Liken - Lichen








A liken (/ˈlaɪkən/ LY-ken yoki, ba'zan Buyuk Britaniyada, /ˈlɪtʃən/, LICH-uz ) kompozitsiyadir organizm kelib chiqadi suv o'tlari yoki siyanobakteriyalar orasida yashash iplar bir nechta qo'ziqorinlar turlari[1] a mutalistik munosabatlar.[2][3][4] Likenler ularning tarkibiy organizmlaridan farqli xususiyatlarga ega. Lishayniklar turli xil ranglarda, o'lchamlarda va shakllarda bo'ladi va ba'zan o'simlikka o'xshaydi, ammo likenlar bunday emas o'simliklar. Likenler mayda, bargsiz shoxlarga ega bo'lishi mumkin (frutikoz ), tekis bargga o'xshash tuzilmalar (folioz ), po'stlog'i kabi sirt ustida yotadigan pufaklar (qobiq ),[5] changga o'xshash ko'rinish (moxov ), yoki boshqa o'sish shakllari.[6]
A makrolixen bu tupga o'xshash yoki bargli liken; boshqa barcha likenlarning muddati tugaydi mikrolixenlar.[2] Bu erda "so'l" va "mikro" hajmga emas, balki o'sish shakliga ishora qiladi.[2] Likenlarning umumiy nomlari so'zni o'z ichiga olishi mumkin mox (masalan, "kiyik moxi ", "Island mox ") va likenler yuzaki ko'rinishi mumkin va moxlar bilan o'sishi mumkin, ammo likenler moxlar yoki har qanday o'simlik bilan bog'liq emas.[4]:3 Likenler o'simliklar singari suv va ozuqa moddalarini singdiradigan ildizlarga ega emas,[7]:2 ammo o'simliklar kabi, ular o'zlarining oziqlanishlarini ishlab chiqaradilar fotosintez.[8] Ular o'simliklarda o'sganda, ular yashamaydilar parazitlar, lekin buning o'rniga o'simliklarni a sifatida ishlating substrat.
Likenler kelib chiqadi dengiz sathi balandga alp balandliklar, ko'plab atrof-muhit sharoitida va deyarli har qanday sirtda o'sishi mumkin.[8] Likenler po'stlog'ida juda ko'paymoqda, barglar, moxlar, boshqa likenlarda,[7] va "havoda yashovchi" shoxlarga osilgan (epifitlar ) ichida yomg'ir o'rmonlari va mo''tadil o'rmonzor. Ular toshlarda, devorlarda o'sadi, qabr toshlari, tomlar, ochiq tuproq yuzalari, kauchuk, suyaklar va tuproq tarkibida biologik tuproq po'stlog'i. Lishayniklarning har xil turlari Yerdagi eng ekstremal muhitda yashashga moslashgan: Arktik tundra, issiq quruq cho'llar, qoyali qirg'oqlar va toksik cüruf uyumlar. Ular hatto qattiq tosh ichida yashab, donalar orasida o'sishi mumkin.
Ma'lumotlarga ko'ra, Yer yuzining 6-8% liklari bilan qoplangan.[9] Likenlarning 20 mingga yaqin turi ma'lum.[10] Ba'zi likonlar jinsiy yo'l bilan ko'payish qobiliyatini yo'qotgan, shunga qaramay davom etmoqda aniqlashtirish.[7][11] Lishayniklarni zamburug'lar, suv o'tlari yoki siyanobakteriyalar boshqalari bilan aloqada bo'lish imkoniyatiga ega bo'lgan nisbatan o'ziga xos miniatyura ekotizimi deb ko'rish mumkin. mikroorganizmlar yanada murakkablashishi mumkin bo'lgan ishlaydigan tizimda kompozitsion organizm.[12][13][14][15]
Likenler bo'lishi mumkin uzoq umr ko'rgan, ba'zilari esa eng qadimgi jonzotlar qatoriga kiradi.[4][16] Ular ko'chkiga o'xshash hodisadan keyin paydo bo'lgan yangi toshlarda o'sadigan birinchi tirik mavjudotlardan biri. Ba'zi lishayniklarning uzoq umr ko'rishlari va sekin va muntazam o'sish sur'atlari hozirgi kunga qadar ishlatilishi mumkin (likenometriya ).
Talaffuz va etimologiya
Amerikalik ingliz tilida "liken" "liken" fe'l bilan bir xil talaffuz qilinadi (/ˈlaɪkeng/). Britaniyalik inglizchada ushbu talaffuz ham, "oshxona" bilan qofiya ham /ˈlɪtʃeng/) ishlatiladi.[17][18][19]
Ingliz tili liken kelib chiqadi Yunoncha χήνiχήν leichēn ("daraxt moxi, liken, liken kabi terining otilishi") orqali Lotin liken.[20][21][22] So'zma-so'z "yalash" degan ma'noni anglatuvchi yunoncha ism fe'ldan kelib chiqqan νiν leichein, "yalamoq".[23][24]
So'z kabi mox, so'z liken an sifatida ishlatiladi sanoqsiz ism, "Liken toshlarda o'sadi" da bo'lgani kabi.
O'sish shakllari
Likenler turli shakl va shakllarda o'sadi (morfologiyalar ). Liken shakli odatda qo'ziqorin filamentlarini tashkil qilish bilan belgilanadi.[25] Reproduktiv bo'lmagan to'qimalar yoki vegetativ tana qismlari deyiladi talus. Likenler talus turi bo'yicha guruhlanadi, chunki talus odatda likenning eng ko'zga ko'ringan qismidir. Talusning o'sish shakllari odatda bir necha asosiy ichki tuzilish turlariga to'g'ri keladi. Likenlarning umumiy nomlari ko'pincha likenga xos bo'lgan o'sish shakli yoki rangidan kelib chiqadi tur.
Liken talusining o'sish shakllarining umumiy guruhlari:
- frutikoz[26][27][28] - tutam yoki ko'p shoxli bargsiz mini-buta kabi o'sib, tik yoki pastga osilgan, uchburchak shoxlari deyarli yumaloq (va)terete ) yoki tekislangan
- folioz[26][27] - 2 o'lchovli, tekis, bargga o'xshash loblarda o'sadi
- qobiq[5][26][27] - sirtga mahkam yopishgan qobiqqa o'xshash (substrat ) qalin bo'yoq kabi
- skuuloza[28] - quyida barg bargiga o'xshash po'stlog'idan hosil bo'lgan, ammo uchlari bo'sh
- moxov[29] - chang
- jelatinli - jele o'xshash
- ipli - ipli yoki o'xshash matli sochlar
- byssoid - aqlli, shunga o'xshash masxara qilingan jun
- tuzilmasdan
Bitta liken turidagi o'sish turlarining xilma-xilligi, o'sish turlarining tavsiflari orasidagi kulrang joylar va o'sish turlari o'rtasida bir-birining ustiga chiqadigan farqlar mavjud, shuning uchun ba'zi mualliflar likenlarni har xil o'sish tavsiflaridan foydalangan holda tasvirlashlari mumkin.
Qisqichbaqasimon liken qariganida, markaz eski quritilgan bo'yoq, eski singan asfalt qoplamasi singari yoki qurigan ko'l tubidagi yorilgan loyning ko'p qirrali "orollari" singari yorila boshlaydi. Bunga borliq deyiladi rimose yoki yolg'iz, va yoriqlar bilan ajratilgan "orol" qismlari deyiladi Areola.[26] Areolalar bir-biridan ajratilgan ko'rinadi, lekin (yoki)[iqtibos kerak ] asosiy "protallus" yoki "gipotalal" bilan bog'langan.[29] Qisqichbaqasimon liken markazdan o'sib chiqqanda va tashqariga chiqqanda paydo bo'ladi, deyiladi qobiq platsodioidi. Areolalarning qirralari substratdan yuqoriga ko'tarilganda, deyiladi skuuloza.[30]:159[28]
Ushbu o'sish shakllari guruhlari aniq belgilanmagan. Folioz likenlari ba'zan tarvaqaylab, frutikoza bo'lib ko'rinishi mumkin. Frutikoz likenlari tekislangan shoxlangan qismlarga ega bo'lishi va bargli ko'rinishi mumkin. Skuamulyoz likenlari qirralari ko'tarilgan joylarda paydo bo'lishi mumkin. Jelatinli likenlar quriganda bargli ko'rinishi mumkin.[30]:159 Ushbu holatlarda ularni ajratib ko'rsatish vositalari quyidagi bo'limlarda keltirilgan.
Ko'paytirish bilan bog'liq tuzilmalar ko'pincha talus yuzasida disklar, tepaliklar yoki silliq chiziqlar ko'rinishida bo'ladi.[7]:4 Talus har doim ham likenning ko'zga ko'rinadigan qismi emas. Ba'zi likonlar o'sishi mumkin ichida donalar orasidagi qattiq tosh (endolitik likenler ), jinsning mevali qismi faqat jinsdan tashqarida o'sib borishi bilan ko'rinadi.[26] Ular rang yoki ko'rinishda dramatik bo'lishi mumkin.[26] Ushbu jinsiy qismlarning shakllari yuqoridagi o'sish shakllari turkumlariga kirmaydi.[26] Ko'zga ko'rinadigan reproduktiv qismlar ko'pincha dumaloq, ko'tarilgan, plastinka yoki diskka o'xshash o'simtalar, qirralari burishgan va quyida bo'limlarda tasvirlangan.
Rang
Likenler turli xil ranglarga ega.[7]:4 Bo'yash odatda fotosintez komponenti bilan belgilanadi.[25] Sariq kabi maxsus pigmentlar usnik kislota, qizil rang, apelsin, sariq va jigarrang ranglarni o'z ichiga olgan likenlarga turli xil ranglarni bering, ayniqsa ochiq va quruq yashash joylarida.[31] Maxsus pigmentlar bo'lmagan taqdirda, likenler odatda namlanganda och yashildan zaytungacha, quruq bo'lsa kulrang yoki kulrang-yashildan jigarranggacha bo'ladi.[31] Buning sababi shundaki, namlik yuzaning terisini keltirib chiqaradi (korteks ) shaffofroq bo'lib, yashil fotobiont qatlamini ochib beradi.[31] Yomg'irdan keyin porloq displeylarda turli rangdagi parchalar "jonlanib" yoki "porlab" turganda, ochiq tosh yuzalarining katta maydonlarini qoplaydigan turli xil rangli likenlar yoki qobiq bilan qoplangan yoki osilgan likenlar ajoyib namoyish bo'lishi mumkin.
Yorug'lik ta'siriga qarab, turli xil rangli likonlar tosh yuzining turli xil qo'shni qismlarida yashashi mumkin.[31] Likenlarning koloniyalari ko'rinishda ajoyib bo'lishi mumkin, ular o'rmonlarda va tabiiy joylarda, masalan, vizual landshaftning aksariyat qismida hukmronlik qiladi. vertikal "bo'yoq" keng tosh yuzlarini qoplagan Yosemit milliy bog'i.[32]
Rang identifikatsiyalashda ishlatiladi.[33]:4 Likenning rangi likenning ho'l yoki quruqligiga qarab o'zgaradi.[33] Identifikatsiya qilish uchun ishlatiladigan rang tavsiflari liken quruqligini ko'rsatadigan rangga asoslangan.[33] Fotosintez sherigi sifatida siyanobakteriyali quruq likenlar quyuq kulrang, jigarrang yoki qora ranglarga ega.[33]
Folioz likenlarining barg barglari osti qismi yuqoridan boshqacha rangga ega (dorsiventral ), ko'pincha jigarrang yoki qora, ba'zan oq. Frutikoz likenining tekislangan "novdalari" bo'lishi mumkin, ular folioz likeniga o'xshaydi, ammo frutikoz likenidagi bargga o'xshash strukturaning pastki qismi bir xil yuqori tomoni sifatida rang. Folioz likenining bargga o'xshash po'stlog'i frutikoz likeniga o'xshab dallanishi mumkin, ammo pastki tomoni boshqacha yuqori tomondan rang.[29]
Ba'zi bir jelatinli likenlarning porlashi tomonidan yaratilgan shilimshiq sekretsiyalar.[25]
Ichki tuzilish va o'sish shakllari
Liken oddiy fotosintez qiluvchi organizmdan iborat, odatda a yashil suv o'tlari yoki siyanobakteriya, qo'ziqorin filamentlari bilan o'ralgan. Odatda, likenning asosiy qismi o'zaro to'qilgan qo'ziqorin filamentlaridan iborat,[34] ipli va jelatinli likenlarda bo'lsa ham[25] bunday emas. Qo'ziqorin a mikobiont. Fotosintez qiluvchi organizm a fotobiont. Algal fotobiontslari deyiladi fikobiontlar.[35] Siyanobakteriyalar fotobiontlari deyiladi siyanobionts.[35]
Ko'payish bilan bog'liq bo'lmagan likenning qismi, likenning "tanasi" yoki "vegetativ to'qimasi" deyiladi. talus. Talus shakli qo'ziqorin yoki yosun alohida o'sadigan har qanday shakldan juda farq qiladi. Talus qo'ziqorinning chaqirilgan filamentlaridan iborat gifalar. Iplar tarvaqaylab o'sadi, so'ngra birlashib to'r hosil qiladi, "bo'lish" deb nomlanadianastomoz ". Qo'ziqorin iplarining meshi zich yoki bo'shashgan bo'lishi mumkin.
Odatda, qo'ziqorin meshi alg yoki atrofini o'rab oladi siyanobakterial hujayralar, ko'pincha ularni liken uyushmalariga xos bo'lgan murakkab qo'ziqorin to'qimalari ichiga qamrab oladi. Tallda zich joylashgan, ko'pincha ikkinchi qo'ziqorin turini o'z ichiga oladigan, zich o'ralgan qo'ziqorin iplaridan himoya qiluvchi "terisi" bo'lishi mumkin yoki bo'lmasligi mumkin,[1] deyiladi a korteks. Frutikoz likenlari "shoxlari" atrofida o'ralgan bitta korteks qatlamiga ega. Folioz likenlari "barg" ning yuqori qismida yuqori korteksga ega, pastki qismida esa alohida pastki korteksga ega. Qisqichbaqasimon va skuamulyoz likenlari faqat yuqori korteksga ega bo'lib, likenning "ichki qismi" ular o'sadigan sirt bilan bevosita aloqada bo'ladi ( substrat). Agar qirralar substratdan yirtilib, tekis va bargga o'xshab ko'rinsa ham, folioz likenlaridan farqli o'laroq, pastki korteksga ega emaslar. Ipli, bussoid, moxov,[29] jelatinli va boshqa likenlarning kortekslari yo'q, ular mavjudlik deb nomlanadi ekortikat.[36]

