Megafauna - Megafauna

Quruqlikda zoologiya, megafauna (dan.) Yunoncha mέγaς megas "katta" va Yangi lotin fauna "hayvonlar hayoti") hudud, yashash joyi yoki geologik davrning yirik yoki ulkan hayvonlarini o'z ichiga oladi. 40 kilogrammdan (90 funt) yuqori vazn ishlatiladi[1] yoki 44 kilogramm (100 funt)[2][3] (ya'ni, a bilan taqqoslanadigan yoki kattaroq massaga ega inson ) yoki a tonna, 1000 kilogramm (2,205 lb)[1][4][5] (ya'ni, ga o'xshash yoki undan kattaroq massaga ega bo'lish ho'kiz ). Ulardan birinchisiga juda ko'p deb o'ylanmagan ko'plab turlar kiradi, masalan oq dumli kiyik va qizil kenguru.
Amalda, akademik va ommabop yozuvlarda uchraydigan eng keng tarqalgan qo'llanma (faqat) uy sharoitida bo'lmagan odamnikidan kattaroq quruq sutemizuvchilarni tasvirlaydi. Bu atama ayniqsa bilan bog'liq Pleistotsen megafauna - quruqlikdagi hayvonlar ko'pincha arxetipik deb hisoblangan zamonaviy o'xshashlaridan kattaroq oxirgi muzlik davri, kabi mamontlar, ularning aksariyati shimoliy Evrosiyoda, Amerika va Avstraliya so'nggi qirq ming yil ichida yo'q bo'lib ketdi.[6] Tirik hayvonlar orasida megafauna atamasi eng katta uchun eng ko'p ishlatiladi mavjud quruqlikdagi sutemizuvchilar fillar, jirafalar, begemot, karkidon va katta sigirlar. Ushbu beshta toifadagi yirik o'txo'rlar orasida sigirlar hozirgi paytda tashqarida topilgan Afrika va janubiy Osiyo, ammo qolganlarning barchasi ilgari kengroq bo'lgan. Megafaunal turlarini parhez turiga qarab ajratish mumkin: megaherbivores (masalan, fillar ), megakarnivorlar (masalan, sherlar ) va kamdan-kam hollarda megaomnivorlar (masalan, ayiqlar ).
Boshqa keng tarqalgan foydalanish ulkan suv turlari uchun, ayniqsa kitlar, kattaroq yovvoyi yoki uy sharoitidagi quruq hayvonlarning har qanday turi antilop va qoramol, shu qatorda; shu bilan birga dinozavrlar va boshqa yo'q qilingan ulkan sudralib yuruvchilar.
Megafauna atamasi umurtqasizlarni ta'riflash uchun juda kamdan-kam qo'llaniladi, ammo u ba'zan yo'q bo'lib ketgan umurtqasiz hayvonlarning ba'zi turlari uchun ishlatilgan, ular bugungi kunda mavjud bo'lgan barcha shu kabi umurtqasizlarning turlaridan ancha katta bo'lgan. 1 m (3 fut) ninachilar ning Karbonli davr.
Ekologik strategiya
Megafauna - eng katta sutemizuvchilar va qushlar ma'nosida - odatda K-strateglar, uzoq umr ko'rish, aholi sonining sekin o'sishi, o'lim darajasi pastligi va (hech bo'lmaganda eng kattasi uchun) kattalarni o'ldirishga qodir tabiiy yirtqichlar kam yoki umuman yo'q.[7] Ushbu xususiyatlar, garchi bunday megafaunaga xos bo'lmagan bo'lsa ham, ularni insonga nisbatan zaiflashtiradi haddan tashqari ekspluatatsiya qisman aholining tiklanish darajasi sustligi sababli.[8][9]
Katta tana o'lchamining evolyutsiyasi
Tananing kattaroq kattaligi evolyutsiyasi bo'yicha o'tkazilgan bir kuzatuv shundaki, ko'pincha nisbatan qisqa vaqt oralig'ida kuzatiladigan tez o'sish sur'atlari ancha uzoq vaqt davomida barqaror bo'lmaydi. Vaqt o'tishi bilan sutemizuvchilarning tana massasi o'zgarishini tekshirganda, ma'lum vaqt oralig'ida maksimal o'sish 0,25 darajagacha ko'tarilgan interval uzunligi bilan miqyosi aniqlandi.[10] Bu maksimal tana hajmining o'sishi tendentsiyasi davomida, anatomik, fiziologik, ekologik, genetik va boshqa cheklovlarning paydo bo'lishini aks ettiradi, deb o'ylashadi. Tana massasining katta pasayishi, masalan, fenomen bilan bog'liq bo'lishi mumkin bo'lgan darajada tezroq o'zgarish tezligi aniqlandi ichki mitti. Yaratilish uzunligiga normalizatsiya qilinganida, tana massasining pasayishining maksimal tezligi o'n baravar o'zgarishi uchun tana massasining o'sishining maksimal tezligidan 30 baravar yuqori ekanligi aniqlandi.[10]
Quruq sutemizuvchilarda

Keyingi Bo'r-paleogen yo'q bo'lib ketish hodisasi taxminan 66 ta parranda bo'lmagan dinozavrlarni yo'q qildi Ma (million yil) oldin quruqlikdagi sutemizuvchilar bo'shliq qoldirgan ekologik joylarni egallash uchun turlicha bo'lganlarida tana kattaligi deyarli eksponent o'sishga uchragan.[11] Hodisa oldidan atigi bir necha kg dan boshlab, maksimal hajmi bir necha million yil o'tgach ~ 50 kg ga, oxirigacha esa ~ 750 kg ga yetdi. Paleotsen. Tana massasining o'sish tendentsiyasi taxminan 40 milya avval (kechqurun) tenglashgandek Eosen ), tana massasi uch darajadan oshganidan keyin fiziologik yoki ekologik cheklovlarga erishilganligini anglatadi.[11] Biroq, avlod uchun kattalashtirish darajasi nuqtai nazaridan qaralganda, eksponent o'sish paydo bo'lguncha davom etganligi aniqlanadi Indrikoterium 30 oy oldin. (Avloddan beri o'lchovlar tana massasi0.259, kattalashgan sari ishlab chiqarish vaqtining ko'payishi log massasi va vaqt chizig'ining chiziqli moslamadan pastga egilishiga olib keladi.)[10]
Megaherbivores oxir-oqibat 10 000 kg dan ortiq tana massasiga erishdi. Ulardan eng kattasi, indrikotirlar va probosidlar, bo'lgan orqa ichak fermentatorlari, ustunlik borligiga ishonishadi oldingi fermentlar juda katta miqdordagi oziq-ovqat mahsulotlarini qabul qilish uchun oshqozon-ichak traktini tezlashtirishi mumkinligi nuqtai nazaridan.[12] Xuddi shunday tendentsiya turli sutemizuvchilar uchun avlod uchun maksimal tana massasining o'sish sur'atlari paydo bo'lganda paydo bo'ladi qoplamalar taqqoslanadi (o'rtacha o'rtacha stavkalar yordamida makroevolyutsion vaqt o'lchovlari). Quruq sutemizuvchilar orasida eng tez o'sish sur'atlari tana massasi0.259 vaqt (ma) yilda sodir bo'lgan perissodaktillar (2,1 nishab), so'ngra kemiruvchilar (1.2) va probosidlar (1.1),[10] bularning barchasi hindgut fermentlaridir. Uchun o'sish darajasi artiodaktillar (0.74) perissodaktillarning uchdan bir qismiga teng edi. Uchun stavka yirtqichlar (0.65) biroz pastroq, ammo primatlar, ehtimol ular tomonidan cheklangan daraxt o'rganilgan sutemizuvchilar guruhlari orasida eng past ko'rsatkich (0,39) bo'lgan.[10]
Bir nechta quruqlikdagi sutemizuvchi hayvonlar evteriya guruhlar (the artiodaktil Andrewsarchus - ilgari a mezonyxid, oksyaenid Sarkastodon va yirtqichlar Amfitsion va Arktod ) barchasi maksimal 1000 kg gacha bo'lgan o'lchamga etdi[11] (yirtqich hayvon Arktoteri va xyaenodontid Simbakubva biroz kattaroq bo'lishi mumkin). Eng katta ma'lum metateryan yirtqich hayvon, Proborxyaena giganteya, aftidan, 600 kg ga etgan, bu chegaraga ham yaqin.[13] Sutemizuvchilar yirtqichlari uchun xuddi shunday nazariy maksimal kattalik sutemizuvchilarning metabolizm tezligi, o'lja olishning energetik harajati va o'ljani qabul qilishning maksimal taxminiy koeffitsienti asosida bashorat qilingan.[14] Shuningdek, sutemizuvchilarning yirtqichlari uchun maksimal kattalik stress bilan cheklangan deb taxmin qilingan humerus eng yuqori tezlikda bardosh bera oladi.[13]
Tananing maksimal kattaligining so'nggi 40 mln. Yil davomida o'zgarishini tahlil qilish shuni ko'rsatadiki, haroratning pasayishi va kontinental quruqlik maydonining ko'payishi tana hajmining oshishi bilan bog'liq. Avvalgi o'zaro bog'liqlik mos keladi Bergmann qoidasi,[15] bilan bog'liq bo'lishi mumkin termoregulyatsiya salqin iqlim sharoitida katta tana massasining afzalligi,[11] yirik organizmlarning oziq-ovqat ta'minotidagi mavsumiylikni engish qobiliyati,[15] yoki boshqa omillar;[15] oxirgi korrelyatsiyani assortiment va resurslarning cheklanganligi bilan izohlash mumkin.[11] Shu bilan birga, ikkita parametr bir-biriga bog'liq (muzliklarning ko'payishi bilan birga dengiz sathining pasayishi tufayli), bu maksimal hajmdagi tendentsiyalarni aniqlashni qiyinlashtiradi.[11]
Dengiz sutemizuvchilarida

Tetrapodlardan beri (birinchi sudralib yuruvchilar, keyinroq sutemizuvchilar Oxirgi Permiyadagi dengizga qaytib, ular o'pkadan foydalangan holda kislorodni samaraliroq iste'mol qilishlari sababli ular dengiz tanasining kattaligi yuqori qismida ustunlik qilishgan.[16][17] Ning ajdodlari turfa yarimaqvat bo'lganiga ishonishadi pakitsetidlar, itlardan kattaroq emas, taxminan 53 million yil oldin (Ma).[18] 40 miloddan ilgari, qarag'aylar uzunligi 20 m va undan ko'proq bo'lgan Basilosaurus, zamonaviy kitlardan ko'p jihatdan farq qiladigan va ular uchun ajdodlardan bo'lmagan cho'zinchoq, serpantinli kit. Buning ortidan, kekiklilarda katta tana kattaligi evolyutsiyasi vaqtincha to'xtab, so'ngra orqaga qaytganga o'xshaydi, ammo mavjud bo'lgan qazilma qoldiqlari cheklangan. Biroq, 31 oy avvalgi davrda (yilda Oligotsen ) hozirgi kungacha tana go'shti massasi sezilarli darajada tez o'sib bordi (o'sish tezligi) tana massasi0.259 million sutkada 3,2 faktorga teng) quruqlikdagi sutemizuvchilarning biron bir guruhi erishgan natijadan.[10] Ushbu tendentsiya barcha zamonlarning eng katta hayvonlariga, zamonaviylariga olib keldi ko'k kit. Cetaceanslarda katta tana hajmining tezroq evolyutsiyasi uchun bir necha sabablar bo'lishi mumkin. Kamroq biomexanik tana hajmining oshishi cheklovlari, tortishish kuchiga qarshi turishdan farqli o'laroq, suvda suspenziya bilan bog'liq bo'lishi mumkin va suzish harakatlari farqli o'laroq quruqlikdagi harakatlanish. Shuningdek, suvning havo bilan taqqoslaganda ko'proq issiqlik quvvati va issiqlik o'tkazuvchanligi oshishi mumkin termoregulyatsiya dengizda katta tana o'lchamining afzalligi endotermlar, ammo kamayib borayotgan daromadlar amal qiladi.[10]
Tishli kitlar orasida tananing maksimal hajmi oziq-ovqat mahsuloti bilan cheklangan ko'rinadi. Kabi kattaroq kattalik sperma va tumshuq kitlar, raqobatbardosh bo'lmagan muhitda nisbatan oson tutiladigan, yirik sefalopod o'ljasiga kirish uchun chuqurroq sho'ng'ishni osonlashtiradi. Odontotsetlarga nisbatan balin kitlarining samaradorligi filtr bilan oziqlantirish Planktonik oziq-ovqat zich bo'lganda kattalashib borishi bilan tarozilar yanada qulayroq bo'lib, kattaroq hajm yanada foydali bo'ladi. The o'pkada ovqatlantirish texnikasi rorquals ga qaraganda ancha tejamkor ko'rinadi qo'chqorni boqish ning balaenid kitlar; oxirgi texnika kamroq zich va yamoq plankton bilan ishlatiladi.[19] Yerning yaqin tarixidagi sovutish tendentsiyasi shamol yordamida ko'plab planktonlarning ko'p joylarini yaratgan bo'lishi mumkin uy-joylar, ulkan kitlarning rivojlanishiga yordam beradi.[19]
Ketaceanslar dengiz sutemizuvchilarining ulkan hajmiga erishgan yagona emas. Eng kattasi yirtqichlar hamma vaqt dengizdir pinnipeds, ularning eng kattasi janubiy fil muhri uzunligi 6 metrga, vazni 5000 kilogrammgacha (11000 funt) yetishi mumkin. Boshqa yirik pinnipedlarga quyidagilar kiradi shimoliy fil muhri 4000 kilogrammda (8,800 funt), morj 2000 kilogrammda (4,400 funt) va Steller dengiz sher 1,135 kilogrammda (2,502 funt). The sireniyalar dengiz sutemizuvchilarning yana bir guruhi bo'lib, ular qarag'aylar yasagan davrda to'liq suv hayotiga moslashgan. Sireniyaliklar fillar bilan chambarchas bog'liq. Eng katta sireniya bu edi Stellerning dengiz sigiri uzunligi 10 metrgacha etgan va og'irligi 8000 dan 10.000 kilogrammgacha (18000 dan 22000 funtgacha) bo'lgan va 18-asrda yo'q bo'lib ketishga ovlangan. Yarim suvli begemot quruqlikdagi sutemizuvchi hayvon bo'lib, u eng katta qarag'aylar bilan chambarchas bog'liq bo'lib, 3200 kilogrammga (7100 funt) erishishi mumkin.
Uchib ketmaydigan qushlarda

