Foraminifera - Foraminifera
| Foraminifera | |
|---|---|
 | |
| Jonli Ammiak tepida (Rotaliida) | |
| Ilmiy tasnif | |
| Domen: | |
| (ochilmagan): | |
| (ochilmagan): | |
| Filum: | |
| Subfilum: | Foraminifera d'Orbigny, 1826 |
| Bo'limlar | |
"Monothalamea "
incertae sedis | |
Foraminifera (/fəˌræməˈnɪfərə/; Lotin "teshik tashuvchilar" uchun; norasmiy deb nomlangan "foramlar") bir hujayrali organizmlar, a a'zolari filum yoki sinf ning ameboid protistlar oqimli donadorligi bilan ajralib turadi ektoplazma oziq-ovqat va boshqa maqsadlarda foydalanish uchun; va odatda tashqi qobiq ("sinov ") turli xil shakllar va materiallar. Sinovlar xitin (ba'zi oddiy nasllarda uchraydi va Matnlar xususan) eng ibtidoiy turga ishoniladi. Foraminiferalarning aksariyati dengizdir, ularning aksariyati dengiz tubida yoki ular ichida yashaydi (ya'ni, mavjud) bentik ), kichikroq soni esa turli xil chuqurlikdagi suv ustunida suzadi (ya'ni, ular mavjud) planktonik ). Ularning ozgina qismi chuchuk suvdan yoki sho'rlangan sharoitdan ma'lum, va kichik subunitni molekulyar tahlil qilish orqali tuproqning juda oz (noakuatik) turlari aniqlangan. ribosomal DNK.[2][3]
Foraminifera odatda a hosil qiladi sinov Yoki bitta yoki bir nechta kameraga ega bo'lishi mumkin bo'lgan qobiq, ba'zilari esa tuzilishi jihatidan juda murakkab bo'lib qoladi.[4] Ushbu chig'anoqlar odatda yasalgan kaltsiy karbonat (CaCO
3) yoki aglutinatsiyalangan cho'kindi zarralari. Ikkala tirik (10000) 50 000 dan ortiq tur tan olinadi[5] va fotoalbom (40,000).[6][7] Ular odatda 1 mm dan kam o'lchamga ega, ammo ba'zilari juda katta eng katta turlar 20 sm gacha.[8]
Zamonaviy ilmiy ingliz tilida bu atama foraminifera ham birlik, ham ko'plik (so'zdan qat'iy nazar) Lotin derivation), va bir yoki bir nechta namunalarni yoki taksonlarni tavsiflash uchun ishlatiladi: uning birlik yoki ko'plik sifatida ishlatilishi kontekstdan aniqlanishi kerak. Guruhni tavsiflash uchun foraminifera tez-tez norasmiy ravishda qo'llaniladi va bu holatlarda odatda kichik harflar mavjud.[9]
O'qish tarixi
Foraminiferaga ma'lum bo'lgan eng qadimgi ma'lumotnoma kelib chiqadi Gerodot, miloddan avvalgi V asrda ularni toshni tashkil etuvchi deb ta'kidlagan Buyuk Giza piramidasi. Ular bugungi kunda nasl vakillari sifatida tan olingan Nummulitlar. Strabon, miloddan avvalgi 1-asrda xuddi shu foraminiferani ta'kidlab, ularning qoldiqlari deb taxmin qilgan yasmiq piramidalarni qurgan ishchilar qoldirgan.[10]
Robert Xuk uning 1665 yilgi kitobida tasvirlangan va tasvirlanganidek, mikroskop ostida foraminiferani kuzatgan Mikrografiya:
Men bir nechta kichkina va bitta kattalashtiruvchi ko'zoynakni sinab ko'rdim va beparvolik bilan oq qum uchastkasini ko'rayotgan edim, shunda donalardan birini aniq qobildoq va gulchambar kabi tasavvur qildim [...] Mikroskopni yaxshi tomonga burab, uni ikkala tomonida va tekis spirali Shell bilan kichik suvli salyangozning Shell-ga o'xshashligini [...][11]
Antoni van Leyvenxuk 1700 yilda tasvirlangan va tasvirlangan foraminiferal testlar, ularni minut kokillalar deb ta'riflagan; uning illyustratsiyasi borligi bilan tanilgan Elphidium.[12] Dastlabki ishchilar foraminiferalarni turkumga kiritdilar Nautilus, ularning aniq o'xshashligini ta'kidlab sefalopodlar. Bu tomonidan tan olingan Lorenz Spengler 1781 yilda ushbu foraminiferada septa teshiklari bor edi, bu esa oxir-oqibat guruhga o'z nomini beradi.[13] Shpengler, shuningdek, foraminiferaning septalari nautilidan farqli ravishda chiqishini va ularning asab naychasi yo'qligini ta'kidladi.[14]

Alcide d'Orbigny, 1826 yilgi ishida ularni bir daqiqali guruh deb hisoblagan sefalopodlar va ularning g'alati morfologiyasini ta'kidladilar, psevdopodiyani tentakllar deb talqin qildilar va juda qisqartirilgan (aslida mavjud bo'lmagan holda) boshni qayd etdilar.[15] U guruhga nom berdi foraminiferlaryoki "teshik ko'taruvchilar", chunki guruh a'zolari o'zlarining qobig'idagi bo'linmalar orasidagi bo'linmalarda farqli o'laroq nautili yoki ammonitlar.[9]
Foraminiferalarning protozoy tabiati birinchi marta tan olingan Dyujardin 1835 yilda.[13] Ko'p o'tmay, 1852 yilda d'Orbigny 72 ta nasli foraminiferani tan olgan holda tasniflash sxemasini ishlab chiqdi va u test shakliga qarab tasnifladi - bu hamkasblar tomonidan qattiq tanqidga uchragan sxema.[12]
X.B. Brady 1884 yilgi monografiyada foraminiferal topilmalar tasvirlangan CHellenjer ekspeditsiya. Brady stratigrafik diapazonga unchalik e'tibor bermasdan, 29 ta subfamilali 10 ta oilani tanidi; uning taksonomiyasi turli xil belgilar taksonomik guruhlarni ajratishi kerak degan fikrni ta'kidladi va shunga o'xshash holda aglutinatsiyalangan va ohakli nasllar o'zaro yaqin aloqada.
Tasniflashning ushbu umumiy sxemasi shu paytgacha saqlanib qoladi Kushmannikiga tegishli 1920 yillarning oxirlarida ishlash. Kushman devor kompozitsiyasini foraminiferalarni tasniflashdagi eng muhim xususiyat deb bilgan; uning tasnifi keng miqyosda qabul qilindi, ammo hamkasblarining "biologik jihatdan sog'lom emasligi" uchun tanqidlarga uchradi.
Kushmanning sxemasi, shunga qaramay, Tamin va Loeblichning 1964 yildagi tasnifiga qadar dominant sxemalar bo'lib qoldi, ular foraminiferalarni sinov devorining mikroyapısı asosida bugungi kunda ham qo'llanilgan umumiy guruhlarga kiritdilar.[12] Ushbu guruhlar yuqori darajadagi tasniflashning turli xil sxemalari bo'yicha turli xil harakatlarga uchragan. Pawlowski (2013) tomonidan molekulyar sistematikadan foydalanish umuman Tappan va Loeblich guruhlarini tasdiqladi, ba'zilari esa polifiletik yoki parafiletik deb topildi; bu ish, shuningdek, asosiy foraminiferal guruhlar o'rtasidagi yuqori darajadagi munosabatlarni aniqlashga yordam berdi.[16]
Taksonomiya
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Pawlowski va boshqalarni ta'qib qilgan Foraminifera filogeniyasi. 2013 yil.[16] Monotalamid buyuradi Astrorxizida va Allogromiida ikkalasi ham parafiletikdir. |
Foraminiferaning taksonomik pozitsiyasi 1854 yilda Shultzedan beri o'zgarib turadi,[17] Foraminiferida buyrug'i deb atagan. Loeblich va Tappan (1992) Foraminiferani sinf sifatida qayta ko'rib chiqdi[18] hozirda odatda ko'rib chiqilganidek.
Foraminiferalar odatda tarkibiga kiritilgan Protozoa,[19][20][21] yoki shunga o'xshash Protoctista-da yoki Protist qirollik.[22][23] Birinchi navbatda asoslangan ishonchli dalillar molekulyar filogenetik, protozoa tarkibidagi asosiy guruhga mansubligi uchun mavjud Rizariya.[19] Rizariya a'zolari o'rtasidagi evolyutsion aloqalar tan olinishidan oldin, Foraminiferalar odatda boshqalari bilan birlashtirilgan amoeboids Granuloreticulosa sinfidagi fil Rhizopodea (yoki Sarcodina) sifatida.
Rizariya muammoli, chunki ular odatdagidan foydalanmasdan, ko'pincha "supergrup" deb nomlanadi taksonomik daraja kabi filum. Kavalier-Smit Rizariyani Protozoa qirolligi tarkibidagi infra-qirollik deb ta'riflaydi.[19]
Ba'zi taksonomiyalar Foraminiferani o'zlari joylashtirgan amoeboid Sarkodinaga tenglashtirgan holda o'zlarining filumiga qo'ydilar.
Morfologik o'zaro bog'liqlik hali qo'llab-quvvatlanmagan bo'lsa-da, molekulyar ma'lumotlar Foraminiferalar bilan chambarchas bog'liqligini ko'rsatadi. Kerkozoa va Radiolariya, ikkalasiga ham murakkab qobiqlarga ega bo'lgan amoeboidlar kiradi; ushbu uchta guruh Rizariyani tashkil qiladi.[20] Biroq, foramlarning boshqa guruhlarga va bir-biriga bo'lgan aniq munosabatlari hali ham to'liq aniq emas. Foraminifera bilan chambarchas bog'liq amatea.[24]
| Mixalevich 2013 yildan taksonomiya[25] |
|---|
* Foraminifera d'Orbigny 1826 yil
|
Anatomiya

Ko'pgina foraminiferalarning eng diqqatga sazovor tomonlari bu qattiq qobiqlar yoki sinovlar. Ular bir nechta kameralardan iborat bo'lishi mumkin va oqsil, cho'kindi zarralari, kalsit, aragonit yoki (bir holda) kremniydan iborat bo'lishi mumkin.[18] Ba'zi foraminiferalarda testlar umuman yo'q.[26] Kabi boshqa qobiqni ajratuvchi organizmlardan farqli o'laroq mollyuskalar yoki mercanlar, foraminiferaning sinovlari ichkarida joylashgan hujayra membranasi ichida protoplazma. Hujayraning organoidlari sinov bo'limi (qismlari) ichida joylashgan bo'lib, sinov teshigi (teshiklari) materialni psevdopodiyadan ichki hujayraga va orqaga o'tkazishga imkon beradi.[27]
Foraminiferal hujayra donador endoplazma va shaffof ektoplazmaga bo'linadi, ulardan a psevdopodial to'r bitta teshikdan yoki sinovdagi ko'plab teshiklardan paydo bo'lishi mumkin. Shaxsiy psevdopodlar xarakterli ravishda ikkala yo'nalishda oqadigan kichik donachalarga ega.[28] Foraminiferalar noyobdir granuloreticulose pseudopodia; ya'ni ularning psevdopodiyasi mikroskop ostida donador bo'lib ko'rinadi; bu psevdopodiya ko'pincha cho'zilib ketadi va bo'linib, bir-biriga qo'shilib ketishi mumkin. Ularni hujayraning ehtiyojlariga qarab kengaytirish va tortib olish mumkin. Pseudopodlar harakatlanish, langar, ekskretsiya, sinov konstruktsiyasi va diatomalar yoki bakteriyalar kabi kichik organizmlardan tashkil topgan oziq-ovqat mahsulotlarini olish uchun ishlatiladi.[29][27]
Sinovlardan tashqari, foraminiferal hujayralar a tomonidan qo'llab-quvvatlanadi sitoskelet mikrotubulalardan iborat bo'lib, ular boshqa amoeboidlarda ko'rinadigan tuzilmasdan erkin joylashtirilgan. Mikrotubulalarni tezda yig'ish va qismlarga ajratish uchun maxsus uyali mexanizmlar rivojlanib, cho'zilgan psevdopodiyaning tez shakllanishi va orqaga tortilishiga imkon beradi.[18]
Gamamontda (jinsiy shaklda) foraminiferalar umuman bitta yadroga ega, agamont (jinssiz shakl) esa ko'p yadroga ega. Hech bo'lmaganda ba'zi turlarda yadrolar dimorfik bo'lib, somatik yadrolarda hosil qiluvchi yadrolarga qaraganda uch barobar ko'proq protein va RNK mavjud. Biroq, yadro anatomiyasi juda xilma-xil ko'rinadi.[30] Ko'p kamerali turlarda yadrolar bitta xonada cheklanishi shart emas. Yadrolar sferik bo'lishi mumkin yoki ko'plab loblarga ega. Odatda yadrolarning diametri 30-50 mm.[31]
Ba'zi foraminiferalarning hujayralarida katta, bo'sh vakuolalar mavjud; ularning aniq maqsadi noma'lum, ammo ular nitrat ombori sifatida ishlashi tavsiya etilgan.[31]
Mitoxondriyalar hujayra bo'ylab bir tekis tarqaladi, ammo ba'zi turlarda ular hujayraning teshiklari va tashqi chekkalari atrofida to'plangan. Bu past kislorodli muhitga moslashish deb faraz qilingan. [31]
Bir nechta turlari ksenofofor kontsentratsiyasining juda yuqori ekanligi aniqlandi radioaktiv izotoplar ularning hujayralarida, har qanday ökaryotning eng yuqori darajasi orasida. Buning maqsadi noma'lum.[32]
Ekologiya
Zamonaviy Foraminifera asosan dengiz organizmidir, ammo tirik odamlar sho'r, chuchuk suvda topilgan[28] va hatto quruqlikdagi yashash joylari.[3] Turlarning aksariyati bentik va yana 40 ta morfospesiya mavjud planktonik.[29] Ammo bu son haqiqiy xilma-xillikning faqat bir qismini aks ettirishi mumkin, chunki genetik jihatdan ajralib turadigan ko'plab turlar morfologik jihatdan ajralib turishi mumkin.[33]
Bentik foraminiferalar odatda mayda donali cho'kindilarda uchraydi, ular qatlamlar orasida faol ravishda harakatlanadi; ammo, ko'plab turlar qattiq tosh substratlarida, dengiz o'tlariga bog'langan yoki cho'kindi yuzasida o'tirgan holda uchraydi.[18]
Planktonik foraminiferalarning ko'p qismi globigerininada, rotaliida ichidagi nasabda uchraydi.[16] Biroq, kamida bitta boshqa rotaliid nasl, Neogallitelliya, mustaqil ravishda planktonik turmush tarzini rivojlantirganga o'xshaydi.[34][35] Bundan tashqari, ba'zi bir yura davri fotoalbomlari mustaqil ravishda planktonik turmush tarzini rivojlantirgan va Robertinidaning a'zolari bo'lishi mumkinligi taxmin qilinmoqda.[36]
Bir qator foramalar bir hujayrali suv o'tlari kabi endosimbionts kabi turli xil nasablardan yashil suv o'tlari, qizil suv o'tlari, oltin suv o'tlari, diatomlar va dinoflagellatlar.[29] Bular mikotrofik foraminiferlar, ayniqsa, ozuqaviy moddalarga kam bo'lgan okean suvlarida keng tarqalgan.[37] Ba'zi formalar kleptoplastik, saqlash xloroplastlar yutilgan suv o'tlaridan o'tkazish uchun fotosintez.[38]
Ko'pchilik foraminiferalar geterotrof bo'lib, mayda organizmlar va organik moddalarni iste'mol qiladi; ba'zi kichik turlar ixtisoslashgan oziqlantiruvchi vositalardir fitodetrit, boshqalari diatomlarni iste'mol qilishga ixtisoslashgan. Ba'zi bentik teshiklar psevodopodiya yordamida cho'kindi va organik zarralar ichida o'zlarini kistlash uchun ishlatiladi.[18] Ba'zi foraminiferalar kabi kichik hayvonlarni o'lja kopepodlar yoki kumushlar; ba'zi teshiklar hatto boshqa teshiklardan ham oldinroq bo'lib, o'ljalarining sinovlarida teshik ochishadi.[39] Ksenofoforlarning bir guruhi o'zlarining sinovlari davomida bakteriyalarni etishtirishni taklif qilishdi.[40] To'xtatib ovqatlantirish guruhda ham keng tarqalgan va hech bo'lmaganda ba'zi turlardan foydalanish mumkin erigan organik uglerod.[18]
Bir nechta foram turlari mavjud parazit, gubkalarni, mollyuskalarni, mercanlarni yoki hatto boshqa foraminiferalarni yuqtirish. Parazitar strategiyalar turlicha; ba'zilari ektoparazit bo'lib, xostdan oziq-ovqat o'g'irlash uchun o'zlarining psevdopodiyalaridan foydalanadilar, boshqalari esa xost egasining qobig'i yoki tanasi devori orqali yorilib, uning yumshoq to'qimalari bilan oziqlanadi.[18]
Foraminiferalarning o'zi ko'plab yirik organizmlar, shu jumladan umurtqasizlar, baliqlar, qirg'oq qushlari va boshqa foraminiferalar tomonidan iste'mol qilinadi. Ammo ba'zi hollarda yirtqichlar organizmlarning o'ziga emas, balki teshik qobig'idagi kaltsiyga ko'proq qiziqish bildirishi mumkin. Bir nechta suv salyangoz turlari tanlangan foraminiferalar bilan oziqlanadi va ko'pincha alohida turlarni afzal ko'rishadi.[41]
Ba'zi bentik foraminiferalar tirik qolish qobiliyatiga ega ekanligi aniqlandi anoksik 24 soatdan ortiq shartlar, bu ularning tanlov qobiliyatiga ega ekanligini ko'rsatadi anaerob nafas olish. Bu cho'kindi-suv chegarasi yaqinidagi o'zgaruvchan kislorodli sharoitlarda omon qolish uchun moslashish sifatida talqin etiladi.[42]
Foraminiferalar okeanning kabi eng chuqur qismlarida uchraydi Mariana xandagi shu jumladan Challenger chuqurligi, ma'lum bo'lgan eng chuqur qismi. Ushbu chuqurlikda, ostida karbonat kompensatsiyasi chuqurligi, sinovlarning kaltsiy karbonati haddan tashqari bosim tufayli suvda eriydi. Challenger chuqurida topilgan Foraminiferalarda karbonat sinovi yo'q, aksincha uning tarkibida organik moddalar mavjud.[43]
Ko'paytirish
Umumiy foraminiferal hayot tsikli o'zgarishni o'z ichiga oladi gaploid va diploid avlodlar, garchi ular asosan shakli jihatidan o'xshash bo'lsa.[17][44] Gaploid yoki gamont dastlab bitta yadro, va ko'plab ishlab chiqarish uchun bo'linadi jinsiy hujayralar, odatda ikkitadan flagella. Diploid yoki agamont ko'p yadroli va undan keyin mayoz yangi gamonts ishlab chiqarish uchun bo'linadi. Bir necha tur jinssiz ko'payish jinsiy avlodlar o'rtasida bentik shakllarda kam uchraydigan narsa emas.[28]

Foraminiferalar ularning ko'payish tsikli bilan bog'liq morfologik dimorfizmni namoyish etadi. Gamont, yoki jinsiy yo'l bilan ko'payadigan gaploid shakli megalosfera- bu uning prolakula, yoki birinchi kamera, mutanosib ravishda katta. Gamont shuningdek A shakl. Gamonts, odatda kattaroq prolokulyatsiyaga ega bo'lishiga qaramay, odatda umumiy sinov diametri agamontlarga qaraganda kichikroq bo'ladi.
Voyaga etganidan keyin gamont bo'linadi mitoz shuningdek, gaploid bo'lgan minglab jinsiy hujayralarni ishlab chiqarish. Ushbu gametalarning barchasi to'liq to'plamga ega organoidlar, va sinovdan zarar ko'rmagan holda atrofdan chiqarib yuboriladi. Gametalar farqlanmaydi sperma va tuxum va turlardan har qanday ikkita jinsiy hujayralar umuman bir-birini urug'lantirishi mumkin.

Ikkita jinsiy hujayralar birlashganda diploid, ko'p yadroli hujayra hosil bo'ladi agamont, yoki B shakl. Gamontdan farqli o'laroq, agamont mikrosferik, mutanosib ravishda kichik birinchi xonaga ega, lekin odatda ko'proq diametrli kameralar ko'proq umumiy diametrga ega. Agamont bu jinssiz ko'payish foraminiferaning fazasi; voyaga etganidan so'ng protoplazma testni butunlay bo'shatadi va uni ajratadi sitoplazma meiotik jihatdan orqali ko'p bo'linish bir qator gaploid naslni hosil qilish uchun. Keyin bu nasllar tarqalishdan oldin megalosferadagi birinchi kamerani hosil qila boshlaydi.
Ba'zi hollarda gaploid yoshi megalosfera shaklida pishib, keyinchalik jinssiz ko'payib, boshqa megalosfera, gaploid nasl hosil qiladi. Bunday holda, birinchi megalosfera shakli shizont yoki A1 shakli, ikkinchisi esa gamont yoki deb nomlanadi A2 shakl.

Kamolot va ko'payish sovuqroq va chuqurroq suvda sekinroq sodir bo'ladi; bu holatlar ham teshiklarning kattalashishiga olib keladi. A shakllari har doimgidan ancha ko'p bo'lib tuyuladi B shakllari, ehtimol ikkita gametaning bir-biriga duch kelishi va muvaffaqiyatli birlashishi ehtimoli kamayganligi sababli.[45][27]
Reproduktiv rejimdagi o'zgarishlar
Turli xil foraminiferal guruhlarda reproduktiv strategiyalarda yuqori xilma-xillik mavjud.
Yilda bir ko'zli turlar, A shakli va B shakl hali ham mavjud. Ko'p qirrali teshiklarning mikrosferik morfida bo'lgani kabi, jinssiz ko'payish B shakli jinsiy ko'payishdan kattaroqdir A shakl.
Oiladagi guruhlar Spirillinidae flagellated o'rniga amoeboid gametlarga ega. Ushbu guruhdagi ko'payishning boshqa jihatlari, odatda, boshqa teshik guruhlariga o'xshashdir.
Ohakli spirillinid Patellina gofrirovkasi ko'pgina boshqa foraminiferalarga qaraganda biroz boshqacha reproduktiv strategiyaga ega. Jinssiz ko'payish B formada butun hujayrani o'rab turgan kist hosil bo'ladi; keyinchalik u bu kist ichida bo'linadi va yosh hujayralar ota-ona testining kalsitini kannibalizatsiya qilib, o'zlarining dastlabki sinov xonasini hosil qiladi. Bular A voyaga etganidan so'ng shakllar to'qqiz kishigacha bo'lgan guruhlarga to'planadi; keyinchalik ular butun guruh atrofida himoya kistasini hosil qiladi. Gametogenez juda kam miqdordagi jinsiy hujayralarni ishlab chiqaradigan ushbu kist ichida paydo bo'ladi. The B kist ichida shakl lichinkalari hosil bo'ladi; hujayralarga bog'lanmagan har qanday yadrolar rivojlanayotgan lichinkalar uchun oziq-ovqat sifatida iste'mol qilinadi. Patellina yilda A xabarlarga ko'ra shakl ikki qavatli, "ortiqcha" va "minus" deb nomlangan jinslar bilan; bu jinslar yadro soni bo'yicha farq qiladi, "ortiqcha" shakli uchta yadroga, "minus" shakli esa to'rt yadroga ega. The B shakli yana kattaroq A shakl.[27][45][39]
Sinovlar

Foraminiferal testlar ichida organizmni himoya qilishga xizmat qiladi. Odatda, qattiq va bardoshli tuzilishi tufayli (boshqa protistlarga nisbatan), foraminiferalarning sinovlari guruh haqidagi ilmiy bilimlarning asosiy manbai hisoblanadi.
Sitoplazmaning tashqariga cho'zilishini ta'minlaydigan testdagi teshiklar teshik deb ataladi.[46] The birlamchi tashqi tomonga olib boruvchi diafragma, har xil turlarda juda ko'p turli xil shakllarga ega, shu jumladan ular bilan chegaralanmagan, dumaloq, yarim oy shaklidagi, yoriq shaklida, kaputli, nurli (yulduzcha shaklidagi), dendritik (tarvaqaylab ketgan). Ba'zi foraminiferalarda "tishli", gardishli yoki labda qilingan asosiy teshiklar mavjud. Faqat bitta asosiy diafragma yoki bir nechta bo'lishi mumkin; ko'plik mavjud bo'lganda, ular klasterli yoki ekvatorial bo'lishi mumkin. Birlamchi teshikdan tashqari, ko'plab foraminiferalar mavjud qo'shimcha teshiklar. Ular relikt teshiklari (oldingi o'sish bosqichidagi o'tgan asosiy teshiklar) yoki noyob tuzilmalar shaklida bo'lishi mumkin.
Sinov shakli turli xil foraminiferalar orasida juda o'zgaruvchan; ular bitta kamerali (bir ko'zli) yoki ko'p kamerali (ko'p ko'zli) bo'lishi mumkin. Ko'p qirrali shakllarda organizm o'sishi bilan yangi kameralar qo'shiladi. Sinov morfologiyalarining xilma-xilligi bir tomonlama va ko'p qirrali shakllarda, shu jumladan spiralli, seriyali va miliolinli va boshqalarda uchraydi.[27]
Ko'pgina foraminiferalar megalosfera va mikrosferik shaxslar ishtirokida o'zlarining sinovlarida dimorfizmni namoyon qiladilar. Ushbu nomlar to'liq organizmning kattaligi haqida gap ketmasligi kerak; aksincha, ular birinchi kameraning o'lchamiga ishora qiladilar yoki prolakula.

Qoldiqlar singari sinovlar qadimgi davrlardan ma'lum Ediakaran davr,[47] va ko'plab dengiz cho'kindi jinslari asosan ulardan iborat. Masalan, Misr piramidalarini tashkil etuvchi ohaktosh deyarli butunlay tashkil topgan nummulitik bentik Foraminifera.[48] Taxminlarga ko'ra Foraminifera rifi yiliga 43 million tonna kaltsiy karbonat ishlab chiqaradi.[49]
Genetik tadqiqotlar yalang'och amyobani aniqladi Retikulomiksa va o'ziga xos ksenofoforlar testlarsiz foraminiferanlar kabi. Yana bir nechta amoeboidlar retikuloza psevdopodlarini ishlab chiqaradi va ilgari ular Granuloreticulosa deb nomlangan, ammo bu endi tabiiy guruh deb hisoblanmaydi va aksariyati endi Cercozoa orasida joylashgan.[50]
Sinov tarkibi
Ularning shakli va tarkibi testlar foramalarni aniqlash va tasniflashning asosiy vositasi. Ko'pchilik ohakli testlarni o'z ichiga oladi kaltsiy karbonat.[28] Kaltsiyali testlar ikkalasidan ham tuzilishi mumkin aragonit yoki kaltsit turlarga qarab; kaltsit sinovlari bo'lganlar orasida test tarkibida magnezium o'rnini bosuvchi yuqori yoki past fraktsiya bo'lishi mumkin.[12] Sinov organik matritsani o'z ichiga oladi, uni ba'zan fotoalbom namunalaridan tiklash mumkin.[12]
Ba'zi tadkikotlar yuqori miqdorni taklif qiladi gomoplaziya foraminiferalarda va na aglutinatsiyalangan, na ohakli foraminiferalar monofil guruhlarni hosil qiladi.[16]
Yumshoq testlar
Ba'zi teshiklarda testlar organik moddalardan, odatda oqsillardan iborat bo'lishi mumkin tektin. Tektin devorlarida cho'kindi zarralari yuzaga erkin yopishgan bo'lishi mumkin.[27] Foram Retikulomiksa butunlay membrana hujayra devoriga ega bo'lgan sinovdan mahrum.[26] Organik devorli teshiklar an'anaviy ravishda "allogromidlar" guruhiga kiritilgan; ammo, genetik tadqiqotlar shuni aniqladiki, bu tabiiy guruhni tashkil qilmaydi.[16]
Aglutinatsiyalangan testlar
Boshqa teshiklarda oqsillar (kollagen bilan bog'liq bo'lishi mumkin), kaltsiy karbonat yoki Temir (III) oksidi.[27][51] Ilgari ushbu shakllar bir xonali "astrorhizidlar" va ko'p kamerali tekstulariidlar sifatida birlashtirilgan. Ammo yaqinda o'tkazilgan genetik tadqiqotlar shuni ko'rsatadiki, "astrorhizidlar" tabiiy guruhlashni tashkil etmaydi, aksincha daraxt daraxtining keng asosini tashkil qiladi.[16]
Textulariid foraminifera, globothalameaning boshqa tirik a'zolaridan farqli o'laroq, aglyutinatsiyalangan testlarga ega; ammo, ushbu sinovlardagi donalar kalsit tsement bilan sementlanadi. Ushbu kalsit tsement kichik (<100nm) globusli nanogrenalardan tashkil topgan, boshqa globotalamenlarga o'xshaydi. Ushbu testlarda ko'plab teshiklar bo'lishi mumkin, yana bir xususiyati ularni globotalameya bilan birlashtiradi.[36]

Aglutinatsiya qiluvchi foraminiferalar qanday zarrachalarni o'zlarining qobig'iga qo'shib olishlariga qarab tanlab olinishi mumkin. Ba'zi turlar tosh zarralarining ma'lum o'lchamlari va turlarini afzal ko'rishadi; boshqa turlari ma'lum biologik materiallarga nisbatan afzalroqdir. Ma'lumki, foraminiferalarning ayrim turlari afzalroq aglyutinatsiyaga uchragan koksolitlar testlarini shakllantirish; boshqalar imtiyozli ravishda foydalanadi echinoderm plitalar, diatomlar, yoki hatto boshqa foraminiferans testlari.[52]
Foraminiferalar Spikulosifon imtiyozli ravishda silika kremini aglutinatlaydi shimgichni spikulalar organik tsementdan foydalanish; u shaklga nisbatan kuchli selektivlikni namoyish etadi, "sopi" ustidagi cho'zilgan va "lampochkasida" qisqartirilgan spikulalardan foydalanadi. Spikulalarni dengiz tubidan ko'tarish vositasi sifatida, shuningdek uning cho'ziluvchanligini cho'zish uchun foydalanadi deb o'ylashadi. psevdopodiya o'ljani qo'lga olish.[51]
Ning aglutinatsiyalangan sinovlari ksenofoforlar diametri 20 sm gacha bo'lgan barcha foraminiferalarning eng kattasi. "Ksenofofor" nomi, "begona jismlarning tashuvchisi" degan ma'noni anglatadi, bu aglutinatsiya qiluvchi odatni anglatadi. Ksenofyoforlar tanadan 63 dan 500 um gacha bo'lgan cho'kindi donalarni oladilar, katta toshlar va mayda loylardan saqlanishadi; cho'kindi turi zarralar aglyutinatsiyalanadigan kuchli omil bo'lib tuyuladi, chunki zarrachalar tarkibiga sulfidlar, oksidlar, vulkanik oynalar va ayniqsa kichikroq foraminiferalarning sinovlari kiradi. Diametri 1,5 sm bo'lgan ksenofiyoforlar butunlay yalang'och holda qayd etilgan va hech qanday sinov o'tkazilmagan.[53]

Kaltsiyli sinovlar
Ohakli sinovlar o'tkazadigan foraminiferalardan kaltsit kristallarining turli xil tuzilmalari topilgan.
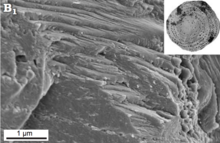
Chinni devorlar Miliolida. Ular buyurtma qilingan tashqi va ichki kalsit qoplamalari bilan tashkil etilgan yuqori magniyli kalsitdan iborat ("ekstraditsiya"va"intradoslar"navbati bilan) va qalin markaziy qatlamni tashkil etuvchi tasodifiy yo'naltirilgan igna shaklidagi kalsit kristallari (" chinni "). Organik ichki qoplama ham mavjud. Tashqi yuzasi chuqurlashtirilgan tuzilishga ega bo'lishi mumkin, ammo u teshiklar bilan teshilmagan." Cornuspirid "miliolidlarida ekstradrosiya etishmasligi aniq.[54][55][36]An'anaviy ravishda "monokristalli" sinov tuzilishi ta'riflangan Spirillinida. Biroq, ushbu testlar yomon tushunilgan va yomon tavsiflangan bo'lib qolmoqda. Ba'zi "monokristalli" spirillinidlar skanerlash elektron mikroskopi bilan kuzatilganda juda kichik kristallardan iborat mozaikadan tashkil topgan sinovlarga ega ekanligi aniqlandi. SEMni kuzatish Patellina sp. chinakam monokristalli sinov, albatta, yuzlari aniq ko'rinishda bo'lishi mumkinligini ko'rsatadi.[36]

Lagenid sinovlar o'nlab mikrometrga etadigan "tola to'plamlaridan" iborat; har bir "to'plam" bitta kalsit kristalidan hosil bo'ladi, kesmasi uchburchak va markazida teshik bor (sinov cho'kmasi artefakti deb o'ylaydi). Bundan tashqari, tolalar to'plamlarining "konus" tuzilishiga biriktirilgan ichki organik qatlam mavjud. Kristalli tuzilish boshqa ohakli foraminiferalarnikidan sezilarli darajada farq qilganligi sababli, u ohakli sinovning alohida evolyutsiyasini ifodalaydi. Lagenidlarning aniq mineralizatsiya jarayoni noaniq bo'lib qolmoqda.[55]

Rotaliid testlar "gialin" deb ta'riflanadi. Ular o'zlari bilan joylashtirilgan pastdan yuqori magnezium kalsitli "nanogrenlardan" hosil bo'ladi C o'qlari sinovning tashqi yuzasiga perpendikulyar. Bundan tashqari, ushbu nanogrenlar qatorlar, ustunlar yoki to'plamlar kabi yuqori darajadagi tuzilishga ega bo'lishi mumkin.[36] Sinov devori xarakterli ravishda ikki qavatli (ikki qavatli) va kichik teshikchalar bilan teshilgan. Sinov devorining tashqi kalsit qatlami "tashqi lamina" deb nomlanadi, ichki kalsit qatlami esa "ichki qoplama" deb nomlanadi; buni sinov ostidagi organik ichki qoplama bilan aralashtirib yubormaslik kerak. Tashqi laminalar va ichki qoplamalar orasida sendvich bo'lib, ikkitasini ajratib turadigan oqsil qatlami bo'lgan "o'rtacha qatlam". O'rtacha qatlam juda o'zgaruvchan; turlarga qarab u aniq belgilangan bo'lishi mumkin, boshqalarda esa keskin chegaralanmagan. O'rta qatlam ichida ba'zi nasllarda cho'kindi zarralari bo'lishi mumkin.[27][56][55]

The Karterinidlar, shu jumladan nasl Karterina va Zaninettiya, sinovning noyob kristalli tuzilishiga ega bo'lib, ularning tasnifini uzoq vaqt murakkablashtirgan. Ushbu turdagi test organik matritsa bilan bog'langan va tarkibida organik moddalarning "qon tomirlari" bo'lgan past magniyli kaltsit spikulalaridan iborat; bu ba'zi tadqiqotchilarning testni aglyutinatsiya qilish kerak degan xulosaga kelishiga olib keldi. Biroq, hayot tadqiqotlari aglutinatsiyani topa olmadi va aslida bu jins cho'kindi zarralari to'planmaydigan sun'iy substratda topilgan.[57] 2014 yilda o'tkazilgan genetik tadqiqotlar natijasida karterinidlar Globothalamea tarkibidagi mustaqil nasl ekanligi aniqlandi va spikula shakllari izchil shakllanib turishi sababli spikulalar ajralib chiqishi g'oyasini qo'llab-quvvatladi. Karterina va Zaninettiya bir xil joydan yig'ilgan (ovoid in Karterina, to'rtburchaklar shaklida yumaloq Zaninettiya).[58]
Hozir yo'q bo'lib ketgan Fusulinidlar an'anaviy ravishda biron bir mikrogranulyar kristallarning sinovlari o'tkazilishida noyob yo'nalishlarga ega va deyarli tsementga ega bo'lmagan noyob hisoblanadi. Biroq, 2017 yilgi tadqiqotlar shuni ko'rsatdiki, taxmin qilingan mikrogranular tuzilish, aslida, qoldiqlarning diagenetik o'zgarishi natijasida yuzaga kelgan va uning o'rniga o'zgartirilmagan fusulinid testlari gialinli tuzilishga ega. Bu shundan dalolat beradiki, guruh Globothalamea.[59]
Robertinidlar bor aragonitik teshiklari bo'lgan sinovlar; bu rotaliidlarning sinovlariga o'xshaydi, chunki ular nanogrenalardan hosil bo'ladi, ammo ular tarkibida va yaxshi tashkil etilgan ustunli domenlarga ega. Dastlabki planktonik teshiklarda aragonitik sinovlar bo'lganligi sababli, bu Robertinida ichida yaqin qarindoshlar bo'lishdan ko'ra, planktonik turmush tarzining alohida evolyutsiyasini aks ettirishi mumkin. Globigerinliklar.[36]
Gialin aragonitik testlari ham mavjud Involutinida.[55]
Silika sinovlari
Bitta tur, Milamellus, opalindan tayyorlangan teshiksiz sinovga ega kremniy.[18] U shakli va tuzilishi bo'yicha odatdagi miliolidlarning sinovlariga o'xshaydi; sinov ichki va tashqi organik qatlamdan, shuningdek cho'zinchoq tayoqchalardan yasalgan o'rta silika qatlamidan iborat. Ushbu kremniy qatlami tashqi, o'rta va ichki subbirliklarga bo'linadi; tashqi va ichki bo'linmalarning har biri taxminan 0,2 mm qalinlikda va uzun o'qlari sinov yuzasiga parallel bo'lgan silika tayoqchalarining subparallel varaqlaridan iborat. O'rta kichik birlik taxminan 18 mm qalinlikda va ochiq maydonda organik tarkibiy qismi bo'lmagan silika tayoqchalarining uch o'lchovli panjarasidan iborat. Ultrastrukturaning miliolidlardan farqi shundaki, novdalar o'rtacha ikki baravar uzunroq va qalinligi ikki baravar ko'p, Milamellus ular qattiq emas, balki ichi bo'sh va, albatta, kalsitdan ko'ra silika sinoviga ega.[60]

Sinov devorining konstruktsiyasi
Yashirin test mavjud bo'lganda, foraminiferal testlarning devorlari ham bo'lishi mumkin nonelellar yoki lamellar.
Nonlamellar devorlari ba'zi foraminiferalarda uchraydi, masalan Karterinida, Spirillinida, va Miliolida. Ushbu shakllarda, yangi kameraning sekretsiyasi avvalgi xonalarga nisbatan boshqa cho'ktirish bilan bog'liq emas. Shunday qilib, sinovda kalsit qatlamlari bilan bog'liq qatlamlar mavjud emas.[56]
Qatlamli devorlari bo'lgan foraminiferalarda yangi kamerani yotqizish avval hosil bo'lgan kameralar ustiga qatlam yotqizilishi bilan birga keladi. This layer may cover all previous chambers, or it may cover only some of them. These layers are known as secondary lamellae.
Foraminifera with lamellar walls can be further broken down into those with monolamellar walls and those with bilamellar devorlar. Monolamellar foraminifera secrete test walls which consist of a single layer, while those of bilamellar foraminifera are double-layered with an organic "median layer", sometimes containing sediment particles. In the case of bilamellar foraminifera, the outer layer is referred to as the "outer lamella" whilst the inner layer is referred to as the "inner lining". Monolamellar forams include the Lagenida, while bilamellar forms include the Rotaliida (including the major planktonic subgroup, the Globigerinina ).[56]
Bilamellar test walls can be further divided into those with septal flaps (a layer of test wall covering the previously-secreted septum) and those lacking septal flaps. Septal flaps are not known to be present in any foraminifera other than those with bilamellar walls.
The presence of a septal flap is often, though not always, associated with the presence of an interlocular space. As the name suggests, this is a small space located between chambers; it may be open and form part of the outer surface of the test, or it may be enclosed to form a void. The layer enclosing the void is formed from different parts of the lamellae in different genera, suggesting an independent evolution of enclosed interlocular spaces in order to strengthen the test.[56]
Evolyutsion tarix
Molecular clocks indicate that the crown-group of foraminifera likely evolved during the Neoproterozoy, between 900 and 650 million years ago; this timing is consistent with Neoproterozoic fossils of the closely related filose amoebae. As fossils of foraminifera have not been found prior to the very end of the Ediakaran, it is likely that most of these Proterozoic forms did not have hard-shelled tests.[61][62]
Due to their non-mineralised tests, "allogromiids " have no fossil record.[61]

The mysterious vendozoans of the Ediacaran period have been suggested to represent fossil ksenofoforlar.[63] Biroq, kashfiyot diagenetically-altered C27 sterollar associated with the remains of Diksoniya cast doubt on this identification and suggest it may instead be an animal.[64] Other researchers have suggested that the elusive trace fossil Paleodictyon va uning qarindoshlar may represent a fossil xenophyophore[65] and noted the similarity of the extant xenophyophore Occultammina to the fossil;[66] however, modern examples of Paleodictyon have not been able to clear up the issue and the trace may alternately represent a burrow or a glass sponge.[67] Supporting this notion is the similar habitat of living xenophyophores to the inferred habitat of fossil graphoglyptids; however, the large size and regularity of many graphoglyptids as well as the apparent absence of xenophyae in their fossils casts doubt on the possibility.[66] As of 2017 no definite xenophyophore fossils have been found.[68]
Test-bearing foraminifera have an excellent fossil record throughout the Fenerozoy eon. The earliest known definite foraminifera appear in the fossil record towards the very end of the Ediacaran; these forms all have agglutinated tests and are unilocular. These include forms like Platisolenitlar va Spirosolenitlar.[69][47]
Single-chambered foraminifera continued to diversity throughout the Cambrian. Some commonly encountered forms include Ammodiskus, Glomospira, Psammosphera, va Turritellella; these species are all agglutinated. They make up part of the Ammodiscina, nasab spirillinids that still contains modern forms.[70][16] Later spirillinids would evolve multilocularity and calcitic tests, with the first such forms appearing during the Trias; the group saw little effects on diversity due to the K-Pg extinction.[71]
The earliest multi-chambered foraminifera are agglutinated species, and appear in the fossil record during the middle Kembriy davr. Due to their poor preservation they cannot be positively assigned to any major foram group.[70]

The earliest known calcareous-walled foraminifera are the Fusulinids, which appear in the fossil record during the Llandoverian epoch of the early Siluriya. The earliest of these were microscopic, planispirally coiled, and evolute; later forms evolved a diversity of shapes including lenticular, globular, and perhaps most famously, elongated rice-shaped forms. Later species of fusulinids grew to much larger size, with some forms reaching 5 cm in length; reportedly, some specimens reach up to 14 cm in length, making them among the largest foraminifera extant or extinct. Fusulinids are the earliest lineage of foraminifera thought to have evolved symbiosis with photosynthetic organisms. Fossils of fusulinids have been found on all continents except Antarktida; they reached their greatest diversity during the Visean davri Karbonli. The group then gradually declined in diversity until finally going extinct during the Permo-triasning yo'q bo'lib ketish hodisasi.[27][71][72]
Davomida Tournaisian epoch of the Carboniferous, Miliolid foraminifera first appeared in the fossil record, having diverged from the spirillinids ichida Tubothalamea. Miliolids suffered about 50% casualties during both the Permo-Triassic and K-Pg extinctions but survived to the present day. Some fossil miliolids reached up to 2 cm in diameter.[71]

Eng qadimgi Lagenid fossils appear during the Moskvalik epoch of the Carboniferous. Seeing little effect due to the Permo-Triassic or K-Pg extinctions, the group diversified through time. Secondarily unilocular taxa evolved during the Jurassic and Cretaceous.
Eng qadimgi Involutinid fossils appear during the Permian; the lineage diversified throughout the Mesozoic of Eurasia before apparently vanishing from the fossil record following the Cenomanian-Turonian Ocean Anoxic Event. The extant group planispirillinidae has been referred to the involutinida, but this remains the subject of debate.[73][71]
The Robertinida first appear in the fossil record during the Anisian epoch of the Triassic. The group remained at low diversity throughout its fossil history; all living representatives belong to the Robertinidae, which first appeared during the Paleotsen.[71]
The first definite Rotaliid fossils do not appear in the fossil record until the Pliensbaxian epoch of the Jurassic, following the Triassic-Jurassic event.[74] Diversity of the group remained low until the aftermath of the Cenomanian-Turonian event, after which the group saw a rapid diversification. Of this group, the planktonic Globigerinina —the first known group of planktonic forams—first appears in the aftermath of the Toarcian Turnover; the group saw heavy losses during both the K-Pg extinction and the Eocene-Oligocene extinction, but remains extant and diverse to this day.[71] An additional evolution of planktonic lifestyle occurred in the Miocene or Pliocene, when the rotaliid Neogallitellia independently evolved a planktonic lifestyle.[34][35]
Paleontological applications


Dying planktonic Foraminifera continuously rain down on the sea floor in vast numbers, their mineralized tests preserved as fossils in the accumulating cho'kindi. Beginning in the 1960s, and largely under the auspices of the Deep Sea Drilling, Ocean Drilling, and International Ocean Drilling Programmes, as well as for the purposes of oil exploration, advanced deep-sea drilling techniques have been bringing up sediment cores bearing Foraminifera fossils.[75] The effectively unlimited supply of these fossil tests and the relatively high-precision age-control models available for cores has produced an exceptionally high-quality planktonic Foraminifera fossil record dating back to the mid-Yura davri, and presents an unparalleled record for scientists testing and documenting the evolutionary process.[75] The exceptional quality of the fossil record has allowed an impressively detailed picture of species inter-relationships to be developed on the basis of fossils, in many cases subsequently validated independently through molecular genetic studies on extant specimens[76]
Because certain types of foraminifera are found only in certain environments, their fossils can be used to figure out the kind of environment under which ancient marine sediments were deposited; conditions such as salinity, depth, oxygenic conditions, and light conditions can be determined from the different habitat preferences of various species of forams. This allows workers to track changing climates and environmental conditions over time by aggregating information about the foraminifera present.[77]
In other cases, the relative proportion of planktonic to benthic foraminifera fossils found in a rock can be used as a proxy for the depth of a given locality when the rocks were being deposited.[78]
Foraminifera have significant application in the field of biostratigrafiya. Due to their small size and hard shells, foraminifera may be preserved in great abundance and with high quality of preservation; due to their complex morphology, individual species are easily recognizable. Foraminifera species in the fossil record have limited ranges between the species' first evolution and their disappearance; stratigraphers have worked out the successive changes in foram assemblages throughout much of the Fenerozoy. As such, the assemblage of foraminifera within a given locality can be analyzed and compared to known dates of appearance and disappearance in order to narrow down the age of the rocks. This allows paleontologists to interpret the age of cho'kindi jinslar qachon radiometrik tanishuv amal qilmaydi.[79] This application of foraminifera was discovered by Alva C. Ellisor 1920 yilda.[80]
Calcareous fossil foraminifera are formed from elements found in the ancient seas where they lived. Thus, they are very useful in paleoklimatologiya va paleoceanografiya. They can be used, as a climate ishonchli vakil, to reconstruct past climate by examining the barqaror izotop ratios and trace element content of the shells (tests). Global temperature and ice volume can be revealed by the isotopes of oxygen, and the history of the uglerod aylanishi and oceanic productivity by examining the stable isotope ratios of carbon;[81] qarang δ18O va -13C. The concentration of trace elements, like magniy (Mg),[82] lityum (Li)[83] va bor (B),[84] also hold a wealth of information about global temperature cycles, continental weathering, and the role of the ocean in the global carbon cycle. Geographic patterns seen in the fossil records of planktonic forams are also used to reconstruct ancient okean oqimlari.
Zamonaviy foydalanish
The neft sanoati relies heavily on mikrofosil such as forams to find potential hydrocarbon deposits.[85]

For the same reasons they make useful biostratigraphic markers, living foraminiferal assemblages have been used as bioindikatorlar in coastal environments, including indicators of coral reef health. Because calcium carbonate is susceptible to dissolution in acidic conditions, foraminifera may be particularly affected by changing climate and okeanning kislotaliligi.

Foraminifera have many uses in neftni qidirish and are used routinely to interpret the ages and paleoenvironments of sedimentary strata in oil wells.[86] Agglutinated fossil foraminifera buried deeply in sedimentary basins can be used to estimate thermal maturity, which is a key factor for petroleum generation. The Foraminiferal ranglanish ko'rsatkichi[87] (FCI) is used to quantify colour changes and estimate burial temperature. FCI data is particularly useful in the early stages of petroleum generation (about 100 °C).
Foraminifera can also be used in arxeologiya ichida provenancing of some stone raw material types. Some stone types, such as ohaktosh, are commonly found to contain fossilised foraminifera. The types and concentrations of these fossils within a sample of stone can be used to match that sample to a source known to contain the same "fossil signature".[88]
Galereya

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Indian Ocean, south-eastern coast of Bali, field width 5.5 mm

Foraminifera of Indian Ocean, south-eastern coast of Bali, field width 5.5 mm

Foraminifera of Indian Ocean, south-eastern coast of Bali, field width 5.5 mm

Foraminifera in Ngapali, Myanmar, field width 5.22 mm

Foraminifera Heterostegina depressa, field width 4.4 mm









Adabiyotlar
- ^ Parfrey, Laura Vogener; Lahr, Daniel J. G.; Knoll, Endryu X.; Katz, Laura A. (2011 yil 16-avgust). "Multigenali molekulyar soatlar bilan erta ökaryotik diversifikatsiya vaqtini baholash". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 108 (33): 13624–13629. Bibcode:2011PNAS..10813624P. doi:10.1073 / pnas.1110633108. PMC 3158185. PMID 21810989.
- ^ Giere, Olav (2009). Meiobenthology: the microscopic motile fauna of aquatic sediments (2-nashr). Berlin: Springer. ISBN 978-3540686576.
- ^ a b Lejzerowicz, Franck; Pawlowski, Jan; Fraissinet-Tachet, Laurence; Marmeisse, Roland (1 September 2010). "Molecular evidence for widespread occurrence of Foraminifera in soils". Atrof-muhit mikrobiologiyasi. 12 (9): 2518–26. doi:10.1111/j.1462-2920.2010.02225.x. PMID 20406290.
- ^ Kennett, J.P.; Srinivasan, M.S. (1983). Neogene planktonic foraminifera: a phylogenetic atlas. Hutchinson Ross. ISBN 978-0-87933-070-5.
- ^ Ald, S.M. va boshq. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ^ Pawlowski, J., Lejzerowicz, F., & Esling, P. (2014). Next-generation environmental diversity surveys of foraminifera: preparing the future. The Biological Bulletin, 227(2), 93-106.
- ^ "World Foraminifera Database".
- ^ Marshall M (3 February 2010). "Zoologger: 'Living beach ball' is giant single cell". Yangi olim.
- ^ a b Lipps JH, Finger KL, Walker SE (October 2011). "What Should We call the Foraminifera" (PDF). Journal of Foraminiferal Research. 41 (4): 309–313. doi:10.2113/gsjfr.41.4.309. Olingan 10 aprel 2018.
- ^ "Foraminifera | Fossil Focus | Time | Discovering Geology | British Geological Survey (BGS)". www.bgs.ac.uk. Olingan 20 iyul 2020.
- ^ "Micrographia, or, Some physiological descriptions of minute bodies made by magnifying glasses ?with observations and inquiries thereupon /by R. Hooke ... : Hooke, Robert, : Free Download, Borrow, and Streaming". Internet arxivi. Olingan 20 iyul 2020.
- ^ a b v d e Sen Gupta, Barun K. (2003), Sen Gupta, Barun K. (ed.), Modern Foraminifera, Springer Netherlands, pp. 7–36, doi:10.1007/0-306-48104-9_2, ISBN 978-0-306-48104-8 Yo'qolgan yoki bo'sh
sarlavha =(Yordam bering);| bob =mensimagan (Yordam bering) - ^ a b BOUDAGHER-FADEL, MARCELLE K. (2018), "Biology and Evolutionary History of Larger Benthic Foraminifera", Evolution and Geological Significance of Larger Benthic Foraminifera (2 ed.), UCL Press, pp. 1–44, ISBN 978-1-911576-94-5, JSTOR j.ctvqhsq3.3
- ^ Hansen, H. (1 January 1981). "On Lorentz Spengler and a neotype for the foraminifer Calcarina spengleri". Iqtibos jurnali talab qiladi
| jurnal =(Yordam bering) - ^ d'Orbigny, Alcide (1826). "Tableau Méthodique de la Classe des Céphalopodes". Annales des Sciences Naturelles, Paris (Série 1). 7: 245–314 – via Biodiversity Heritage Library.
- ^ a b v d e f g Pawlowski, Jan; Holzmann, Maria; Tyszka, Jarosław (1 April 2013). "New supraordinal classification of Foraminifera: Molecules meet morphology". Dengiz mikropaleontologiyasi. 100: 1–10. Bibcode:2013MarMP.100....1P. doi:10.1016/j.marmicro.2013.04.002. ISSN 0377-8398.
- ^ a b Loeblich Jr, A.R.; Tappan, H. (1964). "Foraminiferida". Part C, Protista 2. Umurtqasiz hayvonlar paleontologiyasi haqida risola. Amerika Geologik Jamiyati. pp. C55–C786. ISBN 978-0-8137-3003-5.
- ^ a b v d e f g h Sen Gupta, Barun K. (2002). Modern Foraminifera. Springer. p. 16. ISBN 978-1-4020-0598-5.
- ^ a b v Cavalier-Smith, T (2004). "Hayotning oltita shohligi" (PDF). Ish yuritish. Biologiya fanlari. 271 (1545): 1251–62. doi:10.1098 / rspb.2004.2705. PMC 1691724. PMID 15306349.
- ^ a b Cavalier-Smith, T (2003). "Protist phylogeny and the high-level classification of Protozoa". Evropa Protistologiya jurnali. 34 (4): 338–348. doi:10.1078/0932-4739-00002.
- ^ Tolweb Cercozoa
- ^ European Register of Marine Species
- ^ eForams-taxonomy Arxivlandi 2011 yil 3 oktyabr Orqaga qaytish mashinasi
- ^ Testate amoebae as environmental indicators (PDF), dan arxivlangan asl nusxasi (PDF) 2016 yil 27-noyabrda, olingan 27 noyabr 2016
- ^ Mikhalevich, V.I. (2013). "New insight into the systematics and evolution of the foraminifera". Mikropaleontologiya. 59 (6): 493–527.
- ^ a b Pawlowski, Jan; Bolivar, Ignacio; Fahrni, Jose F.; Vargas, Colomban De; Bowser, Samuel S. (1999). "Molecular Evidence That Reticulomyxa Filosa Is A Freshwater Naked Foraminifer". Eukaryotik mikrobiologiya jurnali. 46 (6): 612–617. doi:10.1111/j.1550-7408.1999.tb05137.x. ISSN 1550-7408. PMID 10568034. S2CID 36497475.
- ^ a b v d e f g h men Saraswati, Pratul Kumar; Srinivasan, M. S. (2016), Saraswati, Pratul Kumar; Srinivasan, M.S. (eds.), "Calcareous-Walled Microfossils", Micropaleontology: Principles and Applications, Springer International Publishing, pp. 81–119, doi:10.1007/978-3-319-14574-7_6, ISBN 978-3-319-14574-7
- ^ a b v d Sen Gupta, Barun K. (1982). "Ecology of benthic Foraminifera". In Broadhead, T.W. (tahrir). Foraminifera: notes for a short course organized by M.A. Buzas and B.K. Sen Gupta. Studies in Geology. 6. University of Tennessee, Dept. of Geological Sciences. 37-50 betlar. ISBN 978-0910249058. OCLC 9276403.
- ^ a b v Hemleben, C .; Anderson, O.R .; Spindler, M. (1989). Zamonaviy Planktonik Foraminifera. Springer-Verlag. ISBN 978-3-540-96815-3.
- ^ Grell, K. G. (1 January 1979). "Cytogenetic systems and evolution in foraminifera". The Journal of Foraminiferal Research. 9 (1): 1–13. doi:10.2113/gsjfr.9.1.1. ISSN 0096-1191.
- ^ a b v Lekieffre, Charlotte; Bernhard, Joan M.; Mabilleau, Guillaume; Filipsson, Helena L.; Meibom, Anders; Geslin, Emmanuelle (1 January 2018). "An overview of cellular ultrastructure in benthic foraminifera: New observations of rotalid species in the context of existing literature". Dengiz mikropaleontologiyasi. 138: 12–32. doi:10.1016/j.marmicro.2017.10.005. ISSN 0377-8398.
- ^ Domanov, M. M. (July 2015). "Natural 226Ra and 232Th radionuclides in xenophyophores of the Pacific Ocean". Xalqaro geokimyo. 53 (7): 664–669. doi:10.1134/S0016702915070034. ISSN 0016-7029. S2CID 127121951.
- ^ Kucera, M.; Darling, K.F. (2002 yil aprel). "Cryptic species of planktonic foraminifera: their effect on palaeoceanographic reconstructions". Philos Trans Royal Soc A. 360 (1793): 695–718. Bibcode:2002RSPTA.360..695K. doi:10.1098/rsta.2001.0962. PMID 12804300. S2CID 21279683.
- ^ a b Ujiié, Yurika; Kimoto, Katsunori; Pawlowski, Jan (December 2008). "Molecular evidence for an independent origin of modern triserial planktonic foraminifera from benthic ancestors". Dengiz mikropaleontologiyasi. 69 (3–4): 334–340. doi:10.1016/j.marmicro.2008.09.003.
- ^ a b Özdikmen, Hüseyin (June 2009). "Substitute names for some unicellular animal taxa (Protozoa" (PDF). Munis Entomology & Zoology. 4 (1): 233–256.
- ^ a b v d e f Dubicka, Zofia (2019). "Chamber arrangement versus wall structure in the high-rank phylogenetic classification of Foraminifera". Acta Palaeontologica Polonica. 64. doi:10.4202/app.00564.2018. ISSN 0567-7920.
- ^ Mikrobial ekologiyaning yutuqlari, 11-jild
- ^ Bernxard, J. M .; Bowser, S.M. (1999). "Dissoksik cho'kindilarning bentik foraminiferalari: xloroplast sekvestratsiyasi va funktsional morfologiyasi". Earth-Science sharhlari. 46 (1): 149–165. Bibcode:1999ESRv ... 46..149B. doi:10.1016 / S0012-8252 (99) 00017-3.
- ^ a b Goldstein, Susan T. (2003), Sen Gupta, Barun K. (ed.), Modern Foraminifera, Springer Netherlands, pp. 37–55, doi:10.1007/0-306-48104-9_3, ISBN 978-0-306-48104-8 Yo'qolgan yoki bo'sh
sarlavha =(Yordam bering);| bob =mensimagan (Yordam bering) - ^ Laureillard, J; Méjanelle, L; Sibuet, M (2004). "Use of lipids to study the trophic ecology of deep-sea xenophyophores". Dengiz ekologiyasi taraqqiyoti seriyasi. 270: 129–140. doi:10.3354/meps270129. ISSN 0171-8630.
- ^ Kalver, Stiven J.; Lipps, Jere H. (2003), Kelley, Patricia H.; Kowalewski, Michał; Hansen, Thor A. (eds.), "Predation on and by Foraminifera", Predator—Prey Interactions in the Fossil Record, Boston, MA: Springer US, pp. 7–32, doi:10.1007/978-1-4615-0161-9_2, ISBN 978-1-4613-4947-1, olingan 30 sentyabr 2020
- ^ Moodley, L.; Hess, C. (1 August 1992). "Tolerance of Infaunal Benthic Foraminifera for Low and High Oxygen Concentrations". Biologik byulleten. 183 (1): 94–98. doi:10.2307/1542410. ISSN 0006-3185. JSTOR 1542410. PMID 29304574.
- ^ Gooday, A.J.; Todo, Y.; Uematsu, K.; Kitazato, H. (July 2008). "New organic-walled Foraminifera (Protista) from the ocean's deepest point, the Challenger Deep (western Pacific Ocean)". Linnean Jamiyatining Zoologik jurnali. 153 (3): 399–423. doi:10.1111/j.1096-3642.2008.00393.x.
- ^ Moore, R.C.; Lalicker, A.G.; Fischer, C.G. (1952). "Ch 2 Foraminifera and Radiolaria". Umurtqasiz hayvonlar qoldiqlari. McGraw-Hill. OCLC 547380.
- ^ a b Haynes, J. R. (18 June 1981). Foraminifera. Springer. ISBN 978-1-349-05397-1.
- ^ Lana, C (2001). "Cretaceous Carterina (Foraminifera)". Dengiz mikropaleontologiyasi. 41 (1–2): 97–102. Bibcode:2001MarMP..41...97L. doi:10.1016/S0377-8398(00)00050-5.
- ^ a b Kontorovich, A. E .; Varlamov, A. I .; Grazhdankin, D. V.; Karlova, G. A .; Klets, A. G.; Kontorovich, V. A .; Saraev, S. V .; Terleev, A. A.; Belyaev, S. Yu .; Varaksina, I. V .; Efimov, A. S. (1 December 2008). "A section of Vendian in the east of West Siberian Plate (based on data from the Borehole Vostok 3)". Rus geologiyasi va geofizikasi. 49 (12): 932–939. Bibcode:2008RuGG ... 49..932K. doi:10.1016 / j.rgg.2008.06.012. ISSN 1068-7971.
- ^ Foraminifera: o'rganish tarixi, London universiteti kolleji, retrieved 20 September 2007
- ^ Langer, M. R.; Silk, M. T. B.; Lipps, J. H. (1997). "Global ocean carbonate and carbon dioxide production: The role of reef Foraminifera". Journal of Foraminiferal Research. 27 (4): 271–277. doi:10.2113/gsjfr.27.4.271.
- ^ Adl, S. M.; Simpson, A. G. B.; Farmer, M. A.; Anderson; va boshq. (2005). "The new higher level classification of Eukaryotes with emphasis on the taxonomy of Protists". Eukaryotik mikrobiologiya jurnali. 52 (5): 399–451. doi:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- ^ a b Maldonado, Manuel; López-Acosta, María; Sitjà, Cèlia; Aguilar, Ricardo; García, Silvia; Vacelet, Jean (10 June 2013). "A giant foraminifer that converges to the feeding strategy of carnivorous sponges: Spiculosiphon oceana sp. nov. (Foraminifera, Astrorhizida)". Zootaxa. 3669 (4): 571–584. doi:10.11646/zootaxa.3669.4.9. hdl:10261/92975. ISSN 1175-5334. PMID 26312358.
- ^ Thomsen, Erik; Rasmussen, Tine L. (1 July 2008). "Coccolith-Agglutinating Foraminifera from the Early Cretaceous and How They Constructed Their Tests". Journal of Foraminiferal Research. 38 (3): 193–214. doi:10.2113/gsjfr.38.3.193. ISSN 0096-1191.
- ^ Levin, Lisa A.; Thomas, Cynthia L. (1 December 1988). "The ecology of xenophyophores (Protista) on eastern Pacific seamounts". Chuqur dengiz tadqiqot qismi A. Okeanografik tadqiqotlar. 35 (12): 2003–2027. Bibcode:1988DSRA...35.2003L. doi:10.1016/0198-0149(88)90122-7. ISSN 0198-0149.
- ^ Jain, Sreepat (2020), Jain, Sreepat (ed.), "Benthic Foraminifera", Fundamentals of Invertebrate Palaeontology: Microfossils, Springer Geology, New Delhi: Springer India, pp. 171–192, doi:10.1007/978-81-322-3962-8_9, ISBN 978-81-322-3962-8
- ^ a b v d Dubicka, Zofia; Ovokki, Kshishtof; Gloc, Michał (1 April 2018). "Micro- and Nanostructures of Calcareous Foraminiferal Tests: Insight from Representatives of Miliolida, Rotaliida and Lagenida". Journal of Foraminiferal Research. 48 (2): 142–155. doi:10.2113/gsjfr.48.2.142. ISSN 0096-1191.
- ^ a b v d Hansen, Hans Jørgen (2003), Sen Gupta, Barun K. (ed.), "Shell construction in modern calcareous Foraminifera", Modern Foraminifera, Dordrecht: Springer Netherlands, pp. 57–70, doi:10.1007/0-306-48104-9_4, ISBN 978-0-306-48104-8
- ^ Machado, Altair; Barros, Facelucia (8 January 2013). "The occurrence of Carterina spiculotesta (Carter, 1877) on an artificial substrate". Tekshirish ro'yxati. 9 (4): 813–814. doi:10.15560/9.4.813. ISSN 1809-127X.
- ^ Pawlowski, Jan; Holzmann, Maria; Debenay, Jean-Pierre (1 October 2014). "Molecular Phylogeny of Carterina Spiculotesta and Related Species from New Caledonia". Journal of Foraminiferal Research. 44 (4): 440–450. doi:10.2113/gsjfr.44.4.440. ISSN 0096-1191.
- ^ Dubicka, Zofia; Gorzelak, Przemysław (9 November 2017). "Unlocking the biomineralization style and affinity of Paleozoic fusulinid foraminifera". Ilmiy ma'ruzalar. 7 (1): 15218. Bibcode:2017NatSR...715218D. doi:10.1038/s41598-017-15666-1. ISSN 2045-2322. PMC 5680253. PMID 29123221.
- ^ Resig, J; Lowenstam, H; Echols, R; Weiner, S (1980). "An extant opaline foraminifer: test ultrastructure, mineralogy, and taxonomy". Special Publications of the Cushman Foundation for Foraminiferal Research. 19: 205–214.
- ^ a b Pawlowski, Jan; Holzmann, Maria; Berney, Cédric; Fahrni, José; Xayr, Endryu J.; Cedhagen, Tomas; Habura, Andrea; Bowser, Samuel S. (30 September 2003). "The evolution of early Foraminifera". Milliy fanlar akademiyasi materiallari. 100 (20): 11494–11498. Bibcode:2003PNAS..10011494P. doi:10.1073/pnas.2035132100. ISSN 0027-8424. PMC 208786. PMID 14504394.
- ^ Groussin, Mathieu; Pawlowski, Jan; Yang, Ziheng (1 October 2011). "Bayesian relaxed clock estimation of divergence times in foraminifera". Molekulyar filogenetik va evolyutsiyasi. 61 (1): 157–166. doi:10.1016/j.ympev.2011.06.008. ISSN 1055-7903. PMID 21723398.
- ^ Seilacher, A. (1 January 2007). "The nature of vendobionts". Geologik Jamiyat, London, Maxsus nashrlar. 286 (1): 387–397. Bibcode:2007GSLSP.286..387S. doi:10.1144/SP286.28. ISSN 0305-8719. S2CID 128619251.
- ^ Bobrovskiy, Ilya; Hope, Janet M.; Ivantsov, Andrey; Nettersheim, Benjamin J.; Hallmann, Christian; Brocks, Jochen J. (21 September 2018). "Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals". Ilm-fan. 361 (6408): 1246–1249. Bibcode:2018Sci...361.1246B. doi:10.1126/science.aat7228. ISSN 0036-8075. PMID 30237355.
- ^ Swinbanks, D. D. (1 October 1982). "Piaeodicton: The Traces of Infaunal Xenophyophores?". Ilm-fan. 218 (4567): 47–49. Bibcode:1982Sci...218...47S. doi:10.1126/science.218.4567.47. ISSN 0036-8075. PMID 17776707. S2CID 28690086.
- ^ a b Levin, Lisa A. (1994). "Paleoecology and Ecology of Xenophyophores". PALAY. 9 (1): 32–41. Bibcode:1994Palai...9...32L. doi:10.2307/3515076. ISSN 0883-1351. JSTOR 3515076.
- ^ Rona, Peter A.; Seilacher, Adolf; de Vargas, Colomban; Xayr, Endryu J.; Bernhard, Joan M.; Bowser, Sam; Vetriani, Costantino; Wirsen, Carl O.; Mullineaux, Lauren; Sherrell, Robert; Frederick Grassle, J. (1 September 2009). "Paleodictyon nodosum: A living fossil on the deep-sea floor". Chuqur dengiz tadqiqotlari II qism: Okeanografiyaning dolzarb tadqiqotlari. Marine Benthic Ecology and Biodiversity: A Compilation of Recent Advances in Honor of J. Frederick Grassle. 56 (19): 1700–1712. Bibcode:2009DSRII..56.1700R. doi:10.1016/j.dsr2.2009.05.015. ISSN 0967-0645.
- ^ Gooday, Andrew J; Holzmann, Maria; Caulle, Clémence; Goineau, Aurélie; Kamenskaya, Olga; Weber, Alexandra A. -T.; Pawlowski, Jan (1 March 2017). "Giant protists (xenophyophores, Foraminifera) are exceptionally diverse in parts of the abyssal eastern Pacific licensed for polymetallic nodule exploration". Biologik konservatsiya. 207: 106–116. doi:10.1016/j.biocon.2017.01.006. ISSN 0006-3207.
- ^ McIlroy, Duncan; Green, O. R.; Brasier, M. D. (2001). "Palaeobiology and evolution of the earliest agglutinated Foraminifera: Platysolenites, Spirosolenites and related forms". Leteya. 34 (1): 13–29. doi:10.1080/002411601300068170. ISSN 1502-3931.
- ^ a b Scott, David B.; Medioli, Franco; Braund, Regan (1 June 2003). "Foraminifera from the Cambrian of Nova Scotia: The oldest multichambered foraminifera". Mikropaleontologiya. 49 (2): 109–126. doi:10.2113/49.2.109. ISSN 1937-2795.
- ^ a b v d e f Tappan, Helen; Loeblich, Alfred R. (1988). "Foraminiferal Evolution, Diversification, and Extinction". Paleontologiya jurnali. 62 (5): 695–714. ISSN 0022-3360. JSTOR 1305391.
- ^ "Fusulinids | GeoKansas". geokansas.ku.edu. Olingan 16 may 2020.
- ^ Czaplewski, John J. "PBDB Navigator". paleobiodb.org. Olingan 16 may 2020.
- ^ Gräfe, K.U. (2005). "Benthic foraminifers and palaeoenvironment in the Lower and Middle Jurassic of the Western Basque-Cantabrian Basin (Northern Spain)". Iberian Geology jurnali. 31 (2): 217–233. S2CID 55664447.
- ^ a b "Nature debates".
- ^ Journal bioinformatics and biology insights, Using the Multiple Analysis Approach to Reconstruct Phylogenetic Relationships among Planktonic Foraminifera from Highly Divergent and Length-polymorphic SSU rDNA Sequences
- ^ Gebhardt, Holger (1 February 1997). "Cenomanian to Turonian foraminifera from Ashaka (NE Nigeria): quantitative analysis and palaeoenvironmental interpretation". Bo'r davridagi tadqiqotlar. 18 (1): 17–36. doi:10.1006/cres.1996.0047. ISSN 0195-6671.
- ^ Báldi, Katalin; Benkovics, László; Sztanó, Orsolya (1 May 2002). "Badenian (Middle Miocene) basin development in SW Hungary: subsidence history based on quantitative paleobathymetry of foraminifera". Xalqaro Yer haqidagi jurnal. 91 (3): 490–504. Bibcode:2002IJEaS..91..490B. doi:10.1007/s005310100226. ISSN 1437-3262. S2CID 129296067.
- ^ Australia, c=AU;o=Australia Government;ou=Geoscience (15 May 2014). "Biostratigraphy". www.ga.gov.au. Olingan 20 iyul 2020.
- ^ Cushman, Joseph A.; Ellisor, Alva C. (1 January 1945). "The Foraminiferal Fauna of the Anahuac Formation". Paleontologiya jurnali. 19 (6): 545–572. JSTOR 1299203.
- ^ Zachos, J.C.; Pagani, M .; Sloan, L.; Tomas, E .; Billups, K. (2001). "Trends, Rhythms, and Aberrations in Global Climate, 65 Ma to Present". Ilm-fan. 292 (5517): 686–693. Bibcode:2001Sci...292..686Z. doi:10.1126/science.1059412. PMID 11326091. S2CID 2365991.
- ^ Branson, Oscar; Redfern, Simon A.T.; Tilishak, Tolek; Sadekov, Aleksey; Langer, Gerald; Kimoto, Katsunori; Elderfield, Henry (December 2013). "The coordination of Mg in foraminiferal calcite". Yer va sayyora fanlari xatlari. 383: 134–141. Bibcode:2013E&PSL.383..134B. doi:10.1016/j.epsl.2013.09.037.
- ^ Misra, S .; Froelich, P. N. (26 January 2012). "Lithium Isotope History of Cenozoic Seawater: Changes in Silicate Weathering and Reverse Weathering". Ilm-fan. 335 (6070): 818–823. Bibcode:2012Sci...335..818M. doi:10.1126/science.1214697. PMID 22282473. S2CID 42591236.
- ^ Hemming, N.G.; Hanson, G.N. (1992 yil yanvar). "Boron isotopic composition and concentration in modern marine carbonates". Geochimica va Cosmochimica Acta. 56 (1): 537–543. Bibcode:1992 yil GeCoA..56..537H. doi:10.1016/0016-7037(92)90151-8.
- ^ Kengash rahbari, R.S .; Cheetham, A.H .; Rowell, A.J. (1987). Fosil umurtqasizlar. Vili. ISBN 978-0865423022.
- ^ Jons, RW (1996). Neftni qidirishda mikropalaeontologiya. Clarendon Press. ISBN 978-0-19-854091-5.
- ^ Makneyl, D.H .; Issler, D.R .; Snoudon, L.R. (1996). Fotoalbom foraminiferlarida ranglarning o'zgarishi, termal etukligi va ko'milish diagenezi. Kanada byulleteni geologik xizmati. 499. Kanada geologik xizmati. ISBN 978-0-660-16451-9.
- ^ Uilkinson, Yan P.; Uilyams, Mark; Yosh, Jeremi R.; Kuk, Samanta R.; Fulford, Maykl G.; Lott, Grem K. (2008 yil 1-avgust). "Silchesterda Rim mozaikasini ishlab chiqarishda ishlatiladigan bo'rning isbotlanganligini baholashda mikrofosillarni qo'llash". Arxeologiya fanlari jurnali. 35 (8): 2415–2422. doi:10.1016 / j.jas.2008.03.010. ISSN 0305-4403.
Tashqi havolalar
- Umumiy ma'lumot
- Kaliforniya universiteti Paleontologiya muzeyi veb-saytida an Foraminiferaga kirish
- Janubiy Florida universiteti tadqiqotchilari tizimni ishlab chiqdilar marhamat atroflarini kuzatish uchun Foraminifera-dan foydalanish
- London Universitet kolleji mikropaleontologiya sayti ko'plab yuqori sifatli, shu jumladan Foraminifera haqida umumiy ma'lumotga ega SEMlar
- Foraminiferal tadqiqotlarda ishlatiladigan atamalarning lug'ati Lukas Xottingerning OA elektron jurnalida chop etilgan lug'ati "Carnets de Géologie - geologiya bo'yicha daftarlar"
- Foraminifera haqida ma'lumot Martin Langerning Mikropaleontologiya sahifasi
- Bentik Foraminifera haqida ma'lumot 2005 yil Urbino Yozgi Paleoklimatologiya maktabidan
- Onlayn flip-kitoblar
- Foraminiferal tadqiqotlarda ishlatiladigan atamalarning lug'ati tomonidan Lukas Xottinger (nashr etilganning muqobil versiyasi "Carnets de Géologie - geologiya bo'yicha daftarlar" )
- Resurslar
- pforams @ mikrotax - planktonik foraminifera taksonomiyasini batafsil bayon qiluvchi onlayn ma'lumotlar bazasi
- The yulduz * qum loyihasi (qismi micro * qamrov doirasi ) - bu Foraminifera haqidagi ma'lumotlarning kooperativ ma'lumotlar bazasi
- 3D modellar tomonidan yaratilgan foramkalar Rentgen tomografiyasi
- XRONOS bor bir nechta Foraminifera manbalari jumladan, a taksonlarni qidirish sahifasi va a mikro-paleo bo'limi NB Ushbu tarkibning aksariyati endi pforams @ mikrotax veb-saytiga kiritilgan
- eForams Foraminifera va foraminiferal chig'anoqlarni modellashtirishga qaratilgan veb-sayt
- Foraminifera galereyasi Yaqinda va qazib olingan Foraminiferalarning turkumi va joylashuvi bo'yicha tasvirlangan katalogi
- "Foraminifera". NCBI taksonomiyasi brauzeri. 29178.