Epstein-Barr virusi - Epstein–Barr virus
| Inson gammaherpesvirusi 4 | |
|---|---|
 | |
| Elektron mikrograf dumaloq ko'rinadigan ikkita Epshteyn-Barr virionlari (virusli zarralar) kapsidlar erkin membrana bilan o'ralgan konvert | |
| Viruslarning tasnifi | |
| (ochilmagan): | Virus |
| Shohlik: | Duplodnaviriya |
| Qirollik: | Heunggongvirae |
| Filum: | Peploviricota |
| Sinf: | Gerviviritsetlar |
| Buyurtma: | Herpesvirales |
| Oila: | Herpesviridae |
| Tur: | Lenfokriptovirus |
| Turlar: | Inson gammaherpesvirusi 4 |
| Sinonimlar[1] | |
| |
The Epstein-Barr virusi (EBV), rasmiy ravishda chaqirilgan Inson gammaherpesvirusi 4, ma'lum bo'lgan to'qqiz kishidan biridir inson gerpesvirus turlari ichida gerpes oilasi, va bu eng keng tarqalganlardan biridir viruslar odamlarda.
Bu eng yaxshi sabab sifatida tanilgan yuqumli mononuklyoz ("mono" yoki "bezgak bezgagi"). Shuningdek, u turli xil zararli bo'lmagan, premalignant va malign Epstein-Barr virusi bilan bog'liq lenfoproliferativ kasalliklar kabi Burkitt limfomasi, gemofagotsitik limfohistiotsitoz,[2] va Xojkin limfomasi; kabi lenfoid bo'lmagan malign kasalliklar oshqozon saratoni va nazofarenks karsinomasi; va bilan bog'liq sharoitlar inson immunitet tanqisligi virusi kabi tukli leykoplakiya va markaziy asab tizimi limfomalar.[3][4] Virus shuningdek, bolalik kasalliklari bilan bog'liq Alice Wonderland sindromida[5] va o'tkir serebellar ataksiya[6] va ba'zi dalillarga asoslanib, ba'zi bir rivojlanish xavfi yuqori otoimmun kasalliklar,[7] ayniqsa dermatomiyozit, tizimli eritematoz, romatoid artrit, Syogren sindromi,[8][9] va skleroz.[10][11][12] Yiliga taxminan 200,000 saraton kasalligi EBVga tegishli deb o'ylashadi.[13][14]
EBV bilan yuqtirish og'iz orqali yuborish orqali sodir bo'ladi tupurik[15] va jinsiy a'zolar sekretsiyasi.
Aksariyat odamlar EBV bilan kasallanib, daromad olishadi adaptiv immunitet. Qo'shma Shtatlarda, besh yoshli bolalarning yarmiga yaqini va kattalarning 90% ga yaqini ilgari yuqtirilgan.[16] Chaqaloqlar EBVga sezgir bo'lib qoladilar onaning antikorlarini himoya qilish yo'qoladi. Ko'pgina bolalar EBV bilan kasallanishadi va bu infektsiyalar odatda hech qanday alomatlarni keltirib chiqarmaydi yoki bolalikning boshqa engil, qisqa kasalliklari bilan ajralib turmaydi. Qo'shma Shtatlarda va boshqa rivojlangan mamlakatlarda ko'p odamlar bolalik davrida EBV yuqtirmaganlar.[17] EBV bilan yuqtirish o'spirinlik davrida yuzaga kelsa, bu sabab bo'ladi yuqumli mononuklyoz 35 dan 50% gacha.[18]
EBV yuqadi B hujayralari immun tizimining va epiteliya hujayralari. EBV boshlangandan so'ng litik infektsiya nazorat ostiga olinadi, EBV kechikish shaxsning B hujayralarida umrining oxirigacha saqlanib qoladi.[15][19]
Belgilari va alomatlari
EBV bilan kasallangan bolalarda kam simptomlar mavjud yoki ular asemptomatik ko'rinishi mumkin, ammo EBV o'spirin yoki kattalar bilan kasallangan bo'lsa, bu sabab bo'lishi mumkin charchoq, isitma, tomoq yallig'langan, shishgan limfa tugunlari bo'ynida, kengaygan taloq, shishgan jigar yoki toshma.[20] Yuqumli kasallikdan keyin Surunkali charchoq sindromi shuningdek, Epstein-Barr infektsiyasi bilan bog'liq.[21]
Virusologiya
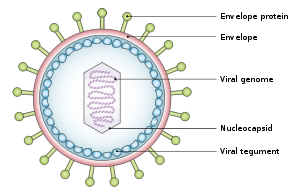
Tuzilishi va genomi
The virus taxminan 122-180 ga teng nm diametrida va tarkibiga kiradi er-xotin spiral ning deoksiribonuklein kislotasi (DNK) tarkibida 172000 ga yaqin tayanch juftliklari va 85 genlar.[15] DNK oqsil bilan o'ralgan nukleokapsid bilan o'ralgan tegument oqsildan iborat bo'lib, u o'z navbatida an bilan o'ralgan konvert ikkalasini ham o'z ichiga oladi lipidlar[22] va sirt proektsiyalari glikoproteinlar uchun zarur bo'lgan mezbon hujayraning infektsiyasi.[22] 2020 yil iyul oyida tadqiqotchilar guruhi virusning nukleokapsidining birinchi to'liq atom modeli haqida xabar berishdi. Ushbu "birinchi to'liq atom modeli [o'z ichiga oladi] ikosahedral kapsid, kapsid bilan bog'liq tegument kompleksi (CATC) va dodekamerik portal - virus genomining translokatsiya apparati."[23][24]
Tropizm
Atama virusli tropizm EBV yuqadigan qaysi hujayra turlarini anglatadi. EBV turli xil hujayra turlarini yuqtirishi mumkin, shu jumladan B hujayralari va epiteliya hujayralari.[25]
Virusli uch qismli glikoprotein komplekslari gHgL gp42 vositachilik B hujayralari membranasi sintezi; gHgL ning ikki qismli komplekslari vositachiligida epiteliya hujayralari membranasi sintezi. B hujayralarida hosil bo'lgan EBV kam miqdordagi gHgLgp42 komplekslariga ega, chunki bu uch qismli komplekslar o'zaro ta'sir qiladi Inson-leykotsit-antigeni II sinf endoplazmatik retikulumdagi B hujayralarida mavjud bo'lgan va parchalanadigan molekulalar. Aksincha, epiteliya hujayralaridan EBV uch qismli komplekslarga boy, chunki bu hujayralar odatda HLA II sinf molekulalarini o'z ichiga olmaydi. Natijada, B hujayralaridan hosil bo'lgan EBV epiteliya hujayralariga, epiteliya hujayralaridan olingan EBV esa B hujayralariga ko'proq yuqadi. Kam bo'lgan viruslar gp42 qismi inson B hujayralari bilan bog'lanish qobiliyatiga ega, ammo yuqtirishga qodir emas.[26]
Replikatsiya davri

Hujayraga kirish
EBV B hujayralariga ham, epiteliya hujayralariga ham zarar etkazishi mumkin. Ushbu ikkita hujayraga kirish mexanizmlari har xil.
