Fosfofruktokinaza 1 - Phosphofructokinase 1
| 6-fosfofruktokinaza | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifikatorlar | |||||||||
| EC raqami | 2.7.1.11 | ||||||||
| CAS raqami | 9001-80-3 | ||||||||
| Ma'lumotlar bazalari | |||||||||
| IntEnz | IntEnz ko'rinishi | ||||||||
| BRENDA | BRENDA kirish | ||||||||
| ExPASy | NiceZyme ko'rinishi | ||||||||
| KEGG | KEGG-ga kirish | ||||||||
| MetaCyc | metabolik yo'l | ||||||||
| PRIAM | profil | ||||||||
| PDB tuzilmalar | RCSB PDB PDBe PDBsum | ||||||||
| Gen ontologiyasi | AmiGO / QuickGO | ||||||||
| |||||||||
| Fosfofruktokinaza | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifikatorlar | |||||||||||
| Belgilar | PFK | ||||||||||
| Pfam | PF00365 | ||||||||||
| Pfam klan | CL0240 | ||||||||||
| InterPro | IPR000023 | ||||||||||
| PROSITE | PDOC00336 | ||||||||||
| SCOP2 | 5pfk / QOIDA / SUPFAM | ||||||||||
| |||||||||||
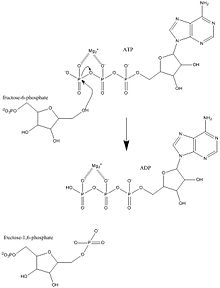
Fosfofruktokinaza-1 (PFK-1) eng muhim tartibga soluvchilardan biridir fermentlar (EC 2.7.1.11 ) ning glikoliz. Bu allosterik ferment 4 subbirlikdan iborat va ko'pchilik tomonidan boshqariladi aktivatorlar va inhibitörler. PFK-1 glikolizning muhim "soddalashtirilgan" bosqichini katalizlaydi fruktoza 6-fosfat va ATP fruktoza 1,6-bifosfat va ADP. Glikoliz anaerob va aerob nafas olish uchun asosdir. Fosfofruktokinaza (PFK) fruktoza-6-fosfatni fruktoza 1,6-bifosfat va ADP ga aylantirish uchun ATPga bog'liq fosforillanishni katalizatori qilganligi sababli u glikolizning muhim tartibga solish bosqichlaridan biri hisoblanadi. PFK glikolizni allosterik inhibisyon orqali boshqarishga qodir va shu bilan hujayra hujayraning energiya talabiga javoban glikoliz tezligini oshirishi yoki kamaytirishi mumkin. Masalan, ATP va ADP ning yuqori nisbati PFK va glikolizni inhibe qiladi. Eukaryotlar va prokaryotlarda PFK regulyatsiyasi o'rtasidagi asosiy farq shundaki, eukaryotlarda PFK fruktoza 2,6-bifosfat bilan faollashadi. Fruktoza 2,6-bifosfatning maqsadi ATP inhibisyonunun o'rnini bosishdir, shuning uchun eukaryotlarning glyukagon va insulin kabi gormonlar tomonidan tartibga solinishi uchun ko'proq sezgir bo'lishiga imkon beradi.[1]
| β-D.-fruktoza 6-fosfat | Fosfofruktokinaza 1 | β-D.-fruktoza 1,6-bifosfat | |
 |  | ||
| ATP | ADP | ||
| Pmen | H2O | ||
| Fruktoza bifosfataza | |||
Tuzilishi
Sutemizuvchilar PFK1 340kd[2] tetramer uch xil bo'linmaning turli xil kombinatsiyalaridan iborat: muskul (M), jigar (L) va trombotsit (P). PFK1 tarkibi tetramer u mavjud bo'lgan to'qima turiga qarab farq qiladi. Masalan, etuk mushak faqat M ni ifodalaydi izozim shuning uchun PFK1 mushaklari faqat M4 homotetramerlaridan iborat. Jigar va buyraklar asosan L izoformini ifoda etadi. Yilda eritrotsitlar, ikkala M va L subbirliklari tasodifiy ravishda tetramerlanib, M4, L4 va fermentning uchta gibrid shaklini hosil qiladi (ML3, M2L2, M3L). Natijada, turli xil izoenzimlar havzalarining kinetik va tartibga solish xususiyatlari subunit tarkibiga bog'liq. PFK faolligi va izoenzim tarkibidagi to'qimalarga xos o'zgarishlar turli xilligiga sezilarli hissa qo'shadi glikolitik va glyukoneogen turli to'qimalar uchun kuzatilgan stavkalar.[3]
PFK1 allosterik ferment bo'lib, tuzilishga o'xshash tuzilishga ega gemoglobin dimerning dimeriga qadar.[4] Har bir dimerning yarmida ATP bog'lanish joyi, qolgan yarmida substrat (fruktoza-6-fosfat yoki (F6P)) bog'lanish joyi va alohida allosterik bog'lanish joyi mavjud.[5]
Tetramerning har bir bo'linmasi 319 ta aminokislotadan iborat bo'lib, ikkita domendan iborat: biri ATP substratni, ikkinchisi fruktoza-6-fosfatni bog'laydi. Har bir domen b bochkadan iborat bo'lib, alfa spirallari bilan o'ralgan silindrsimon b varag'iga ega.
Har bir faol saytning har bir kichik birligining qarama-qarshi tomonida dimerdagi bo'linmalar orasidagi interfeysda allosterik sayt joylashgan. ATP va AMP ushbu sayt uchun raqobatlashadi. N-terminal domeni ATP ni bog'laydigan katalitik rolga ega va C-terminal tartibga soluvchi rolga ega [6]
Mexanizm
PFK1 allosterik ferment bo'lib, uning faolligini simmetriya modeli allosterizm[7] bu erda fermentativ faol bo'lmagan T holatidan faol R holatiga kelishilgan o'tish mavjud. F6P R holatiga yuqori yaqinlik bilan bog'lanadi, ammo T holati fermenti emas. PFK1 bilan bog'langan har bir F6P molekulasi uchun ferment asta-sekin T holatidan R holatiga o'tadi. Shunday qilib, F6P kontsentratsiyasining ko'payishiga qarshi PFK1 faolligini tuzadigan grafik quyidagicha qabul qilinadi sigmasimon an'anaviy ravishda allosterik fermentlar bilan bog'langan egri shakli.
PFK1 oilasiga tegishli fosfotransferazlar va u b-fosfatning ATP dan fruktoza-6-fosfatga o'tishini katalizlaydi. PFK1 faol sayt ATP-Mg2 + va F6P ulanish joylarini o'z ichiga oladi. Substratni biriktirish bilan bog'liq ba'zi taklif qilingan qoldiqlar E. coli PFK1 tarkibiga kiradi Asp127 va Arg171.[8] Yilda B. stearotermofil PFK1, Arg162 qoldig'ining musbat zaryadlangan yon zanjiri, F6P ning salbiy zaryadlangan fosfat guruhi bilan vodorod bilan bog'langan tuz ko'prigini hosil qiladi, bu o'zaro ta'sir R holatini T holatiga nisbatan barqarorlashtiradi va qisman homotrop ta'sir F6P majburiyligi. T holatida ferment konformatsiyasi biroz siljiydi, shunday qilib Arg162 ilgari egallagan bo'shliq bilan almashtiriladi Glu161. Qo'shni aminokislota qoldiqlari orasidagi almashinuv F6P ning fermentni bog'lash qobiliyatini inhibe qiladi.
Kabi allosterik aktivatorlar AMP va ADP allosterik joy bilan bog'lanib, ferment tarkibidagi tarkibiy o'zgarishlarni keltirib chiqarish orqali R holatini hosil bo'lishiga yordam beradi. Xuddi shunday, ATP va kabi inhibitorlar PEP bir xil allosterik joy bilan bog'lanib, T holatining shakllanishini osonlashtiradi va shu bilan ferment faolligini inhibe qiladi.
1-uglerodning gidroksil kislorodi ATP ning beta fosfatiga nukleofil hujum qiladi. Ushbu elektronlar ATP ning beta va gamma fosfatlari orasidagi angidrid kislorodga suriladi.[9][10]
Tartibga solish
PFK1 sutemizuvchilarning glikolitik yo'lidagi eng muhim nazorat joyidir. Ushbu qadam keng qamrovli tartibga solinishi kerak, chunki u nafaqat yuqori darajada eksergonik ostida fiziologik sharoit, shuningdek, bu qat'iy qadam bo'lgani uchun - glikolitik yo'lga xos bo'lgan birinchi qaytarilmas reaktsiya. Bu glyukoza va boshqasini aniq nazorat qilishga olib keladi monosaxaridlar galaktoza va fruktoza glikolitik yo'ldan pastga tushish. Ushbu ferment reaktsiyasidan oldin, glyukoza-6-fosfat potentsial pastga qarab yurishi mumkin pentoza fosfat yo'li, yoki uchun glyukoza-1-fosfatga aylantiriladi glikogenez.
PFK1 bu allosterik tarzda ning yuqori darajasi bilan inhibe qilinadi ATP ammo AMP ATP ning inhibitiv ta'sirini qaytaradi. Shuning uchun hujayra ATP / AMP nisbati pasayganda fermentning faolligi oshadi. Shunday qilib, energiya zaryadi tushganda glikoliz rag'batlantiriladi. PFK1-da ATP uchun turli xil affinitatsiyali ikkita sayt mavjud, ular ikkalasi ham substrat va inhibitor.[2]
PFK1, shuningdek, ATP ning inhibitiv ta'sirini kuchaytiradigan past pH darajalari bilan inhibe qilinadi. PH mushaklar ishlayotganda tushadi anaerob va ortiqcha miqdorda ishlab chiqarish sut kislotasi (sut kislotasi o'zi pH pasayishiga sabab bo'lmasa ham[11]). Ushbu inhibitiv ta'sir mushakni juda ko'p kislota to'planishidan kelib chiqadigan zararlardan himoya qilishga xizmat qiladi.[2]
Nihoyat, PFK1 allosterik tarzda inhibe qilinadi PEP, sitrat va ATP. Fosfoenolpiruv kislotasi quyi oqimdagi mahsulotdir glikolitik yo'l. Krebs tsikli fermentlari maksimal tezligiga yaqinlashganda sitrat paydo bo'lishiga qaramay, sitrat normal fiziologik sharoitda PFK-1 ni inhibe qilish uchun etarli konsentratsiyaga to'planib qoladimi, bu shubhali.[iqtibos kerak ]. ATP kontsentratsiyasining ko'payishi energiyaning ortiqcha ekanligini ko'rsatadi va PFK1da allosterik modulyatsiya joyiga ega bo'lib, u PFK1 ning substratga yaqinligini pasaytiradi.
PFK1 bu allosterik tarzda ning yuqori konsentratsiyasi bilan faollashadi AMP, lekin eng kuchli aktivator fruktoza 2,6-bifosfat, shuningdek, fruktoza-6-fosfatdan ishlab chiqariladi PFK2. Demak, F6P ning ko'pligi yuqori konsentratsiyaga olib keladi fruktoza 2,6-bifosfat (F-2,6-BP). F-2,6-BP ning bog'lanishi PFK1 ning F6P ga yaqinligini oshiradi va ATP ning inhibitiv ta'sirini pasaytiradi. Bu glyukoza ko'p bo'lganida glikoliz tezlashishi sababli, ilgari stimulyatsiya qilishning misoli.[2]
PFK faolligi sintezning repressiyasi orqali kamayadi glyukagon. Glyukagon faollashadi oqsil kinazasi A bu esa o'z navbatida kinaz faolligini o'chiradi PFK2. Bu F6P dan F-2,6-BP har qanday sintezini qaytaradi va shu bilan PFK1ni o'chiradi.
PFK1ni aniq tartibga solish oldini oladi glikoliz va glyukoneogenez bir vaqtning o'zida sodir bo'lishidan. Biroq, mavjud substrat velosipedda harakatlanish F6P va F-1,6-BP o'rtasida. Fruktoza-1,6-bifosfataza (FBPase) F-1,6-BP gidrolizini F6P ga qaytaradi, teskari reaktsiya PFK1 tomonidan katalizlanadi. Glikoliz paytida oz miqdordagi FBPaza faolligi va glyukoneogenez paytida ba'zi PFK1 faolligi mavjud. Ushbu tsikl metabolik signallarni kuchaytirish bilan bir qatorda ATP gidrolizida issiqlik hosil bo'lishiga imkon beradi.
