon MJ, Mielczarek O, Robertson FC, Hubbard KE, Webb AA (2013). "Photosynthetic entrainment of the Arabidopsis thaliana circadian clock". Tabiat. 502 (7473): 689–692. Bibcode:2013Natur.502..689H. doi:10.1038/nature12603. PMC 3827739. PMID 24153186.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Farré EM, Kay SA (2007). "PRR7 protein levels are regulated by light and the circadian clock in Arabidopsis". O'simlik jurnali. 52 (3): 548–560. doi:10.1111/j.1365-313X.2007.03258.x. PMID 17877705.
- ^ Veleri S, Wülbeck C (May 2004). "Unique self-sustaining circadian oscillators within the brain of Drosophila melanogaster". Xalqaro xronobiologiya. 21 (3): 329–42. doi:10.1081/CBI-120038597. PMID 15332440. S2CID 15099796.
- ^ Yoshii T, Hermann-Luibl C, Helfrich-Förster C (2015). "Circadian light-input pathways in Drosophila". Communicative & Integrative Biology. 9 (1): e1102805. doi:10.1080/19420889.2015.1102805. PMC 4802797. PMID 27066180.
- ^ Boothroyd CE, Young MW (2008). "The in(put)s and out(put)s of the Drosophila circadian clock". Nyu-York Fanlar akademiyasining yilnomalari. 1129 (1): 350–7. Bibcode:2008NYASA1129..350B. doi:10.1196/annals.1417.006. PMID 18591494. S2CID 2639040.
- ^ Grima B, Lamouroux A, Chélot E, Papin C, Limbourg-Bouchon B, Rouyer F (November 2002). "The F-box protein slimb controls the levels of clock proteins period and timeless". Tabiat. 420 (6912): 178–82. Bibcode:2002Natur.420..178G. doi:10.1038/nature01122. PMID 12432393. S2CID 4428779.
- ^ Ko HW, Jiang J, Edery I (December 2002). "Role for Slimb in the degradation of Drosophila Period protein phosphorylated by Doubletime". Tabiat. 420 (6916): 673–8. Bibcode:2002Natur.420..673K. doi:10.1038/nature01272. PMID 12442174. S2CID 4414176.
- ^ Helfrich-Förster C (March 2005). "Neurobiology of the fruit fly's circadian clock". Genlar, miya va o'zini tutish. 4 (2): 65–76. doi:10.1111/j.1601-183X.2004.00092.x. PMID 15720403. S2CID 26099539.
- ^ Lalchhandama K (2017). "The path to the 2017 Nobel Prize in Physiology or Medicine". Ilmiy qarash. 3 (Suppl): 1–13.
- ^ a b v Leloup JC, Goldbeter A (February 1998). "A model for circadian rhythms in Drosophila incorporating the formation of a complex between the PER and TIM proteins". Biologik ritmlar jurnali. 13 (1): 70–87. doi:10.1177/074873098128999934. PMID 9486845. S2CID 17944849.
- ^ Goldbeter A (September 1995). "A model for circadian oscillations in the Drosophila period protein (PER)". Ish yuritish. Biologiya fanlari. 261 (1362): 319–24. Bibcode:1995RSPSB.261..319G. doi:10.1098/rspb.1995.0153. PMID 8587874. S2CID 7024361.
- ^ a b Goldbeter A (November 2002). "Computational approaches to cellular rhythms". Tabiat. 420 (6912): 238–45. Bibcode:2002Natur.420..238G. doi:10.1038/nature01259. PMID 12432409. S2CID 452149.
- ^ "Biological Clock in Mammals". BioInteractive. Xovard Xyuz tibbiyot instituti. Olingan 5 may 2015.
- ^ Welsh DK, Takahashi JS, Kay SA (March 2010). "Suprachiasmatic nucleus: cell autonomy and network properties". Fiziologiyaning yillik sharhi. 72: 551–77. doi:10.1146/annurev-physiol-021909-135919. PMC 3758475. PMID 20148688.
- ^ Pfeffer M, Korf HW, Wicht H (March 2018). "Synchronizing effects of melatonin on diurnal and circadian rhythms". Umumiy va qiyosiy endokrinologiya. 258: 215–221. doi:10.1016/j.ygcen.2017.05.013. PMID 28533170.
- ^ Kalpesh J. "Wellness With Artificial Light". Olingan 11 yanvar 2016.