a) korteks qattiq to'qilgan qo'ziqorin iplarining tashqi qatlami (gifalar )
b) bu fotobiont qatlami fotosintezga ega yashil suv o'tlari
v) medulla ichida bo'shashgan gifalar
d) mahkam to'qilgan pastki korteks
e) gifalarni bog'lash ildizpoyalar bu erda qo'ziqorin substratga yopishadi
Frutikoz, folioz, kustoza va skuamulyoz likenlari odatda uch xil to'qimalarga ega, farqlangan qo'ziqorin filamentlarining turli xil zichligiga ega bo'lish orqali.[34] Liken atrof-muhit bilan aloqa qiladigan yuqori qatlam a deb ataladi korteks.[34] Korteks zich qilib to'qilgan, o'ralgan va bir-biriga yopishtirilgan (aglutinatsiyalangan ) qo'ziqorin iplari.[34] Zich o'rash korteksni himoya qiluvchi "teriga" o'xshatib, boshqa organizmlarni chetga surib qo'yadi va quyida joylashgan qatlamlarga quyosh nurlari intensivligini pasaytiradi.[34] Korteks qatlami bir necha yuz mikrometrgacha (mkm) qalinligi (millimetrdan kam) bo'lishi mumkin.[37] Korteks ustiga hujayralar emas, balki sekretsiyalar epikorteksi qo'shilishi mumkin, qalinligi 0,6-1 mm. ba'zi likonlar.[37] Ushbu sekretsiya qatlamida teshiklar bo'lishi mumkin yoki bo'lmasligi mumkin.[37]
Korteks qatlami ostida - deb nomlangan qatlam mavjud fotobiontik qatlam yoki simbion qatlami.[27][34] Simbiont qatlami zichroq o'ralgan qo'ziqorin filamentlariga ega, ularga fotosintez sherigi singdirilgan.[34] Kamroq zichroq o'rash fotosintez paytida bargning anatomiyasiga o'xshash havo aylanishiga imkon beradi.[34] Fotobiontning har bir hujayrasi yoki hujayralari guruhi, odatda, gifalar bilan alohida o'raladi va ba'zi hollarda gustorium.[25] Qisqichbaqasimon va foliozli likenlarda fotobiontik qatlamdagi suv o'tlari qo'ziqorin filamentlari orasida tarqalib, quyida joylashgan qatlamga o'tishda kamayadi. Frutikoz likenlarida fotobiontik qatlam quyida joylashgan qatlamdan keskin farq qiladi.[25]
Simbiont qatlami ostidagi qavat deyiladi medulla. Medulla qo'ziqorin iplari bilan yuqoridagi qatlamlarga qaraganda kamroq zich joylashgan. Folioz likenlarida odatda bo'lgani kabi Peltigera,[30]:159 pastki korteks deb nomlangan qo'ziqorin filamentlarining yana bir zich qadoqlangan qatlami.[29][34] Ildizga o'xshash qo'ziqorin tuzilmalari deyiladi ildizpoyalar (odatda )[30]:159 likenni substratga yopishtirish yoki bog'lab qo'yish uchun pastki korteksdan o'sadi.[2][29] Frutikoz likenlari "poya" va "novdalar" atrofida butun o'ralgan bitta korteksga ega.[30] Medulla eng pastki qatlam bo'lib, shoxsimon talus uchun paxtasimon oq ichki yadro hosil qilishi yoki ichi bo'sh bo'lishi mumkin.[30]:159 Qisqichbaqasimon va skuamulyoz likenlari pastki korteksga ega emas va medulla to'g'ridan-to'g'ri aloqada bo'ladi substrat liken o'sadi.
Qisqichbaqasimon izolatli likenlarda, izolalarning qirralari substratdan tozalanadi va bargli ko'rinadi. Skuamulyoz likenlarida liken talalining substratga yopishmagan qismi ham bargli ko'rinishi mumkin. Ammo bu bargli qismlarda pastki po'stlog'i yo'q, bu esa qobiq va skuamulyoz likenlarini folioz likenlaridan ajratib turadi.[34] Aksincha, folioz likenlari substratga qarama-qarshi lichinka singari yassilangan bo'lib ko'rinishi mumkin, ammo bargga o'xshash loblarning aksariyati substratdan yuqoriga ko'tarilishi mumkin, chunki u mahkam o'ralgan pastki korteks bilan ajralib turadi.[29]
Jelatinli,[30]:159 byssoid va moxov likenlari korteksga ega emas (ular) ekortikat ), va umuman olganda, faqat simbiont qatlamiga o'xshash farqlanmagan to'qimalarga ega.[iqtibos kerak ]
Ikkala yashil alg o'z ichiga olgan likenlarda va siyanobakteriyalar, siyanobakteriyalar yuqori yoki pastki yuzada kichik pustulalarda tutilishi mumkin. sefalodiya.
Pruiniya yuqori sirt ustida oqartuvchi qoplama.[38] An epinekral qatlam "aniqlanmagan shoxli o'lik qo'ziqorin gifasi qatlami lumina suv o'tlari qatlami ustidagi korteks ichida yoki yaqinida ".[38]
2016 yil avgust oyida makrolixenlarning to'qimalarida bir nechta qo'ziqorin turlari borligi haqida xabar berilgan edi.[1]
Fiziologiya
Simbiyotik munosabat
Likenlar - qishloq xo'jaligini kashf etgan qo'ziqorinlar
— Trevor Govard[39]
Liken - bu paydo bo'lgan kompozitsion organizm suv o'tlari yoki siyanobakteriyalar iplar orasida yashash (gifalar ) ning qo'ziqorinlar o'zaro manfaatli simbiyotik munosabatlar. Qo'ziqorinlar suv o'tlari yoki siyanobakteriyalar orqali hosil bo'lgan uglevodlardan foyda ko'radi fotosintez. Yosunlar yoki siyanobakteriyalar, atrof-muhitdan namlik va ozuqa moddalarini to'playdigan va (odatda) unga langar beradigan qo'ziqorin filamentlari tomonidan atrof-muhitdan himoyalanish orqali foyda oladi. Liken tarkibidagi ba'zi fotosintetik sheriklar liken tashqarisida yashashi mumkin bo'lsa-da, liken simbiyotik assotsiatsiyasi ikkala sherikning ekologik doirasini kengaytiradi, shu bilan liken uyushmalarining ko'pgina tavsiflari ularni simbiyotik deb ta'riflaydi. Biroq, simbiyotik bo'lsa-da, munosabatlar ehtimol emas mututeristik, chunki suv o'tlari o'zlarining nomutanosib shakar miqdoridan voz kechishadi (pastga qarang). Ikkala sherik ham suv va mineral ozuqalarni asosan atmosferadan, yomg'ir va chang orqali oladi. Qo'ziqorin sherigi algni suvni ushlab, mineral ozuqa moddalarini olish uchun ko'proq maydon sifatida xizmat qiladi va ba'zi holatlarda olingan minerallarni ta'minlaydi. substrat. Agar a siyanobakteriya mavjud bo'lib, ba'zi uch tomonlama likenlarda bo'lgani kabi, yashil suvo'tga qo'shimcha ravishda asosiy sherik yoki boshqa simbiont sifatida ular atmosfera azotini tuzatish, yashil suv o'tlari faoliyatini to'ldiruvchi.
Uch xil naslda qo'ziqorin sherigi mitoxondriyal energiya ishlab chiqarishda asosiy funktsiyalarga ega bo'lgan atp9 mitokondriyal genini mustaqil ravishda yo'qotdi. Yo'qotish qo'ziqorinlarni o'zlarining simbionlariga to'liq bog'liq qiladi.[40]
Alg yoki siyanobakterial hujayralar fotosintez va o'simliklardagi kabi, ular kamaytirish atmosfera karbonat angidrid ikkala simbiontni oziqlantirish uchun organik uglerod qandlariga. Fikobiontlar (suv o'tlari) hosil qiladi shakar spirtlari (ribitol, sorbitol va eritritol ), ular mikobiont (qo'ziqorin) tomonidan so'riladi.[35] Cyanobionts ishlab chiqaradi glyukoza.[35] Litsenziyalangan qo'ziqorin hujayralari fotosintez mahsulotlaridan fotobiontni "oqishi" mumkin, keyin ular qo'ziqorin tomonidan so'rilishi mumkin.[7]:5
Likenlarning ko'pchiligi, ehtimol ularning aksariyati, simbiyotik munosabatda bo'lib, Cyphobasidiales deb nomlangan basidiomitset xamirturushlar buyrug'i bilan yashaydilar. Ushbu uchinchi sherikning yo'qligi laboratoriyada liken etishtirishning qiyinchiliklarini tushuntirishi mumkin. Xamirturush xujayralari liken tallining xarakterli korteksini shakllantirish uchun javobgardir va uning shakli uchun ham muhim bo'lishi mumkin.[41]
Alga yoki siyanobakteriyalarning qo'ziqorin bilan liken birikmasi tabiiy ravishda yoki madaniy ravishda o'z-o'zidan o'sadigan qo'ziqorin, alga yoki siyanobakteriyalarning tarkibiy qismiga qaraganda juda boshqacha shakli (morfologiyasi), fiziologiyasi va biokimyosiga ega. Tana (talus ) ko'pgina likenlarning qo'ziqorinlari yoki alg alohida o'sadiganlaridan farq qiladi. Laboratoriyada fotobiont bo'lmagan holda o'stirilganda, liken qo'ziqorinlari qo'ziqorin filamentlarining strukturasiz, farqlanmagan massasi sifatida rivojlanadi (gifalar ). Agar tegishli sharoitda uning fotobionti bilan birlashtirilsa, fotobiont bilan bog'liq bo'lgan xarakterli shakl paydo bo'ladi, bu jarayonda morfogenez.[4] Bir nechta ajoyib holatlarda bitta liken zamburug'i yashil suv o'tlari yoki siyanobakterial simbiont bilan birlashganda juda xilma-xil ikkita liken shaklida paydo bo'lishi mumkin. Tabiiyki, ushbu muqobil shakllar dastlab bir-biriga o'xshash tarzda o'sib borguniga qadar turli xil turlar deb qaraldi.
Likenlarning muvaffaqiyatli namunalari ekanligiga dalillar simbiyoz likenler sayyoramizning deyarli barcha yashash joylari va geografik hududlarida mavjud bo'lishi haqiqatdir.[12] Yashil yosunlarning ikki turidagi ikkita tur barcha likenlarning 35% dan ko'prog'ida uchraydi, ammo kamdan-kam hollarda o'zlarining likenidan tashqarida yashovchilarni topish mumkin.[42]
Bir vaqtning o'zida bitta qo'ziqorin sherigi turli xil iqlim sharoitida bir-biridan ustun bo'lgan ikkita yashil suv o'tlari sherigiga ega bo'lsa, bu bir vaqtning o'zida bir nechta fotosintetik sherikga ega bo'lishini ko'rsatishi mumkin, bu likenning yashash joylari va geografik joylashuvlarining keng doiralarida mavjud bo'lishi mumkin.[12]
Lichenning kamida bitta shakli, Shimoliy Amerikadagi soqolga o'xshash likonlar, ikkita emas, balki uchta simbiyotik sherikdan iborat: ascomitsetous qo'ziqorin, fotosintezli alg va kutilmaganda basidiomitsetli xamirturush.[43]
Fikobiontlar faqat suv bug'lari bilan shakarlarning aniq chiqishiga ega bo'lishi mumkin.[35] Tsalyobiontlarning fotosintez qilishlari uchun tall suyuq suv bilan to'yingan bo'lishi kerak.[35]
Yosunlar shakarlarni ishlab chiqaradi, ular qo'ziqorin tomonidan diffuziya orqali so'rilgan maxsus qo'ziqorin gifalariga singib ketadi appressiya yoki gustoriya suv o'tlari hujayralarining devori bilan aloqa qilish.[44] Appressoriya yoki haustoriya suv o'tlari xujayrasi devorlarining o'tkazuvchanligini oshiradigan moddalarni ishlab chiqarishi va devorlarga kirib borishi mumkin.[44] Yosunlar qo'ziqorin tufayli shakar ishlab chiqarishning 80% gacha yo'qotishi mumkin.[44]
Ekologiya
Liken uyushmalari misol bo'lishi mumkin mutalizm, komensalizm yoki hatto parazitizm, turlarga qarab. Liken simbiozining mavjudligini ko'rsatadigan dalillar mavjud parazit yoki komensalistik, dan ko'ra mututeristik.[45] Fotosintetik sherik tabiatda qo'ziqorin sherigidan mustaqil ravishda mavjud bo'lishi mumkin, aksincha emas. Fotobiont hujayralari muntazam ravishda yo'q qilinadi ozuqa moddasi almashish. Assotsiatsiya davom etishi mumkin, chunki fotobiont hujayralarining ko'payishi ularning yo'q bo'lish tezligiga mos keladi.[45] The qo'ziqorin suv o'tlari hujayralarini o'rab oladi,[8] ko'pincha ularni liken uyushmalariga xos bo'lgan murakkab qo'ziqorin to'qimalari ichiga qamrab oladi. Ko'p turlarda qo'ziqorin alg hujayra devoriga kirib boradi,[8] penetratsion qoziqlar hosil qilish (gustoriya ) tomonidan ishlab chiqarilganlarga o'xshash patogen zamburug'lar xost bilan oziqlanadigan.[28][46] Siyanobakteriyalar laboratoriya sharoitida liken tarkibiga kirgandan ko'ra, yolg'iz qolganda tezroq o'sishi mumkin.
Miniatyura ekotizimi va golobiont nazariyasi
Lishayniklarda simbioz shu qadar muvozanatliki, lishayniklar o'zlari va o'zlari uchun nisbatan mustaqil miniatyura ekotizimi deb hisoblangan.[12][13] Lishayniklar yanada murakkab simbiotik tizimlar bo'lishi mumkin, ular tarkibiga fotosentetik bo'lmagan bakterial jamoalar kiradi, ular boshqa funktsiyalarni sherik sifatida bajaradilar. holobiont.[14][15]
Ko'pgina likenler atrof-muhitning buzilishiga juda sezgir va ularni arzonroq ishlatish mumkin[8] baholash havoning ifloslanishi,[47][48][49] ozon tükenme va metall bilan ifloslanish. Lishayniklar tayyorlashda ishlatilgan bo'yoqlar, atirlar,[50] va an'anaviy dorilar. Bir nechta liken turlari hasharotlar tomonidan iste'mol qilinadi[8] yoki kiyik kabi katta hayvonlar.[51] Likenler atrof-muhit ko'rsatkichlari yoki bio-ko'rsatkichlar sifatida keng qo'llaniladi. Havo oltingugurt dioksidi bilan juda yomon ifloslanganida, hech qanday liken bo'lmasligi mumkin; faqat ba'zi yashil suv o'tlari bu sharoitlarga toqat qila oladi. Agar havo toza bo'lsa, unda buta, tukli va bargli likenler ko'payadi. Bir nechta liken turlari ifloslanishning yuqori darajasiga toqat qila oladi va odatda shahar joylarda, yo'laklarda, devorlarda va daraxt qobig'ida uchraydi. Eng sezgir lishayniklar buta va bargli, eng bardoshli lishayniklar esa tashqi ko'rinishiga ko'ra po'stloqdir. Sanoatlashtirishdan beri ko'plab buta va bargli likenler Ramalina, Usnea va Lobariya turlari juda cheklangan diapazonga ega, ko'pincha ular eng toza havoga ega bo'lgan joylar bilan chegaralanadi.
Likenikolozli qo'ziqorinlar
Ba'zi qo'ziqorinlarni faqat tirik holda topish mumkin kuni likenlar kabi majburiy parazitlar. Ular quyidagicha nomlanadi likenikoloz zamburug'lar, va liken ichida yashovchi qo'ziqorinlardan farqli turlar; shuning uchun ular likenning bir qismi deb hisoblanmaydi.[52]
Suvga reaktsiya
Namlik korteksni yanada shaffof qiladi.[7]:4 Shunday qilib, suv o'tlari namlik mavjud bo'lganda fotosintez o'tkazishi mumkin va boshqa paytlarda himoya qilinadi. Korteks yanada shaffof bo'lsa, suv o'tlari yanada aniqroq namoyon bo'ladi va liken yashilroq ko'rinadi.
Metabolitlar, metabolit tuzilmalari va bioaktivlik
Likenler kuchli antioksidant faollikni ko'rsatishi mumkin.[53][54] Ikkilamchi metabolitlar ko'pincha kristall sifatida yotadi apoplast.[55] Ikkilamchi metabolitlar ba'zi substratlarni boshqalaridan ustun qo'yishda rol o'ynaydi deb o'ylashadi.[55]
Ba'zida likenlarda qo'ziqorinlardan tayyorlangan tuzilmalar mavjud metabolitlar, masalan, qisqichbaqasimon likonlar ba'zan a ga ega polisakkarid[tushuntirish kerak ] korteksdagi qatlam.[iqtibos kerak ]
O'sish darajasi
Lishayniklar odatda muntazam, ammo juda sekin o'sish sur'ati yiliga millimetrdan kam.
Qisqichbaqasimon likenlarda chekka bo'ylab eng faol o'sish sodir bo'lgan joy.[30]:159 Ko'pgina qobiq lishayniklari yiliga atigi 1-2 mm o'sadi.
Hayot davomiyligi
Likenler bo'lishi mumkin uzoq umr ko'rgan, ba'zilari esa eng qadimgi tirik organizmlar qatoriga kiritilgan.[4][16] Umrni o'lchash qiyin, chunki "bir xil" individual likenni aniqlaydigan narsa aniq emas.[56] Likenler vegetativ ravishda "bir xil" liken deb ta'riflangan yoki bo'lmasligi mumkin bo'lgan bir bo'lakni sindirish yo'li bilan o'sadi va ikkita liken birlashib, keyin "bir xil" likenga aylanadi.[56] An Arktika "xarita liken" deb nomlangan turlar (Rhizocarpon geographicum ) 8600 yilda, aftidan dunyodagi eng keksa tirik organizm sanaladi.[57]
Atrof-muhitdagi stressga javob
O'simliklar va hayvonlardagi oddiy suvsizlanishdan farqli o'laroq, likonlar a to'liq quruq davrda tana suvining yo'qolishi.[8] Likenler juda past darajalarda omon qolishga qodir suv tarkib (poikilohidrik ).[58]:5–6 Ular yana suv paydo bo'lganda, ular tezda so'riladi, yumshoq va go'shtli bo'ladi.[8] Suvsizlanish davridan keyin membranalarni qayta tuzish bir necha daqiqa yoki undan ko'proq vaqtni talab qiladi.[iqtibos kerak ]
Sinovlarda liken omon qoldi va ajoyib natijalarni ko'rsatdi moslashish qobiliyati ning fotosintez faoliyati ichida simulyatsiya vaqti 34 kun ichida Mars sharoiti tomonidan qo'llab-quvvatlanadigan Mars simulyatsiyasi laboratoriyasida (MSL) Germaniya aerokosmik markazi (DLR).[59][60]
The Evropa kosmik agentligi likenlarning kosmosda himoyasiz yashashi mumkinligini aniqladi. Madridning Complutense universiteti xodimi Leopoldo Sancho boshchiligidagi ikki xil liken turi -Rhizocarpon geographicum va Xanthoria elegans - kapsulada muhrlangan va Rossiyaning "Soyuz" raketasida 2005 yil 31 mayda uchirilgan. Orbitada bo'lganidan keyin kapsulalar ochilib, likenlar keng o'zgaruvchan harorat va kosmik nurlanish bilan kosmik vakuumga bevosita ta'sir ko'rsatdi. 15 kundan so'ng, likonlar erga qaytarildi va fotosintez qilish qobiliyatlari o'zgarmas ekanligi aniqlandi.[61][62]
Ko'paytirish va tarqatish
Vegetativ ko'payish

