Dengiz prokaryotlari - Marine prokaryotes
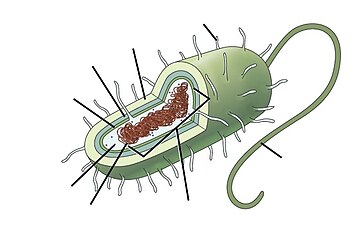
Dengiz prokaryotlari dengizdir bakteriyalar va dengiz arxey. Ular yashash joylari bilan belgilanadi prokaryotlar yashaydiganlar dengiz muhiti, ya'ni sho'r suv dengizlar yoki okeanlar yoki sho'r qirg'oq suvi daryolar. Hammasi uyali hayot shakllari prokaryotlarga va eukaryotlarga bo'linishi mumkin. Eukaryotlar bor organizmlar uning hujayralarida a yadro ichida joylashgan membranalar, prokaryotlar esa membrana ichida o'ralgan yadrosi bo'lmagan organizmlardir.[1][2][3] The uch domenli tizim ning hayotni tasniflash yana bir bo'linishni qo'shadi: prokaryotlar hayotning ikkita sohasiga bo'linadi, mikroskopik bakteriyalar va mikroskopik arxealar, qolgan hamma narsalar, eukaryotlar esa uchinchi domenga aylanadi.[4]
Prokaryotlar muhim rol o'ynaydi ekotizimlar kabi parchalovchilar ozuqa moddalarini qayta ishlash. Ba'zi prokaryotlar patogen, o'simliklar va hayvonlarda kasallik va hatto o'limga olib keladi.[5] Dengiz prokaryotlari muhim darajalar uchun javobgardir fotosintez Okeanda sodir bo'lgan, shuningdek, muhim velosiped uglerod va boshqalar ozuqa moddalari.[6]
Prokaryotlar butun umr davomida yashaydi biosfera. 2018 yilda u jami hisoblangan biomassa sayyoradagi barcha prokaryotlarning 77 milliardga teng edi tonna uglerod (77 Gt C) Bu arxeylar uchun 7 Gt C va bakteriyalar uchun 70 Gt S dan iborat. Ushbu ko'rsatkichlar sayyoramizdagi hayvonlar uchun umumiy biomassaning taxminiy qariyb 2 Gt S ga va odamlarning umumiy biomassasining 0,06 Gt S ga baholanishi bilan taqqoslanishi mumkin.[7] Bu degani, arxa kollektiv ravishda odamlarning kollektiv biomassasidan 100 baravar, bakteriyalar esa 1000 martadan ko'proq.
Uning 600 million yillik hayoti davomida Yer yuzida hayotning aniq dalillari mavjud emas. Hayot bo'lganida, u dengiz prokaryotlari tomonidan 3200 million yil davomida hukmronlik qildi. Shaklida yanada murakkab hayot toj eukaryotlari ga qadar paydo bo'lmadi Kembriya portlashi shunchaki 500 million yil oldin.[8]
Evolyutsiya
The Yer taxminan 4,54 milliard yoshda.[9][10][11] Ning dastlabki tortishuvsiz dalillari Yerdagi hayot kamida 3,5 milliard yil oldin,[12][13] davomida Earxey Geologik davrdan keyingi davr qobiq avvalgi erigan ergashgandan keyin qattiqlasha boshladi Hadean Eon. Mikrobial mat fotoalbomlar 3,48 milliard yoshli odamda topilgan qumtosh yilda G'arbiy Avstraliya.[14][15]


O'tmishdagi turlar ham o'zlarining evolyutsion tarixlarini qayd etishgan. Qoldiqlar, hozirgi organizmlarning qiyosiy anatomiyasi bilan bir qatorda, morfologik yoki anatomik yozuvlarni tashkil etadi.[16] Paleontologlar zamonaviy va yo'q bo'lib ketgan turlarning anatomiyalarini taqqoslash orqali ushbu turlarning nasl-nasabini aniqlashlari mumkin. Ammo bu yondashuv qattiq tana qismlari bo'lgan, masalan, chig'anoqlar, suyaklar yoki tishlar kabi organizmlar uchun eng muvaffaqiyatli hisoblanadi. Bundan tashqari, bakteriyalar va arxeylar kabi prokaryotlar umumiy morfologiyalarning cheklangan to'plamiga ega bo'lganligi sababli, ularning qoldiqlari ajdodlari haqida ma'lumot bermaydilar.
Prokaryotlar Yerda taxminan 3-4 milliard yil yashagan.[17][18] Hech qanday aniq o'zgarishlar yo'q morfologiya yoki keyingi bir necha milliard yil ichida ushbu organizmlarda uyali tashkilot paydo bo'lgan.[19] Eukaryotik hujayralar 1,6-2,7 milliard yil oldin paydo bo'lgan. Hujayra tuzilishidagi navbatdagi katta o'zgarish bakteriyalarni ökaryotik hujayralar yutib yuborganida, deb nomlangan kooperativ birlashmasida sodir bo'ldi endosimbioz.[20][21] Keyin yutib yuborilgan bakteriyalar va mezbon hujayra koevolyutsiyaga uchragan, bakteriyalar esa mitoxondriyaga yoki gidrogenozomalar.[22] Yana bir yutish siyanobakterial o'xshash organizmlar suv o'tlari va o'simliklarda xloroplastlarning paydo bo'lishiga olib keldi.[23]