Virusli B hujayralariga kirish uchun glikoprotein gp350 hujayra retseptorlari bilan bog'lanadi CD21 (CR2 nomi bilan ham tanilgan).[27] Keyin virus glikoprotein gp42 uyali bilan o'zaro ta'sir qiladi MHC II sinf molekulalar. Bu tetiklaydi birlashma hujayra membranasi bilan virusli konvertdan iborat bo'lib, EBV ning B hujayraga kirishiga imkon beradi.[22] Shuningdek, ma'lum bo'lgan inson CD35 komplement retseptorlari 1 (CR1), gp350 / 220 uchun qo'shimcha biriktiruvchi omil bo'lib, EBV ning CD21 manfiy hujayralariga, shu jumladan pishmagan B hujayralariga kirishi uchun marshrutni ta'minlay oladi. EBV infektsiyasi CD35 ekspressionini pasaytiradi.[28]
Epiteliya hujayralariga kirish uchun BMRF-2 virusli oqsili uyali D1 bilan o'zaro ta'sir qiladi integrallar. Keyin gH / gL virusli oqsillari hujayra bilan o'zaro ta'sir qiladi avβ6 /avβ8 integrallar. Bu tetiklaydi birlashma epiteliya hujayrasi membranasi bilan virusli konvertning, EBV ning epiteliya hujayrasiga kirishiga imkon beradi.[22] B-hujayraning kirib kelishidan farqli o'laroq, epiteliya hujayralariga kirishga virus glikoprotein gp42 to'sqinlik qiladi.[27]
EBV hujayraga kirgandan so'ng, virusli kapsid eriydi va virus genomi u tomonga ko'chiriladi hujayra yadrosi.
Litik replikatsiya
The litik tsikl yoki samarali infeksiya natijasida yuqumli kasallik hosil bo'ladi virionlar. EBV ham B hujayralarida, ham epiteliya hujayralarida litik replikatsiyaga uchrashi mumkin. B hujayralarida litik replikatsiya odatda faqat qayta faollashgandan so'ng amalga oshiriladi kechikish. Epiteliya hujayralarida litik replikatsiya ko'pincha to'g'ridan-to'g'ri keladi virusli kirish.[22]
Litik replikatsiya paydo bo'lishi uchun virusli genom chiziqli bo'lishi kerak. Yashirin EBV genomi daireseldir, shuning uchun u litik reaktivatsiya jarayonida chiziqli bo'lishi kerak. Litik replikatsiya paytida virusli DNK polimeraza virus genomini nusxalash uchun javobgardir. Bu xujayrali DNK polimeraza virus genomini nusxa ko'chiradigan kechikish bilan farq qiladi.[22]
Lytic gen mahsulotlari ketma-ket uchta bosqichda ishlab chiqariladi: darhol, erta va kech.[22]Zudlik bilan erta litik genlari mahsulotlari rol o'ynaydi transaktivatorlar, takomillashtirish ifoda keyingi litik genlarning Darhol erta litik gen mahsulotlariga kiradi BZLF1 (shuningdek, uning geni bilan bog'liq bo'lgan Zta, EB1 deb nomlanadi ZEBRA) va BRLF1 (uning mahsulot geni bilan bog'liq Rta).[22]Erta litik genlari mahsulotlarida replikatsiya, metabolizm va blokada kabi ko'plab funktsiyalar mavjud antigenni qayta ishlash. Erta litik gen mahsulotlariga kiradi BNLF2.[22]Va nihoyat, kech litik geni mahsulotlari, masalan, tarkibiy rollarga ega bo'lgan oqsillarga moyil VCA, bu virusni hosil qiladi kapsid. BCRF1 kabi boshqa kech litik gen mahsulotlari EBVga immunitet tizimidan qochishga yordam beradi.[22]
EGCG, ichida polifenol yashil choy, DNK, gen transkripsiyasi va oqsillar darajasida EBV spontan litik infektsiyani inhibe qilish ishida bir vaqtning o'zida va dozaga bog'liq usul; EBV litik genlarining ekspressioni Zta, Rta, va erta antijen kompleksi EA-D (tomonidan chaqirilgan Rta), ammo juda barqaror EBNA-1 EBV infektsiyasining barcha bosqichlarida topilgan gen ta'sirlanmaydi.[29] Maxsus inhibitorlar (yo'llarga) shuni ko'rsatmoqdalar Ras / MEK / MAPK yo'li BZLF1 va EBV litik infektsiyasiga hissa qo'shadi PI3-K BRLF1 orqali o'tish yo'li, ikkinchisi BRLF1 qobiliyatini butunlay bekor qiladi adenovirus vektori EBV infektsiyasining litik shaklini keltirib chiqarish.[29] Bundan tashqari, ba'zi bir genlarning faollashuvi, boshqalari esa yashirin yuqtirilgan B-hujayralarni ikkitasini qo'llash orqali immunitetni yo'q qilishni qanday boshlashni aniqlash uchun o'rganilmoqda. TPA yoki natriy butirat.[29]
Kechikish
Litik replikatsiyadan farqli o'laroq, kechikish virionlarni ishlab chiqarishga olib kelmaydi.[22]Buning o'rniga, EBV genomining doiraviy DNKsi hujayra yadrosi sifatida epizod va uyali telefon orqali ko'chiriladi DNK polimeraza.[22]Kechikish davrida EBV genlarining faqat bir qismi mavjud ifoda etilgan.[15][30]Yashirin EBV o'z genlarini kechikish dasturlari deb nomlanadigan uchta naqshning birida ifodalaydi. EBV yashirin ravishda davom etishi mumkin B hujayralari va epiteliya hujayralari, lekin ikki xil hujayrada turli xil kechikish dasturlari mumkin.