Ko'pgina likenler jinssiz ravishda ko'payadi, yoki parchalanib o'z-o'zidan o'sib chiqadi (vegetativ ko'payish ) yoki tarqalishi orqali diasporalar qo'ziqorin hujayralari bilan o'ralgan bir necha alg hujayralarini o'z ichiga oladi.[2] Tallda farqlanishning nisbatan kamligi sababli diasporaning shakllanishi va vegetativ ko'payish o'rtasidagi chiziq ko'pincha xiralashgan. Frutikoz likenlari osonlikcha mumkin[iqtibos kerak ] bo'lakdan yangi likenlar o'sishi mumkin (vegetativ ko'payish ).[iqtibos kerak ] Ko'p likenler quriganida bo'laklarga bo'linib, shamol ta'sirida tarqalib, namlik qaytib kelganda o'sishni davom ettiradi.[63][64] Soreiya (singular: "soredium") soraliya deb nomlangan tuzilmalarda hosil bo'lgan qo'ziqorin filamentlari bilan o'ralgan suv o'tlari hujayralarining kichik guruhlari bo'lib, ulardan soredia shamol bilan tarqalishi mumkin.[2] Isidiya (birlik: "izidiy") mexanik tarqalish uchun ajralib chiqadigan tarladan tarvaqaylab ketgan, tikanli, cho'zilgan, o'sib chiqqan.[2] Liken tarqaladi (diasporalar ) odatda ikkala sherikning hujayralarini o'z ichiga oladi, ammo "chekka turlar" deb ataladigan qo'ziqorin tarkibiy qismlari "yadro turlari" tomonidan tarqalgan alg hujayralariga tayanadi.[65]
Jinsiy ko'payish

Ko'paytirish bilan bog'liq tuzilmalar ko'pincha talus yuzasida disklar, tepaliklar yoki silliq chiziqlar ko'rinishida bo'ladi.[7]:4 Fotobiontda jinsiy reproduktsiya tanlangan deb ta'kidlangan bo'lsa-da, meiotik faoliyatni (jinsiy reproduktsiya) ko'rsatadigan kuchli dalillar mavjud Trebuksiya.[66][67] Ko'p liken zamburug'lari boshqa qo'ziqorinlar singari jinsiy yo'l bilan ko'payadi va hosil bo'lgan sporalarni hosil qiladi mayoz va gametalarning birlashishi. Tarqoqlikdan so'ng, bunday qo'ziqorin sporalari funktsional liken paydo bo'lishidan oldin mos keladigan alg sherigi bilan uchrashishi kerak.
Ba'zi liken zamburug'lari tegishli Basidiomitsetalar (basidiolixenalar) va ishlab chiqarish qo'ziqorin ularning qarindoshlariga o'xshash reproduktiv tuzilmalarga o'xshash.
Liken zamburug'larining aksariyati tegishli Ascomitsetes (askolikalar). Askolikenlar orasida sporlar deb nomlangan spora hosil qiluvchi inshootlarda ishlab chiqariladi ascomata.[7] Ascomataning eng keng tarqalgan turlari bu apotetsiya (ko'plik: apothecia) va peritetsiy (ko'plik: perithecia).[7]:14 Apotexiya odatda liken talalining yuqori yuzasida joylashgan stakan yoki plastinkaga o'xshash disklardir. Apotexiya disklar singari chayqalgan chiziqli segmentlar kabi shakllanganda, ular deyiladi lirellar.[7]:14 Perithecia shakli liken tallasi to'qimalariga botgan kolbalar shaklida bo'lib, ular kolbadan chiqib ketadigan sporalari uchun kichik teshikka ega va liken yuzasida qora nuqta kabi ko'rinadi.[7]:14
Eng keng tarqalgan uchta sport tanasi turi ko'tarilgan disklardir apotexiya (singular: apothecium), shishaga o'xshash stakan, tepasida kichik teshik bor peritexiya (birlik: perithecium) va piknidiya (birlik: piknidiy), peritetsiyaga o'xshash, ammo asci bo'lmagan (an.) ascus zamburug'laridagi jinsiy sporalarni o'z ichiga olgan va chiqaradigan tuzilishdir Ascomycota ).[68]
Apothetsiyada spora hosil qiluvchi hujayralar qatlami bor, ular deyiladi asci (singular: ascus), va odatda talus to'qimasidan farq qiladi.[7]:14 Apotetsiya tashqi chekkaga ega bo'lganda, chekka deyiladi imtiyoz.[7]:14 Eksipel rangli talus to'qimalariga o'xshash rangga ega bo'lganda, apotesiya yoki liken deyiladi leanorin, tur vakillariga o'xshash ma'no Lekanora.[7]:14 Eksipel uglerod singari qorayganda, u deyiladi lesidin turkum a'zolariga o'xshash ma'no Lesidiya.[7]:14 Marj rangsiz yoki rangsiz bo'lsa, u deyiladi biatorin.[7]:14

