Garudimimus - Garudimimus
| Garudimimus | |
|---|---|
 | |
| Ichida o'rnatilgan holotip Mo'g'uliston tabiiy tarix muzeyi (noto'g'ri kaudal vertebraga e'tibor bering; holotipda faqat to'rttasi qayd etilgan) | |
| Ilmiy tasnif | |
| Qirollik: | Animalia |
| Filum: | Chordata |
| Klade: | Dinozavrlar |
| Klade: | Saurischia |
| Klade: | Theropoda |
| Klade: | Maniraptoriformes |
| Klade: | †Ornitomimozauriya |
| Tur: | †Garudimimus Barsbold, 1981 |
| Tur turlari | |
| †Garudimimus qisqartiradi Barsbold, 1981 yil | |
Garudimimus (ma'nosi "Garuda taqlid ") - bu ornitomimozavr yashagan Osiyo davomida Kechki bo'r. Ushbu tur 1981 yilda Sovet-Mo'g'ul ekspeditsiyasi tomonidan topilgan bitta namunadan ma'lum Bayan Shireh shakllanishi va rasmiy ravishda o'sha yili tasvirlangan Rinchen Barsbold; yagona tur Garudimimus qisqartiradi. Ning anatomik xususiyatlari haqida bir nechta talqinlar Garudimimus namunaning orqa ekspertizalarida o'tkazilgan, ammo ularning aksariyati uni 2005 yilda har tomonlama qayta ko'rib chiqishda tanqid qilingan. Keng ta'riflanmagan ornitomimozavr tipik joyda saqlanib qolgan Garudimimus jinsning qo'shimcha namunalarini ifodalashi mumkin.
Faqatgina ma'lum bo'lgan namunasi Garudimimus uzunligi 3,5 m (11 ft) bo'lgan va vazni 98 kg (216 lb) bo'lgan o'rtacha hayvon edi. Bu bazal va kelib chiqadigan xususiyatlar aralashgan ornitomimozavr edi; ibtidoiy ornitomimozavrlardan farqli o'laroq, ikkala yuqori va pastki jag'lar edi tishsiz, ko'pincha o'ziga xos xususiyati ko'proq olingan ornitomimidlar. Garudimimus orqa oyoqlari nisbatan qisqa va baquvvat, oyoqlari mustahkam va qisqargan ilia. Oyoqning to'rt barmog'i bor edi, birinchisi juda qisqargan, ornitomimidlar esa uch barmoqli bo'lib, birinchi barmoq yo'qolgan. Tishsiz bosh suyagi boshqa avlodlarga qaraganda dumaloq uchi bilan tugaydigan juda to'g'ri jag'larga ega. Ilgari ushbu ibtidoiy ornitomimozaurian a ko'z yoshi Boshsuyagi tepasida, ko'z teshigi oldida "shox". Biroq, yagona namunani qayta tavsiflash shuni ko'rsatdiki, bu struktura shunchaki buzilgan chap edi prefrontal suyak. Yana bir dastlabki talqin metatarsus bilan tiklangan edi arktometatarsalian holat.
1981 yilda birinchi tavsif bilan, Garudimimus o'z oilasida ibtidoiy ornitomimozauriya sifatida aniqlandi. Biroq, yangi namunalarining tavsifi bilan Deinocheirus 2014 yilda, bu ikkinchisi bo'lganligi aniqlandi opa takson ning Garudimimusichida guruhlash Deinocheiridae - yugurish yoki epchil harakatlarga moslashtirilmagan ornitomimozavrlar. Ammo bu joylashtirish ham mumkin emasligi aniqlandi. Tos kamari va orqa oyoqlari Garudimimus oyoq mushaklarining tez ishlaydigan ornitomimidlar singari yaxshi rivojlanmaganligini ko'rsating kursor imkoniyatlar. Ornitomimosauriyaning boshqa a'zolari singari, Garudimimus shoxli tumshug'i bilan kompensatsiya qilingan, ozaygan tishlash kuchiga ega bo'lgan yeyuvchi / o'txo'r edi.
Kashfiyot tarixi

1981 yilda Sovet-Mo'g'ul ekspeditsiyasi paytida Gobi sahrosi, nisbatan kichik teropod skeleti Bayshi Tsavdagi guruh tomonidan Kechki bo'r Bayan Shireh shakllanishi, Janubi-sharqiy Mo'g'uliston. Qoldiqlar quyidagicha kataloglanadi MPC-D 100/13 (dastlab GIN 100/13) va juda to'liq va bo'g'inli skeletni ifodalaydi. Xuddi shu yili ushbu namunani mo'g'ullar rasmiy va qisqacha ta'rifladilar paleontolog Rinchen Barsbold sifatida holotip yangi nomlangan tur turlari Garudimimus qisqartiradi. The umumiy ism, Garudimimus, havolani birlashtiradi Garuda mo'g'uldan kelgan afsonaviy qanotli mavjudotlar Buddist mifologiyasi va a Lotin mimus (taqlid ma'nosini anglatadi). The aniq ism lotin tilidan olingan brevis (qisqa ma'nosini anglatadi) va pes (oyoq ma'nosini anglatadi), kalta degani metatarsus. Barsbold aniqlandi Garudimimus ornitomimosaurian taksoni sifatida, lekin uning ornitomimidlarga qaraganda ancha ibtidoiy ekanligini ta'kidlab, unga o'z oilasini, ya'ni Garudimimidae.[1]
Barsbold 1983 yilda, so'ngra 1990 yilda holotip namunasining qo'shimcha qoldiqlarini tasvirlab berdi Halszka Osmolska.[2][3] Tomonidan erta talqin qilingan Filipp J. Kurri va Deyl Alan Rassel MPC-D 100/13 metatarsusini kim deb atagan Oviraptor sp. va uni an bilan rekonstruksiya qildi arktometatarsal holat.[4] Tomas R. Xolts Ushbu talqinni kuzatib, metatarsus arktometatarsalian bo'lishi mumkin edi va saqlanib qolgani uchun shunchaki disartikulyatsiya qilingan edi.[5] Arkometatarsalian holatini qo'llab-quvvatlash, Currie va Devid A. Ebert ning bu qismini da'vo qilgan Arxeornitomimus material tegishli edi Garudimimus taxmin qilingan arktometatarsalian holatiga asoslanib tarixiy I raqam va II, III va IV metatarsallarning nisbati. Ular metatarsallar ezilib, metatarsal III metatarsning ekstansor yuzasidan orqaga qarab o'rnatilishini ta'kidladilar.[6] 1994 yilda Xolts metatarsus o'rtasida ba'zi o'xshashliklarni taklif qildi Garudimimus va Chirostenotlar.[7] Xuddi shu yili Peres-Moreno hamkasblari bilan ibtidoiy ornitomimozavrlarni tasvirlab berdi Pelecanimimus va holotip namunasida tepalik borligini aniqladi. Ular shunga o'xshash xususiyatni da'vo qilishdi Garudimimus u burun shoxi bilan ifodalangan.[8]

Yoshitsugu Kobayashi va Barsbold 2005 yilda har tomonlama qayta ko'rib chiqishda holotipni kuzatgan bosh suyagi to'liq, 8 bachadon bo'yni umurtqalari (shu jumladan atlas va o'qi ), 9 orqa umurtqalari, 6 sakral vertebra va 4 kaudal vertebra, biroz qovurg'alar, ikkalasi ham ilia, pablar, femora, tibiae, fibulae va deyarli to'liq chap pes voyaga etmagan shaxsning. Ushbu qayta tahrirlash, oldingi orbitadagi shoxning aslida parchalangan chap prefrontal suyak ekanligini ko'rsatgan va metatarsusning zarar ko'rmaganligini tasdiqlagan ko'plab oldingi bayonotlarni rad etdi. taponomik buzilish va arktometatarsal bo'lmagan. Bundan tashqari, Kobayashi ilgari arkometatarsalian holatiga ega emasligi bilan bir qatorda metatarsal nisbatlar boshqacha ekanligini ko'rsatdi Arxeornitomimus.[9] Garchi Garudimimus faqat MPC-D 100/13 dan ma'lum, qo'shimcha namunalar potentsial ravishda yiriklar orasida bo'lishi mumkin suyak suyagi Bayshi Tsavdagi kamida beshta kishidan va Bayon Shireh qatlamidagi boshqa joylardan tasvirlanmagan bir nechta ornitomimozavrlardan tashkil topgan.[10][11][12]
Tavsif

