Dengiz viruslari - Marine viruses

| Serialning bir qismi |
| Plankton |
|---|
 |
Dengiz viruslari yashash joylari bilan belgilanadi viruslar topilgan dengiz muhiti, ya'ni sho'r suv dengizlar yoki okeanlar yoki sho'r qirg'oq suvi daryolar. Viruslar kichik yuqumli moddalar faqat mumkin takrorlash tiriklar ichida hujayralar a mezbon organizm, chunki buning uchun ular xostning replikatsiya texnikasiga muhtoj.[4] Ular barcha turlarini yuqtirishlari mumkin hayot shakllari, dan hayvonlar va o'simliklar ga mikroorganizmlar, shu jumladan bakteriyalar va arxey.[5]
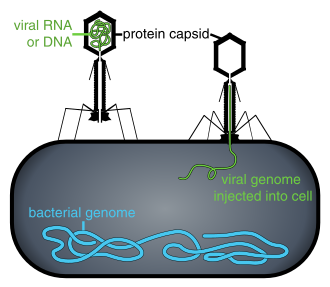
Hujayra ichida bo'lmaganda yoki hujayraga yuqish jarayonida viruslar mustaqil zarrachalar shaklida mavjud virionlar. Virion tarkibiga a kiradi genom (uzoq molekulalar genetik ma'lumotni ikkalasi shaklida olib yuradiganlar DNK yoki RNK ) bilan o'ralgan kapsid (a oqsil genetik materialni himoya qiladigan palto). Ushbu virus zarralarining shakllari oddiydan farq qiladi spiral va ikosahedral ba'zi virus turlari uchun shakllari boshqalari uchun yanada murakkab tuzilmalargacha. Ko'pgina virus turlarida virionlar mavjud bo'lib, ular bilan ko'rish mumkin emas optik mikroskop. O'rtacha virion o'rtacha qiymatning yuzdan bir qismiga teng bakteriya.
Bir choy qoshiq dengiz suvi odatda o'n million dengiz virusini o'z ichiga oladi. Ushbu viruslarning aksariyati bakteriofaglar yuqtiradigan va yo'q qiladigan dengiz bakteriyalari va o'sishini nazorat qilish fitoplankton tagida dengiz oziq-ovqat tarmog'i. Bakteriofaglar o'simliklar va hayvonlar uchun zararsizdir, ammo dengiz ekotizimlarini tartibga solish uchun juda muhimdir. Ular uchun asosiy mexanizmlarni etkazib berish okean uglerodini qayta ishlash va ozuqa moddalari. Deb nomlanuvchi jarayonda virusli shunt, o'lik bakterial hujayralardan ajralib chiqadigan organik molekulalar yangi bakteriyalar va alglarning o'sishini rag'batlantiradi. Xususan, bakteriyalarni viruslar bilan parchalanishi (lizis ) kuchayishi ko'rsatilgan azot aylanish jarayoni va fitoplankton o'sishini rag'batlantirish. Virusli faollik ham ta'sir qiladi biologik nasos, bu jarayon sekvestrlar uglerod chuqur okeanda. Okeanlarda nafas olish miqdorini oshirib, viruslar bilvosita atmosferadagi karbonat angidrid miqdorini 3 ga kamaytirish uchun javobgardir. gigatonnalar yiliga uglerod
Dengiz mikroorganizmlari umumiy miqdorning taxminan 70 foizini tashkil qiladi dengiz biomassasi. Taxminlarga ko'ra, dengiz viruslari har kuni ushbu biomassaning 20 foizini o'ldiradi. Viruslar zararli moddalarni tezda yo'q qilish uchun mas'ul bo'lgan asosiy vositalardir alg gullaydi ko'pincha boshqalarni o'ldiradi dengiz hayoti. Okeanlardagi viruslar soni offshor va suvga chuqurroq tushib boradi, bu erda xo’jayin organizmlari kamroq. Viruslar bu muhim tabiiy vositadir genlarni uzatish ko'payadi turli xil turlari orasida genetik xilma-xillik va evolyutsiyani boshqaradi. Viruslar Yerdagi hayotning so'nggi umumbashariy ajdodlari davrida bakteriyalar, arxeylar va eukariotlar xilma-xilligidan oldin erta evolyutsiyada asosiy rol o'ynagan deb o'ylashadi. Viruslar hanuzgacha Yerdagi o'rganilmagan genetik xilma-xillikning eng yirik sohalaridan biri hisoblanadi.
Fon


Hozirgi kunda viruslar qadimiy va hayotning divergentsiyasini oldindan belgilaydigan kelib chiqishi sifatida tan olingan uchta domen.[6] Ular hayot bor joyda topiladi va, ehtimol, tirik hujayralar paydo bo'lganidan beri mavjud bo'lgan.[7] Viruslarning kelib chiqishi hayotning evolyutsion tarixi noma'lum, chunki ular fotoalbomlarni hosil qilmaydi. Molekulyar texnika viruslarning DNK yoki RNKlarini taqqoslash uchun ishlatiladi va ularning paydo bo'lishini tekshirishning foydali vositasidir.[8] Ba'zi viruslar bo'lishi mumkin rivojlangan dan plazmidlar - hujayralar o'rtasida harakatlanishi mumkin bo'lgan DNK qismlari, boshqalari esa bakteriyalardan rivojlangan bo'lishi mumkin. Evolyutsiyada viruslar muhim vosita hisoblanadi gorizontal genlarning uzatilishi, bu esa ko'payadi genetik xilma-xillik.[9]
Viruslarning shakli ekanligi to'g'risida fikrlar turlicha hayot yoki tirik organizmlar bilan ta'sir o'tkazadigan organik tuzilmalar.[10] Ba'zilar ularni hayot shakli deb hisoblashadi, chunki ular genetik materialni olib yuradilar, o'zlarini yig'ish orqali o'zlarining bir nechta nusxalarini yaratadilar va rivojlanib boradilar. tabiiy selektsiya. Ammo ularning hayotiyligini hisoblash uchun odatda zarur deb hisoblangan uyali tuzilish kabi asosiy xususiyatlar mavjud emas. Viruslar ba'zi bir xususiyatlarga ega, ammo ularning hammasiga ham ega emasligi sababli replikator sifatida ta'rif berilgan[11] va "hayot chekkasidagi organizmlar" sifatida.[12]
Okeandagi viruslar mavjudligi orqali aniqlandi elektron mikroskopi va epifluoresans mikroskopi ekologik suv namunalari va undan keyin metagenomik madaniyatsiz virus namunalaridan namuna olish.[13][14] Dengiz viruslari, garchi mikroskopik bo'lsa ham va umuman e'tiborga olinmaydi olimlar yaqin vaqtgacha okeandagi eng ko'p va xilma-xil biologik mavjudotlardir. Viruslarning taxminiy miqdori 10 ga teng30 okeanda yoki 1 dan 100000x10 gacha6 per mililitr.[4] Dengiz viruslari miqdorini aniqlash dastlab transmissiya elektron mikroskopi yordamida amalga oshirilgan, ammo uning o'rnini epifloresans yoki oqim sitometriyasi.[15]
Bakteriofaglar

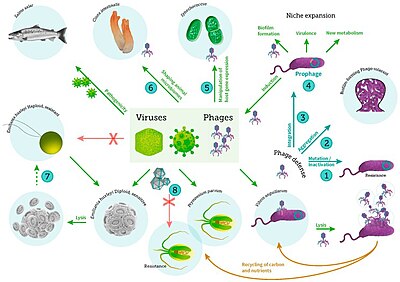
Bakteriofaglar, ko'pincha shunchaki chaqiriladi fajlar, viruslar parazit bakteriyalar. Dengiz fajlari parazit dengiz bakteriyalari kabi siyanobakteriyalar.[16] Ular dengiz muhitida eng ko'p uchraydigan biologik mavjudot bo'lgan turli xil viruslar guruhidir, chunki ularning xostlari, bakteriyalari odatda dengizdagi hujayralar soni bo'yicha dominant hisoblanadi. Okeanlarda bakteriyalarga qaraganda o'n baravar ko'p faglar mavjud,[17] dengiz suvining mililitri uchun 250 million bakteriofag darajasiga etish.[18] Ushbu viruslar bog'lanish orqali o'ziga xos bakteriyalarni yuqtiradi sirt retseptorlari molekulalari va keyin katakka kirish. Qisqa vaqt ichida, ba'zi hollarda bir necha daqiqa, bakterial polimeraza virusli mRNKni oqsilga o'tkazishni boshlaydi. Ushbu oqsillar hujayra ichidagi yangi virionlar, yangi virionlarni yig'ilishiga yordam beradigan yordamchi oqsillar yoki hujayra lizisida ishtirok etadigan oqsillarga aylanadi. Virusli fermentlar hujayra membranasining parchalanishiga yordam beradi va in'ektsiyadan yigirma daqiqa o'tgach, uch yuz fagani takrorlashi mumkin bo'lgan faglar mavjud.[19]
Bakteriyalar begona DNKni yo'q qiladigan fermentlarni ishlab chiqarish orqali o'zlarini bakteriofaglardan himoya qiladi. Ushbu fermentlar cheklash endonukleazalari, bakteriofaglar bakterial hujayralarga kiritadigan virusli DNKni kesib tashlang.[20] Bakteriyalarda foydalanadigan tizim ham mavjud CRISPR o'tmishda bakteriyalar bilan aloqa qilgan viruslar genomlari parchalarini saqlab qolish uchun ketma-ketliklar, bu ularga virusning ko'payishini blokirovka qilishga imkon beradi RNK aralashuvi.[21][22] Ushbu genetik tizim bakteriyalarni ta'minlaydi erishilgan immunitet infektsiyaga.[23]
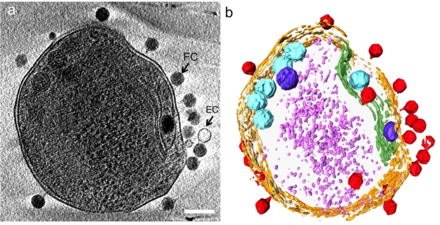
b) hujayra devorini to'q sariq rangda, plazma membranasini och sariq rangda, tilakoid membranani yashil rangda, karboksizomalarni siyan rangda, polifosfat tanasini ko'k rangda, hujayraning yon tomonlarida yoki tepasida adsorbsiyalangan faglarni qizil rangda, va och binafsha rangdagi sitoplazmatik granulalar (ehtimol asosan ribosomalar).[24]