Qush bo'lmagan dinozavrlarning yo'q bo'lib ketishiga olib keladigan barcha sutemizuvchilarning boshlang'ich kattaligi kichik bo'lganligi sababli, sutemizuvchi bo'lmagan umurtqali hayvonlar gigantizm evolyutsiyasi uchun taxminan o'n million yillik imkoniyatga ega edi (paleosen davrida) juda ko'p raqobatlashmasdan.[20] Ushbu oraliqda, tepalik yirtqichi Martlar ko'pincha sudralib yuruvchilar tomonidan ishg'ol qilingan, masalan, quruqlik timsohlar (masalan, Pristichampsus ), katta ilonlar (masalan: Titanoboa ) yoki varanid kaltakesaklar yoki parvoz qilmaydigan qushlar tomonidan[11] (masalan, Paleopsilopterus (Janubiy Amerikada). Bu megafaunalning uchmaydigan o'tqayadigan davri gastornitid qushlar Shimoliy yarim sharda rivojlanib, parvozsiz paleognatlar katta hajmgacha rivojlandi Gondvanan er massalari va Evropa. Gastornitidlar va hech bo'lmaganda uchib ketmaydigan paleognat qushlarining nasl-nasabi Evropada paydo bo'lgan, ikkala nasl ham yirik o'txo'rlar uchun joylarni egallagan, sutemizuvchilar 45 kg dan past bo'lgan (bu kabi boshqa quruqliklardan farqli o'laroq). Shimoliy Amerika va Osiyo, ilgari yirik sutemizuvchilar evolyutsiyasini ko'rgan) va ulardagi eng yirik Evropa tetrapodlari bo'lgan Paleotsen.[21]
Parvozsiz paleognatlar ratitlar, an'anaviy ravishda o'zlarining uchib ketgan kichik qarindoshlari, Neotropik qalamli. Biroq, yaqinda o'tkazilgan genetik tadqiqotlar shuni ko'rsatdiki, kalamush daraxtlari qalamchali daraxt ichida joylashgan va opa-singillar guruhi yo'q bo'lib ketgan moa Yangi Zelandiya.[20][22][23] Xuddi shunday, kichik kivi Yangi Zelandiyaning yo'q bo'lib ketgan singil guruhi ekanligi aniqlandi fil qushlari Madagaskar.[20] Ushbu topilmalar shuni ko'rsatadiki parvozsizlik va gigantizm bir necha bor mustaqil ravishda paydo bo'ldi parallel evolyutsiya.
Yirtqich megafaunal uchmaydigan qushlar ko'pincha sutemizuvchilar bilan erta raqobatlasha olishgan Kaynozoy. Keyinchalik, kaynozoyda, ular rivojlangan yirtqichlar tomonidan ko'chirilib, vafot etishdi. Shimoliy Amerikada bathornitidlar Parakrax va Bathornis tepalik yirtqichlari bo'lgan, ammo tomonidan yo'q bo'lib ketgan Ilk miosen. Janubiy Amerikada, tegishli fosuratsidlar metatherian bilan dominant yirtqich joylarni bo'lishdi sparassodontlar Senozoyning aksariyat davrida, ammo Shimoliy Amerikadan evteriya yirtqichlari kelganidan keyin pasayib ketdi va oxir-oqibat yo'q bo'lib ketdi ( Buyuk Amerika almashinuvi ) davomida Plyotsen. Buning aksincha, katta o'txo'r parvozsiz ratitlar hozirgi kungacha saqlanib qolgan.
Biroq, kaynozoyikning uchib ketmaydigan qushlarining hech biri, shu jumladan yirtqichlar Brontornis, ehtimol omnivorous Dromornis[24] yoki o'txo'rlar Vorombe, har doim 500 kg dan kattaroq massaga o'sgan va shu bilan hech qachon sutemizuvchilarning eng katta yirtqich hayvonlari kattaligiga, hatto eng katta sutemizuvchi o'txo'rlarnikiga teng bo'lmagan. Qushlarning tuxum qobig'ining qalinligi tuxum massasi bilan mutanosib ravishda ko'payib borayotganligi, qushlarning kattaligiga yuqori chegarani beradi, degan fikrlar mavjud.[25][eslatma 1] Eng katta turlari Dromornis, D. stirtoni, maksimal parranda tana massasiga erishgandan keyin yo'q bo'lib ketgan va keyinchalik marsupial tomonidan ustun bo'lgan diprotodontlar bu bir necha baravar kattalikka aylandi.[28]
Katta toshbaqalarda
Gigant toshbaqalar kechning muhim tarkibiy qismlari edi Kaynozoy megafaunalar, har bir qutbsiz qit'ada kelguniga qadar mavjud gominanlar.[29][30] Ma'lum bo'lgan eng katta quruq toshbaqa edi Megalokelis atlasi, ehtimol 1000 kg vaznga ega bo'lgan hayvon.
Ba'zi avvalgi suvda yashovchi testudinlar, masalan. dengiz Archelon bo'r va chuchuk suvlar Stupendemys Miosenning vazni 2000 kg dan ortiq bo'lgan.
Megafaunalning yo'q bo'lib ketishi
Vaqt va mumkin bo'lgan sabablar
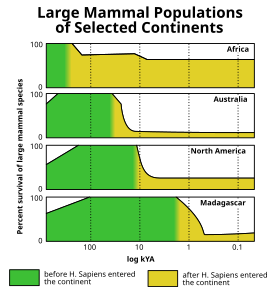
The Golotsenning yo'q bo'lib ketishi (Shuningdek qarang To'rtlamchi davrda yo'q bo'lib ketish hodisasi ) oxirida sodir bo'lgan oxirgi muzlik davri muzlik davri (a.k.a. Würm muzligi kabi ko'plab yirik muzlik davri sutemizuvchilar, qachon junli mamontlar, ichida yo'q bo'lib ketdi Amerika va shimoliy Evroosiyo. Shimoliy Amerikada yo'q bo'lib ketish hodisasini tahlil qilish, bu katta hayvonlar uchun selektivligi bilan kaynozoy erlarining yo'q bo'lib ketish impulslari orasida noyob ekanligini aniqladi.[31](Shakl 10) Turli xil nazariyalar yo'q bo'lib ketish to'lqini bilan bog'liq odam ovi, Iqlim o'zgarishi, kasallik, a g'ayritabiiy ta'sir yoki boshqa sabablar. Biroq, oxirigacha bu yo'q bo'lib ketish Pleystotsen so'nggi 50.000 yil davomida Yer yuzining ko'p qismida sodir bo'lgan megafaunal yo'qolib ketish impulslaridan biri edi. Afrika va janubiy Osiyo (bu erda mahalliy megafauna zamonaviy odamlar bilan birgalikda rivojlanish imkoniyatiga ega edi) nisbatan kamroq ta'sir ko'rsatdi. Oxirgi joylar megafaunani, xususan, sekinroq harakatlanadigan turlarni (asta-sekin zaiflashgan megafauna sinfini) asta-sekin yo'q qilish bilan qiynashgan. ulkan toshbaqalar ), so'nggi bir necha million yil ichida.[32][33]
Materikdan tashqarida Afro-Evroosiyo, ushbu megafaunal yo'q bo'lib ketish, odamlarning dunyoning ilgari yashamagan hududlariga tarqalishini chambarchas taqqoslaydigan va iqlim tarixi bilan umuman bog'liqligini ko'rsatmaydigan (so'nggi geologik vaqt oralig'idagi uchastkalar bilan tasavvur qilish mumkin bo'lgan) juda o'ziga xos quruqlik-quruqlik naqshini kuzatib bordi. kabi iqlim belgilarining dengiz kislorod izotoplari yoki atmosferadagi karbonat angidrid darajasi ).[34][35] Avstraliya[36] va yaqin atrofdagi orollar (masalan, Flores[37]) avval 46000 yil oldin urilgan, keyin esa Tasmaniya taxminan 41,000 yil oldin (taxminan 43,000 yil oldin Avstraliyaga quruqlik ko'prigi tashkil etilgandan so'ng),[38][39][40] Yaponiya taxminan 30,000 yil oldin,[41] Shimoliy Amerika 13000 yil oldin,[2-eslatma] Janubiy Amerika taxminan 500 yil o'tgach,[43][44] Kipr 10000 yil oldin,[45][46] The Antil orollari 6000 yil oldin,[47][48] Yangi Kaledoniya[49] va yaqin atrofdagi orollar[50] 3000 yil oldin, Madagaskar 2000 yil oldin,[51] Yangi Zelandiya 700 yil oldin,[52] The Maskarenlar 400 yil oldin,[53] va Qo'mondon orollari 250 yil oldin.[54] Dunyoning deyarli barcha ajratilgan orollari qirg'in paydo bo'lganidan keyin qisqa vaqt ichida sodir bo'ladigan o'xshash misollarni keltirishi mumkin edi odamlar, garchi bu orollarning aksariyati, masalan Gavayi orollari, hech qachon quruqlikdagi megafauna bo'lmagan, shuning uchun ularning yo'q bo'lib ketgan hayvonot dunyosi kichikroq edi.[34][35]
Vaqtni tahlil qilish Holarktika So'nggi 56000 yil ichida megafaunallarning yo'q bo'lib ketishi va yo'q bo'lib ketishi bunday hodisalar ichida to'planish tendentsiyasini ko'rsatdi. interstadiallar, keskin isish davri, lekin faqat odamlar mavjud bo'lganda. Odamlar migratsiya va rekolonizatsiya jarayonlariga to'sqinlik qilgan bo'lishi mumkin, aks holda megafaunal turlarining iqlim o'zgarishiga moslashishi mumkin edi.[55] Hech bo'lmaganda ba'zi sohalarda, interstadiallar inson populyatsiyasining kengayish davri bo'lgan.[56]
Tahlil Sporormiella so'ngi 130 ming yilni tashkil etgan botqoq cho'kindi yadrolarida qo'ziqorin sporu (asosan megaherbivorlarning tezidan kelib chiqadi). Linch krateri yilda Kvinslend, Avstraliya shuni ko'rsatdiki, ushbu mintaqadagi megafauna deyarli 41 ming yil oldin, bir vaqtning o'zida yo'q bo'lib ketgan iqlim o'zgarishi minimal edi; o'zgarish ko'mirning ko'payishi bilan birga o'tdi va undan keyin yomg'ir o'rmonidan yong'inga chidamli holatga o'tdi sklerofil o'simlik. O'zgarishlarning yuqori aniqlikdagi xronologiyasi odam ovining o'zi megafaunani yo'q qildi va keyinchalik floradagi o'zgarish brauzerlarning yo'q qilinishi va yong'inning ko'payishi natijasida yuzaga kelgan degan farazni qo'llab-quvvatlaydi.[57][58][59][60] Yong'inning ko'payishi megafaunaning yo'q bo'lib ketishiga bir asrga yaqin vaqt qoldi va, ehtimol, ko'rish to'xtatilgandan so'ng yoqilg'ining to'planishi natijasida yuzaga keldi. Keyingi bir necha asrlarda o't o'sdi; sklerofill o'simliklari yana bir asr kechikishi bilan ko'paygan va sklerofil o'rmoni yana ming yildan so'ng rivojlangan.[59] Taxminan 120,000 va 75,000 yil oldin ob-havoning o'zgargan ikki davrida, bu erda yanada sovuq va quruq sharoitga o'tishga javoban sklerofill o'simliklari ko'paygan; ushbu epizodlarning ikkalasi ham megafaunal ko'pligiga sezilarli ta'sir ko'rsatmadi.[59] Pleistosen megafaunasining yo'q bo'lib ketishida odam ovchilarining aybdorligi to'g'risida shunga o'xshash xulosalar uchib ketmaydigan avstraliyalik qushning katta miqdordagi tuxum qobig'i parchalarini tahlil qilish natijasida olingan yuqori aniqlikdagi xronologiyalardan kelib chiqqan. Genyornis newtoni,[61][62][60] ning tahlilidan Sporormiella sharqiy Shimoliy Amerikadagi ko'ldan qo'ziqorin sporu[63][64] konlarini o'rganishdan Shasta er yalqovi Amerikaning janubi-g'arbiy qismida yarim o'ndan ziyod g'orda qolgan go'ng.[65][66]
Odamlarning davom etayotgan ovi va atrof-muhit buzilishi qo'shimcha narsalarga olib keldi yaqin o'tmishda megafaunallarning yo'q bo'lib ketishi va yaratdi keyingi yo'q bo'lib ketishning jiddiy xavfi yaqin kelajakda (quyida keltirilgan misollarga qarang). Odamlar tomonidan, birinchi navbatda, go'sht uchun to'g'ridan-to'g'ri o'ldirish, zamonaviy megafaunal tanazzulning eng muhim omilidir.[67][68]
Boshqa bir qator ommaviy qirilib ketish Erning geologik tarixida ilgari sodir bo'lgan bo'lib, unda o'sha paytdagi megafaunaning bir qismi yoki hammasi yo'q bo'lib ketgan. Mashhur, ichida Bo'r-paleogen yo'q bo'lib ketish hodisasi qush bo'lmagan dinozavrlar va boshqa ko'plab yirik sudralib yuruvchilar yo'q qilindi. Biroq, avvalgi ommaviy qirg'inlar global bo'lgan va megafauna uchun unchalik tanlanmagan; ya'ni boshqa turdagi ko'plab turlar, shu jumladan o'simliklar, dengiz umurtqasizlar[69] va plankton ham yo'q bo'lib ketdi. Shunday qilib, avvalgi voqealar, umumiy holatdagi buzilishlar tufayli yuzaga kelgan bo'lishi kerak biosfera.
Megafaunani yo'q qilish oqibatlari
Oziq moddalarni tashishda ta'siri