A "podium "(ko'plik: podetiya ) - bu talumdan ko'tarilgan meva tanasining likenlangan poyaga o'xshash tuzilishi, qo'ziqorin hosil qiladigan ba'zi qo'ziqorinlar bilan bog'liq apotetsiya.[27] Reproduktiv to'qimalarning bir qismi bo'lganligi sababli, podetiya asosiy tananing (talus) qismi deb hisoblanmaydi, lekin ingl.[27] Podium shoxlangan, ba'zan esa kosaga o'xshash bo'lishi mumkin. Ular odatda qo'ziqorinni ko'taradilar piknidiya yoki apotexiya yoki ikkalasi ham.[27] Ko'p liken bor apotexiya yalang'och ko'z bilan ko'rinadigan.[2]
Ko'pgina likenler mo'l-ko'l jinsiy tuzilmalarni ishlab chiqaradi.[69] Ko'pgina turlar faqat jinsiy spora orqali tarqaladi.[69] Masalan, qobiq lishayniklari Grafis skriptlari va Ochrolechia parella simbiyotik vegetativ tarqalish hosil qilmaydi. Buning o'rniga, bu turlarning liken hosil qiluvchi qo'ziqorinlari o'z-o'zini urug'lantirish orqali jinsiy ravishda ko'payadi (ya'ni ular gomotalik ). Ushbu naslchilik tizimi og'ir muhitda muvaffaqiyatli ko'paytirishni ta'minlashi mumkin.[69]
Mazaedia (singular: mazaedium) a ga o'xshash apothecia tikuvchilik pimi ichida (pin liken ) s, bu erda meva tanasi mayda sopi ustiga o'tirgan, kosachasimon eksiple bilan o'ralgan, bo'shashgan ascosporesning jigarrang yoki qora massasi.[7]:15
Taksonomiya va tasnif
Likenler qo'ziqorin komponenti bo'yicha tasniflanadi. Liken turlariga bir xil ilmiy nom berilgan (binomial ism ) liken tarkibidagi qo'ziqorin turlari sifatida. Likenler qo'ziqorinlarni tasniflash sxemalariga kiritilmoqda. Yosun o'zining ilmiy nomiga ega, bu liken yoki qo'ziqorin bilan hech qanday aloqasi yo'q.[70] Taxminan 13500-17000 liken turlari aniqlangan.[44] Ma'lum bo'lgan qo'ziqorin turlarining deyarli 20% likenler bilan bog'liq.[44]
"Likenlangan qo'ziqorin"butun likenni yoki faqat qo'ziqorinni nazarda tutishi mumkin. Bu kontekstsiz chalkashliklarni keltirib chiqarishi mumkin. Muayyan qo'ziqorin turlari turli xil suv o'tlari turlari bilan likenlarni hosil qilishi mumkin, chunki ular turli xil liken turlari bo'lib tuyuladi, ammo ular hali ham tasniflanadi ( 2014 yildan boshlab) xuddi shu liken turlari kabi.[71]
Ilgari, ba'zi liken taksonomistlari likenlarni o'zlarining bo'linmalariga joylashtirdilar Mikofikofa, ammo bu amaliyot endi qabul qilinmaydi, chunki tarkibiy qismlar alohida-alohida nasablar. Askolikalar ham, basidiolixenlar ham shakllanmaydi monofiletik o'zlarining qo'ziqorin filasida nasabnomalar, lekin ular har bir filum ichida bir nechta asosiy yoki asosan liken hosil qiluvchi guruhlarni hosil qiladi.[72] Basidiolixenlardan ham g'ayrioddiy qo'ziqorin Geosiphon pyriforme, a'zosi Glomeromikota bu hujayralar ichida siyanobakterial simbiontni qamrab olishi bilan o'ziga xosdir. Geosifhon odatda liken deb hisoblanmaydi va uning o'ziga xos simbiozi ko'p yillar davomida tan olinmagan. Jins yanada yaqinroq ittifoqdosh endomikorizal avlodlar. Qo'ziqorinlar Verrucariales bilan dengiz likenlarini hosil qiladi jigarrang suv o'tlari Petroderma maculiforme,[73] va bilan simbiyotik munosabatda bo'lish dengiz o'tlari kabi (toshbo'ron ) va Blidingia minima, bu erda suv o'tlari dominant tarkibiy qismlardir. Qo'ziqorinlar toshbo'ronlarga havo ta'sirida qurib qolishiga qarshi turishga yordam beradi deb o'ylashadi.[74][75] Bundan tashqari, likenler ham foydalanishlari mumkin sariq-yashil suv o'tlari (Heterokokk ) ularning simbiyotik sherigi sifatida.[76]
Lichens mustaqil ravishda tarix davomida bir necha marta suv o'tlari va siyanobakteriyalar bilan bog'langan qo'ziqorinlardan paydo bo'lgan.[77]
Qo'ziqorinlar
Likenning qo'ziqorin tarkibiy qismi mikobiont. Mikobiont an bo'lishi mumkin Ascomycete yoki Basidiomitset.[10] Bog'langan likenler ham deyiladi askolikalar yoki basidiolixenalar navbati bilan. Sifatida yashash simbiont liken qo'ziqorini uchun zarur bo'lgan ozuqaviy moddalarni olishning muvaffaqiyatli usuli hisoblanadi, chunki barcha qo'ziqorin turlarining taxminan 20% ushbu hayot tarziga ega bo'lgan.[78]
Turli xil sheriklar bilan ma'lum qo'ziqorin simbionti tomonidan ishlab chiqarilgan talli o'xshash bo'lishi mumkin,[iqtibos kerak ] va ikkinchi darajali metabolitlar bir xil,[iqtibos kerak ] ko'rsatuvchi[iqtibos kerak ] liken morfologiyasini aniqlashda qo'ziqorin dominant rolga ega ekanligi. Ammo turli xil fotobiontlarga ega bo'lgan bir xil mikobiont ham juda xilma-xil o'sish shakllarini hosil qilishi mumkin.[71] Likenler ma'lumki, unda ikki yoki hatto uchta alg turlari bilan bog'liq bo'lgan bitta qo'ziqorin mavjud.
Garchi har bir liken talusi odatda bir hil bo'lib ko'rinsa-da, ba'zi dalillar qo'ziqorin tarkibiy qismi ushbu turning bir nechta genetik shaxsidan iborat bo'lishi mumkin degan fikrga o'xshaydi.[iqtibos kerak ]
Ikki yoki undan ortiq qo'ziqorin turlari o'zaro ta'sirlashib, bir xil likenni hosil qilishi mumkin.[79]
Quyidagi jadvalda buyurtmalar va oilalar liken hosil qiluvchi turlarni o'z ichiga olgan qo'ziqorinlar.
Photobionts
The fotosintez partner in a lichen is called a fotobiont. The photobionts in lichens come from a variety of simple prokaryotik va ökaryotik organizmlar. In the majority of lichens the photobiont is a green alga (Xlorofit ) yoki a siyanobakteriya. In some lichens both types are present. Algal photobionts are called fikobiontlar, while cyanobacterial photobionts are called cyanobionts.[35] According to one source, about 90% of all known lichens have phycobionts, and about 10% have cyanobionts,[35] while another source states that two thirds of lichens have green algae as phycobiont, and about one third have a cyanobiont.[28] Approximately 100 species of photosynthetic partners from 40[35] genera and five distinct classes (prokaryotic: Siyanofeylar; ökaryotik: Trebouxiophyceae, Feofitlar, Xlorofitlar ) liken hosil qiluvchi zamburug'lar bilan bog'langanligi aniqlandi.[80]
Umumiy suv o'tlari photobionts are from the genera Trebouxia, Trentepoxiya, Pseudotrebouxia, yoki Mirmecia. Trebouxia is the most common genus of green algae in lichens, occurring in about 40% of all lichens. "Trebouxioid" means either a photobiont that is in the genus Trebouxia, yoki shu tur vakiliga o'xshaydi va shuning uchun ehtimol sinf a'zosi Trebouxiophyceae.[27] Yashil yosunlarning eng ko'p tarqalgan ikkinchi turi Trentepoxiya.[28] Overall, about 100 species of eukaryotes are known to occur as photobionts in lichens. Barcha suv o'tlari, ehtimol tabiatda ham, likenda ham mustaqil ravishda mavjud bo'lishi mumkin.[79]
A "siyanolihen " is a lichen with a siyanobakteriya as its main photosynthetic component (photobiont).[81] Most cyanolichen are also ascolichens, but a few basidiolichen like Diktyonema va Acantholichen have cyanobacteria as their partner.[82]
The most commonly occurring cyanobacterium tur bu Nostok.[79] Boshqalar[28] umumiy siyanobakteriya photobionts are from Scytonema.[10] Ko'p siyanollar mayda va qora bo'lib, bor ohaktosh substrat sifatida.[iqtibos kerak ] Boshqa siyanol guruhi jele likenlari avlod Kollema yoki Leptogium are gelatinous and live on moist soils. Another group of large and foliose turlari, shu jumladan Peltigera, Lobariya va Degeliya are grey-blue, especially when dampened or wet. Many of these characterize the Lobarion g'arbiy Britaniyada, masalan, Kelt yomg'ir o'rmoni. Strains of cyanobacteria found in various cyanolichens are often closely related to one another.[83] They differ from the most closely related free-living strains.[83]
Liken assotsiatsiyasi yaqin simbiozdir. U ikkala sherikning ham ekologik doirasini kengaytiradi, lekin tabiiy muhitda ularning o'sishi va ko'payishi uchun har doim ham majburiy emas, chunki ko'pgina alg simbionlari mustaqil ravishda yashashi mumkin. Yosunlarning yorqin namunasi Trentepoxiya, which forms orange-coloured populations on tree trunks and suitable rock faces. Liken tarqaladi (diaspores ) typically contain cells from both partners, although the fungal components of so-called "fringe species" rely instead on algal cells dispersed by the "core species".[65]
The same cyanobiont species can occur in association with different fungal species as lichen partners.[84] The same phycobiont species can occur in association with different fungal species as lichen partners.[35] More than one phycobiont may be present in a single thallus.[35]
A single lichen may contain several algal genotiplar.[85][86] These multiple genotypes may better enable response to adaptation to environmental changes, and enable the lichen to inhabit a wider range of environments.[87]
Controversy over classification method and species names
There are about 20,000 known lichen turlari.[10] But what is meant by "species" is different from what is meant by biological species in plants, animals, or fungi, where being the same species implies that there is a common ajdodlar nasli.[10] Because lichens are combinations of members of two or even three different biological shohliklar, these components kerak bor boshqacha ancestral lineage from each other. By convention, lichens are still called "species" anyway, and are classified according to the species of their fungus, not the species of the algae or cyanobacteria. Lichens are given the same scientific name (binomial ism ) as the fungus in them, which may cause some confusion. The alga bears its own scientific name, which has no relationship to the name of the lichen or fungus.[70]
Depending on context, "lichenized fungus" may refer to the entire lichen, or to the fungus when it is in the lichen, which can be grown in culture in isolation from the algae or cyanobacteria. Some algae and cyanobacteria are found naturally living outside of the lichen. The fungal, algal, or cyanobacterial component of a lichen can be grown by itself in culture. When growing by themselves, the fungus, algae, or cyanobacteria have very different properties than those of the lichen. Lichen properties such as growth form, physiology, and biochemistry, are very different from the combination of the properties of the fungus and the algae or cyanobacteria.
The same fungus growing in combination with different algae or cyanobacteria, can produce lichens that are very different in most properties, meeting non-DNA criteria for being different "species". Historically, these different combinations were classified as different species. When the fungus is identified as being the same using modern DNA methods, these apparently different species get reclassified as the bir xil species under the current (2014) convention for classification by fungal component. This has led to debate about this classification convention. These apparently different "species" have their own independent evolutionary history.[2][71]
There is also debate as to the appropriateness of giving the same binomial name to the fungus, and to the lichen that combines that fungus with an alga or cyanobacterium (sinekdoxa ). This is especially the case when combining the same fungus with different algae or cyanobacteria produces dramatically different lichen organisms, which would be considered different species by any measure other than the DNA of the fungal component. If the whole lichen produced by the same fungus growing in association with different algae or cyanobacteria, were to be classified as different "species", the number of "lichen species" would be greater.
Turli xillik
Likenlangan qo'ziqorinlarning eng ko'p miqdori Ascomycota, taxminan 40% turlar bunday assotsiatsiyani tashkil qiladi.[70] Ushbu likenlangan zamburug'larning ba'zilari yashaydigan litsenziyalanmagan zamburug'lar bilan buyurtmalarda uchraydi saprotroflar yoki o'simlik parazitlari (masalan, Leotiales, Dothideales va Pezizales ). Boshqa liken qo'ziqorinlari faqat beshtasida uchraydi buyurtmalar unda barcha a'zolar ushbu odat bilan shug'ullanadilar (Buyurtmalar Grafidales, Gyalectales, Peltigerales, Pertusariales va Teloschistales ). Umuman olganda, likenlarning taxminan 98% ascomitset mikobiontga ega.[88] Ascomycota yonida eng ko'p likenlangan qo'ziqorinlar tayinlanmaganlarda uchraydi qo'ziqorinlar nomukammal, a catch-all category for fungi whose sexual form of reproduction has never been observed.[iqtibos kerak ] Comparatively few Basidiomitsetalar likenlangan, ammo ular qatoriga kiradi agariklar kabi turlari Likenomfaliya, klavarioid qo'ziqorinlari kabi turlari Multiklavula va kortitsioid qo'ziqorinlari kabi turlari Diktyonema.
Identification methods
Lichen identification uses growth form and reactions to chemical tests.
The outcome of the "Pd test" is called "Pd", which is also used as an abbreviation for the chemical used in the test, para-fenilendiamin.[27] If putting a drop on a lichen turns an area bright yellow to orange, this helps identify it as belonging to either the genus Cladonia yoki Lecanora.[27]
Evolution and paleontology
The fossil record for lichens is poor.[89] The extreme habitats that lichens dominate, such as tundra, mountains, and deserts, are not ordinarily conducive to producing fossils.[89][90] There are fossilized lichens embedded in amber. The fossilized Anziya is found in pieces of amber in northern Europe and dates back approximately 40 million years.[91] Lichen fragments are also found in fossil leaf beds, such as Lobariya from Trinity County in northern California, USA, dating back to the early to middle Miosen.[92]
The oldest fossil lichen in which both symbiotic partners have been recovered is Winfrenatiya, an early zygomycetous (Glomeromycotan ) lichen symbiosis that may have involved controlled parasitism,[iqtibos kerak ] is permineralized in the Rini Chert of Scotland, dating from early Dastlabki devoncha, about 400 million years ago.[93] The slightly older fossil Spongiofiton has also been interpreted as a lichen on morphological[94] and isotopic[95] grounds, although the isotopic basis is decidedly shaky.[96] Bu isbotlangan Siluriya -Devoniy fotoalbomlar Nematothallus[97] va Prototaksitlar[98] were lichenized. Thus lichenized Ascomycota va Basidiomycota were a component of Erta silur -Devoniy terrestrial ecosystems.[99][100] Newer research suggests that lichen evolved after the evolution of land plants.[101]
The ancestral ecological state of both Ascomycota va Basidiomycota ehtimol edi saprobism, and independent lichenization events may have occurred multiple times.[102][103] In 1995, Gargas and colleagues proposed that there were at least five independent origins of lichenization; three in the basidiomycetes and at least two in the Ascomycetes.[104] However, Lutzoni et al. (2001) indicate that lichenization probably evolved earlier and was followed by multiple independent losses. Some non-lichen-forming fungi may have secondarily lost the ability to form a lichen association. As a result, lichenization has been viewed as a highly successful nutritional strategy.[105][106]
Lichenized Glomeromikota may extend well back into the Precambrian. Lichen-like fossils consisting of coccoid cells (siyanobakteriyalar ?) and thin filaments (mucoromycotinan Glomeromikota ?) are permineralized in marine fosforit ning Doushantuo shakllanishi Xitoyning janubida. These fossils are thought to be 551 to 635 million years old or Ediakaran.[107] Ediakaran akritarxlar also have many similarities with Glomeromycotan vesicles and spores.[108] Bundan tashqari, bu da'vo qilingan Ediakaran qoldiqlari shu jumladan Diksoniya,[109] were lichens,[110] although this claim is controversial.[111] Endosimbiyotik Glomeromikota comparable with living Geosifhon may extend back into the Proterozoy in the form of 1500 million year old Horodyskia[112] and 2200 million year old Diskagma.[113] Discovery of these fossils suggest that fungi developed symbiotic partnerships with photoautotrophs long before the evolution of vascular plants, though the Ediacaran lichen hypothesisis largely rejected due to an inappropriate definition of lichens based on taphonomy and substrate ecology.[114]
Ecology and interactions with environment
Substrates and habitats

Lichens cover about 7% of the planet's surface and grow on and in a wide range of substrates and habitats, including some of the most extreme conditions on earth.[115] They are abundant growing on bark, leaves, and hanging from branches "living on thin air" (epifitlar ) ichida yomg'ir o'rmonlari va mo''tadil o'rmonzor. They grow on bare rock, walls, gravestones, roofs, and exposed soil surfaces. They can survive in some of the most extreme environments on Earth: Arktik tundra, hot dry cho'llar, rocky coasts, and toxic cüruf uyumlari. They can live inside solid rock, growing between the grains, and in the soil as part of a biologik tuproq qobig'i in arid habitats such as deserts. Some lichens do not grow on anything, living out their lives blowing about the environment.[2]
When growing on mineral surfaces, some lichens slowly decompose their substrate by chemically degrading and physically disrupting the minerals, contributing to the process of ob-havo by which rocks are gradually turned into soil. While this contribution to weathering is usually benign, it can cause problems for artificial stone structures. For example, there is an ongoing lichen growth problem on Rushmor tog'idagi milliy yodgorlik that requires the employment of mountain-climbing conservators to clean the monument.
Lichens are not parazitlar on the plants they grow on, but only use them as a substrate to grow on. The fungi of some lichen species may "take over" the algae of other lichen species.[8][116] Lichens make their own food from their photosynthetic parts and by absorbing minerals from the environment.[8] Lichens growing on leaves may have the appearance of being parasites on the leaves, but they are not. However, some lichens, notably those of the genus Diplosxistlar are known to parasitise other lichens. Diploschistes muscorum starts its development in the tissue of a host Cladonia turlari.[46]:30[28]:171
In the arctic tundra, lichens, together with moxlar va jigar jigarlari, make up the majority of the zamin qoplamasi, which helps insulate the ground and may provide forage for grazing animals. Misol "Kiyik moxi ", which is a lichen, not a moss.[8]
A crustose lichen that grows on rock is called a saxicolous lichen.[27][30]:159 Crustose lichens that grow on the rock are epilitik, and those that grow immersed inside rock, growing between the crystals with only their fruiting bodies exposed to the air, are called endolithic lichens.[26][30]:159[81] A crustose lichen that grows on bark is called a kortikolli liken.[30]:159 A lichen that grows on wood from which the bark has been stripped is called a lignikolous liken.[36] Lichens that grow immersed inside plant tissues are called endophloidic lichens yoki endophloidal lichens.[26][30]:159 Lichens that use leaves as substrates, whether the leaf is still on the tree or on the ground, are called epiphyllous yoki foliicolous.[35] A terrikololi liken grows on the soil as a substrate. Many squamulous lichens are terricolous.[30]:159 Umbillicate lichens are foliose lichens that are attached to the substrate at only one point.[26] A sersuv liken is not attached to a substrate at all, and lives its life being blown around by the wind.
Lichens and soils
In addition to distinct physical mechanisms by which lichens break down raw stone, recent studies indicate lichens attack stone chemically, entering newly chelated minerals into the ecology.
The lichen exudates, which have powerful chelating capacity, the widespread occurrence of mineral neoformation, particularly metal oxalates, together with the characteristics of weathered substrates, all confirm the significance of lichens as chemical weathering agents.[117]
Over time, this activity creates new fertile soil from lifeless stone.
Lichens may be important in contributing nitrogen to soils in some deserts through being eaten, along with their rock substrate, by snails, which then defecate, putting the nitrogen into the soils.[118] Lichens help bind and stabilize soil sand in dunes.[2] In deserts and semi-arid areas, lichens are part of extensive, living biological soil crusts, essential for maintaining the soil structure.[2] Lichens have a long fossil record in soils dating back 2.2 billion years.[113]
Ecological interactions
Lichens are kashshof turlar, among the first living things to grow on bare rock or areas denuded of life by a disaster.[2] Lichens may have to compete with plants for access to sunlight, but because of their small size and slow growth, they thrive in places where higher plants have difficulty growing. Lichens are often the first to settle in places lacking soil, constituting the sole vegetation in some extreme environments such as those found at high mountain elevations and at high latitudes.[119] Some survive in the tough conditions of deserts, and others on frozen soil of the Arctic regions.[120]
A major ecophysiological advantage of lichens is that they are poikilohydric (poikilo- variable, gidroksidi- relating to water), meaning that though they have little control over the status of their hydration, they can tolerate irregular and extended periods of severe quritish. Like some moxlar, jigar jigarlari, ferns, and a few "resurrection plants ", upon desiccation, lichens enter a metabolic suspension or stasis (known as cryptobiosis ) in which the cells of the lichen symbionts are dehydrated to a degree that halts most biochemical activity. In this cryptobiotic state, lichens can survive wider extremes of temperature, radiation and drought in the harsh environments they often inhabit.