dengiz bilan o'zaro aloqada bo'lish Proxlorokokk MED4 bakteriyasi
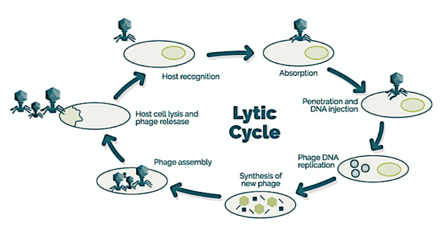
→ biriktirma: faj xujayraning yuzasiga yopishadi
→ penetratsiya: fag DNKni hujayra membranasi orqali yuboradi
→ transkripsiya: xujayraning DNKsi parchalanib, hujayraning metabolizmi
fag biosintezini boshlashga yo'naltirilgan
→ biosintez: fag DNKsi hujayra ichida takrorlanadi
→ pishib etish: takrorlangan material to'liq shakllangan virusli faglarga yig'iladi
→ lizis: yangi hosil bo'lgan fajlar yuqtirilgan hujayradan ajralib chiqadi
(bu jarayonda o'zi yo'q qilinadi) yangi xost hujayralarini izlash uchun [25]
Mikroblar Yer ekotizimlarini qo'llab-quvvatlovchi ozuqaviy o'zgarishlarni boshqaradi,[26] va bu mikroblarni yuqtiradigan viruslar mikroblar sonining ko'payishini va xilma-xilligini modulyatsiya qiladi.[27][24] The siyanobakteriya Proxlorokokk, eng ko'p kislorodli fototrof Yer yuzida uglerod ishlab chiqarishning ulkan ulushiga hissa qo'shadi va ko'pincha mililitrda 100000 hujayradan ko'proq zichlikka etadi. oligotrofik va mo''tadil okeanlar.[28] Demak, virusli (siyanofag) infektsiya va lizis Proxlorokokk global muhim tarkibiy qism hisoblanadi uglerod aylanishi. Siyanofaglar mezbonlarning o'limini keltirib chiqarishda ekologik rolidan tashqari, genlarni, shu jumladan yadro fotosintez genlarini birgalikda tanlash va almashish orqali o'z egalarining metabolizmiga va evolyutsiyasiga ta'sir qiladi.[24]


Uzoq muddatga, dumli faglar buyurtmaning Caudovirales organizmlarning soni va xilma-xilligi bo'yicha dengiz ekotizimlarida hukmronlik qilgandek edi.[16] Biroq, yaqinda o'tkazilgan tadqiqotlar natijasida quyruqsiz viruslar ko'plab chuqurliklarda va okean mintaqalarida hukmronlik qilmoqda.[29] Ushbu dumsiz faglar dengiz bakteriyalariga ham zarar etkazadi va oilalarni o'z ichiga oladi Kortikoviridae,[30] Inoviridae,[31] Microviridae [32] va Avtolykiviridae.[33][34][35][36]
Archaeal viruslar
Arxey viruslari ichida ko'payadi arxey: bu g'ayrioddiy va ba'zan o'ziga xos shakllarga ega bo'lgan ikki simli DNK viruslari.[37][38] Ushbu viruslar batafsil o'rganilgan termofil arxeylar, ayniqsa buyurtmalar Sulfoloballar va Termoproteallar.[39] Ushbu viruslardan himoya qilish RNK aralashuvini o'z ichiga oladi takrorlanadigan DNK arxe genomlari tarkibidagi viruslar genlari bilan bog'liq bo'lgan ketma-ketliklar.[40][41] Ko'pgina arxeylarda viruslardan himoya qiluvchi CRISPR-Cas tizimlari mavjud. Bular arxeylarga virusli DNK qismlarini saqlab qolish imkoniyatini beradi, keyinchalik ular RNK aralashuviga o'xshash jarayon yordamida virus tomonidan keyingi infektsiyalarni nishonga olish va yo'q qilish uchun ishlatiladi.[42]
Qo'ziqorin viruslari
Mikoviruslar, shuningdek, mikofaglar deb nomlanuvchi, yuqtiradigan viruslardir qo'ziqorinlar. Qo'ziqorin hujayralarining infektsiyasi hayvon hujayralaridan farq qiladi. Qo'ziqorinlarda xitindan yasalgan qattiq hujayra devori bor, shuning uchun ko'pchilik viruslar hujayra devoridagi shikastlanishdan keyingina bu hujayralarga kirib borishi mumkin.[43]
- Qarang; Nerva, L .; Ciuffo, M .; Vallino, M .; Margariya, P .; Varese, G.C .; Gnavi, G .; Turina, M. (2016). "Dengiz qo'ziqorinlari to'plamidan virusli va plazmidli simbiontlarni aniqlash va tavsiflash uchun bir nechta yondashuvlar". Viruslarni o'rganish. 219: 22–38. doi:10.1016 / j.virusres.2015.10.028. hdl:2318/1527617. PMID 26546154.
Eukaryot viruslari

Dengiz protistlari
2015 yilga kelib, taxminan 40 ta virus ta'sir ko'rsatmoqda dengiz protistlari ajratilgan va tekshirilgan, ularning aksariyati mikro suv o'tlari viruslari.[44] Ushbu dengiz protisti viruslarining genomlari juda xilma-xildir.[45][46] Dengiz yosunlari oiladagi viruslar bilan yuqishi mumkin Fitodnaviridae. Ular katta (100-560 kb) ikki zanjirli DNK viruslari bilan ikosahedral shaklidagi kapsidlar. 2014 yilga kelib, oilada oltita naslga bo'lingan 33 tur aniqlandi,[47][48] deb nomlanuvchi super viruslar guruhiga kiradi nukleotsitoplazmatik yirik DNK viruslari. Dalillar 2014 yilda chop etilgan bo'lib, ba'zi bir turlarni taklif qilmoqda Fitodnaviridae ilgari ishonilganidek, nafaqat alg turlaridan ko'ra odamlarga yuqishi mumkin.[49] Ushbu oilaga mansub ko'pgina nasllar hujayra retseptorlari orqali mezbon hujayraga kiradi endotsitoz va yadroda takrorlanadi.

Fitodnaviridae ularning gidroksidi xujayralarining o'sishi va unumdorligini tartibga solish orqali muhim ekologik rollarni o'ynaydi. Algal turlari Heterosigma akashiwo va tur Xrizoxromulina Baliqchilikka zarar etkazishi mumkin bo'lgan zich gullarni hosil qilishi mumkin, natijada akvakultura sanoatida yo'qotishlarga olib keladi.[50] Heterosigma akashiwo virusi (HaV) mikrobial vosita sifatida ushbu alg turlari tomonidan ishlab chiqarilgan toksik qizil oqimlarning takrorlanishini oldini olish uchun foydalanish uchun tavsiya etilgan.[51] The koksolitovirus Emiliania huxleyi virusi 86, ulkan ikki qatorli DNK virusi, hamma joyda yuqadi koksolitofora Emiliania huxleyi.[47][48] Ushbu virus dengiz viruslari orasida ma'lum bo'lgan eng katta genomlardan biriga ega.[52] Fitodnaviridae chuchuk suvlar va dengiz alglari turlarining o'limiga va lizisiga olib keladi, suvda organik uglerod, azot va fosforni ajratib, mikroblar tsikli uchun oziq moddalar beradi.[53]
Virus-prokaryot nisbati, VPR, ko'pincha viruslar va xostlar o'rtasidagi munosabatlarning ko'rsatkichi sifatida ishlatiladi. Tadqiqotlar VPRni dengiz mikroblarining mahsuldorligi, o'limi va biogeokimyoviy velosipedda virus ta'sirini bilvosita aniqlash uchun ishlatgan.[54] Biroq, ushbu taxminlarni amalga oshirishda olimlar VPRni 10: 1 deb hisoblashadi, median VPRni er usti okeanida kuzatgan.[54][17] Haqiqiy VPR joylashuvga qarab juda farq qiladi, shuning uchun VPR virusli faollik yoki mo'l-ko'llik uchun to'g'ri proksi bo'lmasligi mumkin, chunki u davolangan.[54][55]
Dengiz umurtqasizlar
bakteriyalar, fitoplankton va baliqlarning virusli infektsiyasi[56]
Dengiz umurtqasizlar virusli kasalliklarga moyil.[57][58][59] Dengiz yulduzi kasallikni isrof qilmoqda ning kasalligi dengiz yulduzi va vaqti-vaqti bilan paydo bo'ladigan, ta'sirlanganlarning ommaviy o'limiga olib keladigan boshqa bir qancha echinodermalar.[60] Ushbu kasallikka chalingan dengiz yulduzlarining 40 ga yaqin turlari mavjud. 2014 yilda ushbu kasallik hozirda "DNK virusi" deb nomlanuvchi bir qatorli DNK virusi bilan bog'liq deb taxmin qilingan edi dengiz yulduzi bilan bog'liq bo'lgan densovirus (SSaDV); ammo, dengiz yulduzi isrof kasalligi to'liq tushunilmagan.[61]
Dengiz umurtqali hayvonlari
Baliq ayniqsa infektsiyaga moyil rabdoviruslar, ular alohida, ammo quturish virusi bilan bog'liq. Rabdovirusning kamida to'qqiz turi turlar, jumladan losos, pike, perch, levrek, karp va cod kabi iqtisodiy ahamiyatga ega kasalliklarni keltirib chiqaradi. Semptomlar orasida anemiya, qon ketish, sustlik va suvning harorati ta'sir qiladigan o'lim darajasi mavjud. Kuluçkalıklarda kasalliklar ko'pincha haroratni 15-18 ° S ga ko'tarish orqali nazorat qilinadi.[62]:442–443 Barcha umurtqali hayvonlar singari, baliqlar ham azoblanadi herpes viruslari. Ushbu qadimiy viruslar o'zlarining xostlari bilan birgalikda rivojlangan va turlarga juda xosdir.[62]:324 Baliqda ular saraton o'smalari va saraton bo'lmagan o'smalarga sabab bo'ladi giperplaziya.[62]:325
1984 yilda, yuqumli losos anemiyasi (ISAv) Norvegiyada topilgan Atlantika lososlari inkubatsiya. Kasallikdagi baliqlarning 80 foizi nobud bo'ldi. Virusli kasallik bo'lgan ISAv hozirgi kunda Atlantika lososida dehqonchilikning hayotiy hayoti uchun katta xavf tug'dirmoqda.[63] Nomidan ko'rinib turibdiki, bu og'irlikni keltirib chiqaradi anemiya yuqtirgan baliq. Baliqning qizil qon hujayralari sutemizuvchilardan farqli o'laroq, DNKga ega va virus bilan yuqishi mumkin. Boshqarish strategiyasiga vaktsina ishlab chiqish va kasallikka genetik qarshilikni yaxshilash kiradi.[64]
Dengiz sutemizuvchilar dengiz virusli infektsiyalariga ham sezgir. 1988 va 2002 yillarda minglab port muhrlari tomonidan Evropada o'ldirilgan phocine distemper virusi.[65] Boshqa ko'plab viruslar, shu jumladan kalitsiviruslar, herpes viruslari, adenoviruslar va parvoviruslar, dengiz sutemizuvchilar populyatsiyasida tarqaladi.[66]
Gigant dengiz viruslari
Ko'pgina viruslarning uzunligi taxminan 20 dan 300 nanometrgacha. Bunga bakteriyalarning uzunligi qarama-qarshi bo'lishi mumkin, bu taxminan 400 nanometrdan boshlanadi. Shuningdek, bor ulkan viruslar, tez-tez chaqiriladi giruslar, odatda uzunligi taxminan 1000 nanometr (bir mikron). Barcha ulkan viruslar filum Nukleotsitoviriko (NCLDV), bilan birgalikda poxviruslar.Ulardan eng kattasi ma'lum Tupanvirus. Gigant virusning bu turi 2018 yilda okean tubida, shuningdek sodali ko'lda topilgan va umumiy uzunligi 2,3 mikrongacha yetishi mumkin.[67]

 Gigant mimivirus
Gigant mimivirus
Kriyo-elektron mikrograf ning CroV ulkan virus [68]
masshtab bar = 0.2µm


Gigant viruslarning kashf etilishi va keyinchalik tavsiflanishi ularning evolyutsion kelib chiqishi bilan bog'liq munozaralarni keltirib chiqardi. Ularning kelib chiqishi uchun ikkita asosiy gipoteza: ular mayda viruslardan, xujayrali organizmlardan DNKni yig'ib olishdan yoki juda murakkab organizmlardan ko'payish uchun o'zini o'zi etarli bo'lmagan hozirgi shaklga aylanganidan iborat.[69] Gigant viruslar qanday murakkab organizmdan ajralib chiqishi mumkinligi ham munozara mavzusi. Bitta taklif, kelib chiqish nuqtasi aslida to'rtinchisini anglatadi domen hayot,[70][71] ammo bu juda arzonlashtirildi.[72][73]
Virofaglar
Viruslar ga tayanadigan kichik, ikki zanjirli DNK viruslari birgalikda infektsiya ning ulkan viruslar. Virofaglar o'zlarining ko'payishi uchun birgalikda yuqadigan ulkan virusning virusli replikatsiya zavodiga tayanadi. Virofaglarning xususiyatlaridan biri shundaki, ularda a parazit birgalikda yuqadigan virus bilan munosabatlar. Ularning ulkan virusga replikatsiya uchun bog'liqligi ko'pincha ulkan viruslarning zararsizlanishiga olib keladi. Virofagiya mezbon organizmning tiklanishi va omon qolishini yaxshilashi mumkin. Boshqalardan farqli o'laroq sun'iy yo'ldosh viruslari, virofaglar a parazit ularning birgalikda yuqadigan virusiga ta'siri. Virofaglar ulkan virusni harakatsiz holga keltirishi va shu bilan mezbon organizmning holatini yaxshilashi kuzatilgan.
Barcha ma'lum bo'lgan virusofaglar oilaga birlashtirilgan Lavidaviridae ("katta virusga bog'liq yoki bog'liq" dan + -viridae ).[74] Birinchi virusofag a sovutish minorasi 2008 yilda Frantsiyaning Parij shahrida. U birgalikda yuqtirgan ulkan virus bilan topilgan, Acanthamoeba castellanii mamavirus (ACMV). Virofagga nom berildi Sputnik va uning replikatsiyasi butunlay ACMV va uning sitoplazmik replikatsiya mexanizmining birgalikda infektsiyasiga bog'liq edi. Sputnik shuningdek, ACMVga inhibitiv ta'sir ko'rsatishi va mezbonning hayotini yaxshilashi aniqlandi. Boshqa xarakterli virusofaglarga Sputnik 2, Sputnik 3, Zamilon va Mavirus.[75][76]
Ushbu virusofaglarning aksariyati tahlil qilish orqali topilgan metagenomik ma'lumotlar to'plamlari. Metagenomik tahlilda DNK ketma-ketliklari bir nechta bioinformatik algoritmlar orqali boshqariladi, ular ma'lum muhim naqsh va xususiyatlarni keltirib chiqaradi. Ushbu ma'lumotlar to'plamida ulkan viruslar va viruslar mavjud. Ular 17 dan 20 gacha bo'lgan ketma-ketliklarni qidirish bilan ajralib turadikbp allaqachon berilgan virusofaglarga o'xshashligi bor. Ushbu virofaglar chiziqli yoki dumaloq ikki simli DNK genomlariga ega bo'lishi mumkin.[77] Madaniyatdagi virusofaglar 40 dan 80 nanometrgacha bo'lgan ikosaedral kapsid zarralariga ega.[78] Virofag zarralari shu qadar kichikki, bu zarralarni ko'rish uchun elektron mikroskopiyadan foydalanish kerak. Metagenomik ketma-ketlikka asoslangan tahlillar taxminan 57 to'liq va qisman virusli genomlarni taxmin qilish uchun ishlatilgan[79] va 2019 yil dekabr oyida turli xil yashash joylaridan 328 ta yuqori sifatli (to'liq yoki deyarli to'liq) genomlarni aniqlash, shu jumladan odamning ichaklari, o'simlik rizosferasi va er osti qatlamlari, 27 ta aniq taksonomik panellardan.[80]



Gigant dengiz virusi CroV yuqtiradi va o'limga olib keladi lizis dengiz zooflagellatining Roenbergensis kafeteryasi.[82] Bu qirg'oq ekologiyasiga ta'sir qiladi, chunki Roenbergensis kafeteryasi suvda uchraydigan bakteriyalar bilan oziqlanadi. Qachon kam sonlar Roenbergensis kafeteryasi keng CroV infektsiyalari tufayli bakterial populyatsiyalar tez o'sib boradi.[83] Ta'siri CroV ning tabiiy populyatsiyalari bo'yicha C. roenbergensis noma'lum bo'lib qolmoqda; ammo, virus juda mezbonga xos ekanligi aniqlandi va boshqa yaqin organizmlarni yuqtirmaydi.[84] Cafeteria roenbergensis, shuningdek, ikkinchi virus bilan yuqtiriladi Mavirus virusi, CroV bilan birgalikda infektsiya paytida.[75] Ushbu virus CroV replikatsiyasiga xalaqit beradi, bu esa omon qolishiga olib keladi C. roenbergensis hujayralar. Mavirus hujayralarining genomiga qo'shilishga qodir C. roenbergensis va shu bilan aholiga immunitet beradi.[76]
Dengiz viruslarining roli
Dengiz viruslari yaqinda juda ko'p o'rganilgan bo'lsa-da, ular allaqachon ko'plab ekotizim funktsiyalari va tsikllarida muhim rol o'ynashi ma'lum. Dengiz viruslari bir qator muhim narsalarni taklif qiladi ekotizim xizmatlari va dengiz ekotizimlarini tartibga solish uchun juda muhimdir.[3] Dengiz bakteriofaglari va boshqa viruslar ta'sir qiladi biogeokimyoviy tsikllar global, ta'minlash va tartibga solish mikroblarning biologik xilma-xilligi, tsikli uglerod dengiz orqali oziq-ovqat tarmoqlari bakteriyalarni oldini olishda juda muhimdir aholi portlashlari.[85]
Virusli shunt

Okeandagi viruslarning dominant egalari dengiz mikroorganizmlari, masalan, bakteriyalardir.[13] Bakteriofaglar o'simliklar va hayvonlar uchun zararsiz bo'lib, dengiz va chuchuk suv ekotizimlarini boshqarishda juda muhimdir.[87] o'limning muhim agentlari hisoblanadi fitoplankton, ning asosi Oziq ovqat zanjiri suv muhitida.[88] Ular suv mikroblari jamoalarida bakteriyalarni yuqtiradi va yo'q qiladi va ularning muhim mexanizmlaridan biridir uglerodni qayta ishlash va dengiz muhitida ozuqa moddalarining aylanishi. O'lik bakterial hujayralardan ajralib chiqadigan organik molekulalar yangi deb ataladigan jarayonda yangi bakterial va alg o'sishini rag'batlantiradi virusli shunt.[89]
Shu tarzda dengiz viruslari samaradorligini oshirish orqali ozuqa tsikllarida muhim rol o'ynaydi biologik nasos. Viruslar sabab bo'ladi lizis tirik hujayralar, ya'ni hujayra membranalarini parchalaydi. Bu kabi birikmalarni chiqaradi aminokislotalar va nuklein kislotalar, ular sirt yaqinida qayta ishlashga moyil.