EBV uchta kechikish dasturidan birini namoyish qilishi mumkin: I kechikish, II kechikish yoki III kechikish. Har bir kechikish dasturi cheklangan, aniq virus to'plamini ishlab chiqarishga olib keladi oqsillar va virusli RNKlar.[31][32]
| Gen ifodalangan | EBNA-1 | EBNA-2 | EBNA-3 A | EBNA-3 B | EBNA-3 C | EBNA-LP | LMP-1 | LMP-2 A | LMP-2 B | EBER |
|---|---|---|---|---|---|---|---|---|---|---|
| Mahsulot | Oqsil | Oqsil | Oqsil | Oqsil | Oqsil | Oqsil | Oqsil | Oqsil | Oqsil | ncRNAlar |
| Kechikish I | + | – | – | – | – | – | – | – | – | + |
| Kechikish II | + | – | – | – | – | + | + | + | + | + |
| Kechikish III | + | + | + | + | + | + | + | + | + | + |
Bundan tashqari, barcha virusli oqsil ekspresiyasi o'chirilgan dastur joylashtirilgan (Kechikish 0).
B hujayralarida uchta kechikish dasturi ham mumkin.[15]B hujayralaridagi EBV kechikishi odatda III kechikishdan II kechikishgacha I kechikishgacha davom etadi. Kechikishning har bir bosqichi B hujayralarining xatti-harakatlariga ta'sir qiladi.[15]Dam olishni yuqtirishda sodda B hujayrasi, EBV kechikish III ga kiradi. III kechikish davrida hosil bo'lgan oqsillar va RNKlar to'plami B hujayrasini proliferatsiya qiluvchi portlashga aylantiradi (B hujayrasini faollashishi deb ham ataladi).[15][22]Keyinchalik, virus gen ekspressionini cheklaydi va II kechikish davriga kiradi. Kechikish II da ishlab chiqarilgan oqsillar va RNKlarning cheklangan to'plami B hujayrasini a ga ajratishga undaydi xotira B hujayrasi.[15][22]Va nihoyat, EBV gen ekspressionini yanada cheklaydi va I kechikish vaqtiga kiradi. EBNA-1 ekspressioni B xotira hujayrasi bo'linib ketganda EBV genomini takrorlashga imkon beradi.[15][22]
Epiteliya hujayralari ichida faqat II kechikish mumkin.[iqtibos kerak ]
Birlamchi infeksiyada EBV orofaringeal epiteliya hujayralarida takrorlanib, B-limfotsitlarda III, II va I kechikishlarni o'rnatadi. B-limfotsitlarning EBV yashirin infektsiyasi virusning davom etishi, keyinchalik epiteliya hujayralarida ko'payishi va yuqumli virusning tupurikka chiqishi uchun zarurdir. B-limfotsitlarning EBV Latency III va II infektsiyalari, epiteliya hujayralarining Latency II infektsiyasi va NK- yoki T-hujayrasining Latency II infektsiyalari yomon xulq-atvorga olib kelishi mumkin, bu EBV genomining bir xilligi va gen ekspressioni bilan belgilanadi.[33]
Qayta faollashtirish
B hujayralaridagi yashirin EBV ga o'tish uchun qayta faollashtirilishi mumkin litik replikatsiya. Bu sodir bo'lishi ma'lum jonli ravishda, lekin uni qo'zg'atadigan narsa aniq ma'lum emas. In vitro, B hujayralaridagi yashirin EBV B hujayra retseptorlarini stimulyatsiya qilish orqali qayta faollashtirilishi mumkin, shuning uchun reaktivatsiya jonli ravishda ehtimol yashirin yuqtirilgan B hujayralari o'zaro bog'liq bo'lmagan infektsiyalarga javob berganida sodir bo'ladi.[22]In vitro, B hujayralaridagi yashirin EBV hujayralarni davolash bilan ham faollashtirilishi mumkin natriy butirat yoki 12-O-Tetradekanoilforbol-13-asetat.[iqtibos kerak ]
B-limfotsitlarning transformatsiyasi
EBV B hujayralarini yuqtirganda in vitro, limfoblastoid oxir-oqibat cheksiz o'sishga qodir bo'lgan hujayra chiziqlari paydo bo'ladi. Ushbu hujayra chiziqlarining o'sish o'zgarishi virusli oqsil ekspressionining natijasidir.
EBNA-2, EBNA-3C va LMP-1 konvertatsiya qilish uchun muhim, EBNA-LP va EBERlar esa bunday emas.[34]
EBV bilan tabiiy infektsiyadan so'ng, virus doimiy infektsiyani o'rnatish uchun gen ekspression dasturlarining bir qismini yoki barchasini bajaradi deb o'ylashadi. Dastlab mezbon yo'qligini hisobga olgan holda immunitet, litik tsikl mezbon ichidagi boshqa (taxmin qilingan) B-limfotsitlarni yuqtirish uchun ko'p miqdorda virionlar ishlab chiqaradi.
Yashirin dasturlar virusli B-limfotsitlarni ko'paytirishi va yuqtirgan hujayralarni virus davom etadigan joylarga olib kelish uchun qayta dasturlaydi va o'zgartiradi. Oxir-oqibat, uy egasi immuniteti rivojlanganda, virus o'z genlarining ko'pini (yoki ehtimol hammasini) o'chirib, davom etadi, faqat vaqti-vaqti bilan yangi virionlarni ishlab chiqarish uchun qayta faollashadi. Vaqti-vaqti bilan virusni qayta faollashishi va virusli gen ekspressionini faollashtiradigan immunitet nazorati olib tashlanadigan hujayralar o'rtasida muvozanat o'rnatiladi.
EBV doimiyligi joyi bo'lishi mumkin ilik. O'zining suyak iligi bilan almashtirilgan EBV-musbat bemorlar EBV-salbiy donordan suyak iligi bilan almashtirilgan. transplantatsiya.[35]
Yashirin antijenler
Barcha EBV yadro oqsillari Cp yoki Wp promotorlaridan boshlanib, transkriptni muqobil ravishda qo'shilishi natijasida genom (an'anaviy nomenklaturada). Genlar tartiblangan EBNA-LP / EBNA-2 / EBNA-3A / EBNA-3B / EBNA-3C / EBNA-1 genom ichida.
Boshlanish kodon EBNA-LP kodlash mintaqasi yadro oqsili transkriptining muqobil qo'shilishi bilan hosil bo'ladi. Ushbu boshlang'ich kodoni bo'lmagan taqdirda, EBNA-2 / EBNA-3A / EBNA-3B / EBNA-3C / EBNA-1 ushbu genlarning qaysi biri stenogramma ichiga muqobil ravishda kiritilganiga qarab ifoda etiladi.
Protein / genlar
| Protein / gen / antigen | Bosqich | Tavsif |
|---|---|---|
| EBNA-1 | yashirin + litik | EBNA-1 oqsili virus genomidagi replikatsiya kelib chiqishiga (oriP) bog'lanib, mezbon hujayraning bo'linishi paytida epizomning ko'payishi va bo'linishida vositachilik qiladi. Bu I guruh kechikishida namoyon bo'lgan yagona virusli oqsil. |
| EBNA-2 | yashirin + litik | EBNA-2 asosiy virus hisoblanadi transaktivator. |
| EBNA-3 | yashirin + litik | Ushbu genlar uy egasini ham bog'laydi RBP-Jκ oqsil. |
| LMP-1 | yashirin | LMP-1 olti oraliqdir transmembran EBV vositasida o'sishni o'zgartirish uchun ham zarur bo'lgan oqsil. |
| LMP-2 | yashirin | LMP-2A / LMP-2B - blokirovka qilish uchun harakat qiladigan transmembran oqsillari tirozin kinaz signal berish. |
| EBER | yashirin | EBER-1 / EBER-2 kichik yadro RNKlari bo'lib, ular ma'lum nukleoprotein zarralari bilan bog'lanib, PKR (dsRNA ga bog'liq serin / treonin oqsil kinaz) bilan bog'lanishni ta'minlaydi va shu bilan uning funktsiyasini inhibe qiladi. EBERlar EBV yuqtirilgan hujayralarida transkripsiyaga uchragan eng ko'p EBV mahsulotidir. Ular odatda gistologik to'qimalarda EBV ni aniqlash uchun maqsad sifatida ishlatiladi.[36] ER-zarralari, shuningdek, o'sishni kuchaytiradigan va sitotoksik T-hujayralarni inhibe qiladigan IL-10 ishlab chiqarishni keltirib chiqaradi. |
| v-snoRNA1 | yashirin | Epstein-Barr virusi snoRNA1 virusning kechikish davrida hosil bo'lgan CD-snoRNA qutisi. V-snoRNA1 virusli DNK polimeraza mRNK ning 3'UTR ga yo'naltirilgan 24 ta nukleotidli RNK bo'laklariga qayta ishlangan miRNA o'xshash kashshof bo'lib xizmat qilishi mumkin.[32] |
| ebv-sisRNA | yashirin | Ebv-sisRNA-1 - III kechikish dasturi davomida hosil bo'lgan barqaror intronik ketma-ketlik RNK. Keyin EBER, bu ushbu dastur davomida virus tomonidan ishlab chiqarilgan eng ko'p tarqalgan uchinchi kichik RNK.[37] |
| miRNAlar | yashirin | EBV mikroRNKlari ikkita transkript bilan kodlangan, bittasi to'plamda BART gen va bitta to'plam BHRF1 klaster. Uchtasi BHRF1 pri-miRNAS (to'rtta miRNA hosil qiluvchi) III kechikish vaqtida ifodalanadi, katta klaster esa BART II turdagi kechikish paytida miRNAlar (20 miRNKgacha) ifoda etiladi. Ushbu miRNAlarning vazifalari hozircha noma'lum. |
| EBV-EA | litik | erta antijen |
| EBV-MA | litik | membrana antijeni |
| EBV-VCA | litik | virusli kapsid antigeni |
| EBV-AN | litik | gidroksidi nukleaza[38] |
EBV ning pastki turlari
EBV ikkita asosiy turga bo'linishi mumkin, EBV turi 1 va EBV turi 2. Ushbu ikkita kichik tip har xil EBNA-3 genlar. Natijada, ikkita kichik tip o'zlarining konvertatsiya qilish qobiliyatlari va qayta faollashish qobiliyatlari bilan ajralib turadi. 1-tip dunyoning aksariyat qismida hukmronlik qiladi, ammo ikkala turi ham teng tarqalgan Afrika. Virusli genomni a bilan kesish orqali EBV 1 turini EBV tip 2 dan ajratish mumkin cheklash fermenti va natijada hazm qilish tartiblarini taqqoslash gel elektroforezi.[22]
Kasallikdagi roli
EBV bir nechta kasalliklarga aloqador bo'lib, shu jumladan yuqumli mononuklyoz,[39] Burkitt limfomasi,[40] Xojkin limfomasi,[41] oshqozon saratoni,[13][42] nazofarenks karsinomasi,[43] skleroz,[10][44][11] va limfomatoid granulomatoz.[45] Xususan, EBV bilan kasallangan B-hujayralar miya lezyonlarida joylashganligi isbotlangan skleroz bemorlar.[11] EBV bilan bog'liq bo'lgan qo'shimcha kasalliklar kiradi Janotti-Crosti sindromi, eritema multiforme, o'tkir jinsiy a'zolar yaralari, og'iz tukli leykoplakiya.[46] Virusli infektsiya, shuningdek, zararli bo'lmagan keng doiraga bog'liq va ko'pincha rivojlanishiga hissa qo'shadi limfoproliferativ kasalliklar og'ir yuqori sezuvchanlik kabi Chivin chaqishi allergiyasi reaktsiyalar,[47] Epstein-Barr virusi musbat mukokutanoz yaralari va hydroa vaktsiniforme kabi xavfli lenfoproliferativ kasalliklar Epstein-Barr virusli musbat Burkitt limfomasi,[48] Epshteyn-Barr virusli musbat Xojkin limfomasi,[49] va birlamchi efuzion limfoma.[50]
Epstein-Barr virusi bilan bog'liq kasalliklarga aloqador alfa-sinuklein birlashtirish (masalan, Parkinson kasalligi, Lewy tanalari bilan demans va bir nechta tizim atrofiyasi ).[51]
Tarix
Epstein-Barr virusi nomi bilan atalgan Maykl Entoni Epshteyn (1921 yil 18-mayda tug'ilgan), hozirda professor Bristol universiteti va Yvonne Barr (1932–2016), 1966 yil Ph.D. bitiruvchisi London universiteti, kim birgalikda kashf etdi[52] va 1964 yilda virus mavjudligi to'g'risida nashr etilgan.[53] 1961 yilda Epstein, a patolog va mutaxassis elektron mikroskopist, "Tropik Afrikadagi eng keng tarqalgan bolalar saratoni - shu paytgacha tan olinmagan sindrom" mavzusidagi ma'ruzada qatnashdi. Ushbu ma'ruza, tomonidan Denis Parsons Burkitt, jarroh Uganda, kasallikning "endemik varianti" (pediatrik shakl) tavsifi edi uning ismini oldi. 1963 yilda Ugandadan namuna yuborilgan Midlseks kasalxonasi madaniyatli bo'lish. Kultivatsiya qilingan hujayralarda virus zarralari aniqlandi va natijalar e'lon qilindi Lanset 1964 yilda Epstein tomonidan, Bert Achong va Barr. Uyali aloqa liniyalari yuborildi Verner va Gertruda Xenl da Filadelfiya bolalar kasalxonasi kim rivojlangan serologik markerlar. 1967 yilda ularning laboratoriyasida texnik mutaxassis mononuklyozni rivojlantirdi va ular saqlangan sarum namunasini solishtirishga muvaffaq bo'lishdi. antikorlar rivojlangan virusga.[54][55][56] 1968 yilda ular EBV to'g'ridan-to'g'ri ishlashini aniqladilar abadiylashtirmoq EBV bilan bog'liq infektsiyalarning ba'zi shakllarini taqlid qilib, infektsiyadan keyin B hujayralari,[57] va virus bilan yuqumli mononuklyoz o'rtasidagi bog'liqlikni tasdiqladi.[58]
Tadqiqot
Nisbatan murakkab virus sifatida EBV hali to'liq o'rganilmagan. Dunyo bo'ylab laboratoriyalar virusni o'rganishni davom ettiradi va uning qo'zg'atadigan kasalliklarini davolashning yangi usullarini ishlab chiqadi. EBVni o'rganishning mashhur usullaridan biri in vitro foydalanishdir bakterial sun'iy xromosomalar.[59] Epshteyn-Barr virusini laboratoriyada doimiy kechikish sharoitida saqlab turish va boshqarish mumkin (bu xususiyat birgalikda ishlatiladi) Kaposi sarkomasi bilan bog'liq bo'lgan herpesvirus, sakkizta odamning herpes viruslaridan biri). Tabiiy xostlarni yuqtirish paytida ko'plab viruslar bu xususiyatga ega deb taxmin qilingan bo'lsada, viruslar hayot aylanishining ushbu qismini o'rganish uchun osonlikcha boshqariladigan tizim mavjud emas. EBVning genomik tadqiqotlari litik reaktivatsiyani va yashirin virusli epizomni boshqarishni o'rganishga muvaffaq bo'ldi.[60]
Faol izlanishlar olib borilayotgan bo'lsa-da Epstein-Barr virusiga qarshi emlash hali mavjud emas. Samarali vaktsinani ishlab chiqarish yiliga dunyo miqyosida 200 minggacha saraton kasalligini oldini olish mumkin.[13] Boshqalar singari inson gerpesvirusi "Epshteyn-Barr pro-preparat yordamida yo'q qilishga imkon berishi mumkin Valatsiklovir, ammo yo'q qilish haqiqatan ham amalga oshirilishini aniqlash uchun qo'shimcha tadqiqotlar o'tkazish kerak.[61]
Shuningdek qarang
- Epstein-Barr virusi infektsiyasi
- Epstein-Barr virusi bilan bog'liq lenfoproliferativ kasalliklar
- Jeyms Korson Niderman, Epstein-Barr virusi yuqumli mononuklyozda qanday yuqishini isbotlagan shifokor
Adabiyotlar
- ^ "ICTV taksonomiyasi tarixi: Inson gammaherpesvirusi 4". Viruslar taksonomiyasi bo'yicha xalqaro qo'mita (ICTV). Olingan 10 yanvar 2019.
- ^ Rezk SA, Zhao X, Vayss LM (sentyabr 2018). "Epstein-Barr virusi (EBV) bilan bog'liq bo'lgan limfoid ko'payishi, 2018 yilgi yangilanish". Inson patologiyasi. 79: 18–41. doi:10.1016 / j.humpath.2018.05.020. PMID 29885408.
- ^ Maeda E, Akaxane M, Kiryu S, Kato N, Yoshikava T, Xayashi N va boshq. (Yanvar 2009). "Epstein-Barr virusi bilan bog'liq kasalliklar spektri: rasmli ko'rib chiqish". Yaponiya radiologiya jurnali. 27 (1): 4–19. doi:10.1007 / s11604-008-0291-2. PMID 19373526. S2CID 6970917.
- ^ Cherry-Peppers G, Daniels CO, Meeks V, Sanders CF, Reznik D (2003 yil fevral). "HAART davridagi og'zaki namoyishlar". Milliy tibbiyot birlashmasi jurnali. 95 (2 ta qo'shimcha 2): 21S-32S. PMC 2568277. PMID 12656429.
- ^ Mastria G, Manchini V, Viganò A, Di Piero V (2016). "Alice in Wonderland sindromi: Klinik va patofiziologik tadqiq". BioMed Research International. 2016: 8243145. doi:10.1155/2016/8243145. PMC 5223006. PMID 28116304.
- ^ Nussinovich M, Prais D, Volovitz B, Shapiro R, Amir J (sentyabr 2003). "Bolalarda yuqumli kasallikdan keyingi o'tkir serebellar ataksiya". Klinik pediatriya. 42 (7): 581–4. doi:10.1177/000992280304200702. PMID 14552515. S2CID 22942874.
- ^ Tussirot E, Roudier J (oktyabr 2008). "Otoimmun kasalliklarda Epstein-Barr virusi". Eng yaxshi amaliyot va tadqiqot. Klinik revmatologiya. 22 (5): 883–96. doi:10.1016 / j.berh.2008.09.007. PMID 19028369.
- ^ Dreyfus DH (2011 yil dekabr). "Autoimmun kasallik: Virusga qarshi yangi davolash usullarining ahamiyati?". Autoimmunity Sharhlari. 11 (2): 88–97. doi:10.1016 / j.autrev.2011.08.005. PMID 21871974.
- ^ Pender MP (2012). "CD8 + T-hujayra etishmovchiligi, Epshteyn-Barr virusi yuqishi, D vitamini etishmovchiligi va avtoimmunitetga qadamlar: birlashtiruvchi gipoteza". Otoimmun kasalliklar. 2012: 189096. doi:10.1155/2012/189096. PMC 3270541. PMID 22312480.
- ^ a b Ascherio A, Munger KL (2010 yil sentyabr). "Epstein-barr virusi infektsiyasi va skleroz: qayta ko'rib chiqish". Neyroimmun farmakologiya jurnali. 5 (3): 271–7. doi:10.1007 / s11481-010-9201-3. PMID 20369303. S2CID 24409610.
- ^ a b v Moreno MA, Or-Geva N, Aftab BT, Xanna R, Croze E, Steinman L, Xan MH (iyul 2018). "MS miya shikastlanishlarida Epstein-Barr virusi infektsiyasining molekulyar imzosi". Nevrologiya. 5 (4): e466. doi:10.1212 / NXI.0000000000000466. PMC 5994704. PMID 29892607.
- ^ Xon, Gulfaraz; Miyashita, Emili M.; Yang, Bin; Babkok, Gregori J.; Torli-Louson, Devid A. (2018 yil 17-noyabr). "Epstein-Barr virusi ko'p miqdordagi skleroz holatlarining miyasida uchraydi va B hujayralariga qaraganda ko'proq ishtirok etishi mumkin". PLOS ONE. San-Fransisko, Kaliforniya: PLOS. 13 (2): e0192109. Bibcode:2018PLoSO..1392109H. doi:10.1371 / journal.pone.0192109. PMC 5796799. PMID 29394264.
- ^ a b v "Epstein-Barr virusi uchun vaktsinani ishlab chiqish global miqyosda 200 minggacha saraton kasalligini oldini olishga imkon beradi". Cancer Research UK (Matbuot xabari). 2014 yil 24 mart. Arxivlandi asl nusxasidan 2017 yil 19 martda.
- ^ Xon, Gulfaraz; Hoshim, Muhammad Javad (2014 yil noyabr). "Epstein-Barr virusidan o'limning global yuki 1990-2010 yillar. Yuqumli vositalar va saraton. London, Angliya: BioMed Central. 9 (1): 38. doi:10.1186/1750-9378-9-38. PMC 4253616. PMID 25473414.
- ^ a b v d e f g h men Amon V, Farrell PJ (2004 yil noyabr). "Epsteyn-Barr virusini kechikishdan reaktivatsiyasi". Tibbiy virusologiya bo'yicha sharhlar. 15 (3): 149–56. doi:10.1002 / rmv.456. PMID 15546128.
- ^ Taxminan 90% kattalarda antikorlar mavjud bo'lib, ular hozirgi yoki o'tmishda EBV infektsiyasini olganligini ko'rsatadi. Arxivlandi 2016-08-08 da Orqaga qaytish mashinasi Yuqumli kasalliklar milliy markazi
- ^ ACP. "Epstein-Barr virusli infektsiyalari: biologiya, patogenezi va boshqaruvi". ACP. Arxivlandi asl nusxasidan 2017 yil 8 dekabrda. Olingan 8 dekabr 2017.
- ^ CDC. "Epstein-Barr virusi va yuqumli mononuklyoz". CDC. Arxivlandi asl nusxasi 2012 yil 20 aprelda. Olingan 29 dekabr 2011.
- ^ Khan G, Miyashita EM, Yang B, Babcock GJ, Thorley-Lawson DA (Avgust 1996). "EBV qat'iyligi in vivo jonli ravishda B hujayra gomeostazining namunasimi?". Immunitet. 5 (2): 173–9. doi:10.1016 / s1074-7613 (00) 80493-8. PMID 8769480.
- ^ "Epstein-Barr virusi (EBV) to'g'risida". Kasalliklarni nazorat qilish va oldini olish markazlari. 14 sentyabr 2016 yil.
- ^ Curr Clin Top yuqumli kasalliklar. 1988; 9: 126-46.
- ^ a b v d e f g h men j k l m n o p q Odumade OA, Hogquist KA, Balfour HH (yanvar 2011). "Birlamchi Epstein-Barr virusli infektsiyalarini tushunish va boshqarish bo'yicha taraqqiyot va muammolar". Klinik mikrobiologiya sharhlari. 24 (1): 193–209. doi:10.1128 / CMR.00044-10. PMC 3021204. PMID 21233512.
- ^ Jia, Liu (2020 yil 17-iyul). "Olimlar Epstein-Bar virusi nukleokapsidining birinchi atom tuzilishini kashf etdilar". phys.org. Olingan 4 oktyabr 2020.
- ^ Li Z, Zhang X, Dong L, Pang J, Xu M, Zhong Q, Zeng MS, Yu X (oktyabr 2020). "Epstein-Barr virusining tegument kapsidining krioEM tuzilishi". Hujayra tadqiqotlari. 30 (10): 873–884. doi:10.1038 / s41422-020-0363-0. PMID 32620850. S2CID 220309464.
- ^ Shannon-Lou, C, Rou, M (fevral, 2014). "Epstein Barr virusiga kirish; o'pish va konjugatsiya". Virusshunoslikning dolzarb fikri. 4: 78–84. doi:10.1016 / j.coviro.2013.12.12.001. PMID 24553068.
- ^ Van X, Xatt-Fletcher LM (1998 yil yanvar). "Glikoprotein gp42 ga ega bo'lmagan Epstein-Barr virusi B hujayralari bilan bog'lanishi mumkin, ammo yuqtirishga qodir emas". Virusologiya jurnali. 72 (1): 158–63. doi:10.1128 / jvi.72.1.158-163.1998. PMC 109360. PMID 9420211.
- ^ a b "Entrez Gen: CR2 komplement komponenti (3d / Epstein Barr virusi) retseptorlari 2". ncbi.nlm.nih.gov/. Arxivlandi asl nusxasidan 2010 yil 5 dekabrda.
- ^ Ogembo JG, Kannan L, Giran I, Nikolson-Weller A, Finberg RW, Tsokos GC, Fingeroth JD (2013 yil fevral). "1 / CD35 tipdagi inson komplementlari retseptorlari - bu Epstein-Barr virusli retseptorlari". Hujayra hisobotlari. 3 (2): 371–85. doi:10.1016 / j.celrep.2013.01.023. PMC 3633082. PMID 23416052.
- ^ a b v Liu S, Li X, Chen L, Yang L, Li L, Tao Y va boshqalar. (2013 yil mart). "(-) - Epigallokatechin-3-gallat inhibitsiyasi Epstein-Barr virusining o'z-o'zidan paydo bo'lgan litik infektsiyasi EBV-musbat hujayralarida ERK1 / 2 va PI3-K / Akt signalizatsiyasini o'z ichiga oladi". Kanserogenez. 34 (3): 627–37. doi:10.1093 / karsin / bgs364. PMID 23180656.
- ^ Torli-Louson DA, Miyashita EM, Xan G (1996 yil may). "Epstein-Barr virusi va B hujayrasi: buning uchun kifoya qiladi". Mikrobiologiya tendentsiyalari. 4 (5): 204–8. doi:10.1016 / s0966-842x (96) 90020-7. PMID 8727601.
- ^ Calderwood MA, Venkatesan K, Xing L, Chase MR, Vazkes A, Xolthaus AM va boshq. (2007 yil may). "Epshteyn-Barr virusi va virusi odam oqsilining o'zaro ta'sir xaritalari". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 104 (18): 7606–11. Bibcode:2007PNAS..104.7606C. doi:10.1073 / pnas.0702332104. PMC 1863443. PMID 17446270. Bu erda ishlatiladigan nomenklatura Kieffga tegishli. Boshqa laboratoriyalar turli nomenklaturalardan foydalanadilar.
- ^ a b Xutzinger R, Feederle R, Mrazek J, Schiefermeier N, Balwierz PJ, Zavolan M va boshq. (Avgust 2009). "Epstein-Barr virusi genomidan kichik nukleolyar RNKning ifodasi va qayta ishlanishi". PLOS patogenlari. 5 (8): e1000547. doi:10.1371 / journal.ppat.1000547. PMC 2718842. PMID 19680535.
- ^ Robertson ES, ed. (2010). Epshteyn-Barr virusi: kechikish va o'zgarish. Caister Academic Press. ISBN 978-1-904455-62-2.
- ^ Yates JL, Uorren N, Sugden B (1985). "Epstein-Barr virusidan olingan plazmidlarning turli xil sutemizuvchilar hujayralarida barqaror replikatsiyasi". Tabiat. 313 (6005): 812–5. Bibcode:1985 yil 313..812Y. doi:10.1038 / 313812a0. PMID 2983224. S2CID 4334367.
- ^ Gratama JW, Oosterveer MA, Zwaan FE, Lepoutre J, Klein G, Ernberg I (noyabr 1988). "Suyak iligi transplantatsiyasi orqali epsteyn-Barr virusini yo'q qilish: virusning kechikish joylariga ta'siri". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 85 (22): 8693–6. Bibcode:1988 yil PNAS ... 85.8693G. doi:10.1073 / pnas.85.22.8693. PMC 282526. PMID 2847171.
- ^ Khan G, Coates PJ, Kangro HO, Slavin G (iyul 1992). "Epstein Barr virusi (EBV) kichik RNKlarni kodladi: oligonukleotid zondlari bilan joyida gibridlanish orqali aniqlash maqsadlari". Klinik patologiya jurnali. 45 (7): 616–20. doi:10.1136 / jcp.45.7.616. PMC 495191. PMID 1325480.
- ^ Moss WN, Steitz JA (2013 yil avgust). "Epstein-Barr virusining genom bo'yicha tahlillari natijasida saqlanib qolgan RNK tuzilmalari va yangi barqaror intronik ketma-ketlik RNK aniqlandi". BMC Genomics. 14: 543. doi:10.1186/1471-2164-14-543. PMC 3751371. PMID 23937650.
- ^ Buisson M, Géoui T, Flot D, Tarbouriech N, Ressing ME, Wiertz EJ, Burmeister WP (avgust 2009). "Ko'prik DNase va RNase faoliyati bilan Epstein-Barr virusi nukleazasining faol joyini kesib o'tadi". Molekulyar biologiya jurnali. 391 (4): 717–28. doi:10.1016 / j.jmb.2009.06.034. PMID 19538972.
- ^ Vays LM, O'Malley D (yanvar 2013). "Xavfsiz limfadenopatiyalar". Zamonaviy patologiya. 26 Qo'shimcha 1 (Qo'shimcha 1): S88-96. doi:10.1038 / modpathol.2012.176. PMID 23281438.
- ^ Pannone G, Zamparese R, Pace M, Pedicillo MC, Cagiano S, Somma P va boshq. (2014). "Burkitt limfomasi patogenezida EBV ning roli: Italiya kasalxonasida o'tkazilgan so'rov". Yuqumli vositalar va saraton. 9 (1): 34. doi:10.1186/1750-9378-9-34. PMC 4216353. PMID 25364378.
- ^ Gandi MK, Tellam JT, Xanna R (2004 yil may). "Epshteyn-Barr virusi bilan bog'liq Xodkin limfomasi". Britaniya gematologiya jurnali. 125 (3): 267–81. doi:10.1111 / j.1365-2141.2004.04902.x. PMID 15086409. S2CID 2355660.
- ^ Yau, TO; Tang, CM; Yu, J (2014 yil 7-iyun). "Epstein-Barr virusi bilan bog'liq oshqozon karsinomasida epigenetik disregulyatsiya: kasallik va davolash usullari". Jahon Gastroenterologiya jurnali. 20 (21): 6448–56. doi:10.3748 / wjg.v20.i21.6448. PMC 4047330. PMID 24914366.
- ^ Dogan S, Hedberg ML, Ferris RL, Rath TJ, Assaad AM, Chiosea SI (2014 yil aprel). "Inson papillomavirusi va Epstein-Barr virusi kam uchraydigan populyatsiyada nazofarenks karsinomasida". Bosh va bo'yin. 36 (4): 511–6. doi:10.1002 / hed.23318. PMC 4656191. PMID 23780921.
- ^ Mechelli R, Manzari C, Policano C, Annese A, Picardi E, Umeton R va boshq. (Mart 2015). "Epstein-Barr virusining genetik variantlari ko'p skleroz bilan bog'liq". Nevrologiya. 84 (13): 1362–8. doi:10.1212 / WNL.0000000000001420. PMC 4388746. PMID 25740864.
- ^ Tagliavini E, Rossi G, Valli R, Zanelli M, Cadioli A, Mengoli MC va boshq. (2013 yil avgust). "Lenfomatoid granulomatoz: bu noyob o'pka lenfoproliferativ jarayoni bilan shug'ullanadigan patologlar uchun amaliy sharh". Patologika. 105 (4): 111–6. PMID 24466760.
- ^ Di Lernia V, Mansuriy Y (oktyabr 2013). "Epstein-Barr virusi va bolalik davrida terining namoyon bo'lishi". Xalqaro dermatologiya jurnali. 52 (10): 1177–84. doi:10.1111 / j.1365-4632.2012.05855.x. PMID 24073903. S2CID 30557756.
- ^ Kyriakidis I, Vasileiou E, Karastrati S, Tragiannidis A, Gompakis N, Hatzistilianou M (dekabr 2016). "Birlamchi EBV infektsiyasi va chivin chaqishiga yuqori sezuvchanlik: voqea bayonoti". Virologica Sinica. 31 (6): 517–520. doi:10.1007 / s12250-016-3868-4. PMID 27900557. S2CID 7996104.
- ^ Navari M, Etebari M, De Falco G, Ambrosio MR, Gibellini D, Leoncini L, Piccaluga PP (2015). "Epstein-Barr virusining mavjudligi immunitet tanqisligi bilan bog'liq Burkitt limfomasida transkripsiyaviy profilga sezilarli ta'sir qiladi". Mikrobiologiya chegaralari. 6: 556. doi:10.3389 / fmicb.2015.00556. PMC 4462103. PMID 26113842.
- ^ Shannon-Lou C, Rikkinson AB, Bell AI (oktyabr 2017). "Epstein-Barr virusi bilan bog'liq lenfomalar". London Qirollik Jamiyatining falsafiy operatsiyalari. B seriyasi, Biologiya fanlari. 372 (1732): 20160271. doi:10.1098 / rstb.2016.0271. PMC 5597738. PMID 28893938.
- ^ Arora N, Gupta A, Sadegi N (iyul 2017). "Birlamchi effuzion limfoma: hozirgi tushunchalar va boshqarish". O'pka tibbiyotidagi hozirgi fikr. 23 (4): 365–370. doi:10.1097 / MCP.0000000000000384. PMID 28399009. S2CID 4514140.
- ^ Woulfe J, Hoogendoorn H, Tarnopolsky M, Muñoz DG (2000 yil noyabr). "Epstein-Barr virusiga qarshi monoklonal antikorlar inson miyasida alfa-sinuklein bilan o'zaro ta'sir qiladi". Nevrologiya. 55 (9): 1398–401. doi:10.1212 / WNL.55.9.1398. PMID 11087792. S2CID 84387269.
- ^ McGrath P (2014 yil 6-aprel). "Saraton virusini aniqlashga kechiktirilgan parvoz yordam berdi". BBC yangiliklari, sog'liqni saqlash. Arxivlandi asl nusxasidan 2015 yil 8 oktyabrda. Olingan 4 noyabr 2015.
- ^ Epstein MA, Achong BG, Barr YM (mart 1964). "Burkitt limfomasidan ekilgan limfoblastlardagi virus zarralari". Lanset. 1 (7335): 702–3. doi:10.1016 / s0140-6736 (64) 91524-7. PMID 14107961.
- ^ Epstein MA (2005). "1. EBV tadqiqotining kelib chiqishi: virusni kashf qilish va tavsiflash". Robertson ES-da (tahrir). Epstein-Barr virusi. Trowbridge: Cromwell Press. 1-14 betlar. ISBN 978-1-904455-03-5. Olingan 18 sentyabr 2010.
- ^ Erle S. Robertson (2005). Epstein-Barr virusi. Horizon Scientific Press. p. 18. ISBN 978-1-904455-03-5. Olingan 3 iyun 2012.
- ^ Miller G (2006 yil 21-dekabr). "Kitoblarni ko'rib chiqish: Epstein-Barr virusi". Nyu-England tibbiyot jurnali. 355 (25): 2708–2709. doi:10.1056 / NEJMbkrev39523.
- ^ Henle V, Henle G (1980). "Epstein-Barr virusi (EBV) bilan bog'liq kasalliklarning epidemiologik jihatlari". Nyu-York Fanlar akademiyasining yilnomalari. 354: 326–31. doi:10.1111 / j.1749-6632.1980.tb27975.x. PMID 6261650. S2CID 30025994.
- ^ Young, LS (2009). Inson va tibbiy virusologiya ish stoli entsiklopediyasi. Boston: Academic Press. 532-533 betlar.
- ^ Delecluse HJ, Feederle R, Behrends U, Mautner J (dekabr 2008). "Epstein-Barr virusiga qarshi immunitetni o'rganishda virusli rekombinantlarning hissasi". Saraton biologiyasi bo'yicha seminarlar. 18 (6): 409–15. doi:10.1016 / j.semcancer.2008.09.001. PMID 18938248.
- ^ Arvey A, Tempera I, Tsay K, Chen XS, Tixmyanova N, Klichinskiy M va boshq. (Avgust 2012). "Epstein-Barr virusi transkriptomi va epigenomining atlasi xost-virusning regulyatsion o'zaro ta'sirini ochib beradi". Cell Host & Microbe. 12 (2): 233–45. doi:10.1016 / j.chom.2012.06.008. PMC 3424516. PMID 22901543.
- ^ Hoshino Y, Katano H, Zou P, Hohman P, Marques A, Tyring SK va boshq. (2009 yil noyabr). "Valatsiklovirin uzoq muddat tatbiq etilishi Epstein-Barr virusi (EBV) bilan zararlangan B hujayralari sonini kamaytiradi, ammo sog'lom ko'ngillilarning B hujayrasiga EBV DNK nusxalari sonini emas". Virusologiya jurnali. 83 (22): 11857–61. doi:10.1128 / JVI.01005-09. PMC 2772668. PMID 19740997.
Tashqi havolalar
Tashqi havolalar
 Bilan bog'liq ommaviy axborot vositalari Epstein-Barr virusi Vikimedia Commons-da
Bilan bog'liq ommaviy axborot vositalari Epstein-Barr virusi Vikimedia Commons-da Bilan bog'liq ma'lumotlar Epstein-Barr virusi Vikipediya sahifalarida
Bilan bog'liq ma'lumotlar Epstein-Barr virusi Vikipediya sahifalarida- Vikidata: mavzu (Scholia )