Holotip namunasi kichik yoshdagi shaxs bo'lsa-da, uning uzunligi taxminan 98 kg (216 funt) og'irlikdagi 3,5 m (11 fut) atrofida bo'lgan deb taxmin qilinadi.[13][14] Xolotipning sub-kattalar holati kattalardagi kattaroq hajmni ko'rsatishi mumkin. Ushbu taksonning ayrim ajralib turadigan xususiyatlariga jag'ning artikulyatsiyasi postorbital barga qaraganda orqaga qarab joylashtirilgan, poydevoridagi teshiklar kiradi. supraoksipital yuqori proektsiyasi, oldingi dumaloq umurtqa pog'onasi nerv tizmalarining lateral yuzalaridagi juftlashgan tushkunliklar va III-1 va III-2 pedal falanjlari lateral yuzasining yuqori uchida chuqur yiv.[9] Bir nechta namunalarning tuklar taassurotlari asosida Ornitomimus, Garudimimus ehtimol shaggy rivojlangan, ratit - tuklar kabi.[15][16]
Boshsuyagi

Bosh suyagi aniqlanadigan kranial elementlar bilan nisbatan yaxshi saqlanib qolgan. Garchi chap tomoni o'ng tomonga qaraganda ko'proq zarar ko'rgan bo'lsa-da, avvalgisi noziklikni saqlaydi sklerotik halqalar. O'ng tomon kamroq shikastlanganligini hisobga olsak, tavsif shu tomonga ko'proq e'tibor qaratdi.[9] Dorsal nuqtai nazardan qaraganda, bosh suyagi kengligi 3,6 sm (36 mm) bo'lgan o'lchovni u yoqdan bu tomonga juda ezadi. Retroformatsiyaga uchraganda, bosh suyagi 4,6 sm (46 mm) kenglik bilan sezilarli darajada kengayadi.[17] Asl ezilgan bosh suyagida zamonaviy bo'lmagan fenestra ba'zi elementlarning siljishi tufayli tartibsiz shaklga ega. Ushbu fenestraning yuqori egriligi kam rivojlangan. Ning qirralari premaxilla bor tishsiz, ingichka va keskin rivojlangan, pastki lateral yuzalarida esa juda ko'p foramina topish mumkin. Ushbu premaxilla tashqi narial ochilishning katta qismi bilan chegaralanadi, ammo orqa tomoni bilan chegaralanmaydi. Yon tomondan qaralganda, prekaksillaning pastki chegarasi to'g'ri va maxilla bilan aloqa burun teshigining orqa uchiga orqaga qarab yotadi. Yon tomondan preaksilla-maxilla tikuv shakllantiradi a to'g'ri burchak yuqori jag'ning pastki chegarasiga. Maksillerning dorsal kengayishi jarayon (suyak kengaytmasi) ning oldingi chegarasidan orqada tugaydi antorbital fossa. The palatal ikkala premaxillaning tokchasida kesuvchi teshik va juft teshik bor. Asosiy tanasining lateral yuzasi maxilla boshqa ornitomimozavrlarda bo'lgani kabi qo'pol. Ham promaksillarar, ham maksillarar fenestralar kam rivojlangan. Antorbital fossa markazida lakrimal bilan maxilla bo'g'imlarining yuqori kengayishi. Yanal ko'rinishda, maxillaning pastki chegarasi ham to'g'ri. Olingan ornitomimidlarda bo'lgani kabi, pastki yuzada ko'plab teshiklar mavjud.[9] Ham prekaksillaning, ham tishlarning tishsiz holati, ularning yuzalarida ko'plab teshiklar paydo bo'lishi ramfoteka (tumshuq) hayotda mavjud edi.[17] The burun suyagi cho'zilgan va tashqi narial teshikning orqa chegarasiga shakl beradi. Frontal aloqa tomon torayib boradi va uning orqa uchi prefrontal va lakrimalning orqa uchlari orasida joylashgan. Ba'zi sirt teshiklari yuqori yuzada mavjud.[9]

The lakrimal suyak yarim T shaklida va oldingi uchi shikastlangan, chunki bu kengayish nazolakrimal kanal tomonidan joylashtirilgan. Yon tomonda va pastki kengaytmaning orqa uchida joylashgan sirt nazolakrimal kanal uchun orqa teshikka ega - bu ham "Gallimimus mongoliensis ". Lakrimalning ichki yuzasida katta uchburchak shaklidagi tushkunlik mavjud. Lakrimalning orqa jarayoni torayib boradi va prefrontaldagi tushkunlikka tushadi. The prefrontal lakrimal asosiy tanasining pastki yuzasiga birlashadigan oldingi jarayonga ega. Prefrontalning ochiq dorsal maydoni lakrimalnikiga qaraganda bir oz kamayadi. Old jarayon ikkiga bo'linadi va ichki shox lateralga qaraganda ancha katta. Orqa nuqtai nazardan, prefrontalning orbital qirrasi pastki qismga qalinlashadi va teshik etishmaydi.[9] Saqlanib qolganidek orbital balandligi 5,95 sm (59,5 mm) bo'lgan katta bo'shliq. Retro deformatsiyaga uchraganda, uning eni 6,1 sm (61 mm) dan biroz kattalashadi.[17] Garchi o'ng tomoni to'liqroq bo'lsa-da, bosh suyagining chap tomonida kamida 11 bo'g'inli skleral plitalar saqlanib qoladi. sklerotik halqa. Halqa radiusi ichkarida taxminan 2,86 sm (28,6 mm) va tashqarida 4,12 sm (41,2 mm) dir. Har bir plastinkaning kengligi taxminan 0,8 sm (8,0 mm). Oldingi uchi jugal umuman saqlanib qolmagan, ko'z yoshi va maxilla uchun tikuvlar yaxshi rivojlanmagan. Jugal oxirigacha uzayadi va lateral ko'rinishda jugal juda tekis va orqada juda qisqa jarayonga ega. Yanal sirt kvadratadugugalning oldingi jarayoni bilan bog'lanish uchun sayoz tushkunlikka ega, ammo u orbitaning orqa chetidan oldin joylashgan.[9]
The kvadratojugal katta va uchburchak shaklidagi suyakdir - aksariyat ornitomimozavrlarning L shaklidagi kvadratadugugalidan farq qiladi. Dorsal jarayonning uchi shikastlangan bo'lsa-da, aftidan oldingisidan uzunroq edi.[9] Orqa yuzada kvadrat o'rta balandlikda joylashgan katta depressiyani saqlaydi. Ushbu depressiya oval shaklida bo'lib, uning uzunligi 1,2 sm (12 mm) va kengligi 0,4 sm (4,0 mm). A pnevmatik foramen depressiyaning pastki qismida ham mavjud.[18] Depressiyaga qadar pastki mintaqaga yaqin to'rtburchakning lateral chegarasida parakuadrat teshiklari uchun trakt mavjud. Ikkalasi ham pastki jag 'kondillari kattaligi teng va yiv bilan yaxshi ajratilgan. The pterygoid uzatma lateral ko'rinishda uchburchak shaklida va pnevmatik teshik darajasida kamida 2,9 sm (29 mm) uzunlikda. Yuqori ko'rinishda frontallar uchburchak bo'lib, frontallarning oldingi tekislangan mintaqasi old tomonga moyil bo'ladi. Orqa uchiga frontallar bitta gumbazga o'xshash tuzilmani hosil qiladi. Ichki tomonlarda parietallar bosh suyagi tomining orqa qismida tekislangan. Boshsuyagi tomi uzoqroqdan ko'ra kengroq. Orqa jarayon qattiqlashadi va shu yo'nalishda rivojlanadi.[9]

The postorbital lateral ko'rinishda yaxshi taqsimlangan qalinlikning keskin rivojlangan suyagi. Frontal bilan tikish orbitaning yuqori orqa mintaqasidan kelib chiqadi va postorbital-parietal tikuv cho'zilib ketadi supratemporal fenestra. Pastki yo'nalishga, pastki jarayon tor. Ning pastki uchlari skuamozal shikastlangan va baland bo'yli oldingi jarayon ichki tomondan postorbitalning orqa jarayoni bilan bog'langan. Skuamozalning ikkala old va pastki kengaytmalari lateral ko'rinishda cho'zilgan. The tishsiz pastki jag'lar juda to'liq va yaxshi saqlanib qolgan. The stomatologik uzunligi 18,3 sm (183 mm) bo'lgan eng uzun mandibular element bo'lib, orqa uchiga qadar qalinlashgan.[9] Sharti bilan; inobatga olgan holda Garudimimus tish tishlari tushirilgan bo'lsa, jag'lar butun og'iz chegarasi bo'ylab o'tmaydi va bu jag'larning ishlashini cheklaydi. Romfotekaning mavjudligi okklyuzion sirt hosil qilish uchun bo'shliqni to'ldirishi mumkin.[17] Tish tishining yuqori chegarasi qirrasi bilan yaxshi rivojlangan va yonma-yon yumaloqlangan. Tish uchining lateral yuzalarida ko'plab teshiklarni topish mumkin. The simfiz (suyak birlashmasi) kam rivojlangan bo'lib, tillararo artikulyatsiya mustahkam bo'lmaganligini ko'rsatmoqda. Ikkala tish suti simfizida ham belkurakka o'xshash yumaloq tokchani keng va chuqur hosil qiladi. Ichki yuzada a Meckelian yiv topish mumkin, ammo uni taloq qoplaydi. Tish tishida ikkita orqa jarayon, biri tashqi mandibularning old chegarasida joylashgan fenestra.[9]
The taloq dentaryning qirrasi orqa uchida eng chuqur va oldingi uchida torayib boradi. Uning orqa qirrasi biroz chuqurlashgan va ichki ko'rinishdan oldinga moyil. Orqa uchi preartikulyarning pastki maydoniga yupqalashadi. Ikkinchi katta mandibular element burchakli. Uning dorsal kengaytmasi biroz tekislanib, pastki jag 'fenestrasining old chetiga qaraganda oldinga ko'proq cho'ziladi. Surangularning orqa tomoni butun tanasi bo'ylab ichki proektsiyani shakllantiradi. Ushbu chekka to tizmasiga o'xshash tuzilishga ega qo'shma kvadratning pastki jag 'kondilasi bilan. Surangularda orqa burchakli teshik teshiklari yo'q. U faqat retroartikulyar jarayonning lateral yuzasining oldingi yarmida qatnashadi. Retroartikulyar jarayon (orqaga qarab suyak kengaytmasi) uning orqa uchida muloyimlik bilan yuqoriga qarab turadi. Surangular bilan taqqoslaganda burchakli kichikroq va tashqi mandibular fenestrani pastki qismiga chegaralaydi. Ushbu suyaklar orasidagi to'g'ri tikuv tashqi mandibular fenestraning orqa uchidan pastki jag 'orqa qismigacha cho'ziladi. Burchakning oldingi jarayoni cho'zilgan va shu yo'nalishda tish bilan to'qnashgan. The preartikulyar katta va gavdali. Ushbu suyakning oldingi uchi tashqi mandibular fenestraning oldingi chetidan orqada yotadi va orqa tomonga kesiladi. The qo'shma to'rtburchakning pastki qistirmalari uchun sirtlar tog 'tizmasiga o'xshash tuzilish bilan yaxshi ajralib turadi. Ushbu tog 'tizmasining yuqori qismlari old va markaz tomonda joylashgan bo'lib, yon tomondan va oxirigacha joylashgan. Ichki yuzasida katta va chuqur pnevmatopora mavjud.[9]
Postkranial skelet

Ko'pchilik bachadon bo'yni umurtqalari eroziya yuzalariga ega va yonma-yon ezilgan; eng to'liq bachadon bo'yni o'qi. Chetga atlas va o'qi, yana oltita umurtqa pog'onasi saqlanib qolgan va ularni uchinchi va oltinchi bachadon bo'yinlariga bog'lash mumkin, ammo oxirgi ikkitasi holati noma'lum. Atlasning qisman intertsentrusi o'qning pastki mintaqasi va chapning yarmiga yaqin joylashgan asab kamari ham mavjud. O'qning markaziy uzunligi ancha qisqa bo'lib, uning uzunligi taxminan 3 sm (30 mm) va balandligi 1,8 sm (18 mm) ni tashkil etadi - ehtimol bu ornitomimozavrlarda ibtidoiy holat. Uning orqa bo'g'im yuzasi konkav va kengligidan balandroq. Markazning lateral yuzalarida pnevmatik fossa yo'q. Nerv umurtqasi baland va juda yumaloq bo'lib, a ligament chandig'i orqa chegarada va eksenel asab kamari markazining o'rta uzunligida pnevmatik chuqurchaga ega. Uchdan beshinchi servikslar bo'g'imlarga ega, ammo faqat asab kamarining o'ng tomonini saqlaydi va oltinchisi bilan to'rtinchi servikallar eng to'liq hisoblanadi. Asab umurtqalari tor va oxirigacha joylashgan. Ikkala so'nggi bo'yin bachadonlari aniqlangan va ularning aniq pozitsiyalari aniq emas, ammo ular cho'zilgan postzigafofizlar ularning orqa qatorga mansubligini taxmin qilish. Shunga qaramay, bu ikkalasining oldingi bachadon bo'yni ettinchi bo'ynidir. To'qqiz ketma-ket orqa umurtqalari saqlanib qolgan va ularga asoslangan Ornitomimus, ehtimol ular to'rtinchi va o'n ikkinchi dorsallarni ifodalaydi. Ular borgan sari kattalashib boradi va dorsallar atrofidagi ba'zi tikuvlar to'liq birlashtirilmaydi, bu holotip namunasi to'liq pishmaganligini anglatadi. Nerv o'murtalari kattaligi bilan bir qatorda, orqa dorsallarda tobora balandroq bo'lib boradi, ammo ularning uzunligi deyarli tengdir. Yanal ko'rinishda, barcha orqa miya oldinga singan, o'n ikkinchi qismdan tashqari oxirigacha ozgina moyil bo'ladi. Orqa umurtqalar bilan orqa santra kattaroqdir amfikoel (kuchli konkav) qirralar va lateral sirtlarda pnevmatizatsiya yo'q. To'rtinchi va beshinchi dorsal markazdagi pastki yuzalarda katta tizmaga o'xshash tuzilmalar mavjud. Sentraning lateral yuzalarida old va orqa qirralarning ham ligament izlari bor.[9]

The sakrum oltitadan iborat sakral vertebra shulardan birinchi va oltinchisining ichki yuzasiga biriktirilgan sakral qovurg'alar mavjud ilium. Sakrumning umumiy uzunligi - 31 sm (310 mm) - deyarli iliumga teng. Ko'pgina sakral santralarning uzunligi to'rtinchi qismdan tashqari taxminan teng. Sakrumning birlashtirilgan qismlari, masalan, birinchi va ikkinchi, uchinchi va to'rtinchi sakrallar mavjud, ammo ba'zi joylar qisman eritilmagan. Birinchi va beshinchi sakralallarning nerv umurtqalari birlashtirilgan va oltinchisining asab orqa miya etishmayapti. Bu tartibsiz suyaklanish kichik yoshdagi shaxsni ko'rsatishi mumkin. Sakral o'murtalarda pnevmatizatsiya yo'q va lateral ko'rinishda ligament izlari birinchi va oltinchi sakral o'murtaning old chegaralarida va beshinchi va oltinchi sakral umurtqalarning orqa qirralarida uchraydi. Birinchi to'rtlik kaudal vertebra markaz va asab kamari orasidagi barcha tikuvlar ochiq holda saqlanadi. Nerv umurtqalari tobora qisqarib boradi va old va orqa chegaralari yivlangan holda to'g'ri va gorizontal orqa chegaralarga ega bo'ladi. Sentraning uzunliklari teng va ularning pastki yuzalarida kaudal umurtqalar ikkala tomonning juft proektsiyalariga ega sulci (yoriqlar) uchun chevron artikulyatsiya.[9] Old dumg'aza umurtqalari nerv chuqurining o'ng tomonida joylashgan chuqur chuqur teshik fossa old nerv kamarining ikkala o'ng yuzasida va yon yuzasida joylashgan, dumg'aza pnevmatik xususiyatini ko'rsatishi mumkin Garudimimus.[19]
Uchinchi va to'rtinchi serviks chevronlari qisman faqat o'zlarining orqa va old qismlari bilan saqlanadi. To'qqiz dorsal qovurg'alar holotipda mavjud va qisqa sil kasalligi va uzoq kapitulum. O'n besh gastraliya parchalar saqlanib qolgan, ammo ularning aniq pozitsiyalari noma'lum. Ular, hech bo'lmaganda, lateral va ichki qismlarga bo'linadi. Ko'pgina sakral qovurg'alar kam rivojlangan. The tos kamari ammo har ikkisi ham yaxshi saqlanib qolgan iskiya mavjud emas. Ikkalasi ham ilia muloyimlik bilan eziladi va iliyaning dorsal qirralari bir-biriga yaqin yotadi, oldingi uchi kanca shaklida bo'ladi. Yanal ko'rinishda iskiyadik pedunkul (iskiumni biriktirish uchun asetabulum orqasidagi kengaytma) xanjar shaklida va uchli va boshqa ornitomimozavrlarning pastki qismidan keng. The brevis fossa (caudofemoralis brevis mushak qo'shilishi) katta, ammo juda sayoz, qo'shimcha ravishda ilium ornitomimidlar kabi keng emas. Cuppedicus fossa (orasidagi chuqurlik preacetabular pichoq va pubik pedunkul ) brevis fossadan kichikroq. Pubis - uzunligi 39 sm (390 mm) bo'lgan katta element. Yanal ko'rinishda yuqori uchi pubis-ischium kontaktining pastki chegarasida ilgakka o'xshash cho'zilgan tuzilishga ega. Uning o'qi deyarli to'g'ri va pubik botinkaning orqa kengaytmasi (oxirida katta proektsiya) oldingi kengaytmaga qaraganda uzunroq. Ikkala pub ham birlashtirilgan.[9]
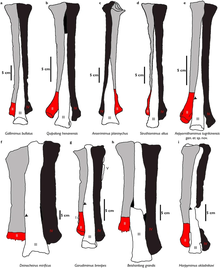
The suyak suyagi o'lchamlari 37,1 sm (371 mm) va pastki ko'rinishdan, umumiy uzunligi femur boshi uning yuqori uchi bo'ylab joylashgan kenglikning yarmidan ko'pi. Oldingi chegarasida kamroq trokanter, katta qo'shimcha trokanterni topish mumkin va bu trochanters sulkus bilan bo'linadi. The to'rtinchi trokanter yuqori uchidan femur uzunligining taxminan 1/3 qismida joylashgan. The tibial lateralning artikulyar o'simtasi kondil orqa yo'nalishda ishlab chiqilgan. The tibiotarsus -femur uzunlik koeffitsienti 1,1 ga teng va uning yuqori uchi bir xil darajada rivojlangan lateral va ichki orqa kondillarga ega. The fibula ga nisbatan bir oz qisqaroq tibia (Mos ravishda 36 sm (360 mm) va 38,8 sm (388 mm)). Ushbu suyak boshqa oyoq elementlariga qaraganda ancha yassilangan va o'qi juda ingichka. Ikkalasi ham astragal va kaltsiy chap tibia bilan bog'langan, ammo kaltseyum ozgina zararlangan ko'rinadi. Pastki qatorlar orasida tarsal saqlanib qolgan elementlar III va IV. Tarsal III uchinchi metatarsalning yuqori yuzasiga va qisman ikkinchi metatarsal bilan og'riydi. Tarsal IV to'rtinchi metatarsalning yuqori sirtining ko'p qismini qoplaydi.[9]
In pes formula, Garudimimus beshga ega bo'lishi bilan ornitomimid holatidan keskin farq qiladi metatarsallar. Metatarsal I - ning birinchi va eng kichik elementi metatarsus yanada tekislangan va qisilgan shaklga ega. Metatarsal I va metatarsal II orasidagi bo'g'im yuzasi ikkinchisining orqa yuzasida joylashgan. Metatarsallar II va IV uzunlik bo'yicha tengsiz va metatarsal III eng uzun (22,9 sm (229 mm)). Metatarsal III ning pastki uchi frontal ko'rinishda kengroq va yuqori qismida mahkamlanmagan, shuning uchunarktometatarsalian holat.[9] Ushbu xususiyat qisman katta bilan taqsimlanadi Beishanlong va Deinocheirus.[20] Metatarsal V - bu juda kamaytirilgan va ingichka element bo'lib, u funktsional ahamiyatga ega emas va metatarsal IV ning orqa yuzasi bilan aniqlanadi. Faqat sakkizta pedal falanjlar va ikkita chap pedal jinsiy bo'lmaganlar holotip bilan mavjud. Oyoqlarda to'rtta ibtidoiy narsa bor edi oyoq barmoqlari birinchisi eng kichigi bilan. Aksariyat falanjlar old va orqa uchlarida dumaloq lateral kondil va konkav old artikulyar yuzalar bilan kengaygan shakliga juda o'xshashdir. Ular ikkinchisidan to'rtinchisiga borgan sari qisqaroq bo'ladilar raqam.[9] Ikkala saqlanib qolgan turli xil jinslar ham takrorlangan shaklga ega, ular ornitomimidlardagi to'g'ri shakldan farq qiladi.[20] To'rtinchi raqamdan kelgan noaniq odam taniqli fleksorga ega sil kasalligi (egiluvchan tendonlar ilova). Ikkinchi raqam ungual kuchli fleksor tuberkulyozi bilan oldingi raqamga o'xshaydi, ammo shakli ancha notekis.[9]
Tasnifi

1981 yilda Barbold tayinlandi Garudimimus alohida Garudimimidae taksondagi g'ayrioddiy xususiyatlarni hisobga olgan holda. Biroq, u ularni aniq ibtidoiy deb aniqladi.[1] 2005 yilda holotipni qayta tavsiflash bilan ko'proq diagnostik belgilar Garudimimus ta'minlandi. Kobayashi va Barsboldning ta'kidlashicha, ushbu takson ibtidoiy va kelib chiqadigan ornitomimozavrlarga xos xususiyatlarni, masalan, to'rt barmoqli oyoqlar va tishsiz jag'lar bilan bo'lishgan. Ularning filogenetik tahlili tiklandi Ornithomimidae sifatida opa takson ning Garudimimus.[9] 2014 yilda uzoq vaqtdan beri sirli bo'lgan yangi va juda to'liq namunalar Deinocheirus Yuong-Nam Li va uning hamkasblari tomonidan tasvirlangan. Qoldiqlar buni ko'rsatdi Deinocheirus aslida ornitomimozavr taksoni bo'lgan, ammo mutlaqo boshqacha edi va shuningdek, yoshini aniqlashga imkon berdi Deinocheiridae oila sifatida, shu jumladan, yaqinroq umumiy ajdodlari bilan birgalikda barcha taksilar Deinocheirus bilan qaraganda Ornitomimus. Li va jamoasi Deinocheiridae uchun katta filogenetik tahlil o'tkazdilar va uchta guruh ushbu guruhda topildi: Beishanlong, Garudimimus va Deinocheirus. Ularning o'zaro munosabatlari asosan oyoq-qo'llarning turli xil anatomik xususiyatlari bilan ifodalangan, ammo oxirgi ikkalasi bir-biri bilan chambarchas bog'liq edi, chunki ularning ikkalasi ham bosh suyagi va tana qoldiqlariga ega. Natijada kladogramma ornitomimozavr dinozavrlari erta bo'r davridagi ikkita asosiy naslga bo'linishini taklif qildi: Deinocheiridae va Ornithomimidae. Tez ishlaydigan ornitomimidlardan farqli o'laroq, deinokeyridlar yugurish uchun qurilmagan va ularning anatomik xususiyatlari Deinocheirus boshqa ornitomimozavrlar bilan taqqoslaganda, ushbu taksonda erishilgan sezilarli kattaliklar bilan izohlash mumkin.[20] Yaqinda, 2019 yilda Xartman va uning hamkasblari yangi paravianni tasvirlab berishdi Hesperornithoides uchun keng filogenetik tahlilni o'tkazish Coelurosauria. Ornitomimozavrlarga nisbatan, Garudimimidae o'z ichiga olgan oila sifatida tiklandi Garudimimus va yaqin qarindoshlari. Garudimimus ning singlisi taksoni sifatida ancha olingan holatda tiklandi Beishanlong. Ushbu tahlil o'zaro yaqin aloqani topdi Deinocheirus va Garudimimus iloji yo'q, chunki avvalgi bazada tiklangan Ornitomimozauriya.[21]
Li va uning hamkasblari uchun chap kladogramma 2014,[20] Hartman va uning hamkasblari uchun to'g'ri kladogramma 2019:[21]
|
|
2020 yilda Serrano-Brañas va jamoasi yangi deinokeyrid turini ta'rifladilar va ularga nom berdilar Paraxenisaurus dan Cerro del Pueblo shakllanishi. Bu guruhning Osiyodan tashqarida topilgan birinchi a'zosi va uning kashf etilishi deinokeyridlar kelib chiqqanligini ko'rsatishi mumkin Laurasiya (shimoliy superkontinent davomida Mezozoy ) yoki ular qutb mintaqalari bo'ylab tarqalib ketganligi Shimoliy yarim shar; shunga o'xshash almashinuv, shuningdek, Osiyo yaqinligiga ega bo'lgan boshqa dinozavrlar guruhida ham sodir bo'lganligi ma'lum Kampanian -Maastrixtiy yoshi. Jamoa topdi Harpimimus juda ibtidoiy deinokeyrid bo'lish, ammo Beishanlong guruhdan tashqarida joylashtirilgan. Garudimimus va Deinocheirus qo'shilishi bilan yaqindan bog'liq taksonlar sifatida yana tiklandi Paraxenisaurus a polotomiya.[22]
Paleobiologiya

Oyoqlarning nisbati, pedalga xos bo'lmagan morfologiya va ilyumning kengayishi Garudimimus tezroq moslashtirilmaganligini taxmin qiling kursor ornitomimidlar, ular kuchli bilan ajralib turadi sonlar va to'g'ri jinssizlar.[9][20] 2011 yilda Lars Shmitz va Ryosuke Motani o'lchamlarini o'lchashdi sklerotik halqa va ko'z teshigi fotoalbomlarda dinozavrlar va pterozavrlarning namunalari, shuningdek ba'zi tirik turlari. Ular buni ta'kidladilar kunduzgi (fotopik) hayvonlar kichikroq sklerotik halqalarga ega, tungi (skotopik) hayvonlar ko'proq kattalashgan halqalarga ega. Katemer (mezopik) hayvonlar - kun bo'yi va tun davomida tartibsiz ravishda faol bo'lganlar - bu ikkita oraliq oralig'ida. Shmitz va Motani ekologik va filogenetik omillarni ajratib, 164 tirik turni o'rganib, diurallik, katemerlik yoki nokturnalilik haqida xulosa chiqarishda ko'z o'lchovlari juda aniq ekanligini payqashdi. tetrapodlar. Natijalar shuni ko'rsatdiki Garudimimus boshqa yirtqich teropodlar asosan tungi bo'lgan kateterli o'txo'r edi. Yirik o'txo'r dinozavrlar katemer bo'lganligi aniqlandi, chunki bu vaqt ko'p bo'lgan em-xashak va termoregulyatsiya omillar. Qushlar va pterozavrlar asosan kunduzgi bo'lib, tungi istisnolardan tashqari. Ular ekologik nish diel faoliyatini rivojlantirishda asosiy harakatlantiruvchi omil bo'lgan degan xulosaga kelishdi.[23] Shu bilan birga, ushbu tahlil Xoll va uning hamkasblari tomonidan Shmit va Motanining usullariga qarshi bahs yuritib, kunduzgi, katemer va tungi hayvonlar o'rtasida skleral anatomiyada juda o'xshashlik borligi haqida bahslashdi. Bundan tashqari, yumshoq to'qimalar kabi shox parda fotoalbom namunalarida aniq bo'lmagan ko'zga qancha yorug'lik tushishini aniqlashda juda muhimdir.[24] Shmitz va Motani ushbu izohga o'zlarining tadqiqotlari va metodologiyalarini mavjud turlarga asoslangan holda yo'q bo'lib ketgan dinozavrlarning xatti-harakatlarini to'g'ri tasniflashlarini tasdiqlagan holda javob berishdi. Ular vizual imkoniyatlarni tiklashda ishonchli manba sifatida skleral halqaning tashqi chegarasini ko'rsatdilar.[25]
2019 yilda Grem M. Xyuz va Jon A. Finarelli tahlil qildi xushbo'y lampochka qancha qushni taxmin qilish uchun zamonaviy qushlar va bir necha yo'q bo'lib ketgan dinozavr turlarining saqlanib qolgan bosh suyaklaridagi nisbati genlar yo'q bo'lib ketgan ushbu turlarning xushbo'y hidli bo'lishida ishtirok etgan bo'lar edi. Ularning tahlillari shuni ko'rsatdiki, ornitomimozavrlarning aksariyatida hidlash sezgirligi nisbatan zaif bo'lgan; Garudimimus hidlash retseptorlarini kodlovchi 417 genga va hidning lampochkaning nisbati 28,8 ga teng bo'lib, bu hidning yomon rivojlanganligini ko'rsatmoqda. Ushbu qiymatlar katta dinozavr turlarida, masalan, ko'proq aniqlanganligini hisobga olsak tirannosauridlar, Xyuz va Finarelli ta'kidlashlaricha, dinozavr nasllari kattalashgan sari, hidlovchi lampochkaning kattaligi ham oshgan, bu esa ularni taklif qilishi mumkin olfaktsiya qush bo'lmagan dinozavrlarda dominant hissiy modal sifatida.[26]
Oziqlantirish odatlari

Kobayashi va Barsbold 2005 yilda pastki jag 'qismida tish tishlari yo'qligi va bir xil shaklga keltirilganligini ta'kidladilar. Garudimimus boshqa ibtidoiy ornitomimozavrlarga qaraganda oziq-ovqat iste'mol qilishda ornitomimidlarga ancha o'xshash edi. Ornitomimidlar singari, yuqori va pastki jag'larning old qismlari Garudimimus ikkalasi ham tumshug'i bilan yopilgan va ovqat yulib olish uchun ishlatilgan bo'lishi mumkin. Bundan tashqari, jag'larning o'rta qismidagi o'tkir qirralar ovqatni kesishga yordam berdi.[9] Bundan tashqari, Garudimimus va qarindoshlar sifatida keng tarqalgan o'txo'r yoki hech bo'lmaganda hamma narsa dinozavrlarni o'ziga xos anatomik xususiyatlari, shu jumladan tumshug'i borligi.[27][23]
2015 yilda Endryu R. Kuff va Emili J. Reyfild chiqish qildilar KT tekshiruvi kuni taponomik jihatdan Bosh suyaklarining buzilgan namunasi Garudimimus, Ornitomimus va Struthiomimus retrodeformatsiya qilish uchun - deformatsiyalangan ob'ektning asl holatini yaratish uchun ishlatiladigan usul. Namunalarning skanerlashi va nisbatan to'liqligi aduktorni qayta tiklashga imkon berdi (harakatni keltirib chiqaruvchi) muskulatura pastki jag'ning, keyin bosh suyaklariga xaritada tushirilgan. Bundan tashqari, remonteka (tumshuq) ning kattaligi a-ga asoslangan holda qayta tiklandi ratit qush, tuyaqush - va katta morflar. O'rganilgan uchta nasl orasida Garudimimus eng past luqma kuchiga ega edi, 19 ishlab chiqarishi aniqlandi N tumshug'ining uchida va xuddi shu uzunlikning o'rta uzunligida 23,9 N. Biroq, bu taksonda eng ko'p o'zgartirilgan mushak bor edi mexanik afzallik va lahzali qo'l - mushaklarning bir qator konfiguratsiyalar bo'yicha ma'lum bir harakatga hissa qo'shishi samaradorligining ko'rsatkichi. Shu nuqtai nazardan, mushaklarning orasidagi eng muhim farq Garudimimus va ikkita ornitomimid topilgan edi, aksariyat mushaklar oxirgi ikkitasida mexanik jihatdan foydaliroq; Bu bosh suyagi uzunroq bo'lganligi sababli yuzaga kelishi mumkin Garudimimus. Cuff and Rayfield ning tishlash kuchi ekanligini ta'kidladilar Garudimimus mandibulasidan foydalanish uchun cheklangan bo'lishi mumkin Struthiomimus rekonstruktsiyalarda yoki MPC-D 100/13 kichik yoshdagi shaxsdir. Taxminan, ular orbitomimozavrlarda ovqatlantirish paytida o'simlik materialini ushlab turish uchun rampoteka bilan birlashtirilgan nisbatan past tishlash kuchlari ishlatilishi mumkin degan kelishuvga kelishdi, bularning hammasi bo'yin muskulaturasi yulib olish uchun etarli kuch berdi. o'simlik.[17]
2019 yilda, Devid J. tugmasi va Lindsay E. Zanno parranda bo'lmagan dinozavrlar orasida o'txo'rlikning ko'plab paydo bo'lishini tahlil qilish uchun 160 mezozoy dinozavr turlari tomonidan taqdim etilgan bosh suyagi biomexanik belgilariga asoslangan katta filogenetik tahlilni o'tkazdi. Ularning natijalari shuni ko'rsatdiki, o'txo'r dinozavrlar asosan ikki xil ovqatlanish rejimiga rioya qilishgan, yoki oziq-ovqat mahsulotlarini ichakda qayta ishlash - nisbatan nozik bosh suyaklari va kam tishlash kuchlari bilan tavsiflanadi - yoki og'iz, bu yuqori tishlash kuchlari kabi keng ishlov berish bilan bog'liq xususiyatlar bilan tavsiflanadi. jag'ning mustahkam mushaklari. Garudimimus va Deinocheirus, bilan birga kaenagnatid, diplodokoid, ornitomimid, trizinosaurid va titanozavr dinozavrlar avvalgi toifaga kirganligi aniqlandi Garudimimus chindan ham past luqma kuchlari bor edi va oziq-ovqat mahsulotlarini qayta ishlashda asosan oshqozoniga tayanardi. Tananing katta o'lchamlari mustaqil ravishda deinokeyridlar va ornitomimidlarda qo'lga kiritildi va kattaroq o'lchamlar o'txo'r hayvonlarga afzalliklarni beradi, masalan, qarshilik kuchayishi ro'za va boshqalar yutish. Demak, bu odatlar deinokeyridlar va ornitomimidlarda ko'proq o'txo'r hayot tarzini ko'rsatishi mumkin. Biroq, Tugma va Zanno o't o'simliklari va hajm o'rtasidagi bog'liqlik har doim ham bir-biriga bog'liq emasligini va ornitomimozavrlarda massa ko'payish tendentsiyasi mavjud emasligini ta'kidladilar. Bundan tashqari, aniq nişler faqat ornitomimozauriya dinozavrlari uchun yomon ekspluatatsiya qilingan Deinocheirus fursatparast omnivoryni namoyish etish. Tugma va Zanno ovqatlanish strategiyasi vaqt o'tishi bilan bog'liq bo'lmagan o'txo'r dinozavrlarda bir-biriga yaqinlashib borgan degan xulosaga kelishdi.[28]
Xuddi shu yili Ali Nabavizoda jag 'mushaklari o'txo'r dinozavrlarda va ba'zilari asosan ortal oziqlantiruvchi, ya'ni jag'larini yuqoriga va pastga siljitish bilan shug'ullanadi, degan xulosaga kelishdi. tropodlar, sauropodomorflar, ibtidoiy ornithischians va ba'zilari stegozavrlar. Nabavizodaning ta'kidlashicha, ornitomimozavrlar ortal oziqlantiruvchi va ularning jag'larini izognatus ko'targanlar, shu bilan har ikki tomonning yuqori va pastki tishlari birdan tiqilib (bir-biriga tegib ketgan). Deinocheirus va Garudimimus jag'larni ochadigan va yopadigan muskullar uchun o'ziga xos biriktirma joylari bor, ular pastga kengaygan, uchburchak shaklidagi adductor mushak tizimining oldinga kengayishiga biriktirilgan. burchakli suyak. Yalang'och jugal gardish bilan birgalikda bu qo'shimcha mushak uchun ko'proq bo'shliq beradi. Bunday cho'zinchoq tumshug'i bilan ikkala deinokeyridning tishlash kuchlari past bo'lib, mexanik ustunligi pasaygan, ammo agar bu mushak oldinga cho'zilsa, bu kuchga kuch qo'shgan bo'lar edi. vaqtinchalik mushak.[29]
Paleo muhit

Ning ma'lum qoldiqlari Garudimimus Oxirgi bo'r davridan topilgan Bayan Shireh shakllanishi Bayshi Tsav joyida.[1] Ushbu shakllanish asosan tomonidan tuzilgan gil toshlar va qumtoshlar bilan konkretsiyalar va xarakterlanadi loy toshlari va sarg'ish-jigarrang o'rta donali qumtoshlar. Kalikaning, flüvial va lakustrinli cho'kindi jinslarning turli joylarda bo'lishi bu qatlamning cho'kindi daryo daryolari va boshqa yirik suv havzalari allyuvial tekislik (baland tog 'daryolari yotqizilgan cho'kindi jinslardan tashkil topgan tekis er) a yarim quruq iqlim.[30][31] Bayshi Tsavning o'zi depozitda saqlangan deb talqin etiladi flüvial atrof-muhit.[32] Foydalanish kaltsit U – Pb o'lchovlari, Bayan Shireh shakllanishining yoshi taxminan 95,9 ± 6,0 milliondan 89,6 ± 4,0 gacha bo'lgan million yil oldin davomida Senomiyalik -Santonian bosqichlar.[33]
Baynshir hududidagi ko'plab shkalaga o'xshash o'zaro faoliyat qatlamlar yirik meandrlarning ko'rsatkichidir va bu suv havzalarining aksariyati sharqiy qismini quritgan bo'lishi mumkin. Gobi sahrosi.[31] Ushbu nam muhit suv havzasi sudralib yuruvchilarni kashf qilish orqali yanada qo'llab-quvvatlanadi toshbaqalar va krokodilomorflar va ostrakodlar.[34][35] Shu kabi qazilma to'plamlari asosida, Bayan Shireh va Iren Dabasu ko'plab mualliflar tomonidan shakllanishlar taklif qilingan.[30][6][36] Bundan tashqari, Iren Dabasu qatlami 95,8 ± 6,2 million yil oldin tuzilgan.[37]

Garudimimus yashash joyini bir nechta dinozavr bilan bo'lishdi fauna, masalan, o'txo'rlar ornithischians Gobihadros, Graciliceratops va Talarurus.[38][39] Boshqa tropodlar o'rta o'lja yirtqichi bilan ifodalanadi Achillobator,[40] va terizinosauridlar Erlikosaurus va Segnozavr.[41] Erketu hayvonot dunyosidagi eng baland o'simlikxo'r bo'lishi mumkin.[42] Bir nechta turlar ajratilgan joy farqi, kabi Erlikosaurus va katta Segnozavr,[41] yoki o'tloq Talarurus va brauzer Tsagantegiya, boshqa ankilozaurid shakllanishidan.[39] Xususida biologik xilma-xillik, Therizinozavrlar eng ko'p bo'lganlar mo'l-ko'l qatlam bo'ylab bir nechta tasvirlangan va ta'riflanmagan namunalari bo'lgan terropodlar.[35] Biroq, tarqalib ketgan hadrosauroid qoldiqlarining yuqori kontsentratsiyasi, xususan, Baynshirda juda katta mo'llikni aks ettiradi;[32] Gobihadros kamida uchta turli joylardan ma'lum.[43] Garchi Garudimimus Hozirda "Bay Shireh Formation" nomi berilgan yagona ornitomimozavr taksoni Bayshi Tsavda mahalliy darajada mo'l-ko'l ornitomimozavrlarni ishlab chiqardi.[44][45][10] Ular orasida norasmiy "Gallimimus mongoliensis " eng to'liq turlardan biri hisoblanadi, ammo bu tur unchalik yoshroqga tegishli emas Gallimimus va butunlay yangi taksonni anglatadi.[46]
Shuningdek qarang
Adabiyotlar
- ^ a b v Barsbold, R. (1981). "Bezzubyye khishchnyye dinozavry Mongolii" [Mo'g'ulistonning tishsiz go'shtli dinozavrlari]. Bitimlar, qo'shma sovet-mo'g'ul paleontologik ekspeditsiyasi (rus tilida). 15: 28−39. S2CID 195060070. Tarjima qilingan qog'oz
- ^ Barsbold, R. (1983). "Xishchnye dinozavry mela Mo'monii" [Mo'g'ulistonning bo'r davridan kelgan go'shtli dinozavrlar] (PDF). Sovet-mo'g'ul qo'shma paleontologik ekspeditsiyasining operatsiyalari (rus tilida). 19: 1−120. Tarjima qilingan qog'oz
- ^ Barsbold, R .; Osmólska, H. (1990). "Ornitomimozauriya". Vayshampelda D. B.; Osmolska, H.; Dodson, P. (tahrir). Dinozavrlar (1-nashr). Berkli: Kaliforniya universiteti matbuoti. 225-244 betlar. ISBN 9780520067271.
- ^ Currie, P. J.; Rassell, D. A. (1988). "Kanadadagi Alberta shtatining Judit daryosidan (Oldman) hosil bo'lgan Chirostenotes pergracilis (Saurischia, Theropoda) osteologiyasi va aloqalari". Kanada Yer fanlari jurnali. 25 (7): 272−286. Bibcode:1988CaJES..25..972C. doi:10.1139 / e88-097.
- ^ Xolts, T. R. (1992). Bo'r davridagi Theropoda metatarsining g'ayrioddiy tuzilishi (Arxosauriya: Dinosauriya: Saurischia) (Doktorlik dissertatsiyasi). Yel universiteti. p. 347.
- ^ a b Currie, P. J.; Eberth, D. A. (1993). "Iren Dabasu qatlamining paleoontologiyasi, sedimentologiyasi va paleoekologiyasi (Yuqori bo'r), Xitoy Xalq Respublikasi, Ichki Mo'g'uliston". Bo'r davridagi tadqiqotlar. 14 (2): 127−144. doi:10.1006 / cres.1993.1011.
- ^ Xolts, T. R. (1994). "Arktometatarsalian pes, bo'r Teropodasi metatarsining g'ayrioddiy tuzilishi (Dinosauria: Saurischia)". Umurtqali hayvonlar paleontologiyasi jurnali. 14 (4): 480−519. doi:10.1080/02724634.1995.10011574. JSTOR 4523590.
- ^ Peres-Moreno, B. P.; Sanz, J. L .; Buskalioni, A. D .; Moratalla, J. J .; Ortega, F.; Rasskin-Gutman, D. (1994). "Ispaniyaning Quyi bo'r davridan noyob ko'p qirrali ornitomimozavr dinozavr" (PDF). Tabiat. 370 (6488): 363−367. Bibcode:1994 yil Natur.370..363P. doi:10.1038 / 370363a0. S2CID 4302245.
- ^ a b v d e f g h men j k l m n o p q r s t siz v Kobayashi, Y .; Barsbold, R. (2005). "Garitimimus brevipes Barsbold, ibtidoiy ornitomimozavrni qayta tekshirish, 1981 yil (Dinosauriya: Theropoda), Mo'g'ulistonning so'nggi bo'r davridan" (PDF). Kanada Yer fanlari jurnali. 42 (9): 1501−1521. Bibcode:2005CaJES..42.1501K. doi:10.1139 / e05-044. hdl:2115/14579.
- ^ a b Chinzorig, T .; Kobayashi, Y .; Saneyoshi, M .; Tsogtbaatar, K .; Batamxatan, Z .; Ryuji, T. (2017). "Mo'g'ulistonning janubi-sharqiy Gobi cho'lining yuqori bo'r davridagi Bayshirshir shaklidagi ikkita yangi ornitomimidlarning (Theropoda, Ornithomimosauria) multitaksik suyagi". Umurtqali hayvonlar paleontologiyasi jurnali. Dastur va tezislar: 97.
- ^ Chinzorig, T .; Kobayashi, Y .; Tsogtbaatar, K .; Currie, P. J.; Takasaki, R .; Tanaka, T .; Iijima, M .; Barsbold, R. (2018). "Mo'g'ulistonning Nememt shakllanishidan kelib chiqqan ornitomimozavrlar: manus morfologik o'zgarishi va xilma-xilligi". Paleogeografiya, paleoklimatologiya, paleoekologiya. 494: 91–100. Bibcode:2018PPP ... 494 ... 91C. doi:10.1016 / j.palaeo.2017.10.031.
- ^ Tsogtbaatar, K. (2019). Mo'g'ulistonning yuqori bo'r davridagi ornitomimozavrlarning (Dinosauriya: Theropoda) evolyutsiyasi, xilma-xilligi va nomutanosibligi. (PDF) (Doktorlik dissertatsiyasi). Xokkaydo universiteti. hdl:2115/74432.
- ^ Temperton, J .; Rozewarne, G.; Kindersli, D. (1993). Kindersli, D. (tahrir). Dinozavrlarning vizual lug'ati. Guvohlarning vizual lug'atlari. Pensilvaniya shtati universiteti: DK CHILDREN. 1-64-betlar. ISBN 9780751310122. LCCN 92053446. OCLC 38325967.
- ^ Zanno, L. E .; Makovicky, P. J. (2013). "O'simlikxo'r teropod dinozavrlarda tana massasining yo'naltirilgan evolyutsiyasi to'g'risida dalillar yo'q". Qirollik jamiyati materiallari B. 280 (1751): 20122526. doi:10.1098 / rspb.2012.2526. PMC 3574415. PMID 23193135.
- ^ Zelenitskiy, D. K .; Terrien, F .; Erikson, G. M .; DeBur, K. L.; Kobayashi, Y .; Eberth, D. A .; Hadfild, F. (2012). "Shimoliy Amerikadan parranda qilingan parranda bo'lmagan dinozavrlar qanotning kelib chiqishi to'g'risida tushuncha beradi". Ilm-fan. 338 (6106): 510–−514. Bibcode:2012Sci ... 338..510Z. doi:10.1126 / science.1225376. PMID 23112330. S2CID 2057698.
- ^ Van der Reest, A. J .; Vulf, A. P.; Currie, P. J. (2016). "Yuqori bo'r dinozavr parki shakllanishidan zich tukli ornitomimid (Dinosauria: Theropoda), Alberta, Kanada". Bo'r davridagi tadqiqotlar. 58: 108−117. CiteSeerX 10.1.1.710.3240. doi:10.1016 / j.cretres.2015.10.004.
- ^ a b v d e Manjet, A. R .; Reyfild, E. J. (2015). "Ornitomimozauriya dinozavr kraniyasining retrodeformatsiyasi va mushaklarning qayta tiklanishi". PeerJ. 3: e1093. doi:10.7717 / peerj.1093. PMC 4512775. PMID 26213655.
- ^ Xendrikx, S.; Araujo, E .; Mateus, O. (2015). "Qushlarga tegishli bo'lmagan tropod kvadrati I: anatomiya va funktsiyalarga umumiy nuqtai nazardan standartlashtirilgan terminologiya". PeerJ. 3: e1245. doi:10.7717 / peerj.1245. PMC 4579021. PMID 26401455.
- ^ Vatanabe, A .; Oltin, M. E. L. G.; Brusatte, S. L.; Benson, R. B. J .; Xoyyer, J .; Devidson, A .; Norell, M. A. (2015). "Ornitomimozavrdagi arxeornitomimusdagi umurtqali pnevmatik kasallik (Dinozavriya: Theropoda) kompyuter tomografiyasi yordamida aniqlandi va Ornitomimozauriyada eksenel pnevmatik kasallikning qayta baholanishi". PLOS ONE. 10 (12): e0145168. Bibcode:2015PLoSO..1045168W. doi:10.1371 / journal.pone.0145168. PMC 4684312. PMID 26682888.
- ^ a b v d e Li, Y.-N .; Barsbold, R .; Currie, P. J.; Kobayashi, Y .; Li, H.-J .; Godefroit, P.; Escuillié, F.; Tsogtbaatar, C. (2014). "Deinocheirus mirificus ulkan ornitomimozavrining uzoq yillik sirlarini echish". Tabiat. 515 (7526): 257−260. Bibcode:2014 yil Noyabr 515..257L. doi:10.1038 / tabiat13874. PMID 25337880. S2CID 2986017.
- ^ a b Xartman, S .; Mortimer, M.; Vahl, V. R.; Lomaks, D. R .; Lippincott, J .; Lovelace, D. M. (2019). "Shimoliy Amerikaning So'nggi Yura davridan kelgan yangi paraviya dinozavri parranda parvozini kechroq sotib olishni qo'llab-quvvatlamoqda". PeerJ. 7: e7247. doi:10.7717 / peerj.7247. PMC 6626525. PMID 31333906.
- ^ Serrano-Brañas, C. I .; Espinosa-Chaves, B.; Makraken, S. A .; Gutieres-Blando, S.; de-Leon, C .; Ventura, J. F. (2020). "Paraxenisaurus normalensis, Cerro del Pueblo shakllanishidan katta deinokeyrid ornitomimozavri (Yuqori bo'r), Meksika, Koaxila". Janubiy Amerika Yer fanlari jurnali. 101: 102610. Bibcode:2020JSAES.101j2610S. doi:10.1016 / j.jsames.2020.102610.
- ^ a b Shmitz, L .; Motani, R. (2011). "Skleral halqa va orbitaning morfologiyasidan kelib chiqqan dinozavrlarda noxushlik". Ilm-fan. 332 (6030): 705−708. Bibcode:2011Sci ... 332..705S. doi:10.1126 / science.1200043. PMID 21493820. S2CID 33253407.
- ^ Hall, M. I .; Kirk, E. C .; Kamilar, J. M .; Carrano, M. T. (2011). "Skleral halqa va orbitaning morfologiyasidan kelib chiqqan dinozavrlarning bexabarligi to'g'risida sharh". Ilm-fan. 334 (6063): 1641. Bibcode:2011 yil ... 334.1641H. doi:10.1126 / science.1208442. PMID 22194558.
- ^ Shmitz, L .; Motani, R. (2011). "Skleral halqa va orbitaning morfologiyasidan kelib chiqqan dinozavrlarning bexabarligi haqidagi sharhga javob". Ilm-fan. 334 (6063): 1641. Bibcode:2011 yil ... 334.1641S. doi:10.1126 / science.1208489.
- ^ Xyuz, G. M .; Finarelli, J. A. (2019). "Dinozavrlarda xushbo'y retseptorlari repertuarining hajmi". Qirollik jamiyati materiallari B. 286 (1904): 20190909. doi:10.1098 / rspb.2019.0909. PMC 6571463. PMID 31185870.
- ^ Barrett, P. M. (2005). "Tuyaqushlarning dinozavrlari parhezi (Theropoda: Ornithomimosauria)". Paleontologiya. 48 (2): 347−358. doi:10.1111 / j.1475-4983.2005.00448.x.
- ^ Tugma, D. J .; Zanno, L. E. (2019). "Qush bo'lmagan dinozavrlarda o'tqazish turlarining xilma-xil rejimlarining takroriy evolyutsiyasi". Hozirgi biologiya. 30 (1): 158−168.e4. doi:10.1016 / j.cub.2019.10.050. PMID 31813611. S2CID 208652510.
- ^ Nabavizadeh, A. (2019). "O'simlikxo'r dinozavrlarda kranial muskulatura: qayta tiklangan anatomik xilma-xillik va ovqatlanish mexanizmlarini o'rganish". Anatomik yozuv. 303 (4): 1104−1145. doi:10.1002 / ar.24283. PMID 31675182. S2CID 207815224.
- ^ a b Jerzykievich, T .; Rassell, D. A. (1991). "Gobi havzasining kech mezozoy stratigrafiyasi va umurtqali hayvonlari". Bo'r davridagi tadqiqotlar. 12 (4): 345–377. doi:10.1016/0195-6671(91)90015-5.
- ^ a b Xiks, J. F .; Brinkman, D. L .; Nichols, D. J .; Watabe, M. (1999). "Mo'g'ulistonning sharqiy Gobi cho'lidagi Bayn Shireh, Burkhont va Xuren Duxdagi Albian - Santonian qatlamlarini paleomagnitik va palinologik tahlillari". Bo'r davridagi tadqiqotlar. 20 (6): 829−850. doi:10.1006 / cres.1999.0188.
- ^ a b Ishigaki, S .; Tsogtbaatar, K .; Saneyoshi, M .; Maynbayr, B .; Aoki, K .; Ulziitseren, S .; Imayama, T .; Takaxashi, A .; Toyoda, S .; Bayardorj, C .; Buyantegsh, B .; Batsux, J .; Purevsuren, B.; Asai, H .; Tsutanaga, S .; Fujii, K. (2016). "Okayama Fan universiteti - Mo'g'uliston Paleontologiya va geologiya instituti qo'shma ekspeditsiyasining 2016 yildagi hisoboti" (PDF). Tabiiy fanlar ilmiy-tadqiqot instituti byulleteni (42): 33−46.
- ^ Kurumada, Y .; Aoki, S .; Aoki, K .; Kato, D .; Saneyoshi, M .; Tsogtbaatar, K .; Uindli, B. F.; Ishigaki, S. (2020). "Mo'g'ulistonning Sharqiy Gobi cho'lida Bayn Shire shakllangan bo'r umurtqali kaltsit U-Pb yoshi: kaliyning yoshni aniqlash uchun foydaliligi". Terra Nova. 32 (4): 246−252. Bibcode:2020TeNov..32..246K. doi:10.1111 / ter.12456.
- ^ Danilov, I. G.; Xirayama, R .; Suxanov, V. B.; Suzuki, S .; Vatabe, M.; Vitek, N. S. (2014). "Mo'g'ulistonning bo'r yumshoq yumshoq qobiqli toshbaqalari (Trionychidae): yangi xilma-xillik, yozuvlar va qayta ko'rib chiqish". Tizimli paleontologiya jurnali. 12 (7): 799–832. doi:10.1080/14772019.2013.847870. S2CID 86304259.
- ^ a b Li, Y. M .; Li, H. J.; Kobayashi, Y .; Carabajal, A. P.; Barsbold, R .; Fiorillo, A. R.; Tsogtbaatar, K. (2019). "Mo'g'ulistonning yuqori bo'r davridagi Bayshirshir shakllanishidan krokodliform yo'lida saqlanib qolgan noan'anaviy lokomotiv harakati va uning paleobiologik oqibatlari". Paleogeografiya, paleoklimatologiya, paleoekologiya. 533 (109353): 2. Bibcode:2019PPP ... 533j9239L. doi:10.1016 / j.palaeo.2019.109239.
- ^ Averianov, A .; Sues, H. (2012). "O'rta va Markaziy Osiyoda so'nggi bo'r davrining kontinental umurtqali birikmalarining o'zaro bog'liqligi" (PDF). Stratigrafiya jurnali. 36 (2): 462−485. S2CID 54210424.
- ^ Guo, Z. X.; Shi, Y. P .; Yang, Y. T .; Tszyan, S. Q .; Li, L. B.; Zhao, Z. G. (2018). "Oxirgi bo'r davridagi Erlian havzasining teskari tomoni (Xitoy NE): Oxotomorsk blokining Sharqiy Osiyo bilan to'qnashuvining oqibatlari" (PDF). Osiyo Yer fanlari jurnali. 154: 49−66. Bibcode:2018JAESc.154 ... 49G. doi:10.1016 / j.jseaes.2017.12.007.
- ^ Tsogtbaatar, K .; Vayshampel, D. B .; Evans, D.C .; Watabe, M. (2019). "Gobi cho'lining (Bo'g'olistonning) so'nggi bo'r davridagi Baynshir shakllanishidan yangi hadrosauroid (Dinosauria: Ornithopoda)". PLOS ONE. 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. doi:10.1371 / journal.pone.0208480. PMC 6469754. PMID 30995236.
- ^ a b Park, J. Y .; Li, Y. N .; Currie, P. J.; Kobayashi, Y .; Koppelxus, E .; Barsbold, R .; Mateus, O .; Li, S .; Kim, S. H. (2020). "Talarurus plicatospineus (Dinosauria: Ankylosauridae) ning qo'shimcha bosh suyaklari va zirhli dinozavrlarning paleobiogeografiyasi va paleoekologiyasi". Bo'r davridagi tadqiqotlar. 108: 104340. doi:10.1016 / j.cretres.2019.104340.
- ^ Pauers, M. A .; Sallivan, C .; Currie, J. J. (2020). "Eudromaeosauria (Dinosauria: Theropoda) tarkibidagi preaksillyar va maksiller belgilarni qayta ko'rib chiqish: Osiyo va Shimoliy Amerika taksonlari orasidagi burun morfologiyasining turlicha tendentsiyalari". Paleogeografiya, paleoklimatologiya, paleoekologiya. 547 (109704): 109704. Bibcode:2020PPP ... 547j9704P. doi:10.1016 / j.palaeo.2020.109704.
- ^ a b Zanno, L. E .; Tsogtbaatar, K .; Chinzorig, T .; Geyts, T. A. (2016). "Segnosaurus galbinensis (Theropoda: Therizinosauria) mandibular anatomiyasi va tish protezining ixtisosliklari". PeerJ. 4: e1885. doi:10.7717 / peerj.1885. PMC 4824891. PMID 27069815.
- ^ Pol, G. S. (2016). Dinozavrlar uchun Princeton Field Guide (2-nashr). Princeton, Nyu-Jersi: Princeton University Press. 223-224-betlar. ISBN 9780691167664.
- ^ Slowiak, J .; Shcgigielski, T.; Ginter, M .; Fostovich-Frelik, L. (2020). "Qutbsiz hadozavrning uzluksiz o'sishi o'rdak dukkakli dinozavrlarning gigantizmini tushuntiradi". Paleontologiya. 63 (4): 579–599. doi:10.1111 / pala.12473.
- ^ Vatabe, M.; Suzuki, S. (2000). "1993 yil Gobi cho'liga Yaponiya-Mo'g'uliston qo'shma paleontologik ekspeditsiyasi to'g'risida hisobot". Hayashibara tabiiy fanlar muzeyi tadqiqot byulleteni. 1: 19−29.
- ^ Vatabe, M.; Suzuki, S. (2000). "Gobi cho'liga Yaponiya-Mo'g'uliston qo'shma paleontologik ekspeditsiyasi to'g'risida hisobot, 1994 y." Hayashibara tabiiy fanlar muzeyi tadqiqot byulleteni. 1: 30−44.
- ^ Kobayashi, Y .; Barsbold, R. (2006). "Mo'g'uliston Nemegt shakllanishidan kelib chiqqan ornitomimidlar" (PDF). Koreya paleontologik jamiyati jurnali. 22 (1): 195−207.
Tashqi havolalar
- Phenome10k-da 3D original bosh suyagi
- Phenome10k-da 3D rekonstruksiya qilingan bosh suyagi
- Youtube-da o'rnatilgan holotip