- ^ [ishonchsiz tibbiy manbami? ] Scheer FA, Wright KP, Kronauer RE, Czeisler CA (August 2007). "Plasticity of the intrinsic period of the human circadian timing system". PLOS ONE. 2 (8): e721. Bibcode:2007PLoSO...2..721S. doi:10.1371/journal.pone.0000721. PMC 1934931. PMID 17684566.
- ^ [ishonchsiz tibbiy manbami? ] Duffy JF, Wright KP (August 2005). "Entrainment of the human circadian system by light". Biologik ritmlar jurnali. 20 (4): 326–38. doi:10.1177/0748730405277983. PMID 16077152. S2CID 20140030.
- ^ Khalsa SB, Jewett ME, Cajochen C, Czeisler CA (June 2003). "A phase response curve to single bright light pulses in human subjects". Fiziologiya jurnali. 549 (Pt 3): 945–52. doi:10.1113/jphysiol.2003.040477. PMC 2342968. PMID 12717008.
- ^ Cromie W (1999-07-15). "Inson biologik soati bir soat orqaga qaytdi". Garvard gazetasi. Olingan 2015-07-04.
- ^ a b Duffy JF, Cain SW, Chang AM, Phillips AJ, Münch MY, Gronfier C, et al. (Sentyabr 2011). "Sex difference in the near-24-hour intrinsic period of the human circadian timing system". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 108 Suppl 3 (Supplement_3): 15602–8. Bibcode:2011PNAS..10815602D. doi:10.1073/pnas.1010666108. PMC 3176605. PMID 21536890.
- ^ a b v Benloucif S, Guico MJ, Reid KJ, Wolfe LF, L'hermite-Balériaux M, Zee PC (April 2005). "Stability of melatonin and temperature as circadian phase markers and their relation to sleep times in humans". Biologik ritmlar jurnali. 20 (2): 178–88. doi:10.1177/0748730404273983. PMID 15834114. S2CID 36360463.
- ^ Adam EK, Quinn ME, Tavernier R, McQuillan MT, Dahlke KA, Gilbert KE (September 2017). "Diurnal cortisol slopes and mental and physical health outcomes: A systematic review and meta-analysis". Psixonuroendokrinologiya. 83: 25–41. doi:10.1016/j.psyneuen.2017.05.018. PMC 5568897. PMID 28578301.
- ^ Baehr EK, Revelle W, Eastman CI (June 2000). "Individual differences in the phase and amplitude of the human circadian temperature rhythm: with an emphasis on morningness-eveningness". Uyqu tadqiqotlari jurnali. 9 (2): 117–27. doi:10.1046/j.1365-2869.2000.00196.x. PMID 10849238. S2CID 6104127.
- ^ "NHLBI Workshop: "Circadian Clock at the Interface of Lung Health and Disease" 28-29 April 2014 Executive Summary". Milliy yurak, o'pka va qon instituti. Sentyabr 2014. Arxivlangan asl nusxasi 2014-10-04 kunlari. Olingan 20 sentyabr 2014.
- ^ Degaute JP, van de Borne P, Linkowski P, Van Cauter E (August 1991). "Quantitative analysis of the 24-hour blood pressure and heart rate patterns in young men". Gipertenziya. 18 (2): 199–210. doi:10.1161/01.hyp.18.2.199. PMID 1885228.
- ^ Quartel L (2014). "The effect of the circadian rhythm of body temperature on A-level exam performance". Undergraduate Journal of Psychology. 27 (1).
- ^ Mohawk JA, Green CB, Takahashi JS (July 14, 2013). "Central and peripheral circadian clocks in mammals". Nevrologiyani yillik sharhi. 35: 445–62. doi:10.1146/annurev-neuro-060909-153128. PMC 3710582. PMID 22483041.
- ^ Id.
- ^ Pendergast JS, Niswender KD, Yamazaki S (January 11, 2012). "Tissue-specific function of Period3 in circadian rhythmicity". PLOS ONE. 7 (1): e30254. Bibcode:2012PLoSO...730254P. doi:10.1371/journal.pone.0030254. PMC 3256228. PMID 22253927.
- ^ Singh M (10 Oct 2013). "Our Skin's Sense Of Time Helps Protect Against UV Damage". Milliy radio. Olingan 19 fevral 2019.
- ^ Abraham U, Granada AE, Westermark PO, Heine M, Kramer A, Herzel H (November 2010). "Coupling governs entrainment range of circadian clocks". Molekulyar tizimlar biologiyasi. 6: 438. doi:10.1038/msb.2010.92. PMC 3010105. PMID 21119632.