Serotonin (5-HT) 5-HT (2A) retseptorlari bilan bog'lanish orqali PFKni ko'paytiradi va PFK tirozin qoldig'ini fosfolipaza S orqali fosforillanishiga olib keladi, bu esa o'z navbatida skelet mushak hujayralarida PFKni qayta taqsimlaydi. PFK glikolitik oqimni boshqargani uchun serotonin glikolizda regulyator rolini o'ynaydi [12]
Genlar
Odamlarda uchta fosfofruktokinaz geni mavjud:
Klinik ahamiyati
Genetik mutatsiya PFKM gen natijalari Tarui kasalligi, bu glikogenni saqlash kasalligi, bu erda ba'zi hujayralar turlaridan foydalanish qobiliyati uglevodlar energiya manbai sifatida buzilgan.[13]
Tarui kasalligi - bu glikogenni saqlash kasalligi, shu jumladan, mushaklarning kuchsizligi (miyopatiya) va jismoniy mashqlar natijasida kelib chiqadigan kramplar va spazmlar, miyoglobinuriya (siydikda miyoglobin borligi, mushaklarning yo'q qilinishini bildiradi) va kompensatsiyalangan gemoliz. ATP glikoliz orqali keraksiz ishlab chiqarilishining oldini olish maqsadida ATF PFKning tabiiy allosterik inhibitori hisoblanadi. Shu bilan birga, Asp (543) Aladagi mutatsiya ATP ning kuchli inhibitiv ta'sirga olib kelishiga olib kelishi mumkin (PFKning inhibitor allosterik bog'lanish joyiga ulanish kuchayganligi sababli).[14][15]
Fosfofruktokinaza mutatsiyasi va saraton: saraton hujayralari tez o'sishi va bo'linishi tufayli energiya talablarini qondirishi uchun ular giperaktiv fosfofruktokinaza 1 fermentiga ega bo'lganda samarali yashaydilar.[16][17] Saraton hujayralari tez o'sib, bo'linib ketganda, dastlab ular qon bilan ta'minlanmaydi va shu bilan gipoksiya (kislorod etishmovchiligi) bo'lishi mumkin va bu tetiklashadi O-GlcNAsilatsiya PFKning 529 serinasida. Ushbu modifikatsiya PFK1 faolligini inhibe qiladi va saraton kasalligining ko'payishini qo'llab-quvvatlaydi, aksincha yuqori PFK1 faolligi saraton kasalligi uchun zarurdir. Buning sababi glyukoza oqimini reaktiv kislorod turlarini zararsizlantirish uchun NADPH hosil qilish uchun pentozfosfat yo'liga yo'naltirish bo'lishi mumkin.[18]
Herpes simplex 1 turi va fosfofruktokinaza: Ba'zi viruslar, jumladan OIV, HCMV va Mayaro, PFK faolligining MOIga bog'liqligi ortishi natijasida glikoliz kabi hujayralardagi metabolik yo'llarga ta'sir qiladi. Herpesning PFK faolligini oshiradigan mexanizm serin qoldiqlarida fermentni fosforlashdir. HSV-1 tomonidan induktsiya qilingan glikoliz ATP tarkibini oshiradi, bu virusning ko'payishi uchun juda muhimdir.[19]
Shuningdek qarang
- PFK2 (fruktoza 6-fosfatni fruktoza 2,6-bifosfatga aylantirilgan joyda yoki aksincha, boshqa joyda o'tkazadi)
- PFP (fruktoza 6-fosfat va fruktoza 1,6-bifosfatni anorganik yordamida reversiv ravishda o'zaro o'zgartiradi pirofosfat ATP o'rniga)
- Fruktoza bifosfataza (fruktoza 1,6-bifosfatni fruktoza 6-fosfatga gidroliz qiladi)
Adabiyotlar
- ^ Usenik A, Legiša M (noyabr 2010). Kobe B (tahrir). "6-fosfofructo-1-kinazda allosterik sitrat bilan bog'lanish joylarining evolyutsiyasi". PLOS ONE. 5 (11): 677–683. doi:10.1371 / journal.pone.0015447. PMC 2990764. PMID 21124851.
- ^ a b v d Stryer L, Berg JM, Timoczko JL (2007). Biokimyo (Oltinchi nashr). San-Fransisko: W.H. Freeman. ISBN 978-0-7167-8724-2.
- ^ Dunaway GA, Kasten TP, Sebo T, Trapp R (1988 yil may). "Inson to'qimalarida fosfofruktokinaza subbirliklarini va izoenzimlarini tahlil qilish". Biokimyo. J. 251 (3): 677–83. doi:10.1042 / bj2510677. PMC 1149058. PMID 2970843.
- ^ PDB: 4pfk; Evans PR, Farrants GW, Hudson PJ (iyun 1981). "Fosfofruktokinaza: tuzilishi va boshqarilishi". Qirollik jamiyatining falsafiy operatsiyalari B. 293 (1063): 53–62. doi:10.1098 / rstb.1981.0059. PMID 6115424. Xulosa – Oyning PDB molekulasi.
- ^ Shirakihara Y, Evans PR (dekabr 1988). "Escherichia coli-dan fosfofruktokinaza kompleksining uning reaksiya mahsulotlari bilan kristalli tuzilishi". J. Mol. Biol. 204 (4): 973–94. doi:10.1016/0022-2836(88)90056-3. PMID 2975709.
- ^ Banaszak K, Mechin I, Obmolova G, Oldxem M, Chang SH, Ruiz T, Radermaxer M, Kopperschläger G, Rypnevski V (mart 2011). "Beyker xamirturushidan va quyon skelet mushaklaridan eukaryotik fosfofruktokinazalarning kristalli tuzilmalari". J Mol Biol. 407 (7): 284–97. doi:10.1016 / j.jmb.2011.01.019. PMID 21241708.
- ^ Peskov K, Goryanin I, Demin O (2008 yil avgust). "Escherichia coli-dan fosfofruktokinaza-1ning kinetik modeli". J Bioinform Comput Biol. 6 (4): 843–67. doi:10.1142 / S0219720008003643. PMID 18763746.
- ^ Hellinga HW, Evans PR (1987). "Escherichia coli fosfofruktokinazasining faol joyidagi mutatsiyalar". Tabiat. 327 (6121): 437–9. doi:10.1038 / 327437a0. PMID 2953977.
- ^ Phong WY, Lin V, Rao SP, Dik T, Alonso S, Pethe K (Avgust 2013). Parish T (tahrir). "Mikobakteriya tuberkulyozida fosfofruktokinaza faolligini tavsiflash funktsional glyukolitik uglerod oqimi gipoksiya ostida toksik metabolik qidiruv moddalar to'planishini cheklash uchun zarurligini aniqladi". PLOS ONE. 8 (2): 1198–206. doi:10.1371 / journal.pone.0056037. PMC 3567006. PMID 23409118.
- ^ Papagianni M, Avramidis N (2012 yil may). "Lactococcus lactis markaziy yo'llarini muhandislik qilish: Lactococcus lactis tarkibidagi Aspergillus nigerdan olingan fosfofruktokinaza (pfk) va alternativ oksidaz (aox1) genlarining funktsional ekspressioni" oksidlanish sharoitida uglerod konversiyasining tezligini yaxshilaydi ". Ferment va mikroblar texnologiyasi. 51 (113): 125–30. doi:10.1016 / j.enzmictec.2012.04.007. PMID 22759530.
- ^ Lindinger, Maykl I.; Kovalchuk, Jon M.; Heigenhauser, George J. F. (2005-09-01). "Skelet mushaklari kislota-asos holatiga fizik-kimyoviy printsiplarni qo'llash". Amerika fiziologiya jurnali. Normativ, integral va qiyosiy fiziologiya. 289 (3): R891-R894. doi:10.1152 / ajpregu.00225.2005. ISSN 0363-6119. PMID 16105823.
- ^ Coelho WS, Sola-Penna M (Yanvar 2013). "Serotonin PLC-PKC-CaMK II- va Yanus kinazga bog'liq signalizatsiya yo'lida 6-fosfofructo-1-kinaz faolligini tartibga soladi". Mol. Hujayra. Biokimyo. 372 (1–2): 211–20. doi:10.1007 / s11010-012-1462-0. PMID 23010892.
- ^ Nakajima H, Raben N, Hamaguchi T, Yamasaki T (mart 2002). "Fosfofruktokinaza etishmovchiligi; o'tmishi, hozirgi va kelajak". Curr. Mol. Med. 2 (2): 197–212. doi:10.2174/1566524024605734. PMID 11949936.
- ^ Bruser A, KirchbergerJ, Schoneberg T (oktyabr 2012). "6-fosfofruktokinaz mushaklarining o'zgargan allosterik regulyatsiyasi Tarui kasalligini keltirib chiqaradi". Biokimyo Biofiz Res Commun. 427 (1): 133–7. doi:10.1016 / j.bbrc.2012.09.024. PMID 22995305.
- ^ Brüser A, Kirchberger J, Schöneberg T (oktyabr 2012). "6-fosfofruktokinaz mushaklarining o'zgargan allosterik regulyatsiyasi Tarui kasalligini keltirib chiqaradi". Biokimyo. Biofiz. Res. Kommunal. 427 (1): 133–7. doi:10.1016 / j.bbrc.2012.09.024. PMID 22995305.
- ^ Gomes LS, Zancan P, Marcondes MC, Ramos-Santos L, Meyer-Fernandes JR, Sola-Penna M, Da Silva D (Fevral 2013). "Resveratrol 6-fosfofructo-1-kinazni inhibe qilish orqali ko'krak bezi saratoni hujayralarining hayotiyligini va glyukoza metabolizmini pasaytiradi". Biochimie. 95 (6): 1336–43. doi:10.1016 / j.biochi.2013.02.013. PMID 23454376.
- ^ Vaz CV, Alves MG, Marques R, Moreira PI, Oliveira PF, Maia CJ, Socorro S (2013 yil fevral). "Androgenga javob beradigan va javob bermaydigan prostata saratoni hujayralari glikolitik metabolizmning aniq profilini namoyish etadi". Int J Biokimyoviy Hujayra Biol. 44 (11): 2077–84. doi:10.1016 / j.biocel.2012.08.013. PMID 22964025.
- ^ Yi V, Klark PM, Meyson DE, Keenan MC, Hill C, Goddard WA, Peters EC, Driggers EM, Hsieh-Wilson LC (Avgust 2012). "Fosfofruktokinaza 1 glikozilatsiyasi hujayralar o'sishi va metabolizmini tartibga soladi". Ilm-fan. 337 (6097): 975–80. doi:10.1126 / science.1222278. PMC 3534962. PMID 22923583.
- ^ Abrantes JL, Alves CM, Kosta J, Almeyda FK, Sola-Penna M, Fontes CF, Souza TM (Avgust 2012). "Herpes simplex 1-turi 6-fosfofructo-1-kinaza (PFK-1) fermenti bilan glikolizni faollashtiradi". Biochim Biofhys Acta. 1822 (8): 1198–206. doi:10.1016 / j.bbadis.2012.04.011. PMID 22542512.
Tashqi havolalar
- Fosfofruktokinaza-1 AQSh Milliy tibbiyot kutubxonasida Tibbiy mavzu sarlavhalari (MeSH)
- Proteopedia.org fosfofruktokinaz
- Da mavjud bo'lgan barcha tarkibiy ma'lumotlarga umumiy nuqtai PDB uchun UniProt: P08237 (ATPga bog'liq 6-fosfofruktokinaza, mushak turi) da PDBe-KB.
- Da mavjud bo'lgan barcha tarkibiy ma'lumotlarga umumiy nuqtai PDB uchun UniProt: Q01813 (ATP ga bog'liq 6-fosfofruktokinaza, trombotsit turi) PDBe-KB.
ATP ADP ATP ADP + + 2 ×  2 ×  2 × 3-fosfogliserat 2 ×  2 × 2-fosfogliserat 2 ×  2 × Fosfoenolpiruvat 2 ×  ADP ATP 2 × Piruvat 2 × |