Virusli faollik biologik nasosning qobiliyatini ham oshiradi ajratuvchi uglerod chuqur okeanda.[66] Lizis hujayra devorlarida mavjud bo'lgan ko'proq hazm bo'lmaydigan uglerodga boy moddalarni chiqaradi, ular chuqur suvlarga eksport qilinadi. Shunday qilib, virusli shunt bilan chuqur suvlarga eksport qilinadigan material, ehtimol u olingan materialga qaraganda ko'proq uglerodga boy.[90][91] Okeanlarda nafas olish miqdorini oshirib, viruslar bilvosita atmosferadagi karbonat angidrid miqdorini uchga kamaytirish uchun javobgardir. gigatonnalar yiliga uglerod[66] Bakteriyalarni viruslar bilan parchalanishi azot aylanishini kuchaytiradi va fitoplankton o'sishini rag'batlantiradi.[92]
The virusli shunt yo'l - bu to'sqinlik qiladigan mexanizm (prokaryotik va ökaryotik ) dengiz mikroblari zarracha bo'lgan organik moddalar (POM) yuqoriga ko'chishdan trofik sathlar ularni qayta ishlash orqali erigan organik moddalar (DOM), bu mikroorganizmlar tomonidan osonlikcha olinishi mumkin. Virusli manevr, mikroblar ekotizimida xilma-xillikni saqlashga yordam beradi, dengiz mikroblarining bir turini mikro muhitda hukmron bo'lishiga yo'l qo'ymaslik.[93] Virusli shunt yo'li bilan qayta ishlangan DOM dengiz DOM ning boshqa asosiy manbalari tomonidan ishlab chiqarilgan miqdor bilan taqqoslanadi.[94]


Viruslar dengiz muhitida eng ko'p tarqalgan biologik mavjudotdir[5] O'rtacha dengiz suvining bir choy qoshig'ida ularning soni o'n millionga yaqin.[96] Ushbu viruslarning aksariyati bakteriofaglar heterotrof bakteriyalarni yuqtirish va siyanofaglar siyanobakteriyalarni yuqtirish. Viruslar mikroblarga nisbatan nisbiy ko'pligi sababli mikrobial tsikldagi mikroorganizmlarni osonlikcha yuqtiradi.[97] Prokaryotik va eukaryotik o'lim uglerod ozuqalarini qayta ishlashga yordam beradi hujayra lizisi. Azotning (xususan, ammoniyning) qayta tiklanishiga oid dalillar mavjud. Ushbu ozuqaviy moddalarni qayta ishlash mikroblarning ko'payishini rag'batlantiradi.[98] Jahon okeanidagi fitoplanktondan olinadigan birlamchi ishlab chiqarishning 25% virusni manevr qilish orqali mikrobial tsiklda qayta ishlanishi mumkin.[99]
Yosunlarning gullashini cheklash
Mikroorganizmlar dengiz biomassasining taxminan 70 foizini tashkil qiladi.[100] Taxminlarga ko'ra, viruslar har kuni ushbu biomassaning 20 foizini o'ldiradi va okeanlarda viruslar bakteriyalar va arxeylardan 15 baravar ko'pdir. Viruslar zararli moddalarni tezda yo'q qilish uchun mas'ul bo'lgan asosiy vositalardir alg gullaydi,[101] ko'pincha boshqa dengiz hayotini o'ldiradi.[102] Olimlar dengiz salohiyatini o'rganmoqdalar siyanofaglar oldini olish yoki orqaga qaytarish uchun ishlatilishi kerak evrofikatsiya. Okeanlardagi viruslar soni offshor va suvga chuqurroq tushib boradi, bu erda xo’jayin organizmlari kamroq.[103]
Genlarning uzatilishi
Dengiz bakteriofaglari ko'pincha o'z ichiga oladi yordamchi metabolik genlar, xostdan kelib chiqqan genlar, virusli infeksiya paytida xujayraning metabolizmini to'ldirish orqali viruslarning ko'payishini qo'llab-quvvatlaydi.[104] Ushbu genlar uglerod, fosfor, oltingugurt va azot kabi ko'plab biogeokimyoviy tsikllarga ta'sir qilishi mumkin.[105][106][107][108]
Viruslar bu muhim tabiiy vositadir genlarni uzatish ko'payadi turli xil turlari orasida genetik xilma-xillik va evolyutsiyani boshqaradi.[9] Viruslar, evolyutsiyada bakteriyalar, arxeylar va eukariotlar xilma-xil bo'lishidan oldin, erta evolyutsiyada asosiy rol o'ynagan deb o'ylashadi. so'nggi universal umumiy ajdod Yerdagi hayot.[109] Viruslar hali ham Yerdagi o'rganilmagan genetik xilma-xillikning eng katta suv omborlaridan biri hisoblanadi.[103]
Dengiz yashash joylari
Sohil bo'ylab
Dengiz sohilidagi yashash joylari quruqlik va okean o'rtasida joylashgan. Ehtimol, bu RNK viruslari ushbu muhitda muhim rol o'ynaydi.[110]
Okean sathida

Dengiz sathidagi yashash joylari atmosfera va okean o'rtasida joylashgan. The biofilmga o'xshash okean sathidagi yashash joylari odatda yashaydigan mikroorganizmlarni yashaydi neuston. Mikro qatlamdagi viruslar, deyiladi virioneuston, yaqinda tadqiqotchilar tomonidan potentsial muhim ekologik ta'sirga ega bo'lgan chegara sirt qatlamlarida sirli biologik mavjudotlar sifatida qiziqish uyg'otdi. Ushbu ulkan havo-suv interfeysi global sirt maydonining 70 foizidan ko'prog'ini o'z ichiga olgan asosiy havo-suv almashinuvi jarayonlari kesishmasida joylashganligini hisobga olsak, bu chuqur ta'sir ko'rsatishi mumkin. dengiz biogeokimyoviy tsikllari, ustida mikrobial tsikl va gaz almashinuvi, shuningdek dengiz oziq-ovqat tarmog'i tuzilishi, dengiz sathidagi mikro qatlamdan kelib chiqqan havodagi viruslarning global tarqalishi va inson salomatligi.[111]
Suv ustunida
Dengiz virusi faoliyati potentsial tushuntirishni taqdim etadi plankton paradoksi tomonidan taklif qilingan Jorj Xatchinson 1961 yilda.[112] Plankton paradoksi shundaki, cheklangan resurslar yaratishi kerak bo'lgan okeanning kichik mintaqalarida ko'plab plankton turlari aniqlangan. raqobatdosh chiqarib tashlash, birgalikda yashaydigan turlarning sonini cheklash.[112] Bunda dengiz viruslari rol o'ynashi mumkin, chunki xostlar bilan potentsial aloqa kuchayishi bilan virusli infektsiya kuchayadi.[4] Shuning uchun viruslar juda ko'p o'sadigan plankton turlarining populyatsiyasini boshqarishi mumkin va bu turlarning xilma-xilligi bilan birga yashashga imkon beradi.[4]
Cho'kindilarda
Dengiz bakteriofaglari muhim rol o'ynaydi chuqur dengiz ekotizimlar. 5x10 gacha12 va 1x1013 chuqur dengiz cho'kindilaridagi kvadrat metrga faglar va ularning ko'pligi ularning soni bilan chambarchas bog'liq prokaryotlar cho'kindi jinslardan topilgan. Cho'kindilarda topilgan prokaryotlarning 80% o'limiga ular javob beradi va bu o'limlarning deyarli barchasi hujayradan kelib chiqadi lizis (yorilish). Bu tirik hujayralardagi azot, uglerod va fosforni erigan organik moddalarga va detritga aylantirishga imkon beradi va bu dengizning chuqur cho'kindilarida ozuqa moddalarining aylanish tezligini oshiradi. Biogeokimyoviy tsikllarda chuqur dengiz cho'kindilarining ahamiyati katta bo'lgani uchun dengiz bakteriofaglari ta'sir ko'rsatadi uglerod, azot va fosfor tsikllari. Ushbu ta'sirlarni aniqroq aniqlash uchun ko'proq tadqiqotlar o'tkazish kerak.[113]
Gidrotermal teshiklarda
Viruslar qismidir gidrotermal shamollatish mikroblar jamiyati va ularning ushbu ekotizimlardagi mikrob ekologiyasiga ta'siri - bu rivojlanayotgan tadqiqot sohasi.[114] Viruslar okeandagi eng keng tarqalgan hayot bo'lib, genetik xilma-xillikning eng katta suv omboriga ega.[101] Ularning infektsiyalari ko'pincha o'limga olib kelishi sababli, ular o'limning muhim manbasini tashkil qiladi va shu bilan biologik okeanografik jarayonlarga keng ta'sir ko'rsatadi, evolyutsiya va biogeokimyoviy velosiped okean ichida.[103] Shamollatish muhitida joylashgan viruslar ko'proq qabul qilinganligini ko'rsatadigan dalillar topildi mututeristik dan parazit ular mavjud bo'lgan o'ta va o'zgaruvchan muhitda omon qolish uchun evolyutsion strategiya.[115] Chuqur dengizdagi gidrotermal shamollatishlarda ko'plab viruslar borligi aniqlandi, bu yuqori virusli ishlab chiqarishni ko'rsatmoqda.[116] Boshqa dengiz muhitida bo'lgani kabi, chuqur dengiz gidrotermal viruslari ko'pligi va xilma-xilligiga ta'sir qiladi prokaryotlar va shuning uchun mikrob biogeokimyoviy aylanishiga ta'sir qiladi lizing takrorlash uchun ularning xostlari.[117] Biroq, ularning o'limi va populyatsiyani boshqarish manbai rolidan farqli o'laroq, viruslar genetik ma'lumotlarning suv omborlari vazifasini bajaradigan prokaryotlarning ekstremal muhitda omon qolishini kuchaytirish uchun ham joylashtirilgan. Shuning uchun atrofdagi stresslar ta'sirida virus atmosferasining mikroorganizmlar bilan o'zaro ta'siri mikroorganizmlarning yashashi uchun xujayrali genlarning tarqalishi orqali yordam beradi deb o'ylashadi. gorizontal genlarning uzatilishi.[118]
Qutbiy mintaqalar
Tarqatish
Viruslar juda xos xususiyatga ega.[119] Dengiz virusi virusning yashaydigan o'sha mintaqasida yashovchi organizmlarni yuqtirish ehtimoli ko'proq.[120] Shuning uchun, biogeografiya virionni yuqtirish qobiliyatining muhim omilidir.
Spatiotemporal va boshqa atrof-muhit gradiyentlari bo'yicha virusli populyatsiyalardagi bu o'zgarishni bilish virusli morfologiya bilan tasdiqlanadi, bu elektron elektron mikroskopi (TEM) bilan belgilanadi. Quyruqsiz viruslar ko'plab chuqurliklarda va okean mintaqalarida dominant bo'lib ko'rinadi, keyin esa Caudovirales myoviruslar, podoviruslar va sifoviruslar.[29] Biroq, oilalarga tegishli viruslar Kortikoviridae,[121] Inoviridae[122] va Microviridae[123] shuningdek, turli xil dengiz bakteriyalarini yuqtirishi ma'lum. Metagenomik dalillar shuni ko'rsatadiki, mikroviruslar (ikosahedral ssDNA faglari) ayniqsa dengiz yashash joylarida keng tarqalgan.[123]
Viruslarning xilma-xilligini baholash uchun metagenomik yondashuvlar ko'pincha mos yozuvlar ketma-ketligining etishmasligi bilan cheklanadi va ko'plab ketma-ketliklar izohsiz qoldiriladi.[124] Shu bilan birga, virusli birikmalar, odatda dengiz suvi namunasini 0,02 um filtrlashdan so'ng hosil bo'lgan virusli fraktsiyani to'g'ridan-to'g'ri ketma-ketligi yoki mikrob metagenomidan virusli birikmalar yoki virusli genomlarni aniqlash uchun bioinformatik yondashuvlar orqali hosil bo'ladi. Virusitor kabi taxminiy virusli birikmalarni aniqlash uchun yangi vositalar[125] va VirFinder,[126] virus bakteriyalarining ko'pligi, ularning joylashuvi va funktsional tarkibini baholashga imkon beradi.[127][128]
Shuningdek qarang
Adabiyotlar
- ^ Bonneyn, S .; Breitbart, M.; Buck, K.N. (2016). "Ferrojan ot gipotezasi: okeandagi temir-virus o'zaro ta'siri". Dengiz fanidagi chegara. 3: 82. doi:10.3389 / fmars.2016.00082. S2CID 2917222.
- ^ Ushbu mayda organizmlarning chindan ham g'alati shakllari bor National Geogaphic ", 2016 yil 12-noyabr.
- ^ a b Shors T (2008). Viruslarni tushunish. Jons va Bartlett nashriyotlari. p. 5. ISBN 978-0-7637-2932-5.
- ^ a b v d Brussaard CP, Baudoux A, Rodriges-Valera F (2016). Stal LJ, Kretoiu MS (tahr.). Dengiz viruslari. Dengiz mikrobiomi. Springer xalqaro nashriyoti. 155-183 betlar. doi:10.1007/978-3-319-33000-6_5. ISBN 9783319329987.
- ^ a b v d Koonin EV, Senkevich TG, Dolja VV (2006). "Qadimgi viruslar dunyosi va hujayralar evolyutsiyasi". Biologiya to'g'ridan-to'g'ri. 1: 29. doi:10.1186/1745-6150-1-29. PMC 1594570. PMID 16984643.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
 Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 2.0 xalqaro litsenziyasi.
Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 2.0 xalqaro litsenziyasi. - ^ Mahy WJ va Van Regenmortel MHV (tahrir). Umumiy virusologiya ish stoli entsiklopediyasi. Oksford: Academic Press; 2009 yil. ISBN 0-12-375146-2. p. 28.
- ^ Iyer LM, Balaji S, Koonin EV, Aravind L (2006). "Nukleo-sitoplazmatik yirik DNK viruslarining evolyutsion genomikasi". Viruslarni o'rganish. 117 (1): 156–84. doi:10.1016 / j.virusres.2006.01.009. PMID 16494962.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Sanjuán R, Nebot MR, Chirico N, Mansky LM, Belshaw R (oktyabr 2010). "Virusli mutatsiya darajasi". Virusologiya jurnali. 84 (19): 9733–48. doi:10.1128 / JVI.00694-10. PMC 2937809. PMID 20660197.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ a b Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Bryussov H (2003). "Faj lateral gen transferi agentlari sifatida". Mikrobiologiyaning hozirgi fikri. 6 (4): 417–24. doi:10.1016 / S1369-5274 (03) 00086-9. PMID 12941415.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Koonin EV, Starokadomskyy P (2016). "Viruslar tirikmi? Replikator paradigmasi eski, ammo yanglish savolga hal qiluvchi nur sochadi". Biologik va biotibbiyot fanlari tarixi va falsafasi bo'yicha tadqiqotlar. 59: 125–134. doi:10.1016 / j.shpsc.2016.02.016. PMC 5406846. PMID 26965225.
- ^ Koonin, E. V .; Starokadomskyy, P. (2016 yil 7 mart). "Viruslar tirikmi? Replikator paradigmasi eski, ammo yanglish savolga hal qiluvchi nur sochadi". Biologik va biotibbiyot fanlari tarixi va falsafasi bo'yicha tadqiqotlar. 59: 125–34. doi:10.1016 / j.shpsc.2016.02.016. PMC 5406846. PMID 26965225.
- ^ Rybicki, RaI (1990). "Organizmlarning hayot chegarasida tasnifi yoki viruslar sistematikasi bilan bog'liq muammolar". Janubiy Afrika jurnali. 86: 182–186.
- ^ a b Mann NH (2005 yil may). "Fagning uchinchi asri". PLOS biologiyasi. 3 (5): e182. doi:10.1371 / journal.pbio.0030182. PMC 1110918. PMID 15884981.
- ^ Wommack KE, Hill RT, Myuller TA, Colwell RR (aprel 1996). "Quyosh nurlarining bakteriofagning hayotiyligi va tuzilishiga ta'siri". Amaliy va atrof-muhit mikrobiologiyasi. 62 (4): 1336–41. doi:10.1128 / AEM.62.4.1336-1341.1996. PMC 167899. PMID 8919794.
- ^ Mari D, Brussaard CP, Thyrhaug R, Bratbak G, Vaulot D (yanvar 1999). "Oqim sitometriyasi orqali madaniyatdagi tabiiy viruslar va tabiiy namunalarni sanab chiqish". Amaliy va atrof-muhit mikrobiologiyasi. 65 (1): 45–52. doi:10.1128 / AEM.65.1.45-52.1999. PMC 90981. PMID 9872758.
- ^ a b Mann, NH (2005-05-17). "Fajning uchinchi asri". PLOS biologiyasi. 3 (5): 753–755. doi:10.1371 / journal.pbio.0030182. PMC 1110918. PMID 15884981.
- ^ a b Wommack KE, Colwell RR (2000 yil mart). "Virioplankton: suv ekotizimidagi viruslar". Mikrobiologiya va molekulyar biologiya sharhlari. 64 (1): 69–114. doi:10.1128 / MMBR.64.1.69-114.2000. PMC 98987. PMID 10704475.
- ^ Bergh O, Borsheim KY, Bratbak G, Heldal M (avgust 1989). "Suv muhitida ko'plab viruslar mavjud". Tabiat. 340 (6233): 467–68. Bibcode:1989 yil natur.340..467B. doi:10.1038 / 340467a0. PMID 2755508. S2CID 4271861.
- ^ Shors 595-97 betlar
- ^ Bickle TA, Krüger DH (iyun 1993). "DNKni cheklash biologiyasi". Mikrobiologik sharhlar. 57 (2): 434–50. doi:10.1128 / MMBR.57.2.434-450.1993. PMC 372918. PMID 8336674.
- ^ Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S va boshq. (2007 yil mart). "CRISPR prokaryotlarda viruslarga qarshi qarshilikni ta'minlaydi". Ilm-fan. 315 (5819): 1709–12. Bibcode:2007 yil ... 315.1709B. doi:10.1126 / science.1138140. hdl:20.500.11794/38902. PMID 17379808. S2CID 3888761.
- ^ Brouns SJ, Jore MM, Lundgren M, Westra ER, Slijkhuis RJ, Snijders AP va boshq. (2008 yil avgust). "Kichik CRISPR RNKlari prokaryotlarda virusga qarshi himoya ko'rsatma". Ilm-fan. 321 (5891): 960–64. Bibcode:2008 yil ... 321..960B. doi:10.1126 / science.1159689. PMC 5898235. PMID 18703739.
- ^ Mojica FJ, Rodriguez-Valera F (sentyabr 2016). "Arxeya va bakteriyalarda CRISPRning kashf etilishi". FEBS jurnali. 283 (17): 3162–69. doi:10.1111 / febs.13766. hdl:10045/57676. PMID 27234458.
- ^ a b v d Murata, K .; Chjan, Q .; Galaz-Montoya, J.G .; Fu, C .; Koulman, M.L .; Osburne, M.S .; Shmid, M.F .; Sallivan, MB; Chisholm, SS; Chiu, V. (2017). "P-SSP7 siyanofagining dengizga adsorbsiyasini ingl Proxlorokokk". Ilmiy ma'ruzalar. 7: 44176. Bibcode:2017 yil NatSR ... 744176M. doi:10.1038 / srep44176. PMC 5345008. PMID 28281671.
- ^ Bakteriofaglar qanday ko'payadi? Barselona universiteti. Qabul qilingan 12 iyul 2020 yil.
- ^ Falkovski, PG.; Fenchel, T .; Delong, EF (2008). "Yerning biogeokimyoviy tsikllarini boshqaradigan mikrob dvigatellari". Ilm-fan. 320 (5879): 1034–1039. Bibcode:2008 yil ... 320.1034F. doi:10.1126 / science.1153213. PMID 18497287. S2CID 2844984.
- ^ Brum, JR .; Sallivan, M.B. (2015). "Qiyinchilikka ko'tarilish: kashfiyotning jadal sur'ati dengiz virusologiyasini o'zgartiradi". Tabiat sharhlari Mikrobiologiya. 13 (3): 147–159. doi:10.1038 / nrmicro3404. PMID 25639680. S2CID 32998525.
- ^ Bouman, X.A .; Ulloa, O .; Scanlan, D.J .; Tsvirglmayer, K .; Li, VK.; Platt, T .; Styuart, V .; Barlow, R .; Let, O .; Klementson, L .; Lutz, V. (2006). "Ning global sirt taqsimotining okeanografik asoslari Proxlorokokk ekotiplar ". Ilm-fan. 312 (5775): 918–921. Bibcode:2006 yil ... 312..918B. doi:10.1126 / science.1122692. PMID 16690867. S2CID 20738145.
- ^ a b Brum JR, Schenck RO, Sallivan MB (sentyabr 2013). "Dengiz viruslarining global morfologik tahlili quyruqsiz viruslarning minimal mintaqaviy o'zgarishini va ustunligini ko'rsatadi". ISME jurnali. 7 (9): 1738–51. doi:10.1038 / ismej.2013.67. PMC 3749506. PMID 23635867.
- ^ Krupovich M, Bamford DH (2007). "Litsik dumsiz dsDNA phage PM2 bilan bog'liq taxminiy profillar suv bakteriyalari genomlarida keng tarqalgan". BMC Genomics. 8: 236. doi:10.1186/1471-2164-8-236. PMC 1950889. PMID 17634101.
- ^ Xue H, Xu Y, Boucher Y, Polz MF (2012). "Vibrio atrof-muhit vabo populyatsiyasi ichida roman filamentli fazaning yuqori chastotasi, VCYϕ". Amaliy va atrof-muhit mikrobiologiyasi. 78 (1): 28–33. doi:10.1128 / AEM.06297-11. PMC 3255608. PMID 22020507.
- ^ Roux S, Krupovich M, Poulet A, Debroas D, Enault F (2012). "Virom o'qishlaridan yig'ilgan 81 ta to'liq genomlar to'plami orqali Microviridae virusli oilasining rivojlanishi va xilma-xilligi". PLOS ONE. 7 (7): e40418. Bibcode:2012PLoSO ... 740418R. doi:10.1371 / journal.pone.0040418. PMC 3394797. PMID 22808158.
- ^ Kauffman, Ketrin M.; Husayn, Fotima A.; Yang, quvonch; Arevalo, Filipp; Braun, Julia M.; Chang, Uilyam K.; Vaninsberghe, Devid; Elsherbini, Jozef; Sharma, Radhey S.; Katler, Maykl B.; Kelli, Libusha; Polz, Martin F. (2018). "Dengiz bakteriyalarini tanib bo'lmaydigan qotillar sifatida dsDNA viruslarining asosiy nasl-nasabi". Tabiat. 554 (7690): 118–122. Bibcode:2018Natur.554..118K. doi:10.1038 / tabiat25474. PMID 29364876. S2CID 4462007.
- ^ Olimlar Jahon Okeanlarida yangi turdagi viruslarni topmoqdalar: Avtolykiviridae, kuni: ilmiy yangiliklar, 25 yanvar 2018 yil
- ^ Okeanda g'alati DNKga ega bo'lgan hech qachon ko'rilmagan viruslar topildi, bo'yicha: fanogohlantirish, 2018 yil 25-yanvar
- ^ NCBI: Avtolykiviridae (oila) - tasniflanmagan dsDNA viruslari
- ^ Lawrence CM, Menon S, Eilers BJ, Bothner B, Khayat R, Duglas T, Young MJ (2009). "Archaeal viruslarning strukturaviy va funktsional tadqiqotlari". Biologik kimyo jurnali. 284 (19): 12599–603. doi:10.1074 / jbc.R800078200. PMC 2675988. PMID 19158076.
- ^ Prangishvili D, Forterre P, Garret RA (2006). "Arxeya viruslari: birlashtiruvchi ko'rinish". Tabiat sharhlari Mikrobiologiya. 4 (11): 837–48. doi:10.1038 / nrmicro1527. PMID 17041631. S2CID 9915859.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Prangishvili D, Garret RA (2004). "Krenarxeal gipertermofil viruslarning juda xilma-xil morfotiplari va genomlari". Biokimyoviy jamiyat bilan operatsiyalar. 32 (Pt 2): 204-8. doi:10.1042 / BST0320204. PMID 15046572.
- ^ Mojica FJ, Díez-Villaseñor C, García-Martiníz J, Soria E (2005 yil fevral). "Muntazam ravishda joylashtirilgan prokaryotik takrorlanishlarning ketma-ketligi xorijiy genetik elementlardan kelib chiqadi". Molekulyar evolyutsiya jurnali. 60 (2): 174–82. Bibcode:2005JMolE..60..174M. doi:10.1007 / s00239-004-0046-3. PMID 15791728. S2CID 27481111.
- ^ Makarova KS, Grishin NV, Shabalina SA, Wolf YI, Koonin EV (mart 2006). "Prokaryotlarda RNK-aralashuvga asoslangan taxminiy immunitet tizimi: prognoz qilingan fermentativ apparatni hisoblash tahlili, eukaryotik RNAi bilan funktsional o'xshashlik va gipotetik ta'sir mexanizmlari". Biologiya to'g'ridan-to'g'ri. 1: 7. doi:10.1186/1745-6150-1-7. PMC 1462988. PMID 16545108.
- ^ van der Oost J, Westra ER, Jekson RN, Wiedenheft B (iyul 2014). "CRISPR-Cas tizimlarining strukturaviy va mexanik asoslarini ochish". Tabiat sharhlari Mikrobiologiya. 12 (7): 479–92. doi:10.1038 / nrmicro3279. PMC 4225775. PMID 24909109.
- ^ Dimmock NJ, Easton AJ, Leppard K (2007). Zamonaviy virusologiyaga kirish (Oltinchi nashr). Blackwell Publishing. p.70. ISBN 978-1-4051-3645-7.
- ^ Tomaru Y, Kimura K va Nagasaki K (2015) "Dengiz protisti viruslari". In: Ohtsuka S, Suzaki T, Horiguchi T, Suzuki N, F emas (eds) Dengiz protistlari sahifalar 501-517. Springer, Tokio. doi:10.1007/978-4-431-55130-0_20. ISBN 978-4-431-55130-0.
- ^ Hyman, Pol; Abedon, Stiven T. (2012). "Kichik burgalar: mikroorganizmlarning viruslari". Scientifica. 2012: 1–23. doi:10.6064/2012/734023. PMC 3820453. PMID 24278736.. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 3.0 xalqaro litsenziyasi.
- ^ Qisqasi, S.M. (2012). "Eukaryotik suv o'tlarini yuqtiradigan viruslar ekologiyasi". Atrof-muhit mikrobiologiyasi. 14 (9): 2253–2271. doi:10.1111 / j.1462-2920.2012.02706.x. PMID 22360532.
- ^ a b "Virusli hudud". ExPASy. Olingan 15 iyun 2015.
- ^ a b ICTV. "Viruslar taksonomiyasi: 2014 yil chiqarilishi". Olingan 15 iyun 2015.
- ^ Yolken, RH; va boshq. (2014). "Xlorovirus ATCV-1 inson orofaringeal virusomining bir qismidir va odam va sichqonlarda kognitiv funktsiyalarning o'zgarishi bilan bog'liq". Proc Natl Acad Sci U S A. 111 (45): 16106–16111. Bibcode:2014 yil PNAS..11116106Y. doi:10.1073 / pnas.1418895111. PMC 4234575. PMID 25349393.
- ^ Tomaru, Yuji; Shirai, Yoko; Nagasaki, Keyto (2008-08-01). "Fikodnavirusning zararli gul hosil qiluvchi raphidofitini yuqtiradigan ekologiyasi, fiziologiyasi va genetikasi. Heterosigma akashiwo". Baliqchilik fani. 74 (4): 701–711. doi:10.1111 / j.1444-2906.2008.01580.x. S2CID 23152411.
- ^ Nagasaki, Keyzo; Tarutani, Kenji; Yamaguchi, Mineo (1999-03-01). "Heterosigma akashiwo virusining o'sish xususiyatlari va uni qizil oqimni boshqarish uchun mikrobiologik vosita sifatida foydalanish". Amaliy va atrof-muhit mikrobiologiyasi. 65 (3): 898–902. doi:10.1128 / AEM.65.3.898-902.1999. PMC 91120. PMID 10049839.
- ^ Eng katta ma'lum bo'lgan virusli genomlar Giantviruses.org. Kirish: 11 iyun 2020.
- ^ Sigee, Devid (2005-09-27). Chuchuk suv mikrobiologiyasi: biologik xilma-xillik va suv muhitida mikroorganizmlarning dinamik o'zaro ta'siri. John Wiley & Sons. ISBN 9780470026472.
- ^ a b v Wigington CH, Sonderegger DL, Brussaard CP, Buchan A, Finke JF, Fuhrman J, Lennon JT, Middelboe M, Suttle CA, Stock C, Wilson WH (2015-08-26). "Jahon okeanida virus va mikrob hujayralari ko'payishi o'rtasidagi munosabatni qayta ko'rib chiqish". bioRxiv: 025544. bioRxiv 10.1101/025544. doi:10.1101/025544.
- ^ Parikka KJ, Le Romancer M, Wauters N, Jacquet S (2017 yil may). "Virus-prokaryot nisbati (VPR) ni aniqlash: turli xil ekotizimlarda virus xosti munosabatlari to'g'risida tushunchalar". Kembrij falsafiy jamiyati biologik sharhlari. 92 (2): 1081–1100. doi:10.1111 / brv.12271. PMID 27113012. S2CID 3463306.
- ^ Midbelbo, M.; Brussaard, C. (2017). "Dengiz viruslari: dengiz ekotizimining asosiy ishtirokchilari". Viruslar. 9 (10): 302. doi:10.3390 / v9100302. PMC 5691653. PMID 29057790.
- ^ T.Jonson, P.T. (1984). "Dengiz umurtqasiz hayvonlarining virusli kasalliklari". Helgoländer Meeresuntersuchungen. 37 (1–4): 65–98. Bibcode:1984HM ..... 37 ... 65J. doi:10.1007 / BF01989296. S2CID 30161955.
- ^ Renault T (2011) "Dengiz mollyuskalarini yuqtiradigan viruslar" In: Hurst CJ (Ed) Virusli ekologiyani o'rganish, 2-jild: Hayvonlar xost tizimlari, John Wiley & Sons. ISBN 9781118024584.
- ^ Arzul, I .; Korbeil, S .; Morga, B .; Renault, T. (2017). "Dengiz mollyuskalarini yuqtiradigan viruslar" (PDF). Umurtqasizlar patologiyasi jurnali. 147: 118–135. doi:10.1016 / j.jip.2017.01.009. PMID 28189502.
- ^ Dawsoni, Solaster. "Isrof qilish sindromi ta'sir qilgan dengiz yulduzlari turlari". Pacificrockyintertidal.org Seastarwasting.org (nd): n. sahifa. Ekologiya va evolyutsion biologiya. Internet.
- ^ "Dengiz yulduzlarini isrof qilish sindromi | MARINe". eeb.ucsc.edu. Olingan 2018-06-03.
- ^ a b v Merfi, FA; Gibbs, EPJ; Horzinek, MC; Studdart MJ (1999). Veterinariya virusologiyasi. Boston: Academic Press. ISBN 978-0-12-511340-3.
- ^ Chili kasalliklarini engishda yordam berish uchun Nyu-Brunsvik Baliq haqida ma'lumot va xizmatlar
- ^ Ma'lumotlar varag'i - Atlantika losos baliqlari tadqiqotlari Arxivlandi 2010 yil 29 dekabr, soat Orqaga qaytish mashinasi Kanadadagi baliqchilik va okeanlar. Qabul qilingan 12 may 2009 yil.
- ^ Hall AJ, Jepson PD, Goodman SJ, Harkonen T (2006). "Shimoliy va Evropa dengizlarida Phocine distemper virusi - ma'lumotlar va modellar, tabiat va oziqlanish". Biologik konservatsiya. 131 (2): 221–29. doi:10.1016 / j.biocon.2006.04.008.
- ^ a b v Suttle CA (oktyabr 2007). "Dengiz viruslari - global ekotizimning asosiy ishtirokchilari". Tabiat sharhlari Mikrobiologiya. 5 (10): 801–12. doi:10.1038 / nrmicro1750. PMID 17853907. S2CID 4658457.