Megafauna ekologik tizimda mineral ozuqa moddalarini lateral tashishda muhim rol o'ynaydi, ularni yuqori joylardan past darajadagi joylarga ko'chirishga intiladi. Ular buni ozuqa moddalarini iste'mol qilish vaqti va uni yo'q qilish yo'li bilan (yoki o'limdan keyin parchalanish orqali) ajratish vaqti orasidagi harakati bilan amalga oshiradilar.[70] Janubiy Amerikada Amazon havzasi Taxminan 12,500 yil oldin sodir bo'lgan megafaunal yo'q bo'lib ketgandan so'ng, bunday lateral diffuziya 98% dan kamaydi.[71][72] Sharti bilan; inobatga olgan holda fosfor mavjudlik mintaqaning aksariyat qismida hosildorlikni cheklaydi, uning havzaning g'arbiy qismidan va toshqin joylardan transportining pasayishiga olib keladi (ikkalasi ham ta'minotni ko'tarilishidan kelib chiqadi) And ) boshqa hududlarga mintaqaning ekologiyasiga sezilarli ta'sir ko'rsatgan deb o'ylashadi va ta'sir hali o'z chegaralariga etib bormagan bo'lishi mumkin.[72] Dengizda chuqurlikda oziqlanadigan turshak va pinnipedlar azotni chuqurlikdan sayoz suvga o'tkazib, kuchaytiradi deb o'ylashadi. okean unumdorligi va faoliyatiga qarshi turish zooplankton, aksincha qilishga moyil bo'lgan.[73]
Metan chiqindilariga ta'siri
Megaherbivores katta populyatsiyalari atmosfera kontsentratsiyasiga katta hissa qo'shishi mumkin metan, bu muhim issiqxona gazi. Zamonaviy kavsh qaytaruvchi hayvon o'txo'rlar metanni yon mahsulot sifatida ishlab chiqaradi oldingi fermentatsiya ovqat hazm qilishda va uni qorayish yoki meteorizm orqali chiqaring. Bugungi kunda yillik 20% atrofida metan chiqindilari chorva mollari metanining chiqarilishidan kelib chiqadi. In Mezozoy, deb taxmin qilingan sauropodlar atmosferaga yiliga 520 million tonna metan chiqarishi mumkin edi,[74] vaqtning iliq iqlimiga hissa qo'shadi (hozirgi zamonga qaraganda 10 ° S gacha issiq).[74][75] Ushbu katta emissiya sauropodlarning ulkan taxmin qilingan biomassasidan kelib chiqadi va alohida o'txo'r hayvonlarning metan ishlab chiqarilishi ularning massasiga deyarli mutanosib ekanligiga ishoniladi.[74]
Yaqinda o'tkazilgan tadqiqotlar shuni ko'rsatdiki, megafaunal o't o'simliklari yo'q bo'lib ketishi kamayishiga olib kelishi mumkin atmosferadagi metan. Ushbu gipoteza nisbatan yangi.[76] Bir tadqiqot natijasida metan chiqindilari tekshirildi bizon egallagan Buyuk tekisliklar Evropa ko'chmanchilari bilan aloqa qilishdan oldin Shimoliy Amerika. Tadqiqot natijalariga ko'ra bizonni olib tashlash yiliga 2,2 million tonnagacha pasayishiga olib keldi.[77] Boshqa bir tadqiqot oxirida atmosferadagi metan kontsentratsiyasining o'zgarishini o'rganib chiqdi Pleystotsen davri Amerikada megafauna yo'q bo'lib ketganidan keyin. Dastlabki odamlar Amerikaga ko'chib o'tgandan keyin taxminan 13000 kishi BP, ularning ovlanishi va boshqa tegishli ekologik ta'sirlar u erda ko'plab megafaunal turlarining yo'q bo'lib ketishiga olib keldi. Hisob-kitoblarga ko'ra, ushbu yo'q bo'lib ketish metan ishlab chiqarishni yiliga 9,6 million tonnaga kamaytirgan. Bu megafaunal metan chiqindilarining yo'qligi, iqlimning boshlanishida keskin iqlimiy sovutishga yordam bergan bo'lishi mumkin. Yosh Dryas.[76] O'sha paytda sodir bo'lgan atmosfera metanining pasayishi, qayd etilgan muz tomirlari, so'nggi yarim million yillikdagi boshqa pasayishdan 2-4 baravar tezroq bo'lib, g'ayrioddiy mexanizm ishlayotganligini ko'rsatdi.[76]
Misollar
Quyida ko'pincha megafauna deb qaraladigan hayvonlarning ba'zi bir muhim misollari keltirilgan ("yirik hayvon" ta'rifi ma'nosida). Ushbu ro'yxat to'liq bo'lishi kerak emas:
- Klade Sinapsida
- Sinf Sutemizuvchilar (filogenetik jihatdan, a qoplama Therapsida ichida; pastga qarang)
- Infraklass Metateriya
- Buyurtma Diprotodontiya
- The qizil kenguru (Macropus rufus) eng katta hayot Avstraliyalik sutemizuvchi va marsupial 85 kg gacha vaznda (187 funt). Biroq, uning yo'q bo'lib ketgan qarindoshi kalta yuzli ulkan kenguru Prokoptodon goliah 230 kg (510 lb) ga yetdi,[78] yo'q bo'lib ketganda diprotodontlar tarixdagi har qanday marsupialning eng katta hajmiga, taxminan 2750 kg (6,060 funt) ga erishdi. Yo'qolib ketgan marsupial sher (Tilakoleo chinnigullari ), 160 kg gacha bo'lgan vaznda (350 lb) har qanday mavjud bo'lgan yirtqich marsupialdan ancha katta edi.
- Buyurtma Diprotodontiya
- Infraklass Evteriya
- Superorder Afroteriya
- Buyurtma Proboscidea
- Fillar quruqlikdagi eng yirik tirik hayvonlardir. Ular va ularning qarindoshlari paydo bo'lishdi Afrika, ammo yaqin vaqtgacha deyarli butun dunyo bo'ylab tarqatilgan. The Afrikalik buta fili (Loxodonta africana) elkasining balandligi 4,3 m (14 fut) gacha va og'irligi 10,4 tonnagacha (11,5 qisqa tonna).[79] Yaqinda yo'q bo'lib ketgan proboskidlar orasida, mamontlar (Mammut) fillarning yaqin qarindoshlari bo'lgan, ammo mastodonlar (Mammut) bir-biridan ancha uzoqda bo'lgan. The dasht mamonti (M. trogontherii) taxminan 10 tonnani tashkil etgani taxmin qilinmoqda, bu uni eng katta hajmga aylantiradi probosid bu quruqlikdagi sutemizuvchilardan keyin ikkinchi o'rinni egallaydi indrikoterinlar.
- Buyurtma Sireniya
- 1500 kg (3300 lb) gacha bo'lgan eng katta sireniya bu G'arbiy Hindiston manati (Trichechus manatus). Stellerning dengiz sigiri (Hydrodamalis gigas) taxminan besh baravar katta bo'lgan, ammo uzoq masofadan topilganidan keyin 27 yil ichida odamlar tomonidan yo'q qilingan Qo'mondon orollari 1741 yilda. Tarixdan oldin bu dengiz sigiri shimoli-sharqiy sohillarda ham yashagan Osiyo va shimoli-g'arbiy Shimoliy Amerika; aftidan, mahalliy ovchilar bu qulayroq joylardan yo'q qilishgan.
- Buyurtma Proboscidea
- Superorder Ksenartra
- Buyurtma Cingulata
- The glyptodontlar katta, og'ir zirhli guruh edi ankilozavr o'xshash xenarthrans yashash bilan bog'liq armadillos. Ular kelib chiqishi Janubiy Amerika paytida Shimoliy Amerikani bosib oldi Buyuk Amerika almashinuvi va oxirida yo'q bo'lib ketdi Pleystotsen davr.[80]
- Buyurtma Pilosa
- Tuproqdagi yalqovlar zamonaviy bilan bog'liq bo'lgan sekin, quruqlikdagi ksenartranslarning yana bir guruhi edi daraxt yalqovlari. Ular xuddi shunday tarixga ega edilar, garchi ular Shimoliy Amerikaga oldinroq etib kelishgan va shimolga tarqalishgan (masalan, Megaloniks ). Eng katta nasl, Megatherium va Eremoteliy, fillar bilan taqqoslanadigan o'lchamlarga etdi.[80]
- Buyurtma Cingulata
- Superorder Evarxontoglires
- Buyurtma Primatlar
- 266 kg (586 lb) gacha bo'lgan eng katta tirik primat bu gorilla (Gorilla beringei va Gorilla gorilla, to'rtta pastki uchtadan uchtasi mavjud juda xavfli ). Yo'qolib ketgan Malagasiya yalqov lemur Arxeoindris yo'q bo'lib ketganda, xuddi shunday hajmga erishdi Gigantopitekus qora ning Janubi-sharqiy Osiyo hali kattaroq deb hisoblanmoqda, garchi dastlabki taxminlardan farqli o'laroq, ehtimol ikki baravar kam (postkranial qoldiqlarning yo'qligi uning hajmini baholashni qiyinlashtiradi).[81] Ba'zi populyatsiyalar arxaik Homo so'nggi paytlarga qaraganda o'rtacha darajada katta edi Homo sapiens;[82][83] masalan, Homo heidelbergensis Afrikaning janubida odatda 2,1 m balandlikda bo'lishi mumkin edi,[84] esa Neandertallar taxminan 30% ko'proq edi.[85]
- Buyurtma Rodentiya
- Mavjud kapyara (Hydrochoerus hydrochaeris) eng yirik yashovchi kemiruvchi Janubiy Amerikaning vazni 80 kg (180 lb) gacha.[86] Yaqinda yo'q bo'lib ketgan Shimoliy Amerikaning bir nechta shakllari kattaroq edi: kapibara Neoxerus pinckneyi (boshqa Neotropik migrant) o'rtacha 40% og'irroq edi; The ulkan qunduz (Kastoroidlar ohioensis) o'xshash edi. Yo'qolib ketgan to'mtoq tishli ulkan xutiya (Amblyrhiza inundata) bir nechta Karib dengizi orollar hali ham kattaroq bo'lishi mumkin edi. Biroq, bir necha million yil oldin Janubiy Amerikada juda katta miqdordagi kemiruvchilar bor edi. Foberomis pattersoni deyarli to'liq skeletdan ma'lum bo'lgan, ehtimol 700 kg (1500 funt) ga etgan. Parchalanib ketgan qoldiqlar shuni ko'rsatmoqda Josephoartigasia monesi 1000 kg (2200 funt) gacha ko'tarildi.
- Buyurtma Primatlar
- Superorder Laurasiatheria
- Buyurtma Yirtqich hayvon
- Hozirgacha mavjud bo'lgan eng katta mushuklar turkumga kiradi Panthera shu jumladan yo'lbars (P. tigris) va sher (P. leo).[87] The Sibir yo'lbarsi (P. t. Altayka) ko'ra eng katta yovvoyi mushuk bo'lishi kerak Bergmann qoidasi, va ba'zilar buni shunday deb hisoblashgan[88][89] ammo bu bahsli.[90] Tarixiy jihatdan yovvoyi Sibir yo'lbarslari hajmi kamaygan va ular hozir Bengal yo'lbarslaridan kichikroq (P. t. Dajla);[91] ammo, Sibir yo'lbarslari hali ham tutqunlikda eng katta yo'lbars bo'lib, vazni 320 kg (710 lb) ga etadi.[92] Panthera turlari bilan ajralib turadi morfologik ularni shovullashga imkon beruvchi xususiyatlar. Yirik qirilib ketgan mushuklarga quyidagilar kiradi Amerika sher (P. atrox) va Janubiy Amerika qichitqi tishli mushuk (Smilodon populyatori ).
- Ayiqlar ning katta yirtqichlaridir kaniform suborder. Eng katta tirik shakllar oq ayiq (Ursus maritimus), tana og'irligi 800 kg (1800 lb) gacha,[93] va deyarli katta Kodiak ayig'i (Ursus arctos middendorffi),[94] Bergmann qoidasiga mos keladi. Arktoteri avgustliklar, yo'q bo'lib ketgan kalta yuzli ayiq Janubiy Amerikadan, bu dunyodagi eng yirik yirtqich sutemizuvchi bo'lib, o'rtacha og'irligi 1600 kg (3500 lb) bo'lgan.[95]
- Muhrlar, dengiz sherlari va morjlar ayiqsimon ajdodlardan kelib chiqqan amfibiya dengiz yirtqichlari. The janubiy fil muhri (Mirounga leonina) ning Antarktika va subantarktika suvlar barcha zamonlardagi eng yirik yirtqich hisoblanadi, buqalar erkaklari maksimal uzunligi 6-7 m (20-23 fut) va maksimal vazni 5000 kg (11000 funt) ga etadi.
- Buyurtma Perissodaktila
- Tapirlar 20 million yil ichida deyarli o'zgarmagan ko'rinadigan qisqa cho'zinchoq tumshug'i va cho'chqaga o'xshash shakli bo'lgan hayvonlarni ko'rib chiqmoqdalar. Ular yashaydilar tropik o'rmonlar Janubi-Sharqiy Osiyo va Janubiy va Markaziy Amerika, hamda so'nggi ikki mintaqaning omon qolgan eng yirik quruqlik hayvonlarini o'z ichiga oladi. To'rt tur mavjud.
- Karkidon bor toq oyoqli tuyoqlilar yasalgan shoxlari bilan keratin, xuddi shu turdagi oqsil sochlarni yaratish Ular 850-3800 kg gacha bo'lgan tirik quruqlikdagi sutemizuvchilar orasida ikkinchi o'rinda turadi. Hozirgacha mavjud bo'lgan beshta turdan uchtasi juda xavfli. Ularning yo'q bo'lib ketishi Markaziy Osiyo qarindoshlar indrikoterinlar zamondagi eng yirik sutemizuvchilar bo'lgan.
 Karkidon, dan Dyurerning yog'och o'ymakorligi
Karkidon, dan Dyurerning yog'och o'ymakorligi
- Buyurtma Artiodactyla
- Jirafalar (Giraffa spp.) balandligi qariyb 6 m (20 fut) ga etgan eng baland yashovchilar. Voyaga etgan erkak uchun o'rtacha og'irlik 1,192 kg (2,628 lb) va kattalar ayol uchun 828 kg (1,825 lb), maksimal og'irligi 1,930 kg (4,250 lb) va erkaklar va ayollar uchun 1,180 kg (2600 lb).
- Sigir tuyoqlilar tirik qolgan eng yirik hayvonlarni o'z ichiga oladi Evropa va Shimoliy Amerika. The suvsar (Bubalis arnee), bizon (Bizon bizoni va B. bonus ) va gaur (Bos gaurus) barchasi 1000 kg dan ortiq vaznga o'sishi mumkin (2200 funt).
- The yarimakvat begemot (Hippopotamus amfibius) buyruqning eng og'ir tirik a'zosi Cetartiodactyla turg'unlardan keyin. Voyaga etganlarning o'rtacha vazni erkaklar va ayollar uchun mos ravishda 1500 kg (3300 funt) va 1300 kg (2900 funt) ni tashkil qiladi, katta erkaklar esa 3200 kg (7100 funt) dan oshadi. Begemot va juda kichikroq juda xavfli pigmiy gippo (Choeropsis liberiensis) deb ishoniladi mavjud bo'lgan eng yaqin qarindoshlar turkumlar. Gipopotamuslar megafaunal turlar qatoriga kiradi eng xavfli odamlarga.[96]
- Infraorder Keteya
- Kitlar, delfinlar va kakliklar dengiz sutemizuvchilaridir. The ko'k kit (Balaenoptera mushaklari) eng katta balin kit va hozirgacha 30 metr (98 fut) bo'lgan eng katta hayvon[97] uzunligi va 170 tonna (190 qisqa tonna)[98] yoki undan ko'p vaznda. The sperma kiti (Fizeter makrosefali) eng katta tishli kit va umurtqali hayvonlar tarixidagi eng yirik yirtqichlardan biri, shuningdek sayyoramizning eng baland va eng dahshatli hayvon (bilan miya kabi besh baravar katta odamlar ). The qotil kit (Orcinus orca) eng katta delfin.
- Buyurtma Yirtqich hayvon
- Superorder Afroteriya
- Infraklass Metateriya
- Buyurtma Pelikozauriya (an'anaviy; parafiletik )
- Kotilrinxus katta, katta tirnoqli, o'txo'r edi kazeid erta Permian Shimoliy Amerika, 6 m (20 fut) va 2 tonnaga etadi.
- Buyurtma Terapsida
- Anteosaurus edi a bosh terish, yarimakvat, yirtqich dinosefali ning O'rta perm Janubiy Afrika. Uzunligi 5-6 metrga etdi va vazni 500-600 kg (1100-1300 funt).[99]
- Lisowicia fil o'lchamidagi (9 tonna) o'txo'r edi kannemeyeriiform dicynodont Oxirgi Trias Evropasi.[100][101]
- Sinf Sutemizuvchilar (filogenetik jihatdan, a qoplama Therapsida ichida; pastga qarang)
- Klade Sauropsida
- Sinf Aves (filogenetik, qoplama Coelurosauria, a takson Saurischia buyrug'i doirasida; pastga qarang)
- Buyurtma Struthioniformes
- The ratitlar qadimiy va xilma-xil guruhdir parvoz qilmaydigan qushlar birinchisining parchalarida topilgan superkontinent Gondvana. Eng katta tirik qush tuyaqush (Struthio tuya) yo'q bo'lib ketganidan oshib ketdi Vorombe ning Madagaskar, (860 kg (1900 lb)) gacha bo'lgan guruhdagi eng og'ir va yo'q bo'lib ketgan ulkan moa (Dinornis) ning Yangi Zelandiya, balandligi 3,4 m (11 fut) gacha ko'tarilgan eng baland. Oxirgi ikkitasi misollar orol gigantizmi.
- Buyurtma Gastornithiformes
- Yo'qolib ketdi dromornitidlar kabi Avstraliyaning Dromornis kattaligi bo'yicha eng katta ratitlarga yaqinlashdi. (Qit'a uchun kichik o'lcham va izolyatsiya tufayli, Avstraliya ba'zan sifatida qaraladi dunyodagi eng katta orol; Shunday qilib, bu turlarni ham gigant deb hisoblash mumkin edi.)
- Buyurtma Katartiformes
- Yo'qolib ketgan kondor o'xshash teratorn Argentavis Janubiy Amerikaning taxmin qilingan qanot oralig'i 5 dan 6 m gacha (16 dan 20 futgacha) va massasi taxminan 70 kg (150 lb) bo'lgan va bu megafaunal uchayotgan qushning eng yaxshi namunasi bo'lgan.
- Buyurtma Struthioniformes
- Sinf Reptiliya (an'anaviy; parafiletik )
- Buyurtma Saurischia
- Saurischian dinozavrlar ning Yura davri va Bo'r o'z ichiga oladi sauropodlar, ma'lum bo'lgan eng uzun (40 m yoki 130 futgacha) va massiv quruqlikdagi hayvonlar (Argentinosaurus 80-100 ga etdimetrik tonna yoki 90-110tonna ), shu qatorda; shu bilan birga tropodlar, eng katta quruqlikdagi yirtqich hayvonlar (Spinosaurus 7-9 tonnagacha o'sdi; shuncha mashhur Tiranozavr, 6,8 tonnagacha).
- Buyurtma Pterosauriya
- Eng kattasi azdarxid pterozavrlar, kabi Hatzegopteryx va Quetzalcoatlus, taxminan 11-12 m (36-39 fut) atrofida qanotlarni ochib, og'irliklari 70-250 kg (150-550 lb) oralig'ida bo'lishi mumkin. Birinchisi uning cho'qqisi yirtqichi bo'lgan deb o'ylashadi orol ekotizimi.
- Buyurtma Timsoh
- Alligatorlar va timsohlar yirik yarimavtali sudralib yuruvchilar, ulardan eng kattasi timsoh sho'r suv (Crocodylus porosus), og'irligi 1,360 kg (3000 lb) gacha o'sishi mumkin. Timsohlarning uzoq ajdodlari va ularning qarindoshlari pseudosuchians (an'anaviy kruotarsanlar ), kech dunyoda hukmronlik qildi Trias, gacha Trias - Yura davridagi yo'q bo'lib ketish hodisasi dinozavrlarning ularni bosib olishlariga imkon berdi. Keyinchalik ular turli xil bo'lib qolishdi Mezozoy, qachon timsoh shakllari kabi Deinosuchus va Sarcosuchus uzunligi 12 m ga etgan. Xuddi shunday katta timsohlar, masalan Mourasuchus va Purussaurus, kabi yaqinda mavjud edi Miosen Janubiy Amerikada.
- Buyurtma Squamata
- Hozirgacha mavjud bo'lgan eng katta narsa kaltakesak, Komodo ajdaho (Varanus komodoensis), yana bir orol giganti, uning yo'q bo'lib ketgan avstraliyalik qarindoshi uzunligi 3 m (10 fut) ga etishi mumkin Megalaniya bu kattalikdan ikki barobardan oshgan bo'lishi mumkin. Bular kaltakesaklarni kuzatish 'dengiz qarindoshlari, mosasaurlar, edi tepalik yirtqichlari kech bo'r dengizlarida.
- Hozirgacha mavjud bo'lgan eng og'ir ilon deb hisoblanadi yashil anakonda (Eunectes murinus), esa retikulyatsiya qilingan piton (Python reticulatus), 8,7 m gacha yoki undan ko'proq, eng uzun deb hisoblanadi. Yo'qolib ketgan avstraliyalik Plyotsen turlari Liyaz, Bluff Downs gigant pitoni, 10 m ga yetdi, esa Paleotsen Titanoboa Janubiy Amerikaning uzunligi 12-15 m gacha va taxminiy og'irligi taxminan 1135 kilogrammni (2500 funt) tashkil etdi.
- Buyurtma Testudinlar
- Eng katta toshbaqa juda xavfli dengiz toshbaqa toshbaqa (Dermochelys coriacea), og'irligi 900 kg (2000 lb) gacha. U boshqasidan ajralib turadi dengiz toshbaqalari uning etishmasligi bilan suyak qobiq. Eng katta er usti chelonianlari - bu ulkan toshbaqalar ning Galapagos orollari (Chelonoidis nigra ) va Aldabra Atoll (Aldabrachelys gigantea ), 300 kg gacha (660 funt). Ushbu toshbaqalar kontinental quruqliklarda keng tarqalgan yirik toshbaqa turlarining assortimentidan omon qolganlarning eng katta qismidir.[29][30] va qo'shimcha orollar[29] pleystotsen davrida.
- Buyurtma Saurischia
- Sinf Aves (filogenetik, qoplama Coelurosauria, a takson Saurischia buyrug'i doirasida; pastga qarang)
- Sinf Amfibiya (keng, ehtimol parafiletik, ma'noda)
- Buyurtma Temnospondiliy (bilan munosabatlar mavjud amfibiyalar tushunarsiz)
- Permiy temnospondil Prionosuchus, ma'lum bo'lgan eng katta amfibiya, uzunligi 9 m ga etgan va timsohga o'xshash suv yirtqichi bo'lgan. Haqiqiy timsohlar paydo bo'lganidan keyin, masalan temnospondillar Koolasuchus (5 m uzunlikdagi) bo'r tomonidan Antarktika mintaqasiga chekingan, yo'q bo'lib ketishdan oldin.
- Buyurtma Temnospondiliy (bilan munosabatlar mavjud amfibiyalar tushunarsiz)
- Sinf Aktinopterygii
- Buyurtma Tetraodontiformes
- Hozirgi eng katta suyakli baliq bo'ladi okean quyosh baliqlari (Mola mola), kattalarning o'rtacha vazni 1000 kg (2200 funt) ni tashkil qiladi. Filogenetik jihatdan "suyakli baliq" bo'lsa-da, uning skeleti birinchi navbatda xaftaga (bu nisbatan engilroq suyak ). U disk shaklidagi tanaga ega va uzun, ingichka bilan o'zini harakatga keltiradi dorsal va anal suyaklar; u birinchi navbatda oziqlanadi meduza. Ushbu uchta jihat bo'yicha (shuningdek, uning kattaligi va sho'ng'in odatlari bo'yicha) u charm toshbaqaga o'xshaydi.
- Buyurtma Lampriformlar
- The bahaybat baliq (Regalecus glesne) eng uzun suyakli baliq bo'lib, 11 m (36 fut) ga etadi.
- Buyurtma Acipenseriformes
- The juda xavfli beluga (Evropa mersinari, Huso huso) 1476 kg gacha bo'lgan (3254 funt) eng katta hisoblanadi baliqlar (ular asosan xaftaga tushadigan) va eng kattasi hisoblanadi anadromoz baliq.
- Buyurtma Siluriformes
- The juda xavfli Mekong ulkan baliqlari (Pangasianodon gigalari), 293 kg gacha (646 lb), ko'pincha eng katta deb hisoblanadi chuchuk suv baliqlari.
- Buyurtma Tetraodontiformes
- Sinf Chondrichthyes
- Buyurtma Lamniformes
- Tirik yirtqich baliqlarning eng katta qismi katta oq akula (Carcharodon karchariaslari), 2240 kg gacha vaznga etadi (4.940 lb). Uning yo'q bo'lib ketgan qarindoshi C. megalodon (bahsli jins ham Carcharodon yoki Carcharocles) dan ko'proq edi kattalik tartibi kattaroq va bu hamma vaqtdagi eng katta yirtqich akula yoki baliq (va umurtqali hayvonlar tarixidagi eng katta yirtqichlardan biri); u kitlarga va boshqalarga o'lja edi dengiz sutemizuvchilar.
- Buyurtma Orectolobiformes
- Hozirgi eng katta nahang, xaftaga tushadigan baliqlar va baliq Umuman olganda kit akulasi (Rhincodon typus), bu og'irliklarga 21,5 tonnadan (47000 funt) oshadi. Balin kitlari singari, bu a filtri oziqlantiruvchi va birinchi navbatda iste'mol qiladi plankton.
- Buyurtma Rajiformes
- Buyurtma Lamniformes
- Sinf Plakodermi
- Buyurtma Arthrodira
- The largest armored fish, Dunkleosteus, arose during the late Devonian. At up to 10 metres (33 ft) in length[102] va 3.6 tonna (4.0 qisqa tonna ) in mass,[103] bu edi hypercarnivorous apex predator ish bilan ta'minlangan suction feeding.[104][105] Its contemporary, Titanichthys, apparently an early filter feeder, rivaled it in size. The arthrodires were eliminated by the environmental upheavals of the Late Devonian extinction, after existing for only about 50 million years.
- Buyurtma Arthrodira
- Sinf Tsefalopoda
- Buyurtma Ammonitida
- Kechki bo'r ammonit Parapuzosia seppenradensis reached a shell diameter of over 2 m.
- Buyurtma Teuthida
- A number of deep ocean creatures exhibit abyssal gigantism. Ular orasida ulkan kalmar (Arxiteutis) va ulkan kalmar (Mesonychoteuthis hamiltoni); both (although rarely seen) are believed to attain lengths of 12 m (39 ft) or more. The latter is the world's largest umurtqasizlar, va eng kattasi bor ko'zlar har qanday hayvon. Both are preyed upon by sperm whales.
- Buyurtma Ammonitida
- Ildiz guruhi Artropoda
- Buyurtma Radiodonta
- Anomalocarids were a group of very early legless marine arthropods that included the largest predators of the Kembriy, kabi Anomalocaris. Erta tomonidan Ordovik they had evolved into giant (for the time) filter feeders, apparently in response to the proliferation of plankton during the Buyuk Ordoviklar biologik xilma-xilligi. Aegirocassis grew to over 2 m in length.[106]
- Buyurtma Eurypterida
- Eurypterids (sea scorpions) were a diverse group of aquatic and possibly amphibious predators that included the most massive artropodlar to have existed. They survived over 200 million years, but finally died out in the Permiy-trias davridagi yo'q bo'lib ketish hodisasi bilan birga trilobitlar and most other forms of life present at the time, including most of the dominant terrestrial therapsids. The Early Devoniy Jaekelopterus reached an estimated length of 2.5 m (8.2 ft), not including its raptorial chelicerae, and is thought to have been a freshwater species.
- Buyurtma Radiodonta

Galereya
Yo'qolib ketdi

Biroz Paleozoy sea scorpions (Eurypterus shown) were larger than a human.

Dunkleosteus was a 10 m (33 ft) long toothless zirhli predatory Devonian platsoderm baliq.

Sail-backed pelycosaur Dimetrodon va temnospondil Eryops Shimoliy Amerikadan Permian.

Leedsichthys, a mid-Yura davri filter feeder fish, may have reached sizes of 7–16.5 m (23–54 ft).

Makronitar sauropodlar; chapdan, Kamarasaurus, Braxiosaurus, Giraffatitan, Evhelopus.

Tiranozavr was a 12.3 m (40 ft) long teropod dinosaur, an apex predator ning west North America.

Osiyo indricothere karkidon Paraceratherium was among the largest land mammals,[107] about twice a bush elephant's mass.

The Kechki miosen teratorn Argentavis of South America had a 7 m (23 ft) wingspan.

Reconstructed jaws of C. megalodon (Baltimor ).

Deinotherium had downward-curving tusks and ranged widely over Afro-Evroosiyo.

Titanis walleri, faqat terror qushi borligi ma'lum bosqinchi North America, was 2.5 m (8.2 ft) tall.

Hippo-sized Diprotodon of Australia, the largest marsupial of all time, became extinct 40,000 years ago.

Megalaniya, a giant carnivorous goanna of Australia, might have grown to 7 metres long.

Glyptodon, from South America's Pleystotsen, was an auto-sized singulat, ning qarindoshi armadillos.

American lions exceeded mavjud sherlar in size and ranged over much of N. America until 11,000 BP.

Woolly mammoths vanished after humans invaded their habitat in Eurasia and N. America.[1]

Haastning burguti, the largest eagle known, attacking moa (which included the tallest bird known).


















Yashash

The sharqiy gorilla is the largest and one of the more endangered primates on the planet.

Eng keng tarqalgan yo'lbarsning pastki turlari, Bengal tigers bor xavf ostida tomonidan brakonerlik va yashash joylarini yo'q qilish.

Polar bears, among the largest ayiqlar (consistent with Bergmann qoidasi ), bor zaif ga Global isish.

The juda xavfli black rhinoceros, up to 3.75 metres (12.3 ft) long, is tahdid qildi by poaching.

Wild Bactrian camels are critically endangered. Their ancestors originated in North America.

Unlike woolly karkidonlar va mamontlar, muskoxen narrowly survived the Quaternary extinctions.[1]

Begemişler, the heaviest and most aquatic juft oyoqli tuyoqlilar, bor kitlar ' closest living relatives.

The sperma kiti, eng kattasi tishli kit and toothed yirtqich, bor biggest brain.

The orca, eng kattasi delfin va to'plami yirtqich, bo'ladi highly intelligent va yashaydi murakkab jamiyatlar.

The kassa, the heaviest non-African bird, can run at 50 km/h through dense yomg'ir o'rmoni.

The timsoh sho'r suv is the largest living sudralib yuruvchi va xavfli predator of humans.

The Komodo ajdaho, an insular giant and the largest kaltakesak, bor serrated teeth and a venomous bite.

The yashil anakonda, suvda yashovchi konstriktor, is the heaviest ilon, weighing up to 97.5 kg (215 lb) or more.

The deep-diving ocean sunfish eng kattasi suyakli baliq, but its skeleton is mostly xaftaga oid.

The Nil perch, one of the largest freshwater fish, is also a damaging invaziv turlar.[3-eslatma]

The great white, the largest macropredatory fish, is more endangered than the yo'lbars.[110]

Examination of a 9 m ulkan kalmar, an abyssal giant va ikkinchi o'rinda sefalopod.


















{kind=link}
{kind=link}
Shuningdek qarang
- Avstraliya megafaunasi
- Bergmann qoidasi
- Xarizmatik megafauna
- Cope qoidasi
- Chuqur dengiz gigantizmi
- Hayvonot dunyosi
- Orol mitti
- Orol gigantizmi
- Largest organisms
- Eng yirik tarixiy organizmlar
- Eng og'ir sutemizuvchilar ro'yxati
- Eng yirik sutemizuvchilar ro'yxati
- Zamonaviy davrda topilgan megafauna ro'yxati
- Megafauna (mifologiya)
- Megafaunal bo'ri
- Megaflora
- Megaherb
- Yangi dunyo pleystotsenining yo'q bo'lib ketishi
- Pleistotsen megafauna
- To'rtlamchi davrda yo'q bo'lib ketish hodisasi
Izohlar
- ^ Nonavian dinosaur size was not similarly constrained because they had a different relationship between body mass and egg size than birds. The 400 kg Aepyornis had larger eggs than nearly all dinosaurs.[26][27]
- ^ Analysis indicates that 35 genera of North American mammals went extinct more or less simultaneously in this event.[42]
- ^ Perspektiv makes the fish appear larger relative to the man standing behind it (another example of a megafaunal species) than it actually is.
Adabiyotlar
- ^ a b v d Stuart, A. J. (November 1991). "Mammalian extinctions in the Late Pleistocene of northern Eurasia and North America". Biologik sharhlar. 66 (4): 453–562. doi:10.1111 / j.1469-185X.1991.tb01149.x. PMID 1801948. S2CID 41295526.
- ^ Martin, P. S. (1984). "Prehistoric overkill: The global model". In Martin, P. S.; Klein, R. G. (eds.). To'rtlamchi qirilishlar: tarixgacha bo'lgan inqilob. Arizona universiteti matbuoti. pp. 354–403. ISBN 978-0-8165-1100-6. OCLC 258362030.
- ^ Martin, P. S.; Steydman, D. (1999-06-30). "Orollar va qit'alarda tarixgacha yo'qolib ketish". MacPhee-da R. D. E (tahrir). Yaqin vaqt ichida yo'q qilish: sabablari, konteksti va oqibatlari. Umurtqali hayvonlarning paleobiologiyasidagi yutuqlar. 2. Nyu-York: Kluwer / Plenum. 17-56 betlar. ISBN 978-0-306-46092-0. OCLC 41368299. Olingan 2011-08-23. see page 17
- ^ Richard A. Farina, Sergio F. Vizcaino, Gerry De Iuliis (2013). "The Great American Biotic Interchange". Megafauna: Janubiy Amerikadagi pleystotsen gigant hayvonlari. Indiana University Press, Bloomington, Indiana. p. 150. ISBN 978-0-253-00230-3.CS1 maint: mualliflar parametridan foydalanadi (havola)
- ^ Bernhard A. Huber, Bradley J. Sinclair, Karl-Heinz Lampe (2005). "Historical Determinants of Mammal Species in Africa". African Biodiversity: Molecules, Organisms, Ecosystems. Springer. p. 294. ISBN 978-0387243153.CS1 maint: mualliflar parametridan foydalanadi (havola)
- ^ Muzlik davri hayvonlari. Illinoys shtati muzeyi
- ^ https://www.britannica.com/science/K-selected-species. Britannica. Qabul qilingan 2017-4-2.
- ^ Barnosky, A. D. (2004-10-01). "Assessing the Causes of Late Pleistocene Extinctions on the Continents". Ilm-fan. 306 (5693): 70–75. Bibcode:2004 yil ... 306 ... 70B. CiteSeerX 10.1.1.574.332. doi:10.1126 / science.1101476. PMID 15459379. S2CID 36156087.
- ^ Bruk, B. V.; Johnson, C. N. (2006). "Selective hunting of juveniles as a cause of the imperceptible overkill of the Australian Pleistocene megafauna". Alcheringa: Avstraliyaning Paleontologiya jurnali. 30 (sup1): 39–48. doi:10.1080/03115510609506854. S2CID 84205755.
- ^ a b v d e f g Evans, A. R.; Jones, D.; Boyer, A. G.; Braun, J. H .; Costa, D. P.; Ernest, S. K. M.; Fitzgerald, E. M. G.; Fortelius, M.; Gittleman, J. L.; Hamilton, M. J.; Harding, L. E.; Lintulaakso, K.; Lyons, S. K.; Okie, J. G.; Saarinen, J. J.; Sibir, R. M .; Smith, F. A.; Stephens, P. R .; Theodor, J. M.; Uhen, M. D. (2012-01-30). "Sutemizuvchilar evolyutsiyasining maksimal darajasi". PNAS. 109 (11): 4187–4190. Bibcode:2012PNAS..109.4187E. doi:10.1073 / pnas.1120774109. PMC 3306709. PMID 22308461.
- ^ a b v d e f g Smith, F. A.; Boyer, A. G.; Braun, J. H .; Costa, D. P.; Dayan, T .; Ernest, S. K. M.; Evans, A. R.; Fortelius, M.; Gittleman, J. L.; Hamilton, M. J.; Harding, L. E.; Lintulaakso, K.; Lyons, S. K.; McCain, C.; Okie, J. G.; Saarinen, J. J.; Sibir, R. M .; Stephens, P. R .; Theodor, J.; Uhen, M. D. (2010-11-26). "The Evolution of Maximum Body Size of Terrestrial Mammals". Ilm-fan. 330 (6008): 1216–1219. Bibcode:2010Sci...330.1216S. CiteSeerX 10.1.1.383.8581. doi:10.1126/science.1194830. PMID 21109666. S2CID 17272200.
- ^ Clauss, M.; Frey, R .; Kiefer, B .; Lechner-Doll, M.; Loylayn, V.; Polster, C .; Ressner, G. E.; Streich, W. J. (2003-04-24). "O'simlikxo'r sutemizuvchilarning tanasining maksimal kattaligi: oldingi ichakdagi morfofiziologik cheklovlar va orqa ichak fermentatorlarining moslashuvi" (PDF). Ekologiya. 136 (1): 14–27. Bibcode:2003 yil Ekol.136 ... 14C. doi:10.1007 / s00442-003-1254-z. PMID 12712314. S2CID 206989975.
- ^ a b Sorkin, B. (2008-04-10). "Yerdagi sutemizuvchi yirtqich hayvonlarda tana massasi bo'yicha biomexanik cheklash". Leteya. 41 (4): 333–347. doi:10.1111 / j.1502-3931.2007.00091.x.
- ^ Carbone, C.; Teacher, A; Rowcliffe, J. M. (2007-01-16). "The Costs of Carnivory". PLOS biologiyasi. 5 (2, e22): 363–368. doi:10.1371/journal.pbio.0050022. PMC 1769424. PMID 17227145.
- ^ a b v Ashton, K. G.; Tracy, M. C.; de Queiroz, A. (October 2000). "Bergmanning qoidasi sutemizuvchilar uchun amal qiladimi?". Amerikalik tabiatshunos. 156 (4): 390–415. doi:10.1086/303400. JSTOR 10.1086/303400. PMID 29592141.
- ^ Webb, J. (2015-02-19). "Evolution 'favours bigger sea creatures'". BBC yangiliklari. BBC. Olingan 2015-02-22.
- ^ Heim, N. A.; Knope, M. L.; Schaal, E. K.; Vang, S. C .; Payne, J. L. (2015-02-20). "Cope's rule in the evolution of marine animals". Ilm-fan. 347 (6224): 867–870. Bibcode:2015Sci...347..867H. doi:10.1126/science.1260065. PMID 25700517.
- ^ Thewissen, J. G. M.; Bajpai, S. (1 January 2001). "Whale Origins as a Poster Child for Macroevolution". BioScience. 51 (12): 1037–1049. doi:10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2. ISSN 0006-3568.
- ^ a b Goldbogen, J. A .; Cade, D. E.; Wisniewska, D. M.; Potvin, J.; Segre, P. S.; Savoca, M. S .; Hazen, E. L.; Czapanskiy, M. F.; Kahane-Rapport, S. R.; DeRuiter, S. L.; Gero, S.; Tønnesen, P.; Gough, W. T.; Hanson, M. B.; Holt, M. M.; Jensen, F. H.; Simon, M .; Stimpert, A. K.; Arranz, P.; Johnston, D. W.; Nowacek, D. P.; Parks, S. E.; Visser, F.; Friedlaender, A. S.; Tyack, P. L.; Madsen, P. T .; Pyenson, N. D. (2019). "Why whales are big but not bigger: Physiological drivers and ecological limits in the age of ocean giants". Ilm-fan. 366 (6471): 1367–1372. Bibcode:2019Sci...366.1367G. doi:10.1126/science.aax9044. hdl:10023/19285. PMID 31831666. S2CID 209339266.
- ^ a b v Mitchell, K. J .; Llamas, B.; Soubrier, J .; Rawlenceent, N. J .; Loyiq, T. H .; Wood, J.; Li, M. S. Y .; Kuper, A. (2014-05-23). "Qadimgi DNK fil qushlarini aniqlaydi va kivi qardosh taksindir va qushlarning ratit evolyutsiyasini aniqlaydi" (PDF). Ilm-fan. 344 (6186): 898–900. Bibcode:2014Sci ... 344..898M. doi:10.1126 / science.1251981. hdl:2328/35953. PMID 24855267. S2CID 206555952.
- ^ Baffetot, E .; Angst, D. (2014 yil noyabr). "Evropaning paleogenida katta uchmaydigan qushlarning stratigrafik tarqalishi va uning paleobiologik va paleogeografik oqibatlari". Earth-Science sharhlari. 138: 394–408. Bibcode:2014ESRv..138..394B. doi:10.1016 / j.earscirev.2014.07.001.
- ^ Phillips MJ, Gibb GC, Crimp EA, Penny D (yanvar 2010). "Tinamoz va moa birgalikda to'planadi: mitoxondriyal genomlar ketma-ketligini tahlil qilish ratitlar orasida parvozning mustaqil yo'qotishlarini aniqlaydi". Tizimli biologiya. 59 (1): 90–107. doi:10.1093 / sysbio / syp079. PMID 20525622.
- ^ Beyker, A. J .; Haddrat O .; Makferson, J.D .; Cloutier, A. (2014). "Parvozsiz kalamushlarda Moa-Tinamou kladegi va adaptiv morfologik konvergentsiyani genomik qo'llab-quvvatlash". Molekulyar biologiya va evolyutsiya. 31 (7): 1686–1696. doi:10.1093 / molbev / msu153. PMID 24825849.
- ^ Murray, Peter F.; Vikers-Rich, Patrisiya (2004). Ajoyib Mihirungs: Avstraliyaning orzu vaqtidagi ulkan parvozsiz qushlar. Indiana universiteti matbuoti. pp. 51, 314. ISBN 978-0-253-34282-9. Olingan 7 yanvar 2012.
- ^ Ibid (2004). p. 212. ISBN 978-0253342829.
- ^ Kenneth Carpenter (1999). Tuxum, uyalar va bolalar dinozavrlari: dinozavrlarning ko'payishiga qarash. Indiana universiteti matbuoti. p.100. ISBN 978-0-253-33497-8. OCLC 42009424. Olingan 6 may 2013.
- ^ Jekson, F. D .; Varricchio, D. J.; Jackson, R. A.; Vila, B.; Chiappe, L. M. (2008). "Comparison of water vapor conductance in a titanosaur egg from the Upper Cretaceous of Argentina and a Megaloolithus siruguei egg from Spain". Paleobiologiya. 34 (2): 229–246. doi:10.1666/0094-8373(2008)034[0229:COWVCI]2.0.CO;2. ISSN 0094-8373.
- ^ Ibid (2004). p. 277. ISBN 978-0253342829.
- ^ a b v Xansen, D. M .; Donlan, C. J .; Griffits, C. J .; Kempbell, K. J. (aprel 2010). "Ekologik tarix va yashirin muhofaza qilish salohiyati: katta va ulkan toshbaqalar taksoni almashtirish uchun namuna sifatida" (PDF). Ecography. 33 (2): 272–284. doi:10.1111 / j.1600-0587.2010.06305.x. Arxivlandi asl nusxasi (PDF) 2011 yil 24 iyulda. Olingan 2011-02-26.
- ^ a b Cione, A. L.; Tonni, E. P.; Soibelzon, L. (2003). "Buzilgan Zig-Zag: Janubiy Amerikada kekozoyning yirik sutemizuvchilar va toshbaqalarning yo'q bo'lib ketishi" (PDF). Rev. Mus. Argentino Cienc. Nat., N.s. 5 (1): 1–19. doi:10.22179 / REVMACN.5.26. ISSN 1514-5158. Arxivlandi asl nusxasi (PDF) 2011 yil 6-iyulda. Olingan 2011-02-06.
- ^ Alroy, J. (1999), "Putting North America's End-Pleistocene Megafaunal Extinction in Context: Large-Scale Analyses of Spatial Patterns, Extinction Rates, and Size Distributions", in MacPhee, R. D. E. (ed.), Extinctions in Near Time: Causes, Contexts, and Consequences, Advances in Vertebrate Paleobiology, 2, New York: Plenum, pp. 105–143, doi:10.1007/978-1-4757-5202-1_6, ISBN 978-1-4757-5202-1, OCLC 41368299
- ^ Corlett, R. T. (2006). "Megafaunal extinctions in tropical Asia" (PDF). Tropinet. 17 (3): 1–3. Olingan 2010-10-04.
- ^ Edmeades, Baz. "Megafauna — First Victims of the Human-Caused Extinction". megafauna.com. (internet-published book with Foreword by Pol S. Martin ). Arxivlandi asl nusxasi 2014-12-25 kunlari. Olingan 2020-02-13.
- ^ a b Martin, P. S. (2005). "Chapter 6. Deadly Syncopation". Mamontlarning alacakaranlığı: muzlik davrining yo'q bo'lib ketishi va Amerikaning qayta tiklanishi. Kaliforniya universiteti matbuoti. 118–128 betlar. ISBN 978-0-520-23141-2. OCLC 58055404. Olingan 2014-11-11.
- ^ a b Burney, D. A .; Flannery, T. F. (2005 yil iyul). "Odamlar bilan aloqada bo'lganidan keyin ellik ming yillik halokatli qirg'inlar" (PDF). Ekologiya va evolyutsiya tendentsiyalari. 20 (7): 395–401. doi:10.1016 / j.tree.2005.04.022. PMID 16701402. Arxivlandi asl nusxasi (PDF) 2010-06-10. Olingan 2014-11-11.
- ^ Roberts, R. G.; Flannery, T. F.; Ayliffe, L. K .; Yoshida, H .; Olley, J. M .; Prideaux, G. J .; Laslett, G. M.; Beyns, A .; Smit, M. A .; Jones, R.; Smit, B. L. (2001-06-08). "So'nggi Avstraliya Megafaunasining yangi davrlari: 46 ming yil oldin qit'ada yo'q bo'lib ketish" (PDF). Ilm-fan. 292 (5523): 1888–1892. Bibcode:2001 yil ... 292.1888R. doi:10.1126 / science.1060264. PMID 11397939. S2CID 45643228. Olingan 2011-08-26.
- ^ Callaway, E. (2016-09-21). "Human remains found in hobbit cave". Tabiat. doi:10.1038/nature.2016.20656. S2CID 89272546.
- ^ Diamond, Jared (2008-08-13). "Paleontologiya: Oxirgi ulkan kenguru". Tabiat. 454 (7206): 835–836. Bibcode:2008 yil natur.454..835D. doi:10.1038 / 454835a. PMID 18704074. S2CID 36583693.
- ^ Turni, C. S. M.; Flannery, T. F.; Roberts, R. G.; Reid, C.; Fifield, L. K.; Higham, T. F. G.; Jacobs, Z.; Kemp, N .; Colhoun, E. A.; Kalin, R. M.; Ogle, N. (2008-08-21). "Tasmaniya, Avstraliyada kechgacha saqlanib qolgan megafauna, ularning yo'q bo'lib ketishiga odamlarning aloqadorligini anglatadi". PNAS. 105 (34): 12150–12153. Bibcode:2008 yil PNAS..10512150T. doi:10.1073 / pnas.0801360105. PMC 2527880. PMID 18719103.
- ^ Roberts, R.; Jacobs, Z. (October 2008). "The Lost Giants of Tasmania" (PDF). Australasian Science. 29 (9): 14–17. Arxivlandi asl nusxasi (PDF) 2011-09-27 da. Olingan 2011-08-26.
- ^ Norton, C. J .; Kondo, Y .; Ono, A .; Chjan, Y .; Diab, M. C. (2009-05-23). "Yaponiyada MIS 3-2 o'tish davrida megafaunal yo'q bo'lib ketishining tabiati". To'rtlamchi xalqaro. 211 (1–2): 113–122. Bibcode:2010QuInt.211..113N. doi:10.1016 / j.quaint.2009.05.002.
- ^ Faith, J. T.; Surovell, T. A. (2009-12-08). "Synchronous extinction of North America's Pleistocene mammals". Milliy fanlar akademiyasi materiallari. 106 (49): 20641–20645. Bibcode:2009PNAS..10620641F. doi:10.1073/pnas.0908153106. PMC 2791611. PMID 19934040.
- ^ Haynes, Gary (2009). "Introduction to the Volume". In Haynes, Gary (ed.). American Megafaunal Extinctions at the End of the Pleistocene. Umurtqali hayvonlarning paleobiologiyasi va paleoantropologiyasi. Springer. 1-20 betlar. doi:10.1007/978-1-4020-8793-6_1. ISBN 978-1-4020-8792-9.
- ^ Fiedel, Styuart (2009). "Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction". In Haynes, Gary (ed.). American Megafaunal Extinctions at the End of the Pleistocene. Umurtqali hayvonlarning paleobiologiyasi va paleoantropologiyasi. Springer. 21-37 betlar. doi:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9.
- ^ Simmons, A. H. (1999). Orol jamiyatida hayvonlarning yo'q bo'lib ketishi: Kiprning gigippopotamus ovchilari. Interdisciplinary Contributions to Archaeology. Kluwer Academic / Plenum nashriyotlari. p. 382. doi:10.1007 / b109876. ISBN 978-0-306-46088-3. OCLC 41712246.
- ^ Simmons, A. H.; Mandel, R. D. (December 2007). "Not Such a New Light: A Response to Ammerman and Noller". Jahon arxeologiyasi. 39 (4): 475–482. doi:10.1080/00438240701676169. JSTOR 40026143. S2CID 161791746.
- ^ Steydman, D.; Martin, P. S.; MacPhee, R. D. E .; Jul, A. J. T .; McDonald, H. G.; Vuds, C. A .; Iturralde-Vinent, M.; Xodgins, G. V. L. (2005-08-16). "Qit'alar va orollarda to'rtinchi davrning yalqovlari asinxron tarzda yo'q bo'lib ketishi". Proc. Natl. Akad. Ilmiy ish. AQSH. 102 (33): 11763–11768. Bibcode:2005 yil PNAS..10211763S. doi:10.1073 / pnas.0502777102. PMC 1187974. PMID 16085711.
- ^ Kuk, S. B.; Dalavos, L. M .; Mychajliw, A. M.; Turvi, S. T .; Upham, N. S. (2017). "Antropogen qirilish G'arbiy Hind sutemizuvchilarining golosen kamayishiga hukmronlik qilmoqda". Ekologiya, evolyutsiya va sistematikaning yillik sharhi. 48 (1): 301–327. doi:10.1146 / annurev-ecolsys-110316-022754.
- ^ Anderson, A .; Qum, C .; Petchey, F.; Loyiq, T. H. (2010). "Yangi Kaledoniyadagi hayvonlarning yo'q bo'lib ketishi va odamlarning yashashi: Pindai g'orlaridagi yangi tadqiqotlarning dastlabki natijalari va natijalari". Tinch okeani arxeologiyasi jurnali. 1 (1): 89–109. hdl:10289/5404.
- ^ Oq, A. V.; Loyiq, T. H .; Xokins, S .; Bedford, S .; Spriggs, M. (2010-08-16). "Megafaunal meiolaniid shoxli toshbaqalar Vanuatu, Tinch okeanining janubi-g'arbiy qismida odamlar yashaguncha omon qoldi". Proc. Natl. Akad. Ilmiy ish. AQSH. 107 (35): 15512–15516. Bibcode:2010PNAS..10715512W. doi:10.1073 / pnas.1005780107. PMC 2932593. PMID 20713711.
- ^ Burney, D. A .; Burney, L. P.; Godfri, L. R .; Jungers, W. L .; Gudman, S. M .; Rayt, H. T .; Jul. A. J. T. (July 2004). "A chronology for late prehistoric Madagascar". Inson evolyutsiyasi jurnali. 47 (1–2): 25–63. doi:10.1016/j.jhevol.2004.05.005. PMID 15288523.
- ^ Xoldauey, R. N .; Jakomb, C. (2000-03-24). "Moasning tezda yo'q bo'lib ketishi (Aves: Dinornithiformes): namuna, sinov va natijalar". Ilm-fan. 287 (5461): 2250–2254. Bibcode:2000Sci ... 287.2250H. doi:10.1126 / science.287.5461.2250. PMID 10731144.
- ^ Janoo, A. (April 2005). "Discovery of isolated dodo bones (Raphus cucullatus (L.), Aves, Columbiformes) from Mauritius cave shelters highlights human predation, with a comment on the status of the family Raphidae Wetmore, 1930". Annales de Paléontologie. 91 (2): 167–180. doi:10.1016 / j.annpal.2004.12.002.
- ^ Anderson, P. K. (July 1995). "Raqobat, yirtqichlik va Steller dengiz sigirining rivojlanishi va yo'q bo'lib ketishi, Hydrodamalis gigas". Dengiz sutemizuvchilar haqidagi fan. 11 (3): 391–394. doi:10.1111 / j.1748-7692.1995.tb00294.x. Arxivlandi asl nusxasi 2011-05-11. Olingan 2011-08-30.
- ^ Kuper, A .; Turney, C.; Hughen, K. A.; Bruk, B. V.; McDonald, H. G.; Bradshaw, C. J. A. (2015-07-23). "Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover". Ilm-fan. 349 (6248): 602–6. Bibcode:2015Sci...349..602C. doi:10.1126/science.aac4315. PMID 26250679. S2CID 31686497.
- ^ Müller, U. C.; Pross, J.; Tzedakis, P. C.; Gamble, C .; Kotthoff, U.; Schmiedl, G.; Wulf, S.; Christanis, K. (February 2011). "The role of climate in the spread of modern humans into Europe". To'rtlamchi davrga oid ilmiy sharhlar. 30 (3–4): 273–279. Bibcode:2011QSRv...30..273M. doi:10.1016/j.quascirev.2010.11.016.
- ^ Biello, D. (2012-03-22). "Big Kill, Not Big Chill, Finished Off Giant Kangaroos". Scientific American news. Olingan 2012-03-25.
- ^ McGlone, M. (2012-03-23). "The Hunters Did It". Ilm-fan. 335 (6075): 1452–1453. Bibcode:2012Sci...335.1452M. doi:10.1126/science.1220176. PMID 22442471. S2CID 36914192.
- ^ a b v Rule, S.; Bruk, B. V.; Xaberle, S. G.; Turni, C. S. M.; Kershaw, A. P. (2012-03-23). "The Aftermath of Megafaunal Extinction: Ecosystem Transformation in Pleistocene Australia". Ilm-fan. 335 (6075): 1483–1486. Bibcode:2012Sci...335.1483R. doi:10.1126/science.1214261. PMID 22442481. S2CID 26675232.
- ^ a b Jonson, C. N .; Alroy, J .; Beeton, N. J .; Bird, M. I.; Bruk, B. V.; Kuper, A .; Gillespi, R.; Errando-Peres, S.; Jacobs, Z.; Miller, G. H .; Prideaux, G. J .; Roberts, R. G.; Rodriges-Rey, M.; Saltre, F.; Turni, C. S. M.; Bradshaw, C. J. A. (2016 yil 10-fevral). "What caused extinction of the Pleistocene megafauna of Sahul?". Qirollik jamiyati materiallari B: Biologiya fanlari. 283 (1824): 20152399. doi:10.1098 / rspb.2015.2399. PMC 4760161. PMID 26865301.
- ^ Miller, G. H .; Magee, J. V.; Jonson, B. J .; Fogel, M. L .; Qoshiqchi, N. A .; Makkullox, M. T .; Ayliffe, L. K. (1999-01-08). "Pleystotsenning yo'q bo'lib ketishi Genyornis newtoni: Avstraliyaning megafaunasiga insonning ta'siri ". Ilm-fan. 283 (5399): 205–208. doi:10.1126 / science.283.5399.205. PMID 9880249.
- ^ Miller, G.; Magee, J .; Smith, M.; Spooner, N.; Beyns, A .; Lehman, S.; Fogel, M.; Jonson, X.; Uilyams, D.; Klark, P .; Florian, C.; Holst, R.; DeVogel, S. (2016-01-29). "Human predation contributed to the extinction of the Australian megafaunal bird Genyornis newtoni ∼47 ka". Tabiat aloqalari. 7: 10496. Bibcode:2016NatCo...710496M. doi:10.1038 / ncomms10496. PMC 4740177. PMID 26823193.
- ^ Johnson, C. (2009-11-20). "Megafaunal Decline and Fall". Ilm-fan. 326 (5956): 1072–1073. Bibcode:2009Sci...326.1072J. doi:10.1126/science.1182770. PMID 19965418. S2CID 206523763.
- ^ Gill, J. L.; Uilyams, J. V.; Jekson, S. T .; Lininger, K. B.; Robinson, G. S. (2009-11-20). "Pleistocene Megafaunal Collapse, Novel Plant Communities, and Enhanced Fire Regimes in North America" (PDF). Ilm-fan. 326 (5956): 1100–1103. Bibcode:2009Sci...326.1100G. doi:10.1126/science.1179504. PMID 19965426. S2CID 206522597.
- ^ Fiedal, Stuart (2009). "Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction". In Haynes, Gary (ed.). American Megafaunal Extinctions at the End of the Pleistocene. Umurtqali hayvonlarning paleobiologiyasi va paleoantropologiyasi. Springer. 21-37 betlar. doi:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9.
- ^ Martin, P. S. (2005). "4-bob. Uydagi yalqovlar". Mamontlarning alacakaranlığı: muzlik davrining yo'q bo'lib ketishi va Amerikaning qayta tiklanishi. Kaliforniya universiteti matbuoti. 78–99 betlar. ISBN 978-0-520-23141-2. OCLC 58055404. Olingan 2014-11-11.
- ^ Milman, Oliver (February 6, 2019). "The killing of large species is pushing them towards extinction, study finds". Guardian. Olingan 13 fevral, 2019.
- ^ Ripple, W. J .; va boshq. (2019). "Are we eating the world's megafauna to extinction?". Tabiatni muhofaza qilish xatlari: e12627. doi:10.1111/conl.12627.
- ^ Alroy, J. (2008-08-12). "Dynamics of origination and extinction in the marine fossil record". PNAS. 105 Suppl 1 (Supplement_1): 11536–11542. Bibcode:2008PNAS..10511536A. doi:10.1073/pnas.0802597105. PMC 2556405. PMID 18695240.
- ^ Bo'ri, A .; Doughty, C. E.; Malhi, Y. (2013). "Lateral Diffusion of Nutrients by Mammalian Herbivores in Terrestrial Ecosystems". PLOS ONE. 8 (8): e71352. Bibcode:2013PLoSO...871352W. doi:10.1371/journal.pone.0071352. PMC 3739793. PMID 23951141.
- ^ Marshall, M. (2013-08-11). "Ecosystems still feel the pain of ancient extinctions". Yangi olim. Olingan 2013-08-12.
- ^ a b Doughty, C. E.; Bo'ri, A .; Malhi, Y. (2013-08-11). "The legacy of the Pleistocene megafauna extinctions on nutrient availability in Amazonia". Tabiatshunoslik. 6 (9): 761–764. Bibcode:2013NatGe ... 6..761D. doi:10.1038 / ngeo1895.
- ^ Roman, J.; Makkarti, JJ (2010). "Kit nasosi: dengiz sutemizuvchilari qirg'oq havzasida birlamchi mahsuldorlikni oshiradi". PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007.
- ^ a b v Uilkinson, D. M .; Nisbet, E. G.; Ruxton, G. D. (2012-05-08). "Could methane produced by sauropod dinosaurs have helped drive Mesozoic climate warmth?". Hozirgi biologiya. 22 (9): R292–R293. doi:10.1016/j.cub.2012.03.042. PMID 22575462. Olingan 2012-05-08.
- ^ "Dinosaur gases 'warmed the Earth'". BBC Nature News. 2012-05-07. Olingan 2012-05-08.
- ^ a b v Smith, F. A.; Elliot, S. M.; Lyons, S. K. (2010-05-23). "Methane emissions from extinct megafauna". Tabiatshunoslik. 3 (6): 374–375. Bibcode:2010NatGe...3..374S. doi:10.1038/ngeo877.
- ^ Kelliher, F. M.; Clark, H. (2010-03-15). "Methane emissions from bison—An historic herd estimate for the North American Great Plains". Qishloq xo'jaligi va o'rmon meteorologiyasi. 150 (3): 473–577. Bibcode:2010AgFM..150..473K. doi:10.1016/j.agrformet.2009.11.019.
- ^ Helgen et. all, Kristofer M. (2006). "Ecological and evolutionary significance of sizes of giant extinct kangaroos" (PDF). Avstraliya Zoologiya jurnali. 54 (4): 293–301. doi:10.1071 / ZO05077 – via si.edu.
- ^ Larramendi, A. (2016). "Shoulder height, body mass and shape of proboscideans" (PDF). Acta Palaeontologica Polonica. 61 (3): 537–574. doi:10.4202 / ilova.00136.2014. S2CID 2092950. Olingan 2018-03-22.
- ^ a b Farina, Richard A.; Vitskaino, Serxio F.; De Iuliis, Gerri (2013 yil 22-may). Megafauna: Janubiy Amerikadagi pleystotsen gigant hayvonlari. Indiana universiteti matbuoti. ISBN 978-0-253-00719-3. OCLC 779244424.
- ^ Chjan, Y .; Harrison, T. (2017). "Gigantopitekus qora: Osiyo pleystosenidagi ulkan maymun qayta ko'rib chiqildi ". Amerika jismoniy antropologiya jurnali. 162 (S63): 153-177. doi:10.1002 / ajpa.23150. PMID 28105715.
- ^ Ruff, C. B .; Trinkaus, E .; Holliday, T. W. (1997-05-08). "Tana massasi va ensefalizatsiya pleystotsen gomosida". Tabiat. 387 (6629): 173–176. Bibcode:1997 yil Natura. 387..173R. doi:10.1038 / 387173a0. PMID 9144286. S2CID 4320413.
- ^ Grayn, F. E .; Jumgers, W. L .; Tobias, P. V.; Pearson, O. M. (1995 yil iyun). "Qoldiqlar Homo femur Berg Aukas, shimoliy Namibiya ". Amerika jismoniy antropologiya jurnali. 97 (2): 151–185. doi:10.1002 / ajpa.1330970207. PMID 7653506.
- ^ Smit, Kris; Burger, Li (2007 yil noyabr). "Bizning hikoyamiz: Inson ajdodlari qoldiqlari". Yalang'och olimlar. Olingan 2011-02-19.
- ^ Kappelman, Jon (1997-05-08). "Ular gigant bo'lishi mumkin". Tabiat. 387 (6629): 126–127. Bibcode:1997 yil Natura. 387..126K. doi:10.1038 / 387126a0. PMID 9144276. S2CID 4328242.
- ^ de Barros Ferraz, K.M.P.M.; Bonax, K .; Verdade, LM (2005). "Kapybaralarda tana massasi va tana uzunligi o'rtasidagi bog'liqlik (Hydrochoerus hydrochaeris)". Biota Neotropica. 5 (1): 197–200. doi:10.1590 / S1676-06032005000100020.
- ^ Kitchener, AC, Breitenmoser-Würsten, C., Eyzirik, E., Gentry, A., Werdelin, L., Wilting, A. va Yamaguchi, N. (2017). "Felidae-ning qayta ko'rib chiqilgan taksonomiyasi: IUCN mushuklari bo'yicha mutaxassislar guruhining mushuklarni tasniflash bo'yicha maxsus guruhining yakuniy hisoboti" (PDF). Mushuk yangiliklari (Maxsus son 11).CS1 maint: mualliflar parametridan foydalanadi (havola)
- ^ Brakefield, Tom (1993). Katta mushuklar: Qudrat shohligi. Voyageur Press. p.44. ISBN 978-0-89658-329-0.
- ^ Nowell, Kristin; Jekson, Piter (1996). Yovvoyi mushuklar: holatni o'rganish va uni muhofaza qilish bo'yicha tadbirlar rejasi (PDF). Gland, Shveytsariya: IUCN / SSC Cat Specialist Group. p. 56. ISBN 978-2-8317-0045-8.
- ^ Kitchener, A. va Yamaguchi, N. (2009). "Yo'lbars nima? Biogeografiya, morfologiya va taksonomiya". Tilsonda R.; Nyhus, P. J. (tahrir). Dunyo yo'lbarslari: fan, siyosat va tabiatni muhofaza qilish Panthera yo'lbarsi. Akademik matbuot. 53-84 betlar. ISBN 978-0-08-094751-8.CS1 maint: mualliflar parametridan foydalanadi (havola)
- ^ Slaght, J. C., Mikel, D. G., Nikolaev, I. G., Goodrich, J. M., Smirnov, E. N., Traylor-Holzer, K., Christie, S., Arjanova, T., Smith, J. L. D. va Karanth, K. U. (2005). "6-bob. Yirtqich hayvonlarning shohi kim? Yovvoyi va asirlikda bo'lgan Amur yo'lbarslarining tana vazniga oid boshqa tarixiy va zamonaviy ma'lumotlar". (PDF). D. G. Mikelda; E. N. Smirnov; J.M.Gudrich (tahrir). Sixote-Alin Zapovednikdagi yo'lbarslar: ekologiya va tabiatni muhofaza qilish (rus tilida). Vladivostok, Rossiya: PSP. 25-35 betlar.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ http://www.ligerworld.com/samson-the-biggest-tiger.html
- ^ DeMaster, D.P .; Stirling, I. (1981 yil 8-may). "Ursus maritimus". Sutemizuvchilar turlari (145): 1–7. doi:10.2307/3504138. JSTOR 3503828.
- ^ Pasitschniak-Arts, M. (1993 yil 23 aprel). "Ursus arctos". Sutemizuvchilar turlari (439): 1–10. doi:10.2307/3503828. JSTOR 3504138.
- ^ Soibelzon, L. H .; Schubert, B. W. (yanvar 2011). "Eng katta taniqli ayiq, Arktoterium angustidens, Argentinaning Pleistosen davridagi Pampean mintaqasidan: ayiqlarning hajmi va parhez tendentsiyalari muhokama qilingan holda ". Paleontologiya jurnali. 85 (1): 69–75. doi:10.1666/10-037.1. S2CID 129585554. Olingan 2011-06-01.
- ^ Svift, E. M. (1997-11-17). "Ularda qanday katta og'izlar bor?: Afrikada gippo bilan ish yuritadigan sayohatchilar ertakni aytib berishga qodir emaslar". Sport Illustrated Vault. Time Inc. Olingan 2011-11-16.
- ^ ^ J. Kalambokidis va G. Shtayger (1998). Moviy kitlar. Voyageur Press. ISBN 0-89658-338-4.
- ^ ^ "Hayvonlarning yozuvlari". Smithsonian National Zoological Park. Qabul qilingan 2007-05-29.
- ^ Anteosaurus Arxivlandi 2016-03-14 da Orqaga qaytish mashinasi. Palaeos.org (2013-04-22)
- ^ Sulej, T .; Niedwiedzki, G. (2019). "Oyoqlari tik, fil bo'yidagi so'nggi trias davridagi sinapsid". Ilm-fan. 363 (6422): 78–80. Bibcode:2019Sci ... 363 ... 78S. doi:10.1126 / science.aal4853. PMID 30467179.
- ^ Sankt Fler, Nikolay (2019 yil 4-yanvar). "Dinozavrlar bilan birga boqilgan sutemizuvchilarning fil o'lchamidagi qarindoshi". The New York Times. Olingan 6 yanvar 2019.
- ^ Palmer, D. (2002 yil 1-iyul). Marshal Illustrated Entsiklopediyasi Dinozavrlar va tarixdan oldingi hayvonlar. Yangi qatorli kitoblar. ISBN 978-1-57717-293-2. OCLC 183092423. Olingan 2013-06-10.
- ^ Monster baliqlari qarshilikni eng kuchli luqma bilan tor-mor qildi. Sidney Morning Herald. 2006 yil 30-noyabr.
- ^ Anderson, P. S.L; Westneat, M. V (2006-11-28). "Oziqlantirish mexanikasi va bosh suyagining tishlash kuchini modellashtirish Dunkleosteus terrelli, qadimiy tepalik yirtqichi ". Biologiya xatlari. 3 (1): 77–80. doi:10.1098 / rsbl.2006.0569. ISSN 1744-9561. PMC 2373817. PMID 17443970.
- ^ Anderson, P.S.L. (2010-05-04). "Artrodire platsodermalarida bosh suyagining kinematik xilma-xilligi va funktsional konvergentsiyasini o'rganish uchun bog'lanish modellaridan foydalanish". Morfologiya jurnali. 271 (8): 990–1005. doi:10.1002 / jmor.10850. ISSN 0362-2525. PMID 20623651. S2CID 46604512.
- ^ Van Roy, P .; Deyli, A. S.; Briggs, D. E. G. (2015 yil 11 mart). "Anomalokaridid magistral gomologiyasi ulkan filtrli oziqlantiruvchi tomonidan aniqlangan". Tabiat. 522 (7554): 77–80. Bibcode:2015 Noyabr 522 ... 77V. doi:10.1038 / tabiat 14256. PMID 25762145. S2CID 205242881.
- ^ Tsubamoto, T. (2012). "Sutemizuvchilardagi astragaldan tana massasini baholash". Acta Palaeontologica Polonica: 259–265. doi:10.4202 / ilova.2011.0067. S2CID 54686160.
- ^ Palmer, D., ed. (1999). Marshal Illustrated Entsiklopediyasi Dinozavrlar va tarixdan oldingi hayvonlar. London: Marshall nashrlari. p. 248. ISBN 978-1-84028-152-1.
- ^ Moyano, S.R .; Jannini, N.P. (2018-10-10). "Tapirusda tug'ruqdan keyingi rivojlanish probozasi bilan bog'liq kranial belgilar (Perissodactyla: Tapiridae) va boshqa mavjud va toshqotgan tuyoqli sutemizuvchilar bilan taqqoslash". Zoologischer Anzeiger. 277 (7554): 143–147. doi:10.1016 / j.jcz.2018.08.005. ISSN 0044-5231.
- ^ Sample, Ian (2010 yil 19-fevral). "Buyuk oq akula yo'lbarsdan ko'ra ko'proq xavflidir, deydi olim". Guardian. Olingan 14 avgust 2013.