- ^ Cao Q, Gery S, Dashti A, Yin D, Zhou Y, Gu J, Koeffler HP (October 2009). "A role for the clock gene per1 in prostate cancer". Saraton kasalligini o'rganish. 69 (19): 7619–25. doi:10.1158/0008-5472.CAN-08-4199. PMC 2756309. PMID 19752089.
- ^ Kawara S, Mydlarski R, Mamelak AJ, Freed I, Wang B, Watanabe H, et al. (2002 yil dekabr). "Low-dose ultraviolet B rays alter the mRNA expression of the circadian clock genes in cultured human keratinocytes". Tergov dermatologiyasi jurnali. 119 (6): 1220–3. doi:10.1046/j.1523-1747.2002.19619.x. PMID 12485420.
- ^ Damiola F, Le Minh N, Preitner N, Kornmann B, Fleury-Olela F, Schibler U (December 2000). "Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus". Genlar va rivojlanish. 14 (23): 2950–61. doi:10.1101/gad.183500. PMC 317100. PMID 11114885.
- ^ Duffy JF, Czeisler CA (June 2009). "Effect of Light on Human Circadian Physiology". Uyqu tibbiyot klinikalari. 4 (2): 165–177. doi:10.1016/j.jsmc.2009.01.004. PMC 2717723. PMID 20161220.
- ^ Czeisler CA, Duffy JF, Shanahan TL, Brown EN, Mitchell JF, Rimmer DW, et al. (Iyun 1999). "Stability, precision, and near-24-hour period of the human circadian pacemaker". Ilm-fan. 284 (5423): 2177–81. doi:10.1126/science.284.5423.2177. PMID 10381883.
- ^ Aldrich MS (1999). Uyqu uchun dori. Nyu-York: Oksford universiteti matbuoti. ISBN 978-0-19-512957-1.
- ^ Wyatt JK, Ritz-De Cecco A, Czeisler CA, Dijk DJ (October 1999). "Circadian temperature and melatonin rhythms, sleep, and neurobehavioral function in humans living on a 20-h day". Amerika fiziologiya jurnali. 277 (4 Pt 2): R1152-63. doi:10.1152/ajpregu.1999.277.4.R1152. PMID 10516257. S2CID 4474347.
- ^ Wright KP, Hull JT, Czeisler CA (December 2002). "Relationship between alertness, performance, and body temperature in humans". Amerika fiziologiya jurnali. Normativ, integral va qiyosiy fiziologiya. 283 (6): R1370-7. CiteSeerX 10.1.1.1030.9291. doi:10.1152/ajpregu.00205.2002. PMID 12388468.
- ^ Zhou X, Ferguson SA, Matthews RW, Sargent C, Darwent D, Kennaway DJ, Roach GD (July 2011). "Sleep, wake and phase dependent changes in neurobehavioral function under forced desynchrony". Uyqu. 34 (7): 931–41. doi:10.5665/SLEEP.1130. PMC 3119835. PMID 21731143.
- ^ Kosmadopoulos A, Sargent C, Darwent D, Zhou X, Dawson D, Roach GD (December 2014). "The effects of a split sleep-wake schedule on neurobehavioural performance and predictions of performance under conditions of forced desynchrony". Xalqaro xronobiologiya. 31 (10): 1209–17. doi:10.3109/07420528.2014.957763. PMID 25222348. S2CID 11643058.
- ^ Grote L, Mayer J, Penzel T, Cassel W, Krzyzanek E, Peter JH, von Wichert P (1994). "Nocturnal hypertension and cardiovascular risk: consequences for diagnosis and treatment". Kardiyovasküler farmakologiya jurnali. 24 Suppl 2: S26-38. PMID 7898092.
- ^ Hershner SD, Chervin RD (2014-06-23). "Kollej o'quvchilari o'rtasida uyquning sabablari va oqibatlari". Uyquning tabiati va fani. 6: 73–84. doi:10.2147 / NSS.S62907. PMC 4075951. PMID 25018659.
- ^ Milner CE, Cote KA (June 2009). "Benefits of napping in healthy adults: impact of nap length, time of day, age, and experience with napping". Uyqu tadqiqotlari jurnali. 18 (2): 272–81. doi:10.1111/j.1365-2869.2008.00718.x. PMID 19645971. S2CID 22815227.
- ^ Lovato N, Lack L (2010). The effects of napping on cognitive functioning. Miya tadqiqotida taraqqiyot. 185. 155–166 betlar. doi:10.1016/B978-0-444-53702-7.00009-9. ISBN 978-0-444-53702-7. PMID 21075238.
- ^ a b v Zelinski EL, Deibel SH, McDonald RJ (March 2014). "The trouble with circadian clock dysfunction: multiple deleterious effects on the brain and body". Neyrologiya va biobehavioral sharhlar. 40 (40): 80–101. doi:10.1016/j.neubiorev.2014.01.007. PMID 24468109. S2CID 6809964.
- ^ Sinert T, Peacock PR (10 May 2006). "Renal Failure, Acute". eMedicine from WebMD. Olingan 2008-08-03.
- ^ Maung SC, El Sara A, Chapman C, Cohen D, Cukor D (May 2016). "Sleep disorders and chronic kidney disease". Jahon nefrologiya jurnali. 5 (3): 224–32. doi:10.5527/wjn.v5.i3.224. PMC 4848147. PMID 27152260.
- ^ Nakano S, Uchida K, Kigoshi T, Azukizawa S, Iwasaki R, Kaneko M, Morimoto S (August 1991). "Circadian rhythm of blood pressure in normotensive NIDDM subjects. Its relationship to microvascular complications". Qandli diabetga yordam. 14 (8): 707–11. doi:10.2337/diacare.14.8.707. PMID 1954805. S2CID 12489921.
- ^ Figueiro MG, Rea MS, Bullough JD (August 2006). "Does architectural lighting contribute to breast cancer?". Journal of Carcinogenesis. 5: 20. doi:10.1186/1477-3163-5-20. PMC 1557490. PMID 16901343.
- ^ Rea MS, Figueiro M, Bullough J (May 2002). "Circadian photobiology: an emerging framework for lighting practice and research". Yoritishni tadqiq qilish va texnologiyasi. 34 (3): 177–187. doi:10.1191/1365782802lt057oa. S2CID 109776194.
- ^ Walmsley L, Hanna L, Mouland J, Martial F, West A, Smedley AR, et al. (2015 yil aprel). "Colour as a signal for entraining the mammalian circadian clock". PLOS biologiyasi. 13 (4): e1002127. doi:10.1371/journal.pbio.1002127. PMC 4401556. PMID 25884537.
- ^ Johnston JD (June 2014). "Physiological responses to food intake throughout the day". Oziqlanish bo'yicha tadqiqotlar. 27 (1): 107–18. doi:10.1017/S0954422414000055. PMC 4078443. PMID 24666537.
- ^ Delezie J, Challet E (December 2011). "Interactions between metabolism and circadian clocks: reciprocal disturbances". Nyu-York Fanlar akademiyasining yilnomalari. 1243 (1): 30–46. Bibcode:2011NYASA1243...30D. doi:10.1111/j.1749-6632.2011.06246.x. PMID 22211891. S2CID 43621902.
- ^ [1][o'lik havola ]
- ^ Circadian Rhythm Disruption and Flying. FAA at https://www.faa.gov/pilots/safety/pilotsafetybrochures/media/Circadian_Rhythm.pdf
- ^ "Jet lag disorder - Symptoms and causes". Mayo klinikasi. Olingan 2019-02-01.
- ^ Zhu L, Zee PC (November 2012). "Circadian rhythm sleep disorders". Nevrologik klinikalar. 30 (4): 1167–91. doi:10.1016/j.ncl.2012.08.011. PMC 3523094. PMID 23099133.
- ^ Oritz-Tuldela E, Martinez-Nicolas A, Diaz-Mardomingo C, Garcia-Herranz S, Pereda-Perez I, Valencia A, Peraita H, Venero C, Madrid J, Rol M. 2014. The Characterization of Biological Rhythms in Mild Cognitive Impairment. BioMed Research International.
- ^ Hardt R (1970-01-01). "The Dangers of LED-Blue light-The Suppression of Melatonin-Resulting in-Insomnia-And Cancers | Robert Hardt". Academia.edu. Olingan 2016-12-24.
- ^ Bedrosian TA, Nelson RJ (January 2017). "Timing of light exposure affects mood and brain circuits". Tarjima psixiatriyasi. 7 (1): e1017. doi:10.1038/tp.2016.262. PMC 5299389. PMID 28140399.
- ^ a b v Logan RW, Williams WP, McClung CA (June 2014). "Circadian rhythms and addiction: mechanistic insights and future directions". Xulq-atvor nevrologiyasi. 128 (3): 387–412. doi:10.1037/a0036268. PMC 4041815. PMID 24731209.
- ^ Prosser RA, Glass JD (June 2015). "Assessing ethanol's actions in the suprachiasmatic circadian clock using in vivo and in vitro approaches". Spirtli ichimliklar. 49 (4): 321–339. doi:10.1016/j.alcohol.2014.07.016. PMC 4402095. PMID 25457753.
- ^ Cha AE (October 2, 2017). "Nobel in physiology, medicine awarded to three Americans for discovery of 'clock genes'". Washington Post. Olingan 2 oktyabr, 2017.
- ^ "The 2017 Nobel Prize in Physiology or Medicine – Press Release". Nobel jamg'armasi. 2017 yil 2-oktabr. Olingan 2 oktyabr, 2017.
Qo'shimcha o'qish
- Aschoff J, ed. (1965). Circadian Clocks. Amsterdam: North Holland Press.
- Avivi A, Albrecht U, Oster H, Joel A, Beiles A, Nevo E (November 2001). "Biological clock in total darkness: the Clock/MOP3 circadian system of the blind subterranean mole rat". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 98 (24): 13751–6. Bibcode:2001PNAS...9813751A. doi:10.1073/pnas.181484498. PMC 61113. PMID 11707566.
- Avivi A, Oster H, Joel A, Beiles A, Albrecht U, Nevo E (September 2002). "Circadian genes in a blind subterranean mammal II: conservation and uniqueness of the three Period homologs in the blind subterranean mole rat, Spalax ehrenbergi superspecies". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 99 (18): 11718–23. Bibcode:2002PNAS...9911718A. doi:10.1073/pnas.182423299. PMC 129335. PMID 12193657.
- Li D, Ma S, Guo D, Cheng T, Li H, Tian Y, et al. (Oktyabr 2016). "Environmental Circadian Disruption Worsens Neurologic Impairment and Inhibits Hippocampal Neurogenesis in Adult Rats After Traumatic Brain Injury". Uyali va molekulyar neyrobiologiya. 36 (7): 1045–55. doi:10.1007/s10571-015-0295-2. PMC 4967018. PMID 26886755.
- Ditty JL, Williams SB, Golden SS (2003). "A cyanobacterial circadian timing mechanism". Genetika fanining yillik sharhi. 37: 513–43. doi:10.1146/annurev.genet.37.110801.142716. PMID 14616072. S2CID 36703896.
- Dunlap JC, Loros J, DeCoursey PJ (2003). Chronobiology: Biological Timekeeping. Sanderlend: Sinayer.
- Dvornyk V, Vinogradova O, Nevo E (March 2003). "Origin and evolution of circadian clock genes in prokaryotes". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 100 (5): 2495–500. Bibcode:2003PNAS..100.2495D. doi:10.1073/pnas.0130099100. PMC 151369. PMID 12604787.
- Koukkari WL, Sothern RB (2006). Introducing Biological Rhythms. Nyu-York: Springer.
- Martino T, Arab S, Straume M, Belsham DD, Tata N, Cai F, et al. (2004 yil aprel). "Day/night rhythms in gene expression of the normal murine heart". Molekulyar tibbiyot jurnali. 82 (4): 256–64. doi:10.1007/s00109-003-0520-1. PMID 14985853. S2CID 871822.
- Refinetti R (2006). Sirkad fiziologiyasi (2-nashr). Boka Raton: CRC Press.
- Takahashi JS, Zatz M (September 1982). "Regulation of circadian rhythmicity". Ilm-fan. 217 (4565): 1104–11. Bibcode:1982Sci...217.1104T. doi:10.1126/science.6287576. PMID 6287576.
- Tomita J, Nakajima M, Kondo T, Iwasaki H (January 2005). "No transcription-translation feedback in circadian rhythm of KaiC phosphorylation". Ilm-fan. 307 (5707): 251–4. Bibcode:2005Sci...307..251T. doi:10.1126/science.1102540. PMID 15550625. S2CID 9447128.
- Moore-Ede MC, Sulzman FM, Fuller CA (1982). The Clocks that Time Us: Physiology of the Circadian Timing System. Kembrij, Massachusets: Garvard universiteti matbuoti. ISBN 978-0-674-13581-9.
Tashqi havolalar
| Ichki ritmlar |  | |
|---|---|---|
| Tashqi tsikllar | ||
| Maydonlar | ||
| Shuningdek qarang |
| |