- ^ Abrahão, Jnatas; Silva, Lorena; Silva, Lyudmila Santos; Xalil, Jak Yaakub Bou; Rodriges, Rodrigo; Arantes, Talita; Assis, Felipe; Boratto, Paulo; Andrade, Migel; Kron, Erna Gessiyen; Ribeyro, Bergmann; Bergier, Ivan; Seligmann, Herve; Gigo, Erik; Kolson, Filipp; Levasyor, Entoni; Kroemer, Gvido; Raul, Dide; Scola, Bernard La (2018 yil 27-fevral). "Kuyruklu ulkan Tupanvirus ma'lum bo'lgan virus atmosferasining eng to'liq tarjima apparatiga ega". Tabiat aloqalari. 9 (1): 749. Bibcode:2018NatCo ... 9..749A. doi:10.1038 / s41467-018-03168-1. PMC 5829246. PMID 29487281.
- ^ Xiao, C .; Fischer, M.G .; Bolotaulo, D.M .; Ulloa-Rondeu, N .; Avila, G.A .; Suttle, Kaliforniya (2017). "Cafeteria roenbergensis virusi kapsidini Cryo-EM rekonstruktsiyasi ulkan viruslar uchun yangi yig'ilish yo'lini taklif qiladi". Ilmiy ma'ruzalar. 7 (1): 5484. Bibcode:2017 yil NatSR ... 7.5484X. doi:10.1038 / s41598-017-05824-w. PMC 5511168. PMID 28710447.
- ^ Bichell RE. "Gigant virus genlarida ularning sirli kelib chiqishi to'g'risida maslahatlar". Hamma narsa ko'rib chiqildi.
- ^ Van Etten JL (2011 yil iyul-avgust). "Gigant viruslar". Amerikalik olim. 99 (4): 304–311. doi:10.1511/2011.91.304.
- ^ Legendre M, Arslan D, Abergel C, Claverie JM (yanvar 2012). "Megavirus genomikasi va hayotning tushunarsiz to'rtinchi domeni". Kommunikativ va integral biologiya. 5 (1): 102–6. doi:10.4161 / cib.18624. PMC 3291303. PMID 22482024.
- ^ Schulz F, Yutin N, Ivanova NN, Ortega DR, Lee TK, Vierheilig J, Daims H, Horn M, Wagner M, Jensen GJ, Kyrpides NC, Koonin EV, Woyke T (aprel 2017). "Tarjima tizimi tarkibiy qismlarining kengaytirilgan komplektli ulkan viruslari" (PDF). Ilm-fan. 356 (6333): 82–85. Bibcode:2017Sci ... 356 ... 82S. doi:10.1126 / science.aal4657. PMID 28386012. S2CID 206655792.
- ^ Bckström D, Yutin N, Yorgensen SL, Dharamshi J, Xoma F, Zaremba-Niedviedzka K, Spang A, Wolf YI, Koonin EV, Ettema TJ (mart 2019). "Chuqur dengiz cho'kindilaridan virus genomlari okean megaviromini kengaytiradi va virusli gigantizmning mustaqil kelib chiqishini qo'llab-quvvatlaydi". mBio. 10 (2): e02497-02418. doi:10.1128 / mBio.02497-18. PMC 6401483. PMID 30837339.
- ^ Duponchel, S; Fischer, MG (mart, 2019). "Viva lavidaviruslari! Gigant DNK viruslarini parazitlashtiradigan virofaglarning beshta xususiyati". PLOS patogenlari. 15 (3): e1007592. doi:10.1371 / journal.ppat.1007592. PMC 6428243. PMID 30897185.
- ^ a b Fischer MG, Suttle CA (aprel 2011). "Katta DNK transpozonlari kelib chiqishidagi virusofag". Ilm-fan. 332 (6026): 231–4. Bibcode:2011 yil ... 332..231F. doi:10.1126 / science.1199412. PMID 21385722. S2CID 206530677.
- ^ a b Fischer MG, Hackl (2016 yil dekabr). "Xost genomining integratsiyasi va virusofag mavirusining ulkan virus ta'sirida reaktivatsiyasi". Tabiat. 540 (7632): 288–91. Bibcode:2016 yil natur.540..288F. doi:10.1038 / tabiat20593. PMID 27929021. S2CID 4458402.
- ^ Katsurakis, Aris; Asvad, Amr (2014). "Gigant viruslar, virusofaglar va ularning qarindoshlari xost genomlarida kelib chiqishi". BMC biologiyasi. 12: 2–3. doi:10.1186 / s12915-014-0051-y. PMC 4096385. PMID 25184667.
- ^ Krupovich, Mart; Kun, Jens; Fischer, Metthias (2015 yil kuzi). "Virofaglar va sun'iy yo'ldosh viruslari uchun tasniflash tizimi" (PDF). Virusologiya arxivi. 161 (1): 233–247. doi:10.1007 / s00705-015-2622-9. PMID 26446887. S2CID 14196910 - Springer orqali.
- ^ Rou, Simon; Chan, Leong-Kit; Egan, Rob; Malmstrom, Reks R.; MakMaxon, Ketrin D. Sallivan, Metyu B. (2017). "Vaqt ketma-ketligi metagenomikasi orqali baholangan virofaglar ekogenomikasi va ularning ulkan viruslari xostlari". Tabiat aloqalari. 8 (1): 858. Bibcode:2017NatCo ... 8..858R. doi:10.1038 / s41467-017-01086-2. ISSN 2041-1723. PMC 5636890. PMID 29021524.
- ^ Paez-Espino, Devid; Chjou, Jingli; Rou, Simon; Nayfax, Stiven; Pavlopulos, Georgios A.; Shuls, Frederik; MakMaxon, Ketrin D. Uolsh, Devid; Voyk, Tanja; Ivanova, Natalya N.; Eloe-Fadrosh, Emili A.; Tring, Susanna G.; Kirpides, Nikos C. (2019-12-10). "Global metagenomika orqali topilgan virusofaglarning xilma-xilligi, evolyutsiyasi va tasnifi". Mikrobiom. 7 (1): 157. doi:10.1186 / s40168-019-0768-5. PMC 6905037. PMID 31823797.
- ^ Duponchel, S; Fischer, MG (2019). "Viva lavidaviruslari! Gigant DNK viruslarini parazitlashtiradigan virofaglarning beshta xususiyati". PLOS Pathog. 15 (3): e1007592. doi:10.1371 / journal.ppat.1007592. PMC 6428243. PMID 30897185.. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Fischer, M. G.; Allen, M. J .; Uilson, V. X.; Suttle, C. A. (2010). "Gendagi ajoyib komplektli ulkan virus dengiz zooplanktonini yuqtiradi" (PDF). Milliy fanlar akademiyasi materiallari. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073/pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Matthias G. Fischer; Michael J. Allen; William H. Wilson; Curtis A. Suttle (2010). "Giant virus with a remarkable complement of genes infects marine zooplankton" (PDF). Milliy fanlar akademiyasi materiallari. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073/pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Massana, Ramon; Javier Del Campo; Christian Dinter; Ruben Sommaruga (2007). "Crash of a population of the marine heterotrophic flagellate Roenbergensis kafeteryasi by viral infection". Atrof-muhit mikrobiologiyasi. 9 (11): 2660–2669. doi:10.1111/j.1462-2920.2007.01378.x. PMID 17922751. S2CID 30191542.
- ^ Waldor MK, Friedman DI, Adhya SL, eds. (2005). Phages: their role in bacterial pathogenesis and biotechnology. Washington DC: ASM Press. pp.450. ISBN 978-1-55581-307-9.
- ^ Käse, Laura; Geuer, Jana K. (2018). "Phytoplankton Responses to Marine Climate Change – an Introduction". YOUMARES 8 - Chegaralar bo'ylab okeanlar: bir-biridan o'rganish. 55-71 betlar. doi:10.1007/978-3-319-93284-2_5. ISBN 978-3-319-93283-5.
- ^ Weitz JS, Wilhelm SW (2013). "An ocean of viruses". Olim. 27 (7): 35–39.
- ^ Suttle CA (September 2005). "Dengizdagi viruslar". Tabiat. 437 (7057): 356–61. Bibcode:2005 yil Natura. 437..356S. doi:10.1038 / nature04160. PMID 16163346. S2CID 4370363.
- ^ Wilhelm SW, Suttle CA (1999). "Viruses and nutrient cycles in the sea: viruses play critical roles in the structure and function of aquatic food webs". BioScience. 49 (10): 781–88. doi:10.2307/1313569. JSTOR 1313569.
- ^ Suttle CA (October 2007). "Marine viruses—major players in the global ecosystem". Tabiat sharhlari Mikrobiologiya. 5 (10): 801–12. doi:10.1038 / nrmicro1750. PMID 17853907. S2CID 4658457.
- ^ Suttle CA (September 2005). "Dengizdagi viruslar". Tabiat. 437 (7057): 356–61. Bibcode:2005 yil Natura. 437..356S. doi:10.1038 / nature04160. PMID 16163346. S2CID 4370363.
- ^ Shelford EJ, Suttle CA (2018). "Virus-mediated transfer of nitrogen from heterotrophic bacteria to phytoplankton". Biogeoscience. 15 (3): 809–15. Bibcode:2018BGeo...15..809S. doi:10.5194/bg-15-809-2018.
- ^ Vaynbauer, Markus G.; va boshq. (2007). "Virusli lizis va boqishdagi protistaning bakterial biomassaga, hosil bo'lishiga va xilma-xilligiga sinergik va antagonistik ta'siri". Atrof-muhit mikrobiologiyasi. 9 (3): 777–788. doi:10.1111 / j.1462-2920.2006.01200.x. PMID 17298376.
- ^ Robinzon, Kerol va Nagappa Ramayah. "Mikrobial heterotrofik metabolizm darajasi mikrobial uglerod nasosini cheklaydi." Amerika ilm-fanni rivojlantirish assotsiatsiyasi, 2011 y.
- ^ Heinrichs, M.E., Mori, C. and Dlugosch, L. (2020) "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". In: YOUMARES 9-The Oceans: Our Research, Our Future , pages 279–297. Springer. doi:10.1007/978-3-030-20389-4_15.
- ^ Dávila-Ramos S, Castelán-Sánchez HG, Martínez-Ávila L, Sánchez-Carbente MD, Peralta R, Hernández-Mendoza A, et al. (2019). "A Review on Viral Metagenomics in Extreme Environments". Mikrobiologiyadagi chegara. 10: 2403. doi:10.3389/fmicb.2019.02403. PMC 6842933. PMID 31749771.
- ^ Fuhrman, Jed A. (1999). "Marine viruses and their biogeochemical and ecological effects". Tabiat. 399 (6736): 541–548. Bibcode:1999Natur.399..541F. doi:10.1038/21119. ISSN 0028-0836. PMID 10376593. S2CID 1260399.
- ^ Tsai, An-Yi, Gwo-Ching Gong, and Yu-Wen Huang. "Importance of the Viral Shunt in Nitrogen Cycling in Synechococcus Spp. Growth in Subtropical Western Pacific Coastal Waters." Terrestrial, Atmospheric & Oceanic Sciences25.6 (2014).
- ^ Vilgelm, Stiven V.; Suttle, Kertis A. (1999). "Viruses and nutrient cycles in the sea: viruses play critical roles in the structure and function of aquatic food webs". BioScience. 49 (10): 781–788. doi:10.2307/1313569. JSTOR 1313569.
- ^ Bar-On, YM; Fillips, R; Milo, R (2018). "Yerdagi biomassaning tarqalishi". PNAS. 115 (25): 6506–6511. doi:10.1073 / pnas.1711842115. PMC 6016768. PMID 29784790.
- ^ a b Suttle, Curtis A. (2005). "Dengizdagi viruslar". Tabiat. 437 (7057): 356–361. Bibcode:2005 yil Natura. 437..356S. doi:10.1038 / nature04160. ISSN 0028-0836. PMID 16163346. S2CID 4370363.
- ^ www.cdc.gov. Zararli algal gullari: Qizil oqim: Uy [Olingan 2014-12-19].
- ^ a b v Suttle, Kurtis A. (2007 yil oktyabr). "Dengiz viruslari - global ekotizimning asosiy ishtirokchilari". Tabiat sharhlari Mikrobiologiya. 5 (10): 801–812. doi:10.1038 / nrmicro1750. ISSN 1740-1526. PMID 17853907. S2CID 4658457.
- ^ Breitbart M, Thompson L, Suttle C, Sullivan M (2007-06-01). "Exploring the Vast Diversity of Marine Viruses" (PDF). Okeanografiya. 20 (2): 135–139. doi:10.5670/oceanog.2007.58.
- ^ Hurwitz BL, U'Ren JM (June 2016). "Viral metabolic reprogramming in marine ecosystems". Mikrobiologiyaning hozirgi fikri. 31: 161–168. doi:10.1016/j.mib.2016.04.002. PMID 27088500.
- ^ Hurwitz BL, Hallam SJ, Sullivan MB (November 2013). "Metabolic reprogramming by viruses in the sunlit and dark ocean". Genom biologiyasi. 14 (11): R123. doi:10.1186/gb-2013-14-11-r123. PMC 4053976. PMID 24200126.
- ^ Anantharaman K, Duhaime MB, Breier JA, Wendt KA, Toner BM, Dick GJ (May 2014). "Sulfur oxidation genes in diverse deep-sea viruses". Ilm-fan. 344 (6185): 757–60. Bibcode:2014Sci...344..757A. doi:10.1126/science.1252229. hdl:1912/6700. PMID 24789974. S2CID 692770.
- ^ Roux S, Hawley AK, Torres Beltran M, Scofield M, Schwientek P, Stepanauskas R, et al. (Avgust 2014). "Ecology and evolution of viruses infecting uncultivated SUP05 bacteria as revealed by single-cell- and meta-genomics". eLife. 3: e03125. doi:10.7554/elife.03125. PMC 4164917. PMID 25171894.
- ^ Forterre P, Philippe H (1999). "The last universal common ancestor (LUCA), simple or complex?". Biologik byulleten. 196 (3): 373–5, discussion 375–7. doi:10.2307/1542973. JSTOR 1542973. PMID 11536914.
- ^ Culley, A.I.; Lang, A.S.; Suttle, Kaliforniya (2006). "Metagenomic analysis of coastal RNA virus communities". Ilm-fan. 312 (5781): 1795–1798. Bibcode:2006Sci...312.1795C. doi:10.1126/science.1127404. PMID 16794078. S2CID 20194876.
- ^ a b Rahlff, Janina (2019). "The Virioneuston: A Review on Viral–Bacterial Associations at Air–Water Interfaces". Viruslar. 11 (2): 191. doi:10.3390/v11020191. PMC 6410083. PMID 30813345.. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ a b Hutchinson GE (1961). "The Paradox of the Plankton". Amerikalik tabiatshunos. 95 (882): 137–145. doi:10.1086/282171. JSTOR 2458386.
- ^ Danovaro R, Dell'Anno A, Corinaldesi C, Magagnini M, Noble R, Tamburini C, Weinbauer M (August 2008). "Major viral impact on the functioning of benthic deep-sea ecosystems". Tabiat. 454 (7208): 1084–7. Bibcode:2008Natur.454.1084D. doi:10.1038/nature07268. PMID 18756250. S2CID 4331430.
- ^ Anderson, Rika E.; Brazelton, Uilyam J.; Baross, Jon A. (2011). "Chuqur er osti biosferasining genetik landshaftiga viruslar ta'sir qiladimi?". Mikrobiologiyadagi chegara. 2: 219. doi:10.3389/fmicb.2011.00219. ISSN 1664-302X. PMC 3211056. PMID 22084639.
- ^ Anderson, Rika E.; Sogin, Mitchell L.; Baross, John A. (2014-10-03). "Metagenomika orqali ochilgan gidrotermal ventilyatsiya ekotizimidagi viruslar, bakteriyalar va arxeylarning evolyutsion strategiyalari". PLOS ONE. 9 (10): e109696. Bibcode:2014PLoSO ... 9j9696A. doi:10.1371 / journal.pone.0109696. ISSN 1932-6203. PMC 4184897. PMID 25279954.
- ^ Ortmann, Elis S.; Suttle, Kurtis A. (2005 yil avgust). "Dengizdagi gidrotermal shamollatish tizimida viruslarning ko'pligi viruslar vositachiligida mikroblarning o'limidan dalolat beradi". Chuqur dengiz tadqiqotlari I qism: Okeanografik tadqiqotlar. 52 (8): 1515–1527. Bibcode:2005 yil DSRI ... 52.1515O. doi:10.1016 / j.dsr.2005.04.002. ISSN 0967-0637.
- ^ Breitbart, Mya (2012-01-15). "Dengiz viruslari: haqiqat yoki jur'at". Dengizchilik fanining yillik sharhi. 4 (1): 425–448. Bibcode:2012ARMS .... 4..425B. doi:10.1146 / annurev-marine-120709-142805. ISSN 1941-1405. PMID 22457982.
- ^ Goldenfeld, Nayjel; Vuz, Karl (2007 yil yanvar). "Biologiyaning navbatdagi inqilobi". Tabiat. 445 (7126): 369. arXiv:q-bio / 0702015. Bibcode:2007 yil natur.445..369G. doi:10.1038 / 445369a. ISSN 0028-0836. PMID 17251963. S2CID 10737747.
- ^ Leggett HC, Buckling A, Long GH, Boots M (October 2013). "Generalism and the evolution of parasite virulence". Ekologiya va evolyutsiya tendentsiyalari. 28 (10): 592–6. doi:10.1016/j.tree.2013.07.002. PMID 23968968.
- ^ Flores CO, Valverde S, Weitz JS (March 2013). "Multi-scale structure and geographic drivers of cross-infection within marine bacteria and phages". ISME jurnali. 7 (3): 520–32. doi:10.1038/ismej.2012.135. PMC 3578562. PMID 23178671.
- ^ Krupovic M, Bamford DH (July 2007). "Litsik dumsiz dsDNA phage PM2 bilan bog'liq taxminiy profillar suv bakteriyalari genomlarida keng tarqalgan". BMC Genomics. 8: 236. doi:10.1186/1471-2164-8-236. PMC 1950889. PMID 17634101.
- ^ Xue H, Xu Y, Boucher Y, Polz MF (January 2012). "Vibrio vabo atrof-muhit populyatsiyasi ichida yangi filamentli fagning yuqori chastotasi, VCY φ". Amaliy va atrof-muhit mikrobiologiyasi. 78 (1): 28–33. doi:10.1128 / AEM.06297-11. PMC 3255608. PMID 22020507.
- ^ a b Roux S, Krupovich M, Poulet A, Debroas D, Enault F (2012). "Virom o'qishlaridan yig'ilgan 81 ta to'liq genomlar to'plami orqali Microviridae virusli oilasining rivojlanishi va xilma-xilligi". PLOS ONE. 7 (7): e40418. Bibcode:2012PLoSO ... 740418R. doi:10.1371 / journal.pone.0040418. PMC 3394797. PMID 22808158.
- ^ Hurwitz BL, Sullivan MB (2013). "The Pacific Ocean virome (POV): a marine viral metagenomic dataset and associated protein clusters for quantitative viral ecology". PLOS ONE. 8 (2): e57355. Bibcode:2013PLoSO...857355H. doi:10.1371/journal.pone.0057355. PMC 3585363. PMID 23468974.
- ^ Roux S, Enault F, Hurwitz BL, Sullivan MB (2015-05-28). "VirSorter: mining viral signal from microbial genomic data". PeerJ. 3: e985. doi:10.7717/peerj.985. PMC 4451026. PMID 26038737.
- ^ Ren J, Ahlgren NA, Lu YY, Fuhrman JA, Sun F (July 2017). "VirFinder: a novel k-mer based tool for identifying viral sequences from assembled metagenomic data". Mikrobiom. 5 (1): 69. doi:10.1186/s40168-017-0283-5. PMC 5501583. PMID 28683828.
- ^ Paez-Espino D, Eloe-Fadrosh EA, Pavlopoulos GA, Thomas AD, Huntemann M, Mikhailova N, et al. (Avgust 2016). "Uncovering Earth's virome". Tabiat. 536 (7617): 425–30. Bibcode:2016Natur.536..425P. doi:10.1038/nature19094. PMID 27533034. S2CID 4466854.
- ^ Coutinho FH, Silveira CB, Gregoracci GB, Thompson CC, Edwards RA, Brussaard CP, et al. (2017 yil iyul). "Marine viruses discovered via metagenomics shed light on viral strategies throughout the oceans". Tabiat aloqalari. 8: 15955. Bibcode:2017NatCo...815955C. doi:10.1038/ncomms15955. PMC 5504273. PMID 28677677.

| Komponentlar |  | |
|---|---|---|
| Virusli hayot aylanishi | ||
| Genetika | ||
| Uy egasi tomonidan | ||
| Boshqalar | ||
| ||