Hayot tarixi bu edi bir hujayrali eukaryotlar, prokaryotlar va arxeylar taxminan 610 million yil oldin okeanlarda ko'p hujayrali organizmlar paydo bo'lguncha Ediakaran davr.[17][25] The ko'p hujayralilik evolyutsiyasi turli xil mustaqil hodisalarda, xilma-xil organizmlarda sodir bo'lgan gubkalar, jigarrang suv o'tlari, siyanobakteriyalar, shilimshiq qoliplari va miksobakteriyalar.[26] 2016 yilda olimlar, taxminan 800 million yil oldin, GK-PID deb nomlangan bitta molekulada kichik genetik o'zgarish organizmlarning bitta hujayra organizmidan ko'plab hujayralardan biriga o'tishiga imkon berganligi haqida xabar berishdi.[27]
Ushbu birinchi ko'p hujayrali organizmlar paydo bo'lganidan ko'p o'tmay, taxminan 10 million yil davomida ajoyib biologik xilma-xillik paydo bo'ldi, bu voqea Kembriya portlashi. Bu erda, ko'pchilik turlari fotoalbomlarda zamonaviy hayvonlar va keyinchalik yo'q bo'lib ketgan noyob nasl-nasablar paydo bo'ldi.[28] Kembriya portlashi uchun turli xil ogohlantiruvchilar, shu jumladan to'planish taklif qilingan kislorod ichida atmosfera fotosintezdan.[29]
Fon

Prokaryot va eukaryot so'zlari yunoncha qaerdan kelgan pro "oldin" degan ma'noni anglatadi, EI "yaxshi" yoki "haqiqiy", va degan ma'noni anglatadi karyon "yong'oq", "yadro" yoki "yadro" degan ma'noni anglatadi.[30][31][32] Demak, etimologik nuqtai nazardan prokaryot "yadrodan oldin" va ökaryot "haqiqiy yadro" degan ma'noni anglatadi.
Prokaryotlar va eukaryotlar o'rtasida hayot shakllarining bo'linishi mikrobiologlar tomonidan qat'iy o'rnatildi Rojer Stanier va C. B. van Niel ularning 1962 yilgi maqolalarida, Bakteriya haqida tushuncha.[33] Ushbu tasnifning bir sababi, keyinchalik tez-tez chaqirilgan narsa edi ko'k-yashil suv o'tlari (endi chaqirildi) siyanobakteriyalar ) o'simliklar qatoriga kirishni to'xtatadi, ammo bakteriyalar bilan guruhlanadi.
1990 yilda Karl Vuz va boshq. tanishtirdi uch domenli tizim.[34][35] Prokaryotlar ikkiga bo'lingan domenlar, arxeyalar va bakteriyalar, eukaryotlar esa o'z-o'zidan domenga aylanadi. Oldingi tasniflardan asosiy farqi - arxeylarning bakteriyalardan ajralishi.



Er yuzidagi hayotning dastlabki dalillari kelib chiqadi biogen uglerod imzolari va stromatolit 3,7 milliard yillik toshlardan topilgan toshqotganliklar.[36][37] 2015 yilda mumkin bo'lgan "qoldiqlar biotik hayot "4,1 milliard yillik toshlardan topilgan.[38][39] 2017 yilda Yer yuzidagi hayotning eng qadimgi shakllari haqidagi taxminiy dalillar toshbo'ron qilingan shaklda xabar qilindi mikroorganizmlar yilda kashf etilgan gidrotermal shamollatish 4,28 milliard yil oldin yashagan bo'lishi mumkin bo'lgan yog'ingarchilik hosil bo'lgan okeanlar 4.4 milliard yil oldin va undan ko'p o'tmay Yerning shakllanishi 4,54 milliard yil oldin.[40][41]
Birgalikda yashashning mikrobial paspaslari bakteriyalar va arxey boshida hayotning ustun shakli bo'lgan Archean Eon va dastlabki evolyutsiyadagi ko'plab muhim qadamlar ushbu muhitda sodir bo'lgan deb o'ylashadi.[42] Evolyutsiyasi fotosintez 3,5 Ga atrofida uning chiqindi mahsuloti ko'payishiga olib keldi kislorod atmosferada, ga olib keladi katta oksijenlanish hodisasi 2.4 Ga atrofida boshlangan.[43]
Eukaryotlarning dastlabki dalillari 1,85 ga teng,[44][45] va ular ilgari bo'lgan bo'lishi mumkin edi, ammo ulardagi kisloroddan foydalanishni boshlaganda ularning xilma-xilligi tezlashdi metabolizm. Keyinchalik, 1,7 ga yaqin, ko'p hujayrali organizmlar bilan paydo bo'la boshladi tabaqalashgan hujayralar ixtisoslashtirilgan funktsiyalarni bajarish.[46]

Prokaryotlarni o'z ichiga olgan havo mikroorganizmlari oqimi sayyoramizni ob-havo tizimlari atrofida, ammo tijorat yo'llari ostida aylantiradi.[50] Ba'zi peripatetik mikroorganizmlar quruqlikdagi chang bo'ronlaridan ko'tarilgan, ammo ko'pchiligi dengiz mikroorganizmlaridan kelib chiqqan dengiz spreyi. 2018 yilda olimlar sayyoramizdagi har kvadrat metrga har kuni yuz millionlab viruslar va o'n millionlab bakteriyalar yotqizilganligini xabar qilishdi.[51][52]
Mikroskopik hayot suv osti dengizining xilma-xilligi va hali ham, masalan, roli uchun juda yaxshi tushunilmagan viruslar dengiz ekotizimlarida.[53] Ko'pincha dengiz viruslari bakteriofaglar, o'simliklar va hayvonlar uchun zararsiz, ammo sho'r va chuchuk suv ekotizimlarini tartibga solish uchun juda muhimdir.[54] Ular suv mikroblari jamoalarida bakteriyalar va arxeylarni yuqtiradi va yo'q qiladi va ularning eng muhim mexanizmi hisoblanadi uglerodni qayta ishlash dengiz muhitida. O'lik bakterial hujayralardan chiqarilgan organik molekulalar yangi bakterial va alg o'sishini rag'batlantiradi.[55] Virusli faollik ham o'z hissasini qo'shishi mumkin biologik nasos, bu jarayon uglerod bu sekvestrlangan chuqur okeanda.[56]
Dengiz bakteriyalari



Bakteriyalar katta qismini tashkil qiladi domen ning prokaryotik mikroorganizmlar. Odatda bir nechta mikrometrlar uzunligi bo'yicha bakteriyalar sharlardan to novda va spiralgacha bo'lgan bir qator shakllarga ega. Bakteriyalar paydo bo'lgan birinchi hayot shakllaridan biri edi Yer va ularning aksariyat qismida mavjud yashash joylari. Bakteriyalar tuproqda, suvda, kislotali issiq buloqlar, radioaktiv chiqindilar,[58] va chuqur qismlari Yer qobig'i. Bakteriyalar ham yashaydi simbiyotik va parazit o'simliklar va hayvonlar bilan munosabatlar.
Bir marta ko'rib chiqildi o'simliklar sinfni tashkil qiladi Shizomitsetalar, bakteriyalar endi quyidagicha tasniflanadi prokaryotlar. Hayvonlarning hujayralaridan va boshqa narsalardan farqli o'laroq eukaryotlar, bakteriyalar hujayralarida a mavjud emas yadro va kamdan-kam port membrana bilan bog'langan organoidlar. Garchi bu atama bakteriyalar an'anaviy ravishda barcha prokaryotlarni o'z ichiga olgan ilmiy tasnif 90-yillarda kashf qilingandan so'ng prokaryotlar organizmlarning ikki xil guruhidan iborat ekanligi o'zgardi rivojlangan qadimiy umumiy ajdodlardan. Bular evolyutsion sohalar deyiladi Bakteriyalar va Arxeya.[59]
Zamonaviy bakteriyalarning ajdodlari bir hujayrali mikroorganizmlar bo'lgan hayotning birinchi shakllari taxminan 4 milliard yil oldin Yerda paydo bo'lishi. Taxminan 3 milliard yil davomida ko'pchilik organizmlar mikroskopik bo'lib, bakteriyalar va arxealar hayotning ustun shakllari bo'lgan.[60][61] Garchi bakterial bo'lsa ham fotoalbomlar kabi mavjuddir stromatolitlar, ularning o'ziga xos etishmasligi morfologiya ularni bakteriyalar evolyutsiyasi tarixini o'rganish yoki ma'lum bir bakteriya turining kelib chiqish vaqtini belgilash uchun foydalanishga to'sqinlik qiladi. Shu bilan birga, bakteriyalarni tiklash uchun genlar ketma-ketligi ishlatilishi mumkin filogeniya va ushbu tadqiqotlar shuni ko'rsatadiki, bakteriyalar birinchi navbatda arxaeal / eukaryotik nasldan ajralib chiqqan.[62] Ikkinchi buyuk evolyutsion divergensiyada, masalan, arxey va eukaryotlarda bakteriyalar ham ishtirok etgan. Bu erda eukaryotlar qadimgi bakteriyalar kirib borishi natijasida paydo bo'lgan endosimbiyotik o'zlari ehtimol bog'liq bo'lgan eukaryotik hujayralar ajdodlari bilan birlashmalar Arxeya.[21][63] Bunga proto-eukaryotik hujayralar yutish bilan bog'liq alfaproteobakterial shakllanadigan simbionlar mitoxondriya yoki gidrogenozomalar, hali ham ma'lum bo'lgan Eukaryoda mavjud. Keyinchalik mitoxondriyani o'z ichiga olgan ba'zi bir ökaryotlar siyanobakteriyalarga o'xshash organizmlarni ham yutib yuborgan. Bu shakllanishiga olib keldi xloroplastlar suv o'tlari va o'simliklarda. Keyinchalik endosimbiotik hodisalardan kelib chiqqan ba'zi suv o'tlari ham mavjud. Bu erda eukariotlar "ikkinchi avlod" plastidiga aylangan eukaryotik suv o'tlarini yutib yubordi.[64][65] Bu sifatida tanilgan ikkilamchi endosimbioz.
Bakteriyalar belgilangan hajmgacha o'sadi va keyin ko'payadi ikkilik bo'linish, shakli jinssiz ko'payish.[66] Optimal sharoitda bakteriyalar juda tez o'sishi va bo'linishi mumkin va bakteriyalar populyatsiyasi har 9,8 daqiqada ikki baravar tez ko'payishi mumkin.[67]
Pelagibakteriyalar va uning qarindoshlari okeandagi eng ko'p tarqalgan mikroorganizmlar bo'lishi mumkin va ular, ehtimol, dunyodagi eng ko'p tarqalgan bakteriyalar deb da'vo qilingan. Ular barcha mikroblarning taxminan 25% ni tashkil qiladi plankton va yozda ular mo''tadil okean yuzasi suvlarida mavjud bo'lgan hujayralarning taxminan yarmini tashkil qilishi mumkin. Umumiy mo'lligi P. ubique qarindoshlari esa taxminan 2 × 10 ga teng28 mikroblar.[68] Biroq, bu haqda xabar berilgan Tabiat 2013 yil fevral oyida bakteriyofag HTVC010P, qaysi hujumlar P. ubique, topilgan va ehtimol sayyoradagi eng keng tarqalgan organizmdir.[69][70]
Rozeobakter shuningdek, okeandagi eng ko'p va ko'p qirrali mikroorganizmlardan biridir. Ular qirg'oqdan ochiq okeangacha va dengiz muzidan dengiz tubigacha bo'lgan turli xil dengiz yashash joylari bo'yicha xilma-xil bo'lib, qirg'oqdagi dengiz bakteriyalarining 25 foizini tashkil qiladi. A'zolari Rozeobakter jinsi muhim rol o'ynaydi dengiz biogeokimyoviy tsikllari va iqlim o'zgarishi, dengizdagi umumiy uglerodning muhim qismini qayta ishlash. Ular aromatik birikmalarni parchalanishiga va iz metallarini olishga imkon beradigan simbiyotik munosabatlarni hosil qiladi. Ular akvakulturada va kvorumlarni aniqlashda keng qo'llaniladi. Yosunlarning gullashi davrida prokaryotik jamoaning 20-30% Rozeobakteriyalardir.[71][72]
Ma'lum bo'lgan eng katta bakteriya - dengiz Thiomargarita namibiensis, yalang'och ko'z bilan ko'rish mumkin va ba'zan 0,75 mm (750 mkm) ga etadi.[73][74]
Siyanobakteriyalar



Siyanobakteriyalar quyosh nurlarini kimyoviy energiyaga aylantirish qobiliyatini rivojlantirgan birinchi organizmlardir. Ular bir hujayralidan tortib to bakteriyalar filomini (bo'linishini) hosil qiladi ipli va o'z ichiga oladi mustamlaka turlari. Ular er yuzining deyarli hamma joylarida uchraydi: nam tuproqda, ham chuchuk suvda, ham dengiz muhitida, hatto Antarktika jinslarida.[76] Xususan, ba'zi turlari okeanda suzib yuruvchi hujayralar singari uchraydi va ular birinchilardan biri bo'lgan fitoplankton.
Fotosintezdan foydalangan dastlabki birlamchi ishlab chiqaruvchilar okeanik edi siyanobakteriyalar taxminan 2,3 milliard yil oldin.[77][78] Molekulyar ajralib chiqishi kislorod tomonidan siyanobakteriyalar fotosintezning yon mahsuloti sifatida Yer atrofidagi global o'zgarishlarni keltirib chiqardi. O'sha paytda kislorod Yerdagi ko'pchilik hayot uchun zaharli bo'lganligi sababli, bu deyarli yo'q bo'lib ketishiga olib keldi kislorodga toqat qilmaydigan organizmlar, a keskin o'zgarish asosiy hayvon va o'simlik turlarining evolyutsiyasini yo'naltirgan.[79]

Kichkina (0.6 µm ) dengiz siyanobakteriyasi Proxlorokokk, 1986 yilda kashf etilgan, bugungi kunda okean tubining muhim qismini tashkil etadi Oziq ovqat zanjiri va ochiq okean fotosintezining katta qismini tashkil etadi[81] va taxminlarga ko'ra Yer atmosferasidagi kislorodning 20%.[82] Ehtimol, bu Yerdagi eng ko'p naslga mansub nasldan nasldan nasldan nasldan nasldan nasldan naslga o'tishi mumkin: bir millilitr dengiz sathidagi suv 100000 hujayradan yoki undan ko'proq hujayradan iborat bo'lishi mumkin.[83]
Dastlab, biologlar tasniflangan siyanobakteriyalar suv o'tlari sifatida va "ko'k-yashil suv o'tlari" deb nomlangan. Yaqinda paydo bo'lgan fikr - siyanobakteriyalar bakteriyalar va shuning uchun ham bir xil emas Qirollik suv o'tlari sifatida. Aksariyat rasmiylar barchani istisno qiladilar prokaryotlar, va shuning uchun suv o'tlari ta'rifidan siyanobakteriyalar.[84][85]
| Tashqi video | |
|---|---|

Siyanobakteriyalar gullaydi o'limga olib kelishi mumkin siyanotoksinlar

Sinekokok, keng tarqalgan dengiz siyanobakteriyasi

Karboksizomalar turlarining ichida ko'p qirrali qorong'u tuzilmalar ko'rinishida Sinekokok



Dengiz arxeyalari

The arxey (Yunoncha qadimiy[87]) tashkil etadi domen va qirollik ning bir hujayrali mikroorganizmlar. Ushbu mikroblar prokaryotlar, demak ular yo'q hujayra yadrosi yoki boshqa membrana bilan bog'langan organoidlar ularning hujayralarida.
Arxeya dastlab quyidagicha tasniflangan bakteriyalar, ammo bu tasnif eskirgan.[88] Arxeoal hujayralar ularni hayotning boshqa ikkita sohasidan ajratib turadigan noyob xususiyatlarga ega, Bakteriyalar va Eukaryota. Arxeya yana ko'p tan olinganlarga bo'linadi fitna. Tasniflash qiyin, chunki ko'pchilik laboratoriyada ajratilmagan va faqat ularni tahlil qilish orqali aniqlangan nuklein kislotalar ularning atrof-muhitidan namunalarda.
Bakteriyalar va arxeylar odatda hajmi va shakli jihatidan bir-biriga o'xshashdir, ammo bir nechta arxa juda g'alati shakllarga ega, masalan, tekis va kvadrat shaklidagi hujayralar Haloquadratum walsbyi.[89] Bakteriyalarga o'xshash morfologik o'xshashligiga qaramay, arxeylar mavjud genlar va bir nechta metabolik yo'llar eukaryotlar bilan chambarchas bog'liq bo'lgan, xususan fermentlar da ishtirok etish transkripsiya va tarjima. Arxeologik biokimyoning boshqa jihatlari o'ziga xosdir, masalan, ularga tayanish efir lipidlari ularning ichida hujayra membranalari, kabi arxeollar. Arxeya eukaryotlarga qaraganda ko'proq energiya manbalaridan foydalanadi: ular organik birikmalar, masalan, shakar, to ammiak, metall ionlari yoki hatto vodorod gazi. Tuzga chidamli arxey ( Haloarxeya ) energiya manbai sifatida quyosh nurlaridan va boshqa arxey turlaridan foydalaning uglerodni tuzatish; ammo, o'simliklardan farqli o'laroq va siyanobakteriyalar, ma'lum arxea turlari ikkalasini ham qilmaydi. Arxeya jinssiz ko'payish tomonidan ikkilik bo'linish, parchalanish, yoki tomurcuklanma; bakteriyalar va eukaryotlardan farqli o'laroq, ma'lum turlar shakllanmaydi sporlar.
Arxeya okeanlarda juda ko'p, arxeylar esa plankton sayyoradagi organizmlarning eng ko'p tarqalgan guruhlaridan biri bo'lishi mumkin. Arxeya Yer hayotining asosiy qismidir va ikkalasida ham rol o'ynashi mumkin uglerod aylanishi va azot aylanishi. Krenarxeota (eotsitlar) - bu dengiz muhitida juda ko'p bo'lgan va uglerodni fiksatsiya qilishda asosiy hissa qo'shganlardan biri bo'lgan arxey filomidir.[90]

Eotsitlar dengiz arxeyasining eng ko'p tarqalgani bo'lishi mumkin

Halobakteriyalar, tuz bilan to'yingan suvdan topilgan, endi arxey deb tan olingan.

Arxeyaning kvadrat shaklidagi tekis hujayralari Haloquadratum walsbyi

Metanosarcina barkeri, ishlab chiqaradigan dengiz arxeyasi metan

Termofillar, kabi Pirolobus fumarii, 100 ° C dan yuqori darajada omon qoling





Nanoarxey ekvivalentlari 2002 yilda a da kashf etilgan dengiz arxeylarining bir turi gidrotermal shamollatish. Bu termofil haroratda taxminan 80 daraja Selsiyda o'sadi. Nanoarxey kabi ko'rinadi majburiy simbiont ustida arxeon Ignikok. O'shandan beri omon qolish uchun u mezbon organizm bilan aloqada bo'lishi kerak Nanoarxey ekvivalentlari lipidlarni sintez qila olmaydi, lekin ularni uy egasidan oladi. Uning hujayralari atigi 400 tani tashkil qiladi nm diametri bo'yicha ma'lum bo'lgan eng kichik uyali organizmlardan biri va ma'lum bo'lgan eng kichik arxeon.[91][92]
Dengiz arxeylari quyidagicha tasniflangan:[93][94][95][96][97]
- Dengiz guruhi I (MG-I yoki MGI): dengiz Thaumarchaeota Idgacha bo'lgan Ia (aka I.a) kichik guruhlari bilan
- Dengiz guruhi II (MG-II): dengiz Euryarchaeota, buyurtma Poseidoniales[98] IId dan IId gacha bo'lgan kichik guruhlar bilan (IIa o'xshash) Poseidoniaceae, IIb o'xshash Talassarxiya )
Parazitlovchi MGII viruslari quyidagicha tasniflanadi magroviruslar - Dengiz guruhi III (MG-III): shuningdek dengiz Euryarchaeota, D dengiz Bentik guruhi D[99]
- Dengiz guruhi IV (MG-IV): shuningdek dengiz Euryarchaeota[100]
Trofik rejim
Prokaryot metabolizmi tasniflanadi ozuqaviy guruhlar uchta asosiy mezon asosida: manbai energiya, elektron donorlar ishlatilgan va manbasi uglerod o'sish uchun ishlatiladi.[101][102]
| Oziqlanish turi | Energiya manbai | Uglerod manbai | Misollar |
|---|---|---|---|
| Fototroflar | Quyosh nuri | Organik birikmalar (fotoheterotroflar) yoki uglerodni biriktirish (fotoavtotroflar) | Siyanobakteriyalar, Yashil oltingugurt bakteriyalari, Xlorofleksi, yoki Binafsha bakteriyalar |
| Litotroflar | Anorganik birikmalar | Organik birikmalar (lithoterotroflar) yoki uglerodni biriktirish (litoautotroflar) | Termodesulfobakteriyalar, Gidrogenfileya, yoki Nitrospira |
| Organotroflar | Organik birikmalar | Organik birikmalar (ximogeterotroflar) yoki uglerodni fiksatsiya qilish (xemoototroflar) | Bacillus, Klostridium yoki Enterobakteriyalar |
Dengiz prokaryotlari uzoq vaqt davomida juda xilma-xildir. Prokaryotlarning metabolizmi eukaryotlarga qaraganda ancha xilma-xildir va bu juda aniq prokaryotik turlarga olib keladi. Masalan, foydalanishdan tashqari fotosintez yoki organik birikmalar eukaryotlar singari energiya uchun dengiz prokaryotlari energiya olishlari mumkin noorganik birikmalar kabi vodorod sulfidi. Bu dengiz prokaryotlarining rivojlanishiga imkon beradi ekstremofillar o'qigan Antarktidaning muz sathidek sovuq muhitda kriyobiologiya, dengiz osti kabi issiq gidrotermal teshiklar yoki (masalan, yuqori sho'rlangan sharoitdahalofillar ).[103] Ba'zi dengiz prokaryotlari yashaydi simbiyotik tarzda boshqa dengiz organizmlarida yoki tanalarida.

- Fototrofiya bakteriyalarni tasniflashda har doim asosiy rol o'ynashi kerak bo'lgan ayniqsa muhim belgidir.[104]
- Aerobik anoksigenik fototrofik bakteriyalar (AAPB) dengizda keng tarqalgan plankton Bu ochiq okean mikroblari jamiyatining 10% dan ortig'ini tashkil qilishi mumkin. Dengiz AAPBlari ikkita dengizda tasniflanadi (Eritrobakter va Rozeobakter ) nasl. Ular, ayniqsa, mo'l-ko'l bo'lishi mumkin oligotrofik ular jamiyatning 24% tashkil etganligi aniqlangan shartlar.[105] Bular geterotrofik energiya ishlab chiqarish uchun nurdan foydalanadigan, ammo karbonat angidridni asosiy uglerod manbai sifatida ishlata olmaydigan organizmlar. Ko'pchilik majburiydir aerob, ya'ni ular o'sishi uchun kislorodni talab qiladi. Hozirgi ma'lumotlar shuni ko'rsatmoqdaki dengiz bakteriyalari bir necha kunlik naslga ega bo'lishiga qaramay, AAPB ning ishlab chiqarish vaqtini ancha qisqartirganligini ko'rsatadigan yangi dalillar mavjud.[106] Sohil va raf suvlarida ko'pincha AAPB miqdori ko'p, ba'zilari esa 13,51% AAPB% ga etadi. Fitoplankton AAPB% ga ham ta'sir qiladi, ammo bu borada ozgina tadqiqotlar olib borilmagan.[107] Ular turli xillarda ham ko'p bo'lishi mumkin oligotrofik sharoitlar, shu jumladan jahon okeanining eng oligotrofik rejimi.[108] Ular global miqyosda taqsimlanadi eyfotik zona va dengiz mikroblari birlashmasining shu paytgacha tan olinmagan tarkibiy qismini ifodalaydi, bu okeanda ham organik, ham noorganik uglerodning aylanishi uchun juda muhimdir.[109]
- Binafsha bakteriyalar:
- Zetaproteobakteriyalar: bor temirni oksidlovchi neytrofil xemolitoautotroflar, dunyo bo'ylab daryolar va dengiz yashash joylarida tarqalgan.
- Vodorod oksidlovchi bakteriyalar aerob va anaeroblarga bo'linadigan fakultativ avtotroflar. Avvalgi foydalanish vodorod sifatida elektron donor va aktseptor sifatida kislorod, ikkinchisi esa sulfat yoki azot dioksididan foydalanadi elektron qabul qiluvchilar.[110]
Harakatlanish
Harakatlanish qobiliyatidir organizm metabolik energiyadan foydalangan holda mustaqil ravishda harakat qilish.
Flagellar harakati
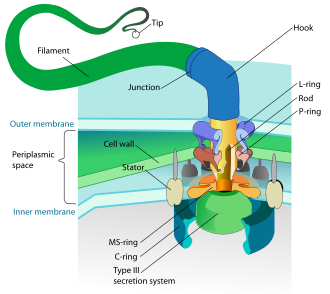

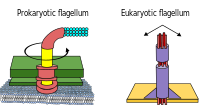
Prokaryotlar, ham bakteriyalar, ham arxeylardan, birinchi navbatda foydalanadilar flagella harakatlanish uchun.
- Bakterial flagella - spiral iplar, ularning har biri a aylanadigan vosita uning tagida soat yo'nalishi bo'yicha yoki teskari yo'nalishda aylanishi mumkin.[112][113][114] Ular bakteriyalar harakatchanligining bir nechta turlaridan ikkitasini ta'minlaydi.[115][116]
- Archaeal flagella deyiladi arxaella va bakterial flagella kabi ishlaydi. Archaellum tizimli ravishda bakterial flagellumga o'xshashdir, ammo u juda ko'p tafsilotlar bilan farq qiladi va noaniq hisoblanadigomologik.[117][111]
Bakteriyalar tomonidan ishlatiladigan aylanma dvigatel modeli an protonlaridan foydalanadi elektrokimyoviy gradient ularning bayroqchalarini harakatga keltirish uchun. Tork bakteriyalar flagellasida flagellum poydevori atrofida proton o'tkazadigan zarralar tomonidan yaratilgan. Flagelning bakteriyalarda aylanish yo'nalishi flagellar motorining perimetri bo'ylab proton kanallarini egallashidan kelib chiqadi.[118]
Ba'zi eukaryotik hujayralar ham flagella-dan foydalanadi - va ular ba'zi protistlar va o'simliklarda va hayvon hujayralarida bo'lishi mumkin. Eukaryotik flagella - bu dumaloq harakat bilan emas, balki oldinga va orqaga kiradigan murakkab uyali proektsiyalar. Prokaryotik flagella rotatorli dvigateldan, eukaryotik flagella esa murakkab sirpanuvchi filament tizimidan foydalanadi. Eukaryotik flagella ATP tomonidan boshqariladigan, prokaryotik flagella esa ATP tomonidan boshqariladigan (arxa) yoki bo'lishi mumkin protonli (bakteriyalar).[119]
Miltillikni tebranish
Miltillikni tebranish sirt ustida harakatlanish uchun ishlatiladigan bakterial harakatchanlikni emaklash shaklidir. Twitching IV tip deb ataladigan sochlarga o'xshash iplarning faolligi bilan bog'liq pili hujayraning tashqi qismidan chiqib, atrofdagi qattiq substratlarga bog'lanib, orqaga tortilib, hujayrani oldinga tortib a tortish uchun kanca.[120][121][122] Ism tebranish harakati mikroskop ostida ko'rib chiqilganda alohida hujayralarning xarakterli siltan va notekis harakatlaridan kelib chiqadi.[123]
Gliding motilligi
Gliding motilligi kabi qo'zg'atuvchi tuzilmalardan mustaqil bo'lgan translokatsiya turidir flagella yoki pili.[124] Gliding mikroorganizmlarning past suvli plyonkalar yuzasi bo'ylab harakatlanishiga imkon beradi. Ushbu harakatlanish mexanizmlari qisman ma'lum. Parvoz qilish tezligi organizmlar orasida turlicha bo'lib, yo'nalishni teskari yo'naltirish qandaydir ichki soat bilan tartibga solinadi.[125] Masalan apikompleksanlar 1-10 mkm / s gacha tez tezlikda harakatlana oladi. Farqli o'laroq Myxococcus xanthus bakteriyalar 5 mm / min tezlikda siljiydi.[126][127]
Ko'chib yuruvchi harakatchanlik
Ko'chib yuruvchi harakatchanlik bakteriyalar populyatsiyasining qattiq yoki yarim qattiq yuzalar bo'ylab tezkor (2-10 mm / s) va muvofiqlashtirilgan translokatsiyasi,[128] va bakterial ko'p hujayralilikning misoli va to'daning harakati. To'qnashuvlar harakati haqida birinchi marta 1972 yilda Xorgen Henrixsen xabar bergan.[129]
Harakatsiz
Harakatsiz turlar o'z kuchlari ostida, atrof-muhit orqali o'zlarini harakatga keltirishga imkon beradigan qobiliyat va tuzilmalarning etishmasligi. Harakatsiz bakteriyalar pichoq naychasida o'stirilganda, ular faqat pichoq chizig'i bo'ylab o'sadi. Agar bakteriyalar harakatchan bo'lsa, chiziq tarqoq bo'lib ko'rinadi va muhitga tarqaladi.[130]
Gaz vakuollari
Ba'zi dengiz prokaryotlari mavjud gaz vakuolalari. Gaz vakuolasi mavjud nanokompyuterlar dengiz bakteriyalari va arxeylarni boshqarishni ta'minlaydigan gazni erkin o'tkazuvchanligi suzish qobiliyati. Ular mil shaklida membrana bilan bog'langan shaklga ega pufakchalar va ba'zi birlarida mavjud plankton prokaryotlar, shu jumladan ba'zilari Siyanobakteriyalar.[131] Hujayralarni suv ustunining yuqori qismida ushlab turish uchun ular ijobiy ishlashga muhtoj bo'lib, ular ishlashni davom ettirishi mumkin fotosintez. Gaz vakuolalari yuqori darajada bo'lgan oqsil qobig'idan iborat hidrofob ichki yuzasi, suv o'tkazmaydigan (va suv bug'ining ichidagi kondensatsiyani to'xtatadigan), lekin ko'pchilik uchun gazlar. Gaz pufakchasi ichi bo'sh tsilindr bo'lgani uchun, atrof qurilganda qulab tushishi mumkin bosim ortadi. Tabiiy seleksiya gaz pufakchasining tuzilishini uning qarshiligini maksimal darajaga ko'tarish uchun juda yaxshi sozlangan buklanish, shu jumladan tashqi mustahkamlovchi oqsil - GvpC, to'qilgan hosepipe ichidagi yashil ip kabi. Gaz pufakchasining diametri va u qulab tushadigan bosim o'rtasida oddiy bog'liqlik mavjud - gaz pufagi qanchalik keng bo'lsa, u zaiflashadi. Shu bilan birga, kengroq gaz pufakchalari samaraliroq bo'lib, tor gaz pufakchalariga qaraganda oqsil birligi uchun ko'proq suzuvchanlikni ta'minlaydi. Turli xil turlar turli xil diametrdagi gaz pufakchalarini ishlab chiqaradi, bu ularga suv ustunining turli chuqurliklarini kolonizatsiya qilishga imkon beradi (tez o'sib boruvchi, yuqori qatlamlarda keng gaz pufakchalari bo'lgan yuqori raqobatbardosh turlar; sekin o'sib boruvchi, qorong'u moslashuvchan, kuchli tor gaz pufakchalari bo'lgan turlar. chuqur qatlamlar).
Hujayra suv ustunidagi balandlikka gaz pufakchalarini sintez qilish orqali erishadi. Hujayra ko'tarilgach, uni ko'paytirishi mumkin uglevod ortgan fotosintez orqali yuk. Juda yuqori va hujayra fotosellash va o'limga olib kelishi mumkin, ammo fotosintez jarayonida hosil bo'lgan uglevod hujayraning zichligini oshiradi va uning cho'kishiga olib keladi. Fotosintez va uglevodlardan hosil bo'lgan uglevodlarning kunlik tsikli katabolizm qorong'i soatlarda hujayraning suv ustunidagi holatini aniq sozlash, uglevod miqdori past bo'lganida va fotosintez zarur bo'lganda uni yuzaga ko'tarish va zararli moddalardan cho'kish uchun etarli bo'ladi. UV nurlanishi hujayraning uglevod miqdori to'ldirilganda. Uglevodning haddan tashqari ko'pligi hujayraning ichki bosimida sezilarli o'zgarishlarni keltirib chiqaradi, bu esa gaz pufakchalari qisilib, qulab tushishiga va hujayraning cho'kib ketishiga olib keladi.
Uchta katta vakuolalar mavjud avlodlar filamentli oltingugurt bakteriyalari, Thioploca, Beggiatoa va Thiomargarita. Sitosol bu nasllarda nihoyatda kamaygan va vakuol hujayraning 40-98 foizini egallashi mumkin.[132] Vakuol tarkibida yuqori konsentratsiyalar mavjud nitrat ionlari va shuning uchun ularni ombor deb o'ylashadi organelle.[133]
Biyolüminesans

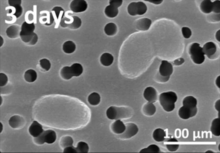
Biolyuminestsent bakteriyalar bor engil ishlab chiqaruvchi bakteriyalar asosan dengiz suvlarida, dengiz cho'kindilarida, chirigan baliqlar yuzasida va dengiz hayvonlari ichaklarida mavjud. Bakterial biolyuminesans keng tarqalmagan bo'lsa-da, quruqlik va chuchuk suv bakteriyalarida ham uchraydi.[120] Ushbu bakteriyalar erkin hayot kechirishi mumkin (masalan Vibrio harveyi ) yoki kabi hayvonlar bilan simbiozda Gavayi Bobtail kalmar (Aliivibrio fischeri ) yoki er usti nematodalar (Photorhabdus luminescens ). Uy egasi organizmlar ushbu bakteriyalarni xavfsiz uy va etarli ovqatlanishni ta'minlaydi. Buning evaziga xostlar bakteriyalar tomonidan ishlab chiqarilgan nurdan kamuflyaj, o'lja va / yoki turmush o'rtog'ini jalb qilish uchun foydalanadilar. Biolyuminestsent bakteriyalar boshqa organizmlar bilan simbiotik aloqalarni rivojlantirdilar, bunda ikkala ishtirokchi ham bir xil darajada foyda ko'rishadi.[135] Bakteriyalarning lyuminesans reaktsiyasidan foydalanishning yana bir sababi kvorumni aniqlash, bakteriyalar hujayralarining zichligiga javoban gen ekspressionini tartibga solish qobiliyati.[136]
The Gavayi bobtail kalmar biolyuminestsent bakteriyalar bilan simbiozda yashaydi Aliivibrio fischeri kalmar mantiyasida maxsus yorug'lik organida yashaydi. Bakteriyalar shakar bilan oziqlanadi aminokislota kalmar tomonidan va evaziga kalmar siluetini pastdan qaralganda yashirish, qarshi yorituvchi u mantiyaning yuqori qismiga tushadigan yorug'lik miqdoriga mos keladi.[137] Kalmar hayvon-bakterial simbiozi uchun namuna organizm bo'lib xizmat qiladi va uning bakteriyalar bilan aloqasi keng o'rganilgan.
Vibrio harveyi novda shaklida, harakatchan (qutb orqali) flagella ) 30 ° dan 35 ° C gacha optimal darajada o'sadigan biolyuminestsent bakteriya. Tropik dengiz suvlarida erkin suzish mumkin, umuman ichak mikroflorasida dengiz hayvonlar, va ham asosiy, ham opportunistik patogen bir qator dengiz hayvonlari.[138] Buning sababi deb o'ylashadi sutli dengizlarning ta'siri, bu erda dengiz suvidan tunda bir tekis ko'k nur taraladi. Ba'zi yorqinliklar taxminan 6000 kvadrat milni (16000 km) qamrab olishi mumkin2).
Magnetotaksis


Magnetotaktik bakteriyalar bo'ylab o'zlarini yo'naltirish magnit maydon chiziqlari ning Yerning magnit maydoni.[140] Ushbu hizalanma ushbu organizmlarga kislorodning optimal konsentratsiyasi mintaqalariga etib borishiga yordam beradi deb ishoniladi.[141] Ushbu vazifani bajarish uchun ushbu bakteriyalar mavjud biomineralizatsiya qilingan organoidlar deb nomlangan magnetozomalar magnit o'z ichiga oladi kristallar. Atrof muhitning magnit xususiyatlariga javoban harakatlanishga moyil bo'lgan mikroorganizmlarning biologik hodisasi ma'lum magnetotaksis. Biroq, ushbu atama ushbu atamani boshqa har qanday qo'llanilishi bilan chalg'itadi Taksilar rag'batlantirish-javob berish mexanizmini o'z ichiga oladi. Dan farqli o'laroq magnetoreseptsiya bakteriyalar tarkibida magnitlangan magnitlar mavjud bo'lib, ular bakteriyalarni hizalanishga majbur qiladi, hattoki o'lik hujayralar ham xuddi kompas ignasi singari hizalanadi.[141]
Dengiz muhiti odatda oqimlar va turbulentlik bilan barqaror yoki davriy harakatda saqlanadigan ozuqa moddalarining past konsentratsiyasi bilan tavsiflanadi. Dengiz bakteriyalari suzish va yo'naltirilgan sezgir-javob tizimlaridan foydalanib, ozuqa gradiyentlarida qulay joylarga ko'chib o'tish strategiyasini ishlab chiqdi. Magnetotaktik bakteriyalar Yerning magnit maydonidan foydalanib oksid-anoksik interfeysga, ularning turg'unligi va tarqalishi uchun eng qulay joy bo'lgan kimyoviy qatlamli cho'kindilarda yoki suv ustunlarida pastga suzishni osonlashtiradi.[142]
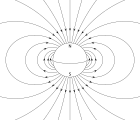
Kengliklariga va bakteriyalar ekvatorning shimolida yoki janubida bo'lishiga qarab, Yer magnit maydoni mumkin bo'lgan ikkita qutblanishdan biriga va yo'nalishi o'zgaruvchan burchak bilan okean tubiga ishora qiladi va umuman kislorodga boy sirtdan uzoqlashadi. . Aerotaksis bakteriyalarning kislorod gradiyentidagi optimal kislorod kontsentratsiyasiga o'tishiga javobdir. Buni turli tajribalar aniq ko'rsatib berdi magnetotaksis va aerotaksis magnetotaktik bakteriyalarda birgalikda ishlaydi. Suv tomchilarida bir tomonlama suzish magnetotaktik bakteriyalar suzish yo'nalishini o'zgartirib, ostida orqaga suzishlari mumkinligi ko'rsatilgan. kamaytirish sharoitlar (optimaldan kam) kislorod kontsentratsiyasi ), oksik sharoitlardan farqli o'laroq (optimal kislorod kontsentratsiyasidan kattaroq).
Morfologiyasidan qat'i nazar, shu paytgacha o'rganilgan barcha magnetotaktik bakteriyalar flagella yordamida harakatchan.[143] Ayniqsa, dengiz magnetotaktik bakteriyalari o'n minglab flagellarni o'z ichiga olishi mumkin bo'lgan aniq flagellar apparatiga ega. Biroq, so'nggi yillarda olib borilgan keng qamrovli izlanishlarga qaramay, magnetotaktik bakteriyalar magnit maydonlarga mos kelishiga javoban o'zlarining flagellar motorlarini boshqaradimi yoki yo'qmi hali aniqlanmagan.[142] Simbiyoz magnetotaktik bakteriyalar bilan tushuntirish sifatida taklif qilingan magnetoreseptsiya ba'zilarida dengiz protistlari.[144] Magnetoreseptsiyaga o'xshash munosabatlar asos bo'lishi mumkinligi to'g'risida tadqiqotlar olib borilmoqda umurtqali hayvonlar shuningdek.[145] Eng qadimgi aniq magnetofosilalar quyidagilardan kelib chiqqan Bo'r janubiy Angliyaning bo'r yotoqlari,[146] magnetofosillarning kamroq ma'lum hisobotlari 1,9 milliard yilgacha tarqaladi Gunflint Chert.[147]
Mikrobial rodopsin

(2) u konfiguratsiyani o'zