Uchinchi sekretsiya tizimini kiriting - Type three secretion system
Uchinchi sekretsiya tizimini kiriting (ko'pincha yoziladi III turdagi sekretsiya tizimi va qisqartirilgan TTSS yoki T3SSdeb nomlangan Injectisome) a oqsil qo'shimchalar bir nechtasida topilgan Gram-manfiy bakteriyalar.
Patogen bakteriyalarda igna o'xshash tuzilish mavjudligini aniqlash uchun sensori zond sifatida ishlatiladi ökaryotik organizmlar va sir bakteriyalarga yordam beradigan oqsillar yuqtirish ularni. Yashirin effektor oqsillari to'g'ridan-to'g'ri bakteriyadan ajralib chiqadi hujayra eukaryotik (xost) hujayraga kirib, ular patogenning omon qolishiga va immunitet ta'siridan xalos bo'lishiga yordam beradigan bir qator ta'sir ko'rsatadi.
Umumiy nuqtai
III turdagi sekretsiya tizimi atamasi 1993 yilda paydo bo'lgan.[1] Ushbu sekretsiya tizimi kamida beshta tizimdan ajralib turadi sekretsiya tizimlari grammusbat bakteriyalarda uchraydi. Ko'pgina hayvon va o'simlik bilan bog'liq bakteriyalar o'xshash T3SSlarga ega. Ushbu T3SS divergent evolyutsiya natijasida o'xshashdir va filogenetik tahlil gram-manfiy bakteriyalar T3SS ni o'tkazishi mumkin bo'lgan modelni qo'llab-quvvatlaydi. gen kassetasi gorizontal ravishda boshqa turlarga. Eng ko'p o'rganilgan T3SS turlari Shigella (sabablari basillyar dizenteriya ), Salmonella (tifo isitmasi ), Escherichia coli (Ichak florasi, ba'zi shtammlar sabab bo'ladi ovqatdan zaharlanish ), Vibrio (gastroenterit va diareya ), Burxolderiya (bezlar ), Yersiniya (vabo ), Xlamidiya (jinsiy yo'l bilan yuqadigan kasallik ), Pseudomonas (yuqtiradi odamlar, hayvonlar va o'simliklar ) va o'simlik patogenlari Erviniya, Ralstoniya va Xanthomonas va o'simlik simbionti Rizobium.
T3SS taxminan 30 xil oqsildan iborat bo'lib, uni eng murakkab sekretsiya tizimlaridan biriga aylantiradi. Uning tuzilishi bakterial bilan juda ko'p o'xshashliklarni ko'rsatadi flagella (uchun ishlatiladigan uzun, qattiq, hujayradan tashqari tuzilmalar harakatchanlik ). T3SSda qatnashadigan ba'zi oqsillar aminokislota flagellar oqsillariga ketma-ketlik homologiyasi. T3SS-ga ega bo'lgan ba'zi bakteriyalar ham flagella va harakatchan (Salmonella, masalan,), ba'zilari esa yo'q (Shigella, masalan; misol uchun). Texnik jihatdan aytganda, III turdagi sekretsiya infektsiyaga bog'liq oqsillarni va flagellar tarkibiy qismlarini ajratish uchun ishlatiladi. Biroq, "III turdagi sekretsiya" atamasi asosan infektsiya apparatlariga nisbatan qo'llaniladi. Bakterial flagellum III turdagi sekretsiya tizimi bilan umumiy ajdodga ega.[2][3]
T3SSs ko'plab patogen bakteriyalarning patogenligi (yuqtirish qobiliyati) uchun juda muhimdir. T3SSdagi nuqsonlar bakteriyani patogen bo'lmagan holga keltirishi mumkin. Gram-manfiy bakteriyalarning ba'zi invaziv bo'lmagan shtammlari T3SSni yo'qotgan, chunki energetik jihatdan qimmat tizim endi ishlatilmayapti.[4] An'anaviy bo'lsa-da antibiotiklar o'tmishda ushbu bakteriyalarga qarshi samarali bo'lgan, antibiotiklarga chidamli shtammlar doimo paydo bo'ladi. T3SS ish uslubini tushunish va unga qaratilgan dori vositalarini ishlab chiqish 1990 yillarning oxiridan beri dunyodagi ko'plab tadqiqot guruhlarining muhim maqsadiga aylandi.
Tuzilishi
| III turdagi sekretsiya tizimi | |
|---|---|
 T3SS igna kompleksi | |
| Identifikatorlar | |
| Belgilar | T3SS |
| TCDB | 1. B.22 |
| OPM superfamily | 348 |
| OPM oqsili | 5tcq |
T3SS ning o'ziga xos belgisi - bu igna[5][6] (umuman olganda, igna kompleksi (Bosimining ko'tarilishi) yoki T3SS apparati (T3SA); ham chaqirdi injektomli qachon ATPase chiqarib tashlandi; pastga qarang). Yashirinishi kerak bo'lgan bakteriyalar oqsillari bakteriyalardan o'tadi sitoplazma igna orqali to'g'ridan-to'g'ri mezbon sitoplazmasiga. Uch membranalar ikkita sitoplazmani ajratib oling: gram-manfiy bakteriyaning ikki qavatli membranasi (ichki va tashqi membranalari) va evkaryotik membrana. Igna juda tanlangan va deyarli o'tkazmaydigan membranalar orqali silliq o'tishni ta'minlaydi. Bitta bakteriya membranasi bo'ylab tarqaladigan bir necha yuz igna kompleksiga ega bo'lishi mumkin. Igna kompleksi barcha patogen bakteriyalarning T3SSs universal xususiyati ekanligi taklif qilingan.[7]
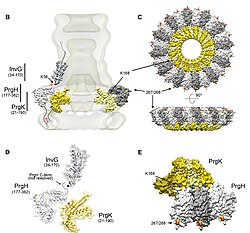
Igna kompleksi bakteriya sitoplazmasidan boshlanib, ikkita membranani kesib o'tib, hujayradan chiqib turadi. Membranaga biriktirilgan qism tayanch (yoki bazal tanasi) T3SS ning. Hujayra tashqari qismi bu igna. Deb nomlangan ichki tayoq ignani tayanch bilan bog'laydi. Igna o'zi, garchi T3SS ning eng katta va eng ko'zga ko'ringan qismi bo'lsa ham, bitta oqsilning ko'plab birliklaridan iborat. Shuning uchun har xil T3SS oqsillarining aksariyati bazani hosil qiluvchi va xostga ajraladigan oqsillardir. Yuqorida ta'kidlab o'tilganidek, igna kompleksi bakterial flagella bilan o'xshashliklarga ega. Aniqrog'i, igna kompleksining asosi tizimli ravishda flagellar tagiga juda o'xshash; ignaning o'zi flagellar kancasına o'xshashdir, bu asosni flagellar filament bilan bog'laydigan tuzilish.[8][9]
Baza bir nechta dumaloq halqalardan iborat bo'lib, yangi igna majmuasida qurilgan birinchi inshootdir. Baza qurib bo'lingandan so'ng, u tashqi oqsillarni (igna) sekretsiya mashinasi bo'lib xizmat qiladi. Butun kompleks qurib bo'lingandan so'ng tizim xujayralarga yuborilishi kerak bo'lgan sekretsiya oqsillariga o'tadi. Igna pastdan yuqoriga qarab qurilgan deb taxmin qilinadi; igna birliklari monomer bir-biriga oqsil qoziq, shunday qilib igna uchidagi birlik oxirgi qo'shilgan bo'ladi. Igna kichik birligi 9 k atrofida bo'lgan eng kichik T3SS oqsillaridan biridirDa. 100-150 subbirlik har bir ignadan iborat.
T3SS ignasi 60−80 atrofida nm uzunligi va tashqi kengligida 8 nm. U minimal uzunlikka ega bo'lishi kerak, shunda boshqa hujayradan tashqari bakterial tuzilmalar (yopishqoq moddalar va lipopolisakkarid masalan, qatlam) sekretsiyaga xalaqit bermaydi. Igna teshigi 3 nm diametrga ega. Katlanmış effektorli oqsillarning ko'pi igna teshigidan o'tish uchun juda katta, shuning uchun ko'p ajratilgan oqsillar ignadan o'tishi kerak ochildi, tomonidan bajarilgan vazifa ATPase strukturaning tagida.[10]
T3SS oqsillari

T3SS oqsillarini uchta toifaga birlashtirish mumkin:
- Strukturaviy oqsillar: taglikni, ichki tayoqni va ignani qurish.
- Effektor oqsillari: xujayra ichiga ajratiladi va infektsiyani rag'batlantiradi / xujayra xujayralarini himoya qilishni to'xtatadi.
- Chaperones: bakterial sitoplazmadagi effektorlarni bog'laydi, ularni birikishdan va tanazzul va ularni igna majmuasi tomon yo'naltiring.
T3SS genlarining aksariyati joylashtirilgan operonlar. Ushbu operonlar ba'zi turlardagi bakterial xromosomada va maxsus ajratilgan joyda joylashgan plazmid boshqa turlarda. SalmonellaMasalan, ko'pgina T3SS genlari to'plangan xromosoma mintaqasi mavjud Salmonella patogenlik oroli (SPI). ShigellaBoshqa tomondan, barcha T3SS genlari joylashgan katta virusli plazmidga ega. Shuni ta'kidlash kerakki, ko'plab patogenlikdagi orollar va plazmidlarda orol / plazmidning yangi turga tez-tez gorizontal ravishda gen o'tkazilishini ta'minlaydigan elementlar mavjud.
Igna orqali chiqarilishi kerak bo'lgan effektor oqsillari tizim tomonidan tan olinishi kerak, chunki ular minglab boshqa oqsillar bilan birga sitoplazmada suzadi. Tanib olish a orqali amalga oshiriladi sekretsiya signali- boshida joylashgan aminokislotalarning qisqa ketma-ketligi (The N-terminali ) oqsilni (odatda dastlabki 20 ta aminokislotada), bu igna kompleksi taniy oladi. Boshqa sekretsiya tizimlaridan farqli o'laroq, T3SS oqsillarining sekretsiya signali hech qachon oqsildan ajralmaydi.
Sekretsiyani induktsiya qilish
Igna bilan xujayraning aloqasi T3SSni sekretsiyani boshlashga olib keladi;[11] ushbu tetik mexanizmi haqida ko'p narsa ma'lum emas (pastga qarang). Konsentratsiyasini pasaytirish orqali sekretsiya ham paydo bo'lishi mumkin kaltsiy ionlari ichida o'sish muhiti (uchun Yersiniya va Pseudomonas; qo'shish orqali amalga oshiriladi chelator kabi EDTA yoki EGTA ) va qo'shib aromatik bo'yoq Kongo qizil o'sish muhitiga (uchun Shigella), masalan; misol uchun. Ushbu usullar va boshqa usullar laboratoriyada III turdagi sekretsiyani sun'iy ravishda qo'zg'atish uchun ishlatiladi.
Xujayra hujayralari bilan aloqa qilishdan tashqari, tashqi belgilar tomonidan sekretsiyani induktsiya qilish ham sodir bo'ladi jonli ravishda, yuqtirilgan organizmlarda. Bakteriyalar bunday signallarni sezadilar harorat, pH, osmolarlik va kislorod darajalari va ularni T3SS-ni faollashtirish to'g'risida "qaror qabul qilish" uchun foydalaning. Masalan; misol uchun, Salmonella -ni takrorlashi va bosib olishi mumkin yonbosh ichak o'rniga ko'richak hayvon ichak. Bakteriyalar ushbu hududlarda mavjud bo'lgan turli xil ionlar tufayli qaerdaligini bilish imkoniyatiga ega; yonbosh ichakni o'z ichiga oladi shakllantirish va atsetat, ko'r ichak esa yo'q. Bakteriyalar bu molekulalarni sezadi, ular ichakda ekanligini aniqlaydi va ularning sekretsiya mexanizmini faollashtiradi. Kabi ko'richakda mavjud bo'lgan molekulalar, masalan propionat va butirat, bakteriyalarga salbiy signal beradi va sekretsiyani inhibe qiladi. Xolesterin, a lipid ko'pgina eukaryotik hujayra membranalarida uchraydi, ichkarida sekretsiyani keltirib chiqarishi mumkin Shigella.
Yuqorida sanab o'tilgan tashqi belgilar sekretsiyani to'g'ridan-to'g'ri yoki genetik mexanizm orqali tartibga soladi. Bir nechta transkripsiya omillari tartibga soluvchi ifoda T3SS genlari ma'lum. T3SS effektorlarini bog'laydigan ba'zi shaperonlar transkripsiya omillari sifatida ham ishlaydi. Teskari aloqa mexanizmi taklif qilingan: bakteriya ajralib chiqmasa, uning effektor oqsillari shaperonlar bilan bog'lanib, sitoplazmada suzadi. Sekretsiya boshlanganda chaperonlar effektorlardan ajralib, ikkinchisi ajralib chiqadi va hujayradan chiqib ketadi. So'ngra yolg'iz chaperonlar transkripsiya omillari vazifasini bajaradi, ularning ta'sir etuvchilarini kodlovchi genlar bilan bog'lanib, ularning transkripsiyasini keltirib chiqaradi va shu bilan ko'proq effektorlar ishlab chiqaradi.
Type3SS injeksisomalariga o'xshash tuzilmalar bakteriyalar sekretsiyasini eukaryotik xostga yoki in vivo jonli ravishda boshqa maqsad hujayralariga etkazib berishga qaratilgan tashqi membrana pufakchalarini bo'shatishga yordam beradigan salbiy bakterial tashqi va ichki membranalarni perçinleme qilish uchun taklif qilingan.[12]
T3SS vositachiligidagi infektsiya
T3SS effektorlari poydevoridagi igna kompleksiga kirib, xujayra xujayrasi tomon igna ichiga kirib boradi. Efaktorlarning xostga kirishining aniq usuli asosan noma'lum. Ilgari, ignaning o'zi mezbon hujayra membranasidagi teshikni teshishga qodir, deb taxmin qilingan edi; ushbu nazariya rad etildi. Hozir aniqki, ba'zi bir efaktorlar umumiy nom bilan atalgan translokatorlar, oldin ajralib chiqadi va teshik yoki kanal hosil qiladi (a translokon) mezbon hujayra membranasida, ular orqali boshqa effektorlar kirishi mumkin. Mutatsiyaga uchragan translokatorlar mavjud bo'lmagan bakteriyalar oqsillarni ajratib olishga qodir, ammo ularni xujayra hujayralariga etkaza olmaydi. Umuman olganda har bir T3SS uchta translokatorni o'z ichiga oladi. Ba'zi translokatorlar ikki baravar rol o'ynaydi; ular teshik hosil bo'lishida qatnashgandan keyin hujayraga kirib, o'z vazifalarini bajaradilar halollik bilan, insof bilan effektorlar.
T3SS effektorlari xost hujayralarini bir necha usul bilan boshqaradi. Eng hayratlanarli ta'sir bu xujayra xujayrasi tomonidan bakteriyani yutishini rag'batlantirishdir. T3SS-larga ega bo'lgan ko'plab bakteriyalar infektsiyani takrorlash va ko'paytirish uchun xujayralarga kirishi kerak. Ular xujayra hujayralariga kiritadigan efektorlar xujayrani bakteriyani yutib yuborishga va uni amalda "eyishga" undaydi. Buning uchun bakterial effektorlar manipulyatsiya qilishadi aktin polimerizatsiya mezbon hujayraning texnikasi. Aktin - ning tarkibiy qismi sitoskelet shuningdek, u harakatchanlikda va hujayra shaklidagi o'zgarishlarda ishtirok etadi. T3SS effektorlari orqali bakteriya xujayraning o'z texnikasidan o'z manfaati uchun foydalana oladi. Bakteriya hujayraga kirgandan so'ng boshqa effektorlarni osonroq ajratishi mumkin va u qo'shni hujayralarga kirib, tezda butunni yuqtirishi mumkin. to'qima.
Shuningdek, T3SS effektorlari xostnikini buzgani ko'rsatilgan hujayra aylanishi va ularning ba'zilari qo'zg'atishga qodir apoptoz. Eng ko'p o'rganilgan T3SS effektorlaridan biri IpaB dan Shigella flexneri. U mezbon hujayra membranasida teshik hosil qiluvchi translokator sifatida ham, efir vazifasini o'tab, mezbon hujayraga zararli ta'sir ko'rsatadi. IpaB ning apoptozni keltirib chiqarishi isbotlangan makrofaglar - hayvon hujayralari immunitet tizimi - ular tomonidan qamrab olgandan keyin.[13] Keyinchalik IpaB o'zaro ta'sir o'tkazish orqali bunga erishishi ko'rsatildi kaspaz 1, ökaryotik hujayralardagi asosiy tartibga soluvchi oqsil.[14]
T3SS effektorlarining yana bir yaxshi tavsiflangan klassi Transkripsiya aktivatoriga o'xshash effektorlardir (TAL effektorlari ) dan Xanthomonas. Ushbu oqsillar o'simliklarga yuborilganda o'simlik hujayrasi yadrosiga kirib, o'simliklarning promotorlari ketma-ketligini bog'lashi va bakteriyalar infektsiyasiga yordam beradigan o'simlik genlarining transkripsiyasini faollashtirishi mumkin.[15] Yaqinda TAL effektor-DNKni aniqlash oddiy kodni o'z ichiga olganligi isbotlandi[16][17] va bu ushbu oqsillar mezbon o'simlik hujayralarida genlarning transkripsiyasini qanday o'zgartirishi mumkinligi haqidagi tushunchani ancha yaxshilaydi.
Hal qilinmagan muammolar
90-yillarning o'rtalaridan beri T3SS bo'yicha yuzlab maqolalar nashr etilgan. Ammo tizim bilan bog'liq ko'plab muammolar hal qilinmagan:
- T3SS oqsillari. Har bir organizmda 10 tadan kam bo'lgan taxminan 30 ta T3SS oqsilidan to'g'ridan-to'g'ri aniqlangan biokimyoviy usullari. Qolganlari, ehtimol kamdan-kam hollarda, ularni aniqlash qiyin bo'lgan va ular nazariy bo'lib qolmoqda (garchi ko'plab T3SS genlari / oqsillarida biokimyoviy emas, balki genetik tadqiqotlar o'tkazilgan bo'lsa ham). Har bir oqsilning lokalizatsiyasi ham to'liq ma'lum emas.
- Igna uzunligi. Yangi igna kerakli uzunlikka etganida bakteriya qanday qilib "bilishi" ma'lum emas. Bir nechta nazariyalar mavjud, ular orasida igna uchi va tagini qandaydir tarzda bog'laydigan "hukmdor oqsil" mavjud. Igna uchiga yangi monomerlar qo'shilishi o'lchagich oqsilini cho'zishi va shu bilan igna uzunligini taglikka ishora qilishi kerak.
- Energetika. Igna ichidagi oqsillarni harakatga keltiruvchi kuch to'liq ma'lum emas. An ATPase T3SS bazasi bilan bog'liq va oqsillarni ignaga yo'naltirishda ishtirok etadi; ammo transport uchun energiya etkazib beradimi-yo'qmi aniq emas.
- Yashirin signal. Yuqorida aytib o'tganimizdek, effektor oqsillarida sekretsiya signalining mavjudligi ma'lum. Signal tizimga T3SS-tashilgan oqsillarni boshqa har qanday oqsildan ajratib olishga imkon beradi. Uning mohiyati, talablari va tanib olish mexanizmi juda yaxshi tushunilmagan, ammo qaysi bakteriyalar oqsillarini III tip sekretsiya tizimi orqali tashish mumkinligini taxmin qilish usullari yaqinda ishlab chiqilgan.[19]
- Sekretsiyani faollashtirish. Bakteriya effektorlarni ajratish vaqti qachon kelganligini bilishi kerak. Keraksiz sekretsiya, biron bir hujayra xujayrasi yaqin joyda bo'lmaganda, energiya va resurslar jihatidan bakteriya uchun behuda bo'ladi. Bakteriya qandaydir tarzda ignaning xujayra xujayrasi bilan aloqasini taniy oladi. Bu qanday amalga oshirilayotgani hali o'rganilmoqda va usul patogenga bog'liq bo'lishi mumkin. Ba'zi nazariyalar xujayra xujayrasi bilan aloqa qilishda igna tuzilishidagi nozik konformatsion o'zgarishni postulyatsiya qiladi; bu o'zgarish balki bazaning sekretsiyani boshlashi uchun signal bo'lib xizmat qilishi mumkin. Tanib olishning bir usuli kashf etilgan Salmonella, bu mezbon hujayra sitosolini sezishga tayanadi pH patogenlik orolida efektorlar sekretsiyasini yoqish uchun 2 ta kodlangan T3SS.[20]
- Chaperonlarni bog'lash. Chaperonlar o'z ta'sirini qachon bog'lashi ma'lum emas (keyin yoki undan keyin) tarjima ) va ular sekretsiya oldidan qanday qilib o'zlarining ta'sir etuvchilardan ajralishi.
- Effektor mexanizmlari. 21-asrning boshidan beri T3SS effektorlarining xostni boshqarish usullari haqida ko'p narsa ochilgan bo'lsa-da, aksariyat effektlar va yo'llar noma'lum bo'lib qolmoqda.
- Evolyutsiya. Yuqorida aytib o'tilganidek, T3SS bakterial flagellum bilan chambarchas bog'liq.[21] Uchta raqobatdosh gipoteza mavjud:[22] birinchidan, flagellum birinchi bo'lib rivojlangan va T3SS shu tuzilishdan kelib chiqqan, ikkinchidan, T3SS avval evolyutsiyalangan va flagellum undan olingan, uchinchidan, ikkala tuzilma umumiy ajdoddan kelib chiqqan. Turli xil senariylar haqida ba'zi tortishuvlar bo'lgan,[2][22] chunki ularning barchasi ikkala tuzilish orasidagi protein homologiyasini va ularning funktsional xilma-xilligini tushuntiradi.[23] Shunga qaramay, so'nggi filogenomik dalillar T3SS flagellumdan genlarni dastlabki yo'qotish va keyinchalik genlarni olishni o'z ichiga olgan jarayon natijasida kelib chiqqanligi haqidagi gipotezani qo'llab-quvvatlamoqda.[24] Oxirgi jarayonning muhim bosqichi T3SS ga sekretinlarni jalb qilish edi, bu hodisa boshqa membranalar bilan bog'liq tizimlardan kamida uch marta sodir bo'lgan.
T3SS oqsillarining nomlanishi

1990-yillarning boshidan boshlab T3SS-ning yangi oqsillari turli xil bakteriyalar turlarida barqaror tezlikda topilmoqda. Qisqartmalar har bir organizmdagi oqsillarning har bir qatori uchun mustaqil ravishda berilgan va nomlar, odatda, oqsilning ishi haqida juda ko'p ma'lumot bermaydi. Mustaqil ravishda turli bakteriyalarda topilgan ba'zi oqsillar keyinchalik isbotlangan gomologik; ammo tarixiy nomlar asosan saqlanib qolgan, bu chalkashliklarni keltirib chiqarishi mumkin bo'lgan fakt. Masalan, SicA, IpgC va SycD oqsillari gomologlardir Salmonella, Shigella va Yersiniyanavbati bilan, lekin ularning nomidagi oxirgi harf ("seriya raqami") buni ko'rsatmaydi.
Quyida T3SS o'z ichiga olgan bir nechta turlarning eng keng tarqalgan oqsil seriyali nomlari haqida qisqacha ma'lumot berilgan. E'tibor bering, ushbu nomlar tarkibiga T3SS texnikasini hosil qiluvchi oqsillar ham, ajratilgan moddalar ham kiradi effektor oqsillari:
- Yersiniya
- Yop: Yersiniya tashqi oqsil
- Ysc: Yersiniya sekretsiya (komponent)
- Ypk: Yersiniya protein kinaz
- Salmonella
- Spa: Antigenning yuzaki namoyishi
- Sic: Salmonella chaperone bosqini
- Sip: Salmonella bosqinchi oqsil
- Prg: PhoP-repressiya qilingan gen
- Inv: Bosqin
- Org: Kislorod bilan boshqariladigan gen
- Ssp: Salmonella- sirli oqsil
- Iag: Invasion bilan bog'liq gen
- Shigella
- Ipg: Invasion plazmid geni
- Ipa: Invasion plazmid antijeni
- Mxi: Ipa ning membranani ifodasi
- Spa: Antigenning yuzaki namoyishi
- Osp: Tashqi Shigella oqsil
- Esherichiya
- Tir: Translokatsiyalangan intimin retseptorlari
- Sentyabr: Sekretsiyasi E. coli oqsillar
- Esc: Esherichiya sekretsiya (komponent)
- Xususan: Esherichiya sekretsiya oqsili
- Ces: Chaperone of E. coli sekretsiya
- Pseudomonas
- Hrp: Yuqori sezuvchanlik va patogenlik
- Hrc: Haddan tashqari sezgir javob saqlanadi (yoki Hrp saqlanib qoladi)
- Rizobium
- Yo'q: Nodulyatsion oqsil
- Rhc: Rizobium saqlanib qolgan
- Bir nechta turlarda:
- Vir: Virulentlik
- "Protochlamydia amoebophila"
- "Sodalis glossinidius"[25]
Ushbu qisqartmalardan keyin harf yoki raqam mavjud. Maktublar odatda "seriya raqami" ni, kashfiyotning xronologik tartibini yoki genning fizik ko'rinish tartibini operon. Raqamlar, kamdan-kam holatlar, tarkibidagi oqsilning molekulyar og'irligini bildiradi kDa. Misollar: IpaA, IpaB, IpaC; MxiH, MxiG, MxiM; Spa9, Spa47.
Barcha T3SSlarda bir nechta asosiy elementlar paydo bo'ladi: igna monomeri, ignaning ichki tayog'i, halqa oqsillari, ikkita translokator, igna uchi oqsili, o'lchagich oqsili (bu igna uzunligini aniqlaydi deb o'ylashadi; yuqoriga qarang) va The ATPase, bu sekretsiya uchun energiya etkazib beradi. Quyidagi jadvalda T3SS o'z ichiga olgan to'rtta bakteriyalar tarkibidagi ushbu asosiy oqsillarning bir qismi ko'rsatilgan:
| ↓ Funktsiya / Jins → | Shigella | Salmonella | Yersiniya | Esherichiya |
|---|---|---|---|---|
| Igna monomeri | MxiH | PrgI | YscF | EscF |
| Ichki tayoq | MxiI | PrgJ | YscI | EscI |
| Igna uchi oqsili | IpaD | SipD | LcrV | EspA |
| Translokator | IpaB | SipB | YopB | EspD |
| Translokator | IpaC | SipC | YopD | EspB |
| Ikki translokator uchun shaperone | IpgC | SicA | SycD | CesD |
| ATPase | Spa 47 | InvC | YscN | SepB (EscN) |
| Hukmdor oqsil | Spa 32 | InvJ | YscP | Orf16 |
| Kommutator | Spa 40 | SpaS | Yscu | EskU |
| Darvozabon | MxiC | InvE | YopN (TyeA) | SepL |
T3SS tadqiqotlarida qo'llaniladigan usullar
T3SS igna komplekslarini izolyatsiyasi
Katta, mo'rt, hidrofob hujayralardagi membrana tuzilmalari ko'p yillar davomida muammo bo'lib kelgan. 1990-yillarning oxiriga kelib, T3SS NClarni ajratib olish uchun bir nechta yondashuvlar ishlab chiqildi. 1998 yilda birinchi NClar ajratilgan Salmonella typhimurium.[26]
Izolyatsiya qilish uchun bakteriyalar katta miqdordagi suyuqlikda o'stiriladi o'sish muhiti ular yetguncha log bosqichi. Ular keyin santrifüj qilingan; The superfant (vosita) tashlanadi va granulalar (bakteriyalar) qayta tiklanadi lizis buferi odatda o'z ichiga oladi lizozim va ba'zan a yuvish vositasi kabi LDAO yoki Triton X-100. Ushbu tampon hujayra devori. Bir necha marta lizis va yuvishdan keyin ochilgan bakteriyalar bir qator ta'sir ko'rsatadi ultrasentrifugalar. Ushbu davolash katta makromolekulyar tuzilmalarni boyitadi va kichik hujayralar tarkibiy qismlarini tashlaydi. Ixtiyoriy ravishda, yakuniy lizat keyingi tozalashga duchor bo'ladi CSCl zichlik gradyenti.
Keyinchalik tozalashdan foydalanish uchun qo'shimcha yondashuv yaqinlik xromatografiyasi. Rekombinant A olib boruvchi T3SS oqsillari oqsil yorlig'i (a histidin yorlig'i, masalan) tomonidan ishlab chiqarilgan molekulyar klonlash va keyin kiritildi (o'zgartirildi ) o'rganilgan bakteriyalarga. Boshlang'ich bosimining izolyatsiyasidan so'ng, yuqorida aytib o'tilganidek, lizat yorliqqa yuqori yaqinligi bo'lgan zarrachalar bilan qoplangan ustun orqali o'tadi (gistidin teglarida: nikel ionlari ). Belgilangan oqsil kolonda saqlanadi va shu bilan butun igna kompleksi. Bunday usullar yordamida yuqori darajadagi poklikka erishish mumkin. Ushbu poklik bosimining ko'tarilishini tavsiflash uchun ishlatilgan ko'plab nozik tahlillar uchun juda muhimdir.
III toifadagi effektorlar 1990-yillarning boshlaridan beri ma'lum bo'lgan, ammo ularni xujayralarga etkazib berish usuli to'liq sir edi. Ko'pchilik o'rtasidagi homologiya flagellar va T3SS oqsillari tadqiqotchilarni flagellarga o'xshash tashqi T3SS tuzilishi mavjudligidan shubha qilishlariga olib keldi. Igna tuzilishini aniqlash va undan keyin ajratish tadqiqotchilarga:
- NCning uch o'lchovli tuzilishini batafsil tavsiflash va shu orqali sekretsiya mexanizmi to'g'risida xulosalar chiqarish (masalan, ignaning tor kengligi sekretsiya oldidan effektorlarni ochishni talab qiladi),
- oqsil tarkibiy qismlarini tahlil qilish, bu izolyatsiya qilingan ignalarni proteomik tahlilga kiritish orqali (pastga qarang),
- turli xil bosimining ko'tarilish qismlariga rollarni belgilash, bu bilan nokaut T3SS genlari, mutatsiyaga uchragan bakteriyalardan NC ni ajratib olish va mutatsiyalar sabab bo'lgan o'zgarishlarni o'rganish.
Mikroskopiya, kristallografiya va qattiq holatdagi NMR
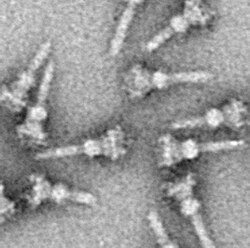
Deyarli barcha oqsillarda bo'lgani kabi, T3SS NC ni vizualizatsiya qilish faqat mumkin elektron mikroskopi. NClarning birinchi rasmlarida (1998) tirik bakteriyalar va tekis, ikki o'lchovli ajratilgan bosim o'tkazgichlarning hujayra devoridan chiqayotgan igna tuzilmalari tasvirlangan.[26] 2001 yilda NClarning tasvirlari Shigella flexneri raqamli ravishda tahlil qilindi va NC ning birinchi yarim 3D tuzilishini olish uchun o'rtacha.[5] NC-larning spiral tuzilishi Shigella flexneri 16-sonli qaror bilan hal qilindi Å foydalanish Rentgen tolaning difraksiyasi 2003 yilda,[27] va bir yildan keyin 17-Å NClarning 3D tuzilishi Salmonella typhimurium nashr etildi.[28] So'nggi yutuqlar va yondashuvlar NC ning yuqori aniqlikdagi 3D tasvirlariga imkon berdi,[29][30] NCning murakkab tuzilishini yanada aniqlashtirish.
Ko'p yillar davomida ko'plab T3SS oqsillari kristallangan. Bularga NC tarkibidagi oqsillar, effektorlar va chaperonlar kiradi. Igna kompleksi monomerining birinchi tuzilishi "Burkholderia pseudomallei" dan BsaL ning NMR tuzilishi va keyinchalik MixH ning kristalli tuzilishi edi. Shigella flexneriikkalasi ham 2006 yilda hal qilingan.[31][32]
2012 yilda rekombinant yovvoyi turdagi igna ishlab chiqarish kombinatsiyasi, qattiq holatdagi NMR, elektron mikroskopi[33] va Rozetta modellashtirish natijasida supramolekulyar interfeyslar va oxir-oqibat atomning to'liq tuzilishi aniqlandi Salmonella typhimurium T3SS ignasi.[34] 80 ta qoldiq PrgI bo'linmalari o'ng burama spiral birikmasini hosil qilganligi, har ikki burilishda taxminan 11 ta bo'linma bilan o'xshashligi ko'rsatilgan. flagellum ning Salmonella typhimurium. Model, shuningdek, igna yuzasida joylashgan kengaytirilgan amino-terminal domenini aniqladi, yuqori konservalangan karboksi terminali esa lümen tomon yo'naladi.[34]
Proteomika
T3SS tarkibiga kiradigan oqsillarni aniqlash uchun bir necha usullardan foydalanilgan. Izolyatsiya qilingan igna komplekslarini ajratish mumkin SDS-PAGE. Bo'yashdan keyin paydo bo'ladigan bantlar jeldan alohida ravishda chiqarilishi va ulardan foydalanib tahlil qilinishi mumkin oqsillarni ketma-ketligi va mass-spektrometriya. NC ning tarkibiy qismlarini bir-biridan ajratish mumkin (masalan, igna qismi tayanch qismidan) va shu fraktsiyalarni tahlil qilish orqali har birida qatnashadigan oqsillarni ajratib olish mumkin. Shu bilan bir qatorda, izolyatsiya qilingan NClarni to'g'ridan-to'g'ri mass spektrometriyasi yordamida tahlil qilish mumkin elektroforez, NC haqida to'liq tasavvurga ega bo'lish uchun proteom.
Genetik va funktsional tadqiqotlar
Ko'pgina bakteriyalardagi T3SS tadqiqotchilar tomonidan boshqarilgan. Shaxsiy manipulyatsiya ta'sirini kuzatish yordamida tizimning har bir tarkibiy qismining roli to'g'risida tushuncha berish mumkin. Manipulyatsiya misollari:
- Bir yoki bir nechta T3SS genlarini yo'q qilish (genlarni nokaut qilish ).
- Haddan tashqari ifoda bir yoki bir nechta T3SS genlari (boshqacha aytganda: ishlab chiqarish jonli ravishda odatdagidan kattaroq miqdordagi T3SS oqsilidan).
- T3SS genlari yoki oqsillarida nuqta yoki mintaqaviy o'zgarishlar. Bu oqsil tarkibidagi o'ziga xos aminokislotalar yoki mintaqalarning funktsiyasini aniqlash uchun amalga oshiriladi.
- Bakteriyalarning bir turidan gen yoki oqsilni boshqasiga kiritish (o'zaro komplementatsiya tahlili). Bu ikkita T3SS o'rtasidagi farqlar va o'xshashliklarni tekshirish uchun amalga oshiriladi.
T3SS komponentlarini manipulyatsiyasi bakteriyalar funktsiyasi va patogenligining bir necha jihatlariga ta'sir qilishi mumkin. Mumkin bo'lgan ta'sirga misollar:
- Hujayra ichidagi patogenlar holatida bakteriyalarning mezbon hujayralarni bosib olish qobiliyati. Buni an yordamida o'lchash mumkin bosqinni tahlil qilish (gentamisindan himoya qilish tahlili ).
- Hujayra ichidagi bakteriyalarning mezbon hujayralar o'rtasida ko'chib yurish qobiliyati.
- Bakteriyalarning mezbon hujayralarni o'ldirish qobiliyati. Buni bir necha usul bilan o'lchash mumkin, masalan LDH - tahlilni bo'shating, unda ferment O'lik hujayralardan oqib chiqadigan LDH uning fermentativ faolligini o'lchash orqali aniqlanadi.
- T3SS ning o'ziga xos oqsilni ajratishi yoki umuman chiqarishi qobiliyati. Buni tahlil qilish uchun suyuq muhitda o'sadigan bakteriyalarda sekretsiya paydo bo'ladi. Keyin bakteriyalar va muhitni santrifüjlash yo'li bilan ajratib olinadi, so'ngra ajratilgan oqsillar borligi uchun o'rtacha fraktsiya (superfatant) tahlil qilinadi. Odatda salgılanan oqsilning chiqarilishini oldini olish uchun unga sun'iy ravishda katta molekula biriktirilishi mumkin. Agar o'sha paytda ajratilmagan oqsil igna kompleksining pastki qismida "tiqilib" qolsa, sekretsiya samarali ravishda bloklanadi.
- Bakteriyalarning buzilmagan igna kompleksini yig'ish qobiliyati. NClarni manipulyatsiya qilingan bakteriyalardan ajratib olish va mikroskopik tekshirish mumkin. Kichkina o'zgarishlarni har doim ham mikroskop yordamida aniqlash mumkin emas.
- Bakteriyalarning tirik hayvonlar yoki o'simliklarni yuqtirish qobiliyati. Hatto manipulyatsiya qilingan bakteriyalar ko'rsatilsa ham in vitro xujayrali hujayralarni yuqtirish imkoniyatiga ega bo'lish uchun, ularning tirik organizmda infektsiyani ushlab turish qobiliyati beparvo bo'lmasligi mumkin.
- Boshqa genlarning ekspression darajalari. Buni bir necha usul bilan, xususan, tahlil qilish mumkin shimoliy blot va RT-PCR. Hammaning ifoda darajalari genom tomonidan tahlil qilinishi mumkin mikroarray. Ko'p turlari III transkripsiya omillari va ushbu usullar yordamida tartibga soluvchi tarmoqlar topildi.
- Bakteriyalarning ko'payishi va yaroqliligi.
T3SS inhibitörleri
T3SS ni inhibe qiladigan bir nechta birikmalar topildi grammusbat bakteriyalar shu jumladan guadinominlar tomonidan tabiiy ravishda ishlab chiqarilgan Streptomitsiyalar turlari.[35] Monoklonal antikorlar T3SS ni ham inhibe qiladigan ishlab chiqilgan.[36]
III turdagi signal peptidini bashorat qilish vositalari
Adabiyotlar
- ^ Salmond GP, Rivz PJ (1993). "Graf-salbiy bakteriyalarda membranalar harakati va oqsil sekretsiyasi". Biokimyo fanlari tendentsiyalari. 18 (1): 7–12. doi:10.1016/0968-0004(93)90080-7. PMID 8438237.
- ^ a b Gophna U, Ron EZ, Graur D (2003 yil iyul). "Bakteriyalarning III tipdagi sekretsiya tizimlari qadimgi va ko'plab gorizontal-uzatish hodisalari bilan rivojlangan". Gen. 312: 151–63. doi:10.1016 / S0378-1119 (03) 00612-7. PMID 12909351.
- ^ Nguyen L, Paulsen IT, Tchieu J, Hueck CJ, Saier MH (aprel 2000). "III turdagi oqsil sekretsiyasi tizimining tarkibiy qismlarini filogenetik tahlillari". Molekulyar mikrobiologiya va biotexnologiya jurnali. 2 (2): 125–44. PMID 10939240.
- ^ Gong X, Vu GP, Bai Y, Yang E, Lyu F, Lu S (2010 yil yanvar). "Salmonella III tip sekretsiya tizimi InvJ, PrgJ, SipC, SipD, SopA va SopB omillarini madaniyatlarda va sichqonlarda differentsial ifodalash". Mikrobiologiya. 156 (Pt 1): 116-27. doi:10.1099 / mic.0.032318-0. PMC 2889428. PMID 19762438.
- ^ a b Bloker A, Jouihri N, Larquet E, Gounon P, Ebel F, Parsot C, Sansonetti P, Allaoui A (2001). "Shigella flexneri" igna kompleksi "ning tuzilishi va tarkibi, uning III tipli sekretsiyasining bir qismi". Mol mikrobiol. 39 (3): 652–663. doi:10.1046 / j.1365-2958.2001.02200.x. PMID 11169106.
- ^ Galan JE, Wolf-Watz H (2006). "Eukaryotik hujayralarga oqsilni III turdagi sekretsiya mashinalari orqali yuborish". Tabiat. 444 (7119): 567–573. Bibcode:2006 yil Noyabr 444..567G. doi:10.1038 / nature05272. PMID 17136086.
- ^ Pallen M. J.; Beyli C. M.; Beatson S. A. (2006). "Flih / Yscl-ga o'xshash oqsillar o'rtasidagi bakteriyalarning III-sekretsiya tizimlari va FoF1 va vakuolyar ATPazlarning ikkinchi sopi tarkibiy qismlari o'rtasidagi evolyutsiya aloqalari". Proteinli fan. 15 (4): 935–940. doi:10.1110 / ps.051958806. PMC 2242474. PMID 16522800.
- ^ Aizawa S (2001). "Bakterial flagella va II tip sekretsiya tizimlari". FEMS mikrobiologiya xatlari. 202 (2): 157–164. doi:10.1111 / j.1574-6968.2001.tb10797.x. PMID 11520608.
- ^ Doolittle W. F.; Zhaxybayeva, Olga (2007). "Evolyutsiya: kamaytiriladigan murakkablik - bakterial flagella uchun holat". Hozirgi biologiya. 17 (13): R510-512. doi:10.1016 / j.cub.2007.05.003. PMID 17610831.
- ^ Akeda Y, Galan JE (2005 yil oktyabr). "III turdagi sekretsiyadagi substratlarning shaperon chiqishi va ochilishi". Tabiat. 437 (7060): 911–5. Bibcode:2005 yil Noyabr 433..911A. doi:10.1038 / nature03992. PMID 16208377.
- ^ Kimbrouz T. G.; Miller S. I. (2000). "Salmonella typhimurium II tip sekretsiya tarkibiy qismlarining igna kompleksi shakllanishiga qo'shgan hissasi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 97 (20): 11008–11013. Bibcode:2000PNAS ... 9711008K. doi:10.1073 / pnas.200209497. PMC 27139. PMID 10984518.
- ^ YashRoy R.C. (2003). "Gram-manfiy patogenlar tomonidan evkaryotik hujayraning intoksikatsiyasi: III turdagi sekretsiya tizimi uchun yangi bakterial tashqi membrana bilan bog'langan nanovesikulyar ekzotsitoz modeli". Xalqaro toksikologiya. 10 (1): 1–9.
- ^ Zychlinsky A, Kenny B, Menard R, Prevost MC, Holland IB, Sansonetti PJ (1994). "IpaB Shigella flexneri tomonidan qo'zg'atilgan makrofag apoptoziga vositachilik qiladi". Mol mikrobiol. 11 (4): 619–627. doi:10.1111 / j.1365-2958.1994.tb00341.x. PMID 8196540.
- ^ Xilbi H, Moss JE, Xersh D, Chen Y, Arondel J, Banerji S, Flavell RA, Yuan J, Sansonetti PJ, Zychlinskiy A (1998). "Shigella tomonidan qo'zg'atilgan apoptoz IpaB bilan bog'langan Caspase-1ga bog'liq". J Biol Chem. 273 (49): 32895–32900. doi:10.1074 / jbc.273.49.32895. PMID 9830039.
- ^ Boch, J .; Bonas, U. (2010). "XanthomonasAvrBs3 III turdagi oilaviy effektlar: kashfiyot va funktsiya". Fitopatologiyaning yillik sharhi. 48: 419–436. doi:10.1146 / annurev-fito-080508-081936. PMID 19400638.
- ^ Moscou, M. J .; Bogdanove, A. J. (2009). "Oddiy shifr TAL effektorlari tomonidan DNKning tan olinishini boshqaradi". Ilm-fan. 326 (5959): 1501. Bibcode:2009 yil ... 326.1501M. doi:10.1126 / science.1178817. PMID 19933106.
- ^ Boch J, Scholze H, Schornack S va boshq. (2009 yil dekabr). "TAL tip III effektorlarining DNK bilan bog'lanish o'ziga xosligi kodini buzish". Ilm-fan. 326 (5959): 1509–12. Bibcode:2009 yil ... 326.1509B. doi:10.1126 / science.1178811. PMID 19933107.
- ^ Shraytt O .; Lefebre, M. D .; Brunner, M. J .; Shmyed, V. X.; Shmidt, A .; Radiklar, J .; Mextler, K .; Galan, J. E .; Marlovits, T. C. (2010). Stebbinlar, C. Erec (tahrir). "Salmonella typhimurium III tip sekretsiya ignalarining kompleks tarkibiy qismlari topologiyasi va tashkil etilishi". PLOS patogenlari. 6 (4): e1000824. doi:10.1371 / journal.ppat.1000824. PMC 2848554. PMID 20368966.
- ^ Grynberg M, Godzik A (2009 yil aprel). Stebbinlar, C. Erec (tahrir). "Signal uchun signal topildi". PLOS Pathog. 5 (4): e1000398. doi:10.1371 / journal.ppat.1000398. PMC 2668190. PMID 19390616.
- ^ Yu XJ va boshqalar. (2010 yil may). "hujayra ichidagi Salmonella orqali pHni aniqlash effektor translokatsiyasini keltirib chiqaradi". Ilm-fan. 328 (5981): 1040–3. Bibcode:2010Sci ... 328.1040Y. doi:10.1126 / science.1189000. hdl:10044/1/19679. PMC 6485629. PMID 20395475.
- ^ Medini D, Covacci A, Donati C (2006 yil dekabr). "Proteinli gomologik tarmoq oilalari III va IV turdagi sekretsiya tizimlarining bosqichma-bosqich diversifikatsiyasini ochib berishdi". PLOS hisoblash. Biol. 2 (12): e173. Bibcode:2006PLSCB ... 2..173M. doi:10.1371 / journal.pcbi.0020173. PMC 1676029. PMID 17140285.
- ^ a b Saier, M (2004). "Bakterial III turdagi oqsil sekretsiyasi tizimlarining evolyutsiyasi". Mikrobiologiya tendentsiyalari. 12 (3): 113–115. doi:10.1016 / j.tim.2004.01.003. PMID 15001186.
- ^ Makken XN, Guttman DS (2008). "III turdagi sekretsiya tizimining rivojlanishi va uning o'simlik-mikroblarning o'zaro ta'sirida ta'siri". Yangi fitol. 177 (1): 33–47. doi:10.1111 / j.1469-8137.2007.02293.x. PMID 18078471.
- ^ Ebbi, Sofi S.; Rocha, Eduardo P. C. (2012-09-01). "Flagellar bo'lmagan III turdagi sekretsiya tizimi bakteriyalar flagellumidan rivojlanib, mezbon hujayralarga moslashgan tizimlarga aylandi". PLOS Genetika. 8 (9): e1002983. doi:10.1371 / journal.pgen.1002983. ISSN 1553-7404. PMC 3459982. PMID 23028376.
- ^ Moran, N. A. (2001 yil 13 fevral). "Hasharotlar ichidagi bakteriyalarni boshqarish". Milliy fanlar akademiyasi materiallari. 98 (4): 1338–1340. doi:10.1073 / pnas.98.4.1338.
- ^ a b Kubori T, Matsushima Y, Nakamura D, Uralil J, Lara-Tejero M, Suxan A, Galan JE, Aizava SI (aprel 1998). "Salmonella typhimurium III tip oqsil sekretsiyasi tizimining supramolekulyar tuzilishi". Ilm-fan. 280 (5363): 602–5. Bibcode:1998Sci ... 280..602K. doi:10.1126 / science.280.5363.602. PMID 9554854.
- ^ Cordes FS, Komoriya K, Larquet E, Yang S, Egelman EH, Bloker A, Lea SM (2003). "Shigella flexneri III tipli sekretsiya tizimi ignasining spiral tuzilishi". J Biol Chem. 278 (19): 17103–17107. doi:10.1074 / jbc.M300091200. PMID 12571230.
- ^ Marlovits TC, Kubori T, Suxan A, Tomas DR, Galan JE, Unger VM (2004). "III turdagi sekretsiya ignalari kompleksini yig'ish bo'yicha tarkibiy tushunchalar". Ilm-fan. 306 (5698): 1040–1042. Bibcode:2004 yil ... 306.1040M. doi:10.1126 / science.1102610. PMC 1459965. PMID 15528446.
- ^ Sani M, Allaoui A, Fusetti F, Oostergetel GT, Keegstra V, Boekema EJ (2007). "Shigella flexneri III tipli sekretsiya apparati igna kompleksining tarkibiy tuzilishi" (PDF). Mikron. 38 (3): 291–301. doi:10.1016 / j.micron.2006.04.007. PMID 16920362.
- ^ Hodkinson JL, Xorsli A, Stabat D, Simon M, Jonson S, da Fonseca kompyuter, Morris E.P., Wall JS, Lea SM, Blocker AJ (2009). "Shigella T3SS transmembran mintaqalarini uch o'lchovli rekonstruktsiya qilish davomida 12 barobar simmetriya va yangi xususiyatlarni ochib beradi". Nat Struct Mol Biol. 16 (5): 477–485. doi:10.1038 / nsmb.1599. PMC 2681179. PMID 19396171.
- ^ Chjan, L; Vang, Y; Picking, WL; Picking, WD; De Guzman, RN (2006 yil 2-iyun). "Burkholderia pseudomallei ning III turdagi sekretsiya ignasi oqsili monomerik BsaL eritmasining tuzilishi". Molekulyar biologiya jurnali. 359 (2): 322–30. doi:10.1016 / j.jmb.2006.03.028. PMID 16631790.
- ^ Deane JE, Roversi P, Kordes FS, Jonson S, Kenjale R, Daniell S, Booy F, Pick WD, Picking WL, Blocker AJ, Lea SM (2006). "III turdagi sekretsiya tizimi ignasining molekulyar modeli: mezbon hujayralarni sezish uchun ta'siri". Proc Natl Acad Sci AQSh. 103 (33): 12529–12533. Bibcode:2006 yil PNAS..10312529D. doi:10.1073 / pnas.0602689103. PMC 1567912. PMID 16888041.
- ^ Galkin VE; Schmied WH; Schraidt O; Marlovits TC va Egelman (2010). "Salmonella typhimurium III sekretsiya tizimi ignasi tuzilishi flagellar tizimidan ajralib chiqishini ko'rsatadi". J Mol Biol. 396 (5): 1392–1397. doi:10.1016 / j.jmb.2010.01.001. PMC 2823972. PMID 20060835.
- ^ a b Loquet A, Sgourakis NG, Gupta R, Giller K, Riedel D, Goosmann C, Grizinger C, Kolbe M, Beyker D, Becker S, Lange A (2012). "III turdagi sekretsiya tizimi ignasining atomik modeli". Tabiat. 486 (7402): 276–279. Bibcode:2012 yil natur.486..276L. doi:10.1038 / tabiat11079. PMC 3598588. PMID 22699623.
- ^ Xolms, T. C .; May, A. E.; Zaleta-Rivera, K .; Ruby, J. G.; Skyues-Koks, P.; Fishbax, M. A .; Derisi, J. L .; Ivatsuki, M .; Uramura, S .; Xosla, C. (2012). "Gvadinomin biosintezi bo'yicha molekulyar tushunchalar: III turdagi sekretsiya tizimining inhibitori". Amerika Kimyo Jamiyati jurnali. 134 (42): 17797–17806. doi:10.1021 / ja308622d. PMC 3483642. PMID 23030602.
- ^ Theuretzbacher U, Piddock LJ (iyul 2019). "An'anaviy bo'lmagan antibakterial terapiya imkoniyatlari va muammolari". Hujayra xosti va mikrob. 26 (1): 61–72. doi:10.1016 / j.chom.2019.06.004. PMID 31295426.
Qo'shimcha o'qish
- Tez tushuncha dan injektoma kimyosini tasvirlab beradi Qirollik kimyo jamiyati
- Xost-patogenning o'zaro ta'siri yilda Pseudomonas shpritslari pv. pomidor va bakterial dog 'kasalligiga olib keladigan pomidor o'simliklari.