Lichens do not have roots and do not need to tap continuous reservoirs of water like most higher plants, thus they can grow in locations impossible for most plants, such as bare rock, sterile soil or sand, and various artificial structures such as walls, roofs and monuments. Many lichens also grow as epifitlar (epi- on the surface, phyte- plant) on plants, particularly on the trunks and branches of trees. When growing on plants, lichens are not parazitlar; they do not consume any part of the plant nor poison it. Lichens produce allelopatik chemicals that inhibit the growth of mosses. Some ground-dwelling lichens, such as members of the subgenus Cladina (reindeer lichens), produce allelopathic chemicals that leach into the soil and inhibit the germination of seeds, spruce and other plants.[121] Stability (that is, longevity) of their substrat is a major factor of lichen habitats. Most lichens grow on stable rock surfaces or the bark of old trees, but many others grow on soil and sand. In these latter cases, lichens are often an important part of soil stabilization; indeed, in some desert ecosystems, vascular (higher) plant seeds cannot become established except in places where lichen crusts stabilize the sand and help retain water.
Lichens may be eaten by some animals, such as kiyik, yashash Arktika mintaqalar. The lichinkalar bir qator Lepidoptera species feed exclusively on lichens. Bunga quyidagilar kiradi oddiy piyoda va marbled beauty. However, lichens are very low in protein and high in carbohydrates, making them unsuitable for some animals. Lichens are also used by the Northern Flying Squirrel for nesting, food, and a water source during winter.
Effects of air pollution

If lichens are exposed to air pollutants at all times, without any bargli parts, they are unable to avoid the accumulation of pollutants. Also lacking stomata va a kutikula, lichens may absorb aerozollar and gases over the entire thallus surface from which they may readily tarqoq to the photobiont layer.[122] Because lichens do not possess roots, their primary source of most elementlar is the air, and therefore elemental levels in lichens often reflect the accumulated composition of ambient air. The processes by which atmospheric deposition occurs include tuman va shudring, gaseous absorption, and dry deposition.[123] Consequently, many environmental studies with lichens emphasize their feasibility as effective biomonitors of atmospheric quality.[122][124][125][126][127][128]
Not all lichens are equally sensitive to havoni ifloslantiruvchi moddalar, so different lichen species show different levels of sensitivity to specific atmospheric pollutants.[129] The sensitivity of a lichen to air pollution is directly related to the energy needs of the mycobiont, so that the stronger the dependency of the mycobiont on the photobiont, the more sensitive the lichen is to air pollution.[130] Upon exposure to air pollution, the photobiont may use metabolic energy for repair of its cellular structures that would otherwise be used for maintenance of its photosynthetic activity, therefore leaving less metabolic energy available for the mycobiont. The alteration of the balance between the photobiont and mycobiont can lead to the breakdown of the symbiotic association. Therefore, lichen decline may result not only from the accumulation of toxic substances, but also from altered nutrient supplies that favor one symbiont over the other.[122]
This interaction between lichens and air pollution has been used as a means of monitoring air quality since 1859, with more systematic methods developed by Uilyam Nylander 1866 yilda.[2]
Insondan foydalanish

Ovqat
Lichens are eaten by many different cultures across the world. Although some lichens are only eaten in times of famine, others are a staple food or even a delicacy. Two obstacles are often encountered when eating lichens: lichen polisakkaridlar are generally indigestible to humans, and lichens usually contain mildly toxic ikkilamchi birikmalar that should be removed before eating. Very few lichens are poisonous, but those high in vulpinic acid yoki usnik kislota zaharli hisoblanadi.[131] Most poisonous lichens are yellow.
Oldin, Iceland moss (Cetraria Islandica) was an important source of food for humans in northern Europe, and was cooked as a bread, porridge, pudding, soup, or salad. Vila (Bryoria fremontii) was an important food in parts of North America, where it was usually pitcooked. Northern peoples in North America and Siberia traditionally eat the partially digested reindeer lichen (Cladina spp.) after they remove it from the Rum of caribou or reindeer that have been killed. Rock tripe (Umbilicaria spp. va Lasalia spp.) is a lichen that has frequently been used as an emergency food in North America, and one species, Umbilicaria esculenta, is used in a variety of traditional Korean and Japanese foods.
Likenometriya
Lichenometry is a technique used to determine the age of exposed rock surfaces based on the size of lichen thalli. Introduced by Beschel in the 1950s,[132] the technique has found many applications. u ishlatilgan arxeologiya, paleontologiya va geomorfologiya. It uses the presumed regular but slow rate of lichen growth to determine the ochiq jinslarning yoshi.[32]:9[133] Measuring the diameter (or other size measurement) of the largest lichen of a species on a rock surface indicates the length of time since the rock surface was first exposed. Liken eski tosh yuzlarida saqlanib qolinishi mumkin[iqtibos kerak ] 10,000 years, providing the maximum age limit of the technique, though it is most accurate (within 10% error) when applied to surfaces that have been exposed for less than 1,000 years.[134] Lichenometry is especially useful for dating surfaces less than 500 years old, as radiokarbonli uchrashuv techniques are less accurate over this period.[135] Likenometriya uchun eng ko'p ishlatiladigan lishayniklar naslga tegishli Rhizocarpon (masalan, tur Rhizocarpon geographicum ) va Xanthoria.
Biologik parchalanish
Lichens have been shown to degrade polyester resins, as can be seen in archaeological sites in the Roman city of Baelo Claudia Ispaniyada.[136] Lichens can accumulate several environmental pollutants such as lead, copper, and radionuclides.[137] Some species of lichen such as Parmelia sulcata va Lobaria pulmonaria, and many in the Cladonia genus have been shown to produce serine proteases capable of the degradation of pathogenic forms of prion protein (PrP), which may be useful in treating contaminated environmental reservoirs.[138][139][140]
As dyes
Many lichens produce secondary compounds, including pigments that reduce harmful amounts of sunlight and powerful toxins that reduce o't o'simliklari or kill bacteria. These compounds are very useful for lichen identification, and have had economic importance as bo'yoqlar kabi cudbear or primitive antibiotiklar.
The pH ko'rsatkichi (indicated acidic or basic) in the litmus test is a dye extracted from the lichen Roccella tinctoria qaynatish orqali.
In the Highlands of Scotland, traditional dyes uchun Harris tweed[2] and other traditional cloths were made from lichens, including the orange Xanthoria parietina and the grey foliaceous Parmelia saxatilis common on rocks known as "crottle".
There are reports dating almost 2,000 years old of lichens being used to make purple and red dyes.[141] Of great historical and commercial significance are lichens belonging to the family Roccellaceae, commonly called orchella weed or orchil. Orcein and other lichen dyes have largely been replaced by synthetic versions.
Traditional medicine and research
Tarixiy jihatdan an'anaviy tibbiyot of Europe, Lobariya pulmonariyasi was collected in large quantities as "Lungwort", due to its lung-like appearance (the imzolar haqidagi ta'limot suggesting that herbs can treat body parts that they physically resemble). Xuddi shunday, Peltigera leykoflebi was used as a supposed cure for po'stloq, due to the resemblance of its cephalodia to the appearance of the disease.[28]
Lichens produce metabolitlar in research for their potential therapeutic or diagnostic value.[142] Some metabolites produced by lichens are structurally and functionally similar to broad-spectrum antibiotics while few are associated respectively to antiseptic similarities.[143] Usnik kislota is the most commonly studied metabolite produced by lichens.[143] It is also under research as an bactericidal agent against Escherichia coli va Staphylococcus aureus.[144]
Estetik murojaat

Colonies of lichens may be spectacular in appearance, dominating the surface of the visual landscape as part of the aesthetic appeal to visitors of Yosemit milliy bog'i va Sequoia milliy bog'i.[32]:2 apelsin va sariq lichens add to the ambience of desert trees, rock faces, tundras, and rocky seashores. Intricate webs of lichens hanging from tree branches add a mysterious aspect to forests. Fruticose lichens are used in model temir yo'l[145] and other modeling hobbies as a material for making miniature trees and shrubs.
Adabiyotda
Erta Midrashik literature, the Hebrew word "vayilafeth" in Rut 3:8 is explained as referring to Rut entwining herself around Boaz like lichen.[146] The tenth century Arab physician, Al-Tamimi, mentions lichens dissolved in sirka va gul suvi being used in his day for the treatment of skin diseases and rashes.[147]
Syujeti Jon Vindxem roman Liken bilan muammo revolves around an anti-aging chemical extracted from a lichen.
Tarix
Although lichens had been recognized as organisms for quite some time, it was not until 1867, when Swiss botanist Simon Shvendener proposed his dual theory of lichens, that lichens are a combination of fungi with algae or cyanobacteria, whereby the true nature of the lichen association began to emerge.[148] Schwendener's hypothesis, which at the time lacked experimental evidence, arose from his extensive analysis of the anatomy and development in lichens, algae, and fungi using a yorug'lik mikroskopi. Many of the leading lichenologists at the time, such as James Crombie va Nylander, rejected Schwendener's hypothesis because the common consensus was that all living organisms were autonomous.[148]
Other prominent biologists, such as Geynrix Anton de Bari, Albert Bernxard Frank, Melchior Treub va Hermann Hellriegel were not so quick to reject Schwendener's ideas and the concept soon spread into other areas of study, such as microbial, plant, animal and human pathogens.[148][149] When the complex relationships between pathogenic microorganisms and their hosts were finally identified, Schwendener's hypothesis began to gain popularity. Further experimental proof of the dual nature of lichens was obtained when Eugen Thomas published his results in 1939 on the first successful re-synthesis experiment.[148]
In the 2010s, a new facet of the fungi-algae partnership was discovered. Toby Spribille and colleagues found that many types of lichen that were long thought to be ascomycete -algae pairs were actually ascomycete-basidiomitset -algae trios.[1][150]
Galereya

Lobariya pulmonariyasi, tree lungwort, lung lichen, lung moss; Upper Bavaria, Germany

Cladonia macilenta var. bacillaris 'Lipstick Cladonia'

Usnea Avstraliya, a fruticose form, growing on a tree branch

Gipogimniya qarz tubulosa bilan Bryoriya sp. va Takermannopsis sp. Kanada qoyalaridagi

Letharia sp. bilan Bryoriya sp. on pine branches near Blackpine Lake, Washington State

Lobaria oregana, commonly called 'Lettuce lichen', in the Hoh yomg'ir o'rmoni, Vashington shtati







A lichen growing on a rock in a Brazilian cloud forest

Xanthoparmelia qarz lavicola, a foliose lichen, on basalt.

Map lichen (Rhizocarpon geographicum ) on rock

Physcia millegrana (a foliose lichen), with an unlichenized polypore fungus (bottom right), on a fallen log.

Kiyik moxi (Cladonia rangiferina)

Crustose lichens on limestone in Alta Murgia-Southern Italy

Cladonia qarz cristatella, a lichen commonly referred to as 'British Soldiers'. Notice the red tips.

A crusty crustose lichen devorda

Lichen on a lilac bush

Foliose lichens on rock growing outward and dying in the center. These lichens are at least several decades old.

Xanthoria sp. lichen on volcanic rock in Craters of the Moon National Monument (Idaho, USA)

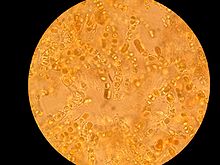
Microscopic view of lichen growing on a piece of concrete dust.[eslatma 1]













Shuningdek qarang
Izohlar
- ^ This was scraped from a dry, concrete-paved section of a drainage ditch. This entire image covers a square that is approximately 1.7 millimeters on a side. The numbered ticks on the scale represent distances of 230 micrometers, or slightly less than 0.25 millimeter.
Adabiyotlar
- ^ a b v d Spribil, Tobi; Tuovinen, Veera; Resl, Philipp; Vanderpool, Dan; Wolinski, Heimo; Aime, M. Catherine; Schneider, Kevin; Stabentheiner, Edith; Toome-Heller, Merje (21 July 2016). "Basidiomycete yeasts in the cortex of ascomycete macrolichens". Ilm-fan. 353 (6298): 488–92. Bibcode:2016Sci...353..488S. doi:10.1126/science.aaf8287. ISSN 0036-8075. PMC 5793994. PMID 27445309.
- ^ a b v d e f g h men j k l m n o "Liken nima?". Avstraliya milliy botanika bog'lari. Arxivlandi asl nusxasi 2014 yil 2-iyulda. Olingan 10 oktyabr 2014.
- ^ Introduction to Lichens – An Alliance between Kingdoms Arxivlandi 2014 yil 22-avgust Orqaga qaytish mashinasi. University of California Museum of Paleontology.
- ^ a b v d e Brodo, Irwin M. and Duran Sharnoff, Sylvia (2001) Shimoliy Amerikaning likenlari. ISBN 978-0300082494.
- ^ a b Galloway, D.J. (1999 yil 13-may). "Lichen Glossary". Avstraliya milliy botanika bog'lari. Arxivlandi asl nusxasi on 6 December 2014.
- ^ Margulis, Lin; Barreno, EVA (2003). "Likenlarga qarab". BioScience. 53 (8): 776. doi:10.1641 / 0006-3568 (2003) 053 [0776: LAL] 2.0.CO; 2.
- ^ a b v d e f g h men j k l m n o p q r Sharnoff, Stephen (2014) Kaliforniya likenlari uchun dala qo'llanmasi, Yel universiteti matbuoti. ISBN 978-0-300-19500-2
- ^ a b v d e f g h men j k Speer, Brian R; Ben Waggoner (May 1997). "Lichens: Life History & Ecology". University of California Museum of Paleontology. Arxivlandi asl nusxasidan 2015 yil 2 mayda. Olingan 28 aprel 2015.
- ^ Asplund, Yoxan; Wardle, David A. (11 October 2016). "How lichens impact on terrestrial community and ecosystem properties". Biologik sharhlar. 92 (3): 1720–1738. doi:10.1111 / brv.12305. hdl:11250/2578209. ISSN 1464-7931. PMID 27730713. S2CID 25453156.
- ^ a b v d e "Lichens: Systematics, University of California Museum of Paleontology". Arxivlandi asl nusxasidan 2015 yil 24 fevralda. Olingan 10 oktyabr 2014.
- ^ Lendemer, J. C. (2011). "Shimoliy Amerika turlarini taksonomik qayta ko'rib chiqish Leprariya s.l. that produce divaricatic acid, with notes on the type species of the genus L. incana". Mikologiya. 103 (6): 1216–1229. doi:10.3852/11-032. PMID 21642343. S2CID 34346229.
- ^ a b v d Casano, L. M.; Del Campo, E. M.; García-Breijo, F. J.; Reig-Armiñana, J; Gasulla, F; Del Hoyo, A; Guéra, A; Barreno, E (2011). "Two Trebouxia liken talliyasida har xil fiziologik ko'rsatkichlarga ega suv o'tlari mavjud Ramalina farinacea. Coexistence versus competition?". Atrof-muhit mikrobiologiyasi (Qo'lyozma taqdim etildi). 13 (3): 806–818. doi:10.1111/j.1462-2920.2010.02386.x. hdl:10251/60269. PMID 21134099.
- ^ a b Honegger, R. (1991) Fungal evolution: symbiosis and morphogenesis, Symbiosis as a Source of Evolutionary Innovation, Margulis, L., and Fester, R. (eds). Cambridge, MA, USA: The MIT Press, pp. 319–340.
- ^ a b Grube, M; Kardinale, M; De Kastro Jr, J. V.; Myuller, H; Berg, G (2009). "Liken simbiozlaridagi bakteriyalar jamoalarining turlarga xos tarkibiy va funktsional xilma-xilligi". ISME jurnali. 3 (9): 1105–1115. doi:10.1038 / ismej.2009.63. PMID 19554038.
- ^ a b Barreno, E., Errera-Kampos, M., Garsiya-Brejo, F., Gasulla, F. va Reig-Arminana, J. (2008) "Ramalinaandagi kortikal tuzilmalar bilan bog'liq bo'lgan fotosintetik bo'lmagan bakteriyalar Usnea Meksikadan kelgan talli "[doimiy o'lik havola ]. Asilomar, Pacific Grove, Kaliforniya, AQSh: tezislar IAL 6- ABLS qo'shma yig'ilishi.
- ^ a b Morris J, Purvis V (2007). Likenler (hayot). London: Tabiat tarixi muzeyi. p. 19. ISBN 978-0-565-09153-8.
- ^ "Liken". tomoshabin.co.uk. 2012 yil 17-noyabr. Arxivlandi asl nusxasidan 2014 yil 23 dekabrda. Olingan 2 noyabr 2014.
- ^ "Liken". Oksford yashash lug'ati. Oksford universiteti matbuoti. Arxivlandi asl nusxasidan 2014 yil 29 avgustda. Olingan 10 yanvar 2018.
- ^ Oksford ingliz lug'atida faqat "liken" talaffuzi keltirilgan: "liken". Oksford ingliz lug'ati (Onlayn tahrir). Oksford universiteti matbuoti. Olingan 10 yanvar 2018. (Obuna yoki ishtirok etuvchi muassasaga a'zolik talab qilinadi.)
- ^ Xarper, Duglas. "liken". Onlayn etimologiya lug'ati.
- ^ liken. Charlton T. Lyuis va Charlz Short. Lotin lug'ati kuni Perseus loyihasi.
- ^ χήνiχήν. Liddel, Genri Jorj; Skott, Robert; Yunoncha-inglizcha leksikon da Perseus loyihasi.
- ^ νiν yilda Liddel va Skott.
- ^ Asalarilar, Robert S. P. (2010). "s.v. Tiλε, λείχω". Yunon tilining etimologik lug'ati. Leyden hind-evropa etimologik lug'ati seriyasi. 1. Lucien van Beek yordamida. Leyden, Boston: Brill. 846-47 betlar. ISBN 9789004174184.
- ^ a b v d e f "Likenler va Bryofitlar, Michigan shtati universiteti, 10-25-99". Arxivlandi asl nusxasi 2011 yil 5 oktyabrda. Olingan 10 oktyabr 2014.
- ^ a b v d e f g h men j Liken so'zlashuvi, Shimoliy Amerika ma'lumotlari, Silviya va Stiven Sharnoff, [1] Arxivlandi 2015 yil 20 yanvar Orqaga qaytish mashinasi
- ^ a b v d e f g h men j k "Alan Silversaydning liken lug'ati (p-z), Alan Silversayd". Arxivlandi asl nusxasidan 2014 yil 31 oktyabrda. Olingan 10 oktyabr 2014.
- ^ a b v d e f g h men Dobson, F.S. (2011). Likenler, ingliz va irland turlarining tasvirlangan qo'llanmasi. Slough, Buyuk Britaniya: Richmond Publishing Co. ISBN 9780855463151.
- ^ a b v d e f g "Foliose likenlari, Liken Talus turlari, Allan Silversayd". Arxivlandi asl nusxasidan 2014 yil 19 oktyabrda. Olingan 10 oktyabr 2014.
- ^ a b v d e f g h men j k l m Shimoliy Amerikaning shimoliy-g'arbiy qismida moxlar likenlari va fernslari, Dale H. Vitt, Janet E. Marsh, Robin B. Bovey, Lone Pine nashriyot kompaniyasi, ISBN 0-295-96666-1
- ^ a b v d "Likenler, Saguaro-Juniper Corporation". Arxivlandi asl nusxasi 2015 yil 10 mayda. Olingan 10 oktyabr 2014.
- ^ a b v Makkun, B .; Grenon, J .; Martin, E .; Mutch, L.S .; Martin, E.P. (2007 yil mart). "Syerra Nevada milliy bog'laridagi boshqaruv masalalariga oid likenlar". Shimoliy Amerika qo'ziqorinlari. 2: 1–39. doi:10.2509 / pnwf.2007.002.003.
- ^ a b v d Michigan Lichens, Julie Jones Medlin, B. Jain Publishers, 1996, ISBN 0877370397, 9780877370390, [2] Arxivlandi 2016 yil 24-noyabr kuni Orqaga qaytish mashinasi
- ^ a b v d e f g h men j Likenler: Morfologiya haqida ko'proq ma'lumot, Kaliforniya universiteti Paleontologiya muzeyi, [3] Arxivlandi 2015 yil 28 fevral Orqaga qaytish mashinasi
- ^ a b v d e f g h men j k l Lichen Photobionts, Nebraska universiteti Omaha Arxivlandi 2014 yil 6 oktyabr kuni Orqaga qaytish mashinasi
- ^ a b "Alan Silversaydning liken lug'ati (g-o), Alan Silversayd". Arxivlandi asl nusxasidan 2014 yil 2 noyabrda. Olingan 10 oktyabr 2014.
- ^ a b v Büdel, B .; Scheidegger, C. (1996). Talus morfologiyasi va anatomiyasi. Liken biologiyasi. 37-64 betlar. doi:10.1017 / CBO9780511790478.005. ISBN 9780511790478.
- ^ a b Heidmarsson, Starri; Heidmarsson, Starri (1996). "Pruina liken turkumidagi taksonomik belgi sifatida Dermatokarpon". Bryolog. 99 (3): 315–320. doi:10.2307/3244302. JSTOR 3244302.
- ^ Sharnoff, Silviya va Sharnoff, Stiven. "Liken biologiyasi va atrof-muhit" Arxivlandi 2015 yil 17 oktyabrda Orqaga qaytish mashinasi. sharnoffphotos.com
- ^ Pogoda, S.S. Qo'riqchilar, K. G.; Lendemer, J. C .; Keyn, N. C .; Tripp, E. A. (2018). "Liken hosil qiluvchi zamburug'larda mitoxondriyal genomlarning murakkabligini pasayishi majburiy simbiozlarning genom me'morchiligiga oydinlik kiritdi - Vili Onlayn kutubxonasi". Molekulyar ekologiya. 27 (5): 1155–1169. doi:10.1111 / mec.14519. PMID 29417658.
- ^ Askomitset makrolixenlarning korteksidagi basidiomitset xamirturushlari - Ilm-fan
- ^ Skaloud, P; Peksa, O (2010). "ITS rDNA va aktinlar ketma-ketligiga asoslangan evolyutsion xulosalar oddiy liken algining xilma-xilligini ochib beradi. Asteroxloris (Trebouxiophyceae, Chlorophyta) ". Molekulyar filogenetik va evolyutsiyasi. 54 (1): 36–46. doi:10.1016 / j.ympev.2009.09.035. PMID 19853051.
- ^ Spribil, Tobi; Tuovinen, Veera; Resl, Filipp; Vanderpool, Dan; Volinski, Xeymo; Aime, M. Ketrin; Shnayder, Kevin; Stabentheiner, Edit; Tome-Xeller, Merje; Thor, Go'ran; Mayrhofer, Helmut (2016 yil 29-iyul). "Askomitset makrolixenlari korteksidagi basidiomitset xamirturushlari". Ilm-fan. 353 (6298): 488–492. Bibcode:2016Sci ... 353..488S. doi:10.1126 / science.aaf8287. ISSN 0036-8075. PMID 27445309.
- ^ a b v d e Ramel, Gordon. "Liken nima?". Earthlife Internet. Arxivlandi asl nusxasidan 2015 yil 19 yanvarda. Olingan 20 yanvar 2015.
- ^ a b Ahmadjon V. (1993). Liken simbiyozi. Nyu-York: John Wiley & Sons. ISBN 978-0-471-57885-7.
- ^ a b Honegger, R. (1988). "Mikobionts". Nash IIIda T.H. (tahrir). Liken biologiyasi. Kembrij: Kembrij universiteti matbuoti (1996 yilda nashr etilgan). ISBN 978-0-521-45368-4.
- ^ Ferry, B. V., Baddeley, M. S. va Xoksuort, D. L. (tahrirlovchilar) (1973) Havoning ifloslanishi va Likenler. Athlone Press, London.
- ^ Rose C. I., Hawksworth D. L. (1981). "Londonning toza havosida likenni rekolonizatsiya qilish". Tabiat. 289 (5795): 289–292. Bibcode:1981 yil 28-iyun ... 289R. doi:10.1038 / 289289a0. S2CID 4320709.
- ^ Xoksuort, D.L. va Rose, F. (1976) Lichenlar ifloslanish monitorlari sifatida. Edvard Arnold, Biologiya instituti seriyasi, 66-son. ISBN 0713125551
- ^ "Oak Moss Absolute Oil, Evernia prunastri, Parfyumeriya fiksatori". Arxivlandi asl nusxasidan 2014 yil 25 dekabrda. Olingan 19 sentyabr 2014.
- ^ Skogland, Terje (1984). "Yovvoyi kiyiklarni boqish uchun mo'ljallangan joy". Ekografiya. 7 (4): 345. doi:10.1111 / j.1600-0587.1984.tb01138.x.
- ^ Louri, Jeyms D .; Diederich, Pol (2003). "Likenikoloz zamburug'lar: o'zaro ta'sir, evolyutsiya va bioxilma-xillik" (PDF). Bryolog. 106: 80. doi:10.1639 / 0007-2745 (2003) 106 [0080: LFIEAB] 2.0.CO; 2. Arxivlandi (PDF) 2011 yil 3 yanvarda asl nusxadan. Olingan 2 may 2011.
- ^ Xagivara K, Rayt PR va boshqalar. (Mart 2015). "Islandiya va Gavayi likenlarining antioksidant xususiyatlarini qiyosiy tahlil qilish". Atrof-muhit mikrobiologiyasi. 18 (8): 2319–2325. doi:10.1111/1462-2920.12850. PMID 25808912. S2CID 13768322.
- ^ Odabasoglu F, Aslan A, Cakir A va boshq. (2005 yil mart). "Antioksidant faollik, ba'zi liken turlarining kuchini va umumiy fenolik tarkibini kamaytiradi". Fitoterapiya. 76 (2): 216–219. doi:10.1016 / j.fitote.2004.05.012. PMID 15752633.
- ^ a b Xak, Markus; Yurgens, Sascha-Rene; Lyushner, Kristof (2010). "Norstikt kislota: uning fizik-kimyoviy xususiyatlari va ushbu depsidon ishlab chiqaradigan likenlarning ekologik afzalliklari o'rtasidagi o'zaro bog'liqlik". Atrof-muhit va eksperimental botanika. 68 (3): 309. doi:10.1016 / j.envexpbot.2010.01.003.
- ^ a b "Yer hayoti tarmog'i, Likenlarda o'sish va rivojlanish". earthlife.net. Arxivlandi asl nusxasi 2015 yil 28 mayda. Olingan 12 oktyabr 2014.
- ^ "Likenler". Milliy park xizmati, AQSh Ichki ishlar vazirligi, Qo'shma Shtatlar hukumati. 2016 yil 22-may. Arxivlandi asl nusxasidan 2018 yil 5 aprelda. Olingan 4 aprel 2018.
- ^ Nash III, Tomas H. (2008). "Kirish". Nash IIIda T.H. (tahrir). Liken biologiyasi (2-nashr). Kembrij: Kembrij universiteti matbuoti. 1-8 betlar. doi:10.1017 / CBO9780511790478.002. ISBN 978-0-521-69216-8.
- ^ Bolduin, Emili (2012 yil 26 aprel). "Lichen Marsning qattiq muhitidan omon qoldi". Skymania yangiliklari. Arxivlandi asl nusxasi 2012 yil 28 mayda. Olingan 27 aprel 2012.
- ^ Sheldrake, Merlin (2020). Chigal hayot: Qo'ziqorinlar bizning dunyomizni qanday yaratadi, ongimizni o'zgartiradi va kelajagimizni qanday shakllantiradi. Bodli Xed. p. 94. ISBN 978-1847925206.
- ^ "ESA - insonning kosmik parvozi va tadqiqoti - Lichen kosmosda omon qoladi". Arxivlandi asl nusxasidan 2010 yil 26 fevralda. Olingan 16 fevral 2010.
- ^ Sancho, L. G.; De La Torre, R .; Xornek, G.; Ascaso, C .; De Los Rios, A .; Pintado, A .; Vierxos, J .; Schuster, M. (2007). "Lishayniklar kosmosda omon qoladi: 2005 yilgi LICHENS tajribasi natijalari". Astrobiologiya. 7 (3): 443–454. Bibcode:2007 AsBio ... 7..443S. doi:10.1089 / ast.2006.0046. PMID 17630840. S2CID 4121180.
- ^ Eyxorn, Syuzan E., Evert, Rey F. va Raven, Piter H. (2005). O'simliklar biologiyasi. Nyu-York: W. H. Freeman and Company. p. 1. ISBN 0716710072.
- ^ Kuk, Rebekka; Makfarland, Kennet (1995). Umumiy botanika 111 laboratoriya qo'llanmasi. Noksvill, TN: Tennessi universiteti. p. 104.
- ^ a b A. N. Ray; B. Bergman; Ulla Rasmussen (2002 yil 31-iyul). Simbiozdagi siyanobakteriyalar. Springer. p. 59. ISBN 978-1-4020-0777-4. Arxivlandi asl nusxasidan 2013 yil 31 dekabrda. Olingan 2 iyun 2013.
- ^ Qonun, R .; Lyuis, D. H. (1983 yil noyabr). "Biotik muhit va jinsiy aloqani saqlash - mutalualistik simbiozlardan ba'zi dalillar". Linnean Jamiyatining Biologik jurnali. 20 (3): 249–276. doi:10.1111 / j.1095-8312.1983.tb01876.x. ISSN 0024-4066.
- ^ Shkaloud, Pavel; Steinova, Yana; Kídká, Tereza; Vancurová, Lucie; Peksa, Ondeyj (2015 yil 4-may). "Suv o'tlari bioxilma-xilligi jumboqini yig'ish: Asterochloris (Trebouxiophyceae, Chlorophyta) turiga oid turlarni delimitatsiya qilish".. Fitologiya jurnali. 51 (3): 507–527. doi:10.1111 / jpy.12295. ISSN 0022-3646. PMID 26986666. S2CID 25190572.
- ^ Ramel, Gordon. "Likenlarning reproduktiv tuzilmalari". Arxivlandi asl nusxasi 2014 yil 28 fevralda. Olingan 22 avgust 2014.
- ^ a b v Murtagh GJ, Dyer PS, Krittenden PD (2000 yil aprel). "Jinsiy aloqa va bitta liken". Tabiat. 404 (6778): 564. Bibcode:2000 yil natur.404..564M. doi:10.1038/35007142. PMID 10766229. S2CID 4425228.
- ^ a b v Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008). Qo'ziqorinlarning lug'ati (10-nashr). Uollingford: CABI. 378-381 betlar. ISBN 978-0-85199-826-8.
- ^ a b v "Shakli va tuzilishi - Stikta va Dendriscocaulon". Avstraliya milliy botanika bog'lari. Arxivlandi asl nusxasi 2014 yil 28 aprelda. Olingan 18 sentyabr 2014.
- ^ Lutzoni, F.; Kauff, F.; Koks, C. J .; Maklafflin, D.; Selio, G.; Dentinger, B .; Padamzi, M .; Xibbett D.; va boshq. (2004). "Qo'ziqorin hayot daraxtini yig'ish: hujayralararo xususiyatlarning rivojlanishi, tasnifi va rivojlanishi". Amerika botanika jurnali. 91 (10): 1446–1480. doi:10.3732 / ajb.91.10.1446. PMID 21652303. S2CID 9432006.
- ^ Sanders, V.B.; Moe, R. L .; Ascaso, C. (2004). "Pirenomitset zamburug'i hosil qilgan intertidal dengiz likenlari Verrucaria tavaresiae (Askomikotina) va jigarrang suv o'tlari Petroderma maculiforme (Phaeophyceae): tall tashkiloti va simbiontning o'zaro ta'siri - NCBI ". Amerika botanika jurnali. 91 (4): 511–22. doi:10.3732 / ajb.91.4.511. PMID 21653406.
- ^ "Qo'ziqorinlar va suv o'tlari o'rtasidagi o'zaro munosabatlar - Nyu-Brunsvik muzeyi". Arxivlandi asl nusxasidan 2018 yil 18 sentyabrda. Olingan 4 oktyabr 2018.
- ^ Miller, Keti Ann; Peres-Ortega, Serxio. "Liken kontseptsiyasiga qarshi kurash: Turgidosculum ulvae - Kembrij". Likenolog. 50 (3): 341–356. doi:10.1017 / S0024282918000117. Arxivlandi asl nusxasidan 2018 yil 7 oktyabrda. Olingan 7 oktyabr 2018.
- ^ Rybalka, N .; Bo'ri, M.; Andersen, R. A .; Fridl, T. (2013). "Xloroplastning kelishuvi - BMC Evolyutsion Biologiya - BioMed Central". BMC evolyutsion biologiyasi. 13: 39. doi:10.1186/1471-2148-13-39. PMC 3598724. PMID 23402662.
- ^ Lutzoni, Fransua; Pagel, Mark; Reeb, Valeri (2001 yil 21-iyun). "Qo'ziqorinlarning asosiy nasllari liken simbiyotik ajdodlaridan olingan". Tabiat. 411 (6840): 937–940. Bibcode:2001 yil Natur.411..937L. doi:10.1038/35082053. PMID 11418855. S2CID 4414913.
- ^ Xoksuort, D.L. (1988). "Zamburug'li-algalli simbiozlarning xilma-xilligi, ularning evolyutsion ahamiyati va likenlarning tabiati". Linnean Jamiyatining Botanika jurnali. 96: 3–20. doi:10.1111 / j.1095-8339.1988.tb00623.x. S2CID 49717944.
- ^ a b v Rikkinen J. (1995). "Chiroyli ranglarning orqasida nima bor? Likenlarning fotobiologiyasini o'rganish". Bryobrothera. 4 (3): 375–376. doi:10.2307/3244316. JSTOR 3244316.
- ^ Fridl, T .; Büdel, B. (1996). "Fotobionts". Nash IIIda T.H. (tahrir). Liken biologiyasi. Kembrij: Kembrij universiteti matbuoti. 9-26 betlar. doi:10.1017 / CBO9780511790478.003. ISBN 978-0-521-45368-4.
- ^ a b "Alan Silversaydning liken lug'ati (a-f), Alan Silversayd". Arxivlandi asl nusxasidan 2014 yil 31 oktyabrda. Olingan 10 oktyabr 2014.
- ^ Hallenbek, Patrik C. (2017 yil 18-aprel). Fototrofik prokaryotlarda zamonaviy mavzular: ekologik va amaliy jihatlar. ISBN 9783319462615. Arxivlandi asl nusxasidan 2018 yil 4 oktyabrda. Olingan 4 oktyabr 2018.
- ^ a b Rikkinen, J. (2002). "Liken gildiyalari tegishli siyanobakterial simbiontlar bilan bo'lishadilar". Ilm-fan. 297 (5580): 357. doi:10.1126 / science.1072961. PMID 12130774. S2CID 35731669.
- ^ O'Brayen, H.; Miadlikowska, J .; Lutzoni, F. (2005). "Liken zamburug'ining bir-biriga yaqin to'rt turi bilan bog'liq simbiyotik siyanobakteriyalar bo'yicha mezbonlarning ixtisoslashuvini baholash Peltigera". Evropa pikologiya jurnali. 40 (4): 363–378. doi:10.1080/09670260500342647. S2CID 4094256.
- ^ Guzov-Krzeminska, B (2006). "Protoparmeliopsis muralis tlichenida fotobionning mavjudligi, uning ITS rDNA tahlillari bilan aniqlangan". Likenolog. 38 (5): 469–476. doi:10.1017 / s0024282906005068.
- ^ Ohmura, Y .; Kavachi, M .; Kasay, F .; Vatanabe, M. (2006). "Vegetativ ravishda ko'payadigan liken tarkibidagi simbiontlarning genetik birikmalari, Parmotrema tinctorum, ITS rDNA ketma-ketliklari asosida "(2006)". Bryolog. 109: 43–59. doi:10.1639 / 0007-2745 (2006) 109 [0043: gcosia] 2.0.co; 2.
- ^ Piercey-Normore (2006). "Likenni hosil qiluvchi asko-miket Evernia mezomorfasi ning multiplegenotiplari bilan bog'laydi Trebouxia jamesii". Yangi fitolog. 169 (2): 331–344. doi:10.1111 / j.1469-8137.2005.01576.x. PMID 16411936.
- ^ Lutzoni, Fransua; Pagel, Mark; Reeb, Valeri (2001). "Qo'ziqorinlarning asosiy nasllari liken simbiyotik ajdodlaridan olingan". Tabiat. 411 (6840): 937–940. Bibcode:2001 yil Natur.411..937L. doi:10.1038/35082053. PMID 11418855. S2CID 4414913.
- ^ a b "Lishayniklar: fotoalbom yozuvlari" Arxivlandi 2010 yil 25 yanvar Orqaga qaytish mashinasi, Kaliforniya universiteti Paleontologiya muzeyi.
- ^ Speer BR, Vagoner B. "Likenlarning fotoalbom yozuvlari". Kaliforniya universiteti Paleontologiya muzeyi. Arxivlandi asl nusxasidan 2010 yil 25 yanvarda. Olingan 16 fevral 2010.
- ^ Poinar Jr., GO. (1992). Amberdagi hayot. Stenford universiteti matbuoti.
- ^ Peterson EB. (2000). "E'tibordan chetda qolgan qoldiq liken (Lobariaceae)". Likenolog. 32 (3): 298–300. doi:10.1006 / lich.1999.0257.
- ^ Teylor, T. N .; Xass, X.; Remi, V.; Kerp, H. (1995). "Eng qadimgi fotoalbom liken". Tabiat. 378 (6554): 244. Bibcode:1995 yil 378..244T. doi:10.1038 / 378244a0. S2CID 4353572. Arxivlandi asl nusxasi 2007 yil 11 yanvarda.
- ^ Teylor WA, Free CB, Helgemo R, Ochoada J (2004). "Spongiofitonning SEM tahlili fotoalbom liken sifatida talqin qilingan". Xalqaro o'simlik fanlari jurnali. 165 (5): 875–881. doi:10.1086/422129. S2CID 85382155.
- ^ Jahren, A.H .; Porter, S .; Kuglich, J.J. (2003). "Devonning dastlabki er usti organizmlarida liken metabolizmi aniqlangan". Geologiya. 31 (2): 99–102. Bibcode:2003 yil Geo .... 31 ... 99J. doi:10.1130 / 0091-7613 (2003) 031 <0099: LMIIED> 2.0.CO; 2. ISSN 0091-7613. S2CID 49241170.
- ^ Fletcher, B. J .; Berling, D. J.; Chaloner, W. G. (2004). "Barqaror uglerod izotoplari va quruqlikdagi Devon organizmining metabolizmi Spongiofiton". Geobiologiya. 2 (2): 107–119. doi:10.1111 / j.1472-4677.2004.00026.x.
- ^ Edvards D; Axe L (2012). "Prototaksitlarning yaqin ittifoqchisi bo'lgan Nematasketumga qo'ziqorin yaqinligi to'g'risida dalillar". Linnean Jamiyatining Botanika jurnali. 168: 1–18. doi:10.1111 / j.1095-8339.2011.01195.x.
- ^ Retallack G.J .; Landing, E. (2014). "Devon magistrallarining yaqinligi va arxitekturasi Prototaksitlar loganii". Mikologiya. 106 (6): 1143–1156. doi:10.3852/13-390. PMID 24990121. S2CID 20013625.
- ^ Karatigin IV; Snigirevskaya NS; Vikulin SV. (2009). "Eng qadimiy quruqlikdagi liken Winfrenatia reticulata : Yangi topilma va yangi talqin ". Paleontologik jurnal. 43 (1): 107–114. doi:10.1134 / S0031030109010110. S2CID 85262818.
- ^ Karatigin IV, Snigirevskaya NS, Vikulin SV (2007). "Dastlabki Devon ekotizimidagi qo'ziqorinlar ishtirokidagi simbiyozning ikki turi". Evropa Mikologlarining XV Kongressi, Sankt-Peterburg, Rossiya, 2007 yil 16-21 sentyabr. 1 (1): 226. Arxivlandi asl nusxasidan 2013 yil 24 aprelda. Olingan 18 fevral 2011.
- ^ "Likenlar olimlarning fikriga qaraganda yoshroqdir - o'simliklardan keyin million yillar o'tgach rivojlangan bo'lishi mumkin". 15 Noyabr 2019. Arxivlandi asl nusxasidan 2019 yil 18 dekabrda. Olingan 18 noyabr 2019.
- ^ Xansen, Karen; Perri, Brayan A.; Dranginis, Endryu V.; Pfister, Donald H. (2013 yil may). "Pironemataceae (Pezizomycetes, Ascomycota) juda xilma-xil stakan-qo'ziqorinlar oilasi filogeniyasi tanlangan hayot tarixi xususiyatlarining aloqalari va evolyutsiyasini aniqlaydi". Molekulyar filogenetik va evolyutsiyasi. 67 (2): 311–335. doi:10.1016 / j.ympev.2013.01.014. ISSN 1055-7903. PMID 23403226.
- ^ Schoch CL; Shon GH; Lopes-Giraldez F; Townsend JP; Miadlikowska J; Hofstetter V; Robbertse B; Matheny PB; va boshq. (2009). "Ascomycota hayot daraxti: keng miqyosdagi filogeniya asosiy reproduktiv va ekologik belgilarning kelib chiqishi va evolyutsiyasini aniqlaydi". Syst. Biol. 58 (2): 224–239. doi:10.1093 / sysbio / syp020. PMID 20525580.
- ^ Gargas, A; Depriest, PT; Grube, M; Tehler, A (1995). "SSU rDNA filogeniyasi tomonidan taklif qilingan qo'ziqorinlarda liken simbiyozlarining ko'p kelib chiqishi". Ilm-fan. 268 (5216): 1492–1495. Bibcode:1995 yil ... 268.1492G. doi:10.1126 / science.7770775. PMID 7770775. S2CID 563601.
- ^ Honegger R. (1998). "Liken simbiozi - bu nimani ajoyib qiladi?" (PDF). Likenolog. 30 (3): 193–212. doi:10.1017 / s002428299200015x. Arxivlandi (PDF) asl nusxasidan 2019 yil 26 aprelda. Olingan 30 yanvar 2019.
- ^ Wedin M, Döring H, Gilenstam G (2004). "Saprotrofiya va likenlashtirish bir xil qo'ziqorin turlarining turli substratlar uchun variantlari sifatida: ekologik plastika va qo'ziqorinlarning turmush tarzi Striktis-Konotrema murakkab ". Yangi fitolog. 16 (3): 459–465. doi:10.1111 / j.1469-8137.2004.01198.x.
- ^ Yuan X, Xiao S, Teylor TN (2005). "600 million yil oldin likenga o'xshash simbioz". Ilm-fan. 308 (5724): 1017–1020. Bibcode:2005 yil ... 308.1017Y. doi:10.1126 / science.1111347. PMID 15890881. S2CID 27083645.
- ^ Retallack G.J. (2015). "Qo'ziqorinlarning prekambriyalik adaptiv nurlanishining akritarx dalili" (PDF). Botanika Pacifica. 4 (2): 19–33. doi:10.17581 / bp.2015.04203 yil. Arxivlandi (PDF) asl nusxasidan 2016 yil 22 dekabrda. Olingan 22 dekabr 2016.
- ^ Retallack GJ. (2007). "Ediakaranning mashhur fotoalbomlari bo'lgan Dikkinsoniyaning o'sishi, parchalanishi va ko'milishi". Alcheringa: Avstraliyaning Paleontologiya jurnali. 31 (3): 215–240. doi:10.1080/03115510701484705. S2CID 17181699.
- ^ Retallack GJ. (1994). "Ediakaran qoldiqlari likenlari bo'lganmi?". Paleobiologiya. 20 (4): 523–544. doi:10.1017 / s0094837300012975. JSTOR 2401233.
- ^ Switek B (2012). "Qarama-qarshi da'vo quruqlikda hayotni 65 million yilga erta qo'yadi". Tabiat. doi:10.1038 / tabiat.2012.12017. S2CID 130305901. Arxivlandi 2013 yil 1 yanvardagi asl nusxadan. Olingan 2 yanvar 2013.
- ^ Retallack, G.J .; Dann, K.L .; Saxby, J. (2015). "Glazier National Park (AQSh, Montana, AQSh) dan Mesoproterozoyik qoldiqlari Horodyskia". Prekambriyen tadqiqotlari. 226: 125–142. Bibcode:2013PreR..226..125R. doi:10.1016 / j.precamres.2012.12.005.
- ^ a b Retallack, G.J .; Krull, E.S .; Takray, G.D .; Parkinson, D. (2013). "Janubiy Afrikadagi paleoproterozoyik (2,2 Ga) paleosoldan urn shaklidagi muammoli qoldiqlar". Prekambriyen tadqiqotlari. 235: 71–87. Bibcode:2013 yil avval..235 ... 71R. doi:10.1016 / j.precamres.2013.05.015.
- ^ Lyuking, Robert; Nelsen, Metyu P. (2018), "Ediacarans, Protolichenlar va Likendan olingan penitsillium", Transformativ paleobotanika, Elsevier, 551-590 betlar, doi:10.1016 / b978-0-12-813012-4.00023-1, ISBN 978-0-12-813012-4, olingan 14 noyabr 2020
- ^ Quruqlikda yashash poygasida likenler o'simliklarni mag'lub etmadi - The New York Times
- ^ "Ifloslanish, o'simlik osti dunyosi". Avstraliya milliy botanika bog'lari. Arxivlandi asl nusxasi 2014 yil 17 fevralda. Olingan 10 oktyabr 2014.
- ^ Chen, Jie; Blyum, Xans-Piter; Beyer, Lotar (2000). "Liken kolonizatsiyasi natijasida hosil bo'lgan toshlarning ob-havosi - sharh" (PDF). KATENA. 39 (2): 121. doi:10.1016 / S0341-8162 (99) 00085-5. Arxivlandi (PDF) asl nusxasidan 2015 yil 2 aprelda. Olingan 21 mart 2015.
- ^ Jons, Klive G.; Shachak, Moshe (1990). "Tosh yeyayotgan salyangozlar cho'l tuprog'ini urug'lantirish". Tabiat. 346 (6287): 839. Bibcode:1990 yil 34-iyun. doi:10.1038 / 346839a0. S2CID 4311333.
- ^ Walker, T. R. (2007). "Labrador, Kanalning boreal o'rmonlarining likenlari: Tekshirish ro'yxati". Evansiya. 24 (3): 85–90. doi:10.1639/0747-9859-24.3.85. S2CID 129100097.
- ^ Oksanen, I. (2006). "Likenlarning ekologik va biotexnologik jihatlari". Amaliy mikrobiologiya va biotexnologiya. 73 (4): 723–734. doi:10.1007 / s00253-006-0611-3. PMID 17082931. S2CID 12172616.
- ^ Lawrey, Jeyms D. (1994). "Liken allelopati: sharh". Inderjitda; K. M. M. Dakshini; Frank A. Eynhellig (tahr.). Allelopatiya. Organizmlar, jarayonlar va dasturlar. ACS simpoziumi seriyasi. 582. Amerika kimyo jamiyati. 26-38 betlar. doi:10.1021 / bk-1995-0582.ch002. ISBN 978-0-8412-3061-3.
- ^ a b v Nash III, Tomas H. (2008). "Havoning ifloslanishiga liken sezgirligi". Nash IIIda T.H. (tahrir). Liken biologiyasi (2-nashr). Kembrij: Kembrij universiteti matbuoti. 299-314 betlar. doi:10.1017 / CBO9780511790478.016. ISBN 978-0-521-69216-8.
- ^ Knops, J.M.H .; Nash, T. H. III; Boucher, V.L .; Schlesinger, W.H. (1991). "Mineral velosiped va epifitik lishayniklar: ekotizim darajasidagi ta'siri". Likenolog. 23 (3): 309–321. doi:10.1017 / S0024282991000452.
- ^ Halonen P, Hyvarinen M, Kauppi M (1993). "Epifitik liken tarkibidagi emissiya bilan bog'liq va elementlarning kontsentratsiyasini takroriy nazorat qilish Gipogimniya fizodlari Finlyandiyaning g'arbiy qismida ". Annales Botanici Fennici. 30: 251–261.
- ^ Walker T. R.; Pystina T. N. (2006). "Rossiyaning Pechora havzasida neft va gaz sanoati tomonidan yuzaga keladigan ifloslanish va ekologik ta'sirlarni kuzatish uchun likenlardan foydalaning". Gersogiya. 19: 229–238.
- ^ Walker T. R.; Krittenden P. D .; Yosh S. D .; Pristina T. (2006). "Rossiyaning shimoliy-sharqiy Pechora havzasida neft va gaz sanoati tufayli ifloslanish ta'sirini baholash". Ekologik ko'rsatkichlar. 6 (2): 369–387. doi:10.1016 / j.ecolind.2005.03.015.
- ^ Walker T. R.; Krittenden P. D .; Yosh S. D. (2003). "Rossiyaning shimoliy-sharqiy qismida, Usa daryosi havzasida kislota chiqindilarining manbalariga nisbatan qishki qor va terrikolous likenlarning kimyoviy tarkibidagi mintaqaviy o'zgarish". Atrof muhitning ifloslanishi. 125 (3): 401–412. doi:10.1016 / s0269-7491 (03) 00080-0. PMID 12826418.
- ^ Vays-Penzias, Piter S.; Bank, Maykl S.; Klifford, Deana L.; Torregrosa, Alisiya; Zheng, Belle; Lin, Vendi; Uilmers, Kristofer C. (26 noyabr 2019). "Dengiz tumaniga tushadigan kirishlar qirg'oqdagi quruqlikdagi oziq-ovqat tarmog'ida metilmerkuriy bioakkumulyatsiyasini kuchaytirmoqda". Ilmiy ma'ruzalar. 9 (1): 17611. Bibcode:2019 NatSR ... 917611W. doi:10.1038 / s41598-019-54056-7. PMC 6879473. PMID 31772229.
- ^ Hogan, C. Maykl (2010). "Abiotik omil". Yer entsiklopediyasi. Vashington, DC: Fan va atrof-muhit bo'yicha milliy kengash. Arxivlandi asl nusxasi 2013 yil 8-iyun kuni. Olingan 27 oktyabr 2013.
- ^ Beltman IH, de Kok LJ, Kuiper PJC, van Hasselt PR (1980). "Epifitik likenlarning yog 'kislotasi tarkibi va xlorofill tarkibi va ularning havoning ifloslanishiga sezgirligi bilan bog'liqligi". Oikos. 35 (3): 321–326. doi:10.2307/3544647. JSTOR 3544647.
- ^ Emmerich R, Giez I, Lange OL, Proksch P (1993). "Liken birikmalarining toksikligi va antifedant faolligi, polifagli o'txo'r hasharotlarga qarshi Spodoptera littoralis". Fitokimyo. 33 (6): 1389–1394. doi:10.1016 / 0031-9422 (93) 85097-B.
- ^ Beschel RE (1950). "Flecten als altersmasstab Rezenter morainen". Zeitschrift für Gletscherkunde und Glazialgeologie. 1: 152–161.
- ^ Curry, R. R. (1969) "Markaziy Syerra Nevada, Kaliforniya shtatining golosenli iqlim va muzlik tarixi", 1-47 betlar, Amerika Geologiya Jamiyati Maxsus Qog'oz, 123, S. A. Shumm va V. C. Bredli, eds.
- ^ Sowers, J. M., Noller, J. S. va Lettis, W. R. (tahr.) (1997) Tanishuv va zilzilalar: to'rtlamchi davr geoxronologiyasini ko'rib chiqish va uni paleosismologiyada qo'llash. AQSh yadroviy tartibga solish komissiyasi, NUREG / CR 5562.
- ^ Innes, J. L. (1985). "Likenometriya". Jismoniy geografiyada taraqqiyot. 9 (2): 187. doi:10.1177/030913338500900202. S2CID 220949784.
- ^ Cappitelli, Francesca; Sorlini, Klaudiya (2008). "Mikroorganizmlar bizning madaniy merosimizni ifodalaydigan narsalarda sintetik polimerlarga hujum qiladi". Amaliy va atrof-muhit mikrobiologiyasi. 74 (3): 564–569. doi:10.1128 / AEM.01768-07. PMC 2227722. PMID 18065627.
- ^ Gadd, Jefri Maykl (2010 yil mart). "Metall, minerallar va mikroblar: geomikrobiologiya va bioremediatsiya". Mikrobiologiya. 156 (Pt 3): 609-643. doi:10.1099 / mic.0.037143-0. PMID 20019082.
- ^ Jonson, Kristofer J; Bennet, Jeyms P; Biro, Stiven M; Dyuk-Velaskes, Xuan Kamilo; Rodrigez, Sintiya M.; Bessen, Richard A; Rok, Tonie E (2011 yil may). Bartz, Jeyson S (tahrir). "Kasallik bilan bog'liq bo'lgan prion oqsilining likenlardan serinli proteinlar bilan parchalanishi". PLOS ONE. 6 (5): e19836. Bibcode:2011PLoSO ... 619836J. doi:10.1371 / journal.pone.0019836. PMC 3092769. PMID 21589935.
- ^ Rodriges, Sintiya M; Bennet, Jeyms P; Jonson, Kristofer J (2012 yil 1-yanvar). "Likenler: antionionlarga qarshi kutilmagan agentlarmi?". Prion. 6 (1): 11–16. doi:10.4161 / pri.6.1.17414. PMC 3338958. PMID 22453171.
- ^ Bennet, Jeyms P; Rodrigez, Sintiya M; Jonson, Kristofer (2012 yil iyul). "Clioniya jinsi likenlari tomonidan Prion oqsilining parchalanishi". Likenolog. 44 (4): 523–531. doi:10.1017 / S0024282912000102.
- ^ Kasselman, Karen Ley; Dekan, Jenni (1999). Yovvoyi rang: [tabiiy bo'yoqlarni tayyorlash va ulardan foydalanish bo'yicha to'liq qo'llanma]. Nyu-York: Watson-Guptill nashrlari. ISBN 978-0-8230-5727-6.
- ^ Myuller, K (2001). "Likenlardan olingan farmatsevtik jihatdan tegishli metabolitlar". Amaliy mikrobiologiya va biotexnologiya. 56 (1–2): 9–10. doi:10.1007 / s002530100684. PMID 11499952. S2CID 11958438.
- ^ a b Morton, E .; Uinters, J. va Smit, L. (2010). "Turli xil davolash qilingan moxlar va likenlarning antiseptik va antibiotik xususiyatlarini tahlil qilish" Arxivlandi 2017 yil 20-avgust Orqaga qaytish mashinasi. Michigan universiteti biologik stantsiyasi
- ^ Bustinza, F. (1952). "Likenlardan antibakterial moddalar". Iqtisodiy botanika. 6 (4): 402–406. doi:10.1007 / bf02984888. S2CID 39335883.
- ^ "Themodelrailroader.com". Arxivlandi asl nusxasi 2014 yil 15 oktyabrda. Olingan 10 oktyabr 2014.
- ^ Rabbi shunday izohladi Xanox Zundel ben Jozef, uning sharhida Ets Yosef ("Jozef daraxti"), ustida Sefer Midrash Rabbah, vol. 2, Nyu-York, 1987 yil, s.v. Rut Rabba 6:3
- ^ Zohar Amar va Yaron Serri, Al-Tamimiy ta'riflaganidek, Isroil va Suriya mamlakati, Ramat-Gan 2004, 56-bet, 108-109 ISBN 965-226-252-8 (Ibroniycha)
- ^ a b v d Honegger R. (2000). "Simon Schwender (1829-1919) va likenlardagi ikkilamchi gipoteza". Bryolog. 103 (2): 307–313. doi:10.1639 / 0007-2745 (2000) 103 [0307: SSATDH] 2.0.CO; 2. ISSN 0007-2745. JSTOR 3244159.
- ^ Treub, Melchior (1873) Onderzoekingen de natuur der lichenen ustidan. Leyden universiteti dissertatsiyasi.
- ^ Yong, Ed (2016 yil 21-iyul), "Montanadagi treyler parkidagi bir yigit 150 yillik biologiyani qanday ag'darib tashlagan", Atlantika, arxivlandi asl nusxasidan 2017 yil 23 iyulda, olingan 23 iyul 2017.
Qo'shimcha o'qish
- Yorgensen, Per M. va Lyuking, Robert (2018 yil aprel). "" Rustici Pauperrimi ": Likenlar to'g'risida Linney afsonasi tuzatildi". Linnean 34(1), 9-12 betlar.
Tashqi havolalar
- LIAS nuri: Dunyoning liken turlari uchun onlayn interaktiv identifikatsiya kaliti
- Kaliforniya universiteti Paleontologiya muzeyi kustoza yoki skuamuloza likenining kesimining mikroskopik tasviri
- Earth Life Internet - turli xil o'sish shakllari uchun ichki liken tuzilmalarining sxematik rasmlari
- Sidney universiteti liken biologiyasi
- Memorial Universitetining NL Nature loyihasi, asosan, likenlarga qaratilgan
- Qishloq xo'jaligini kashf etgan qo'ziqorinlar
- Britaniya likenlari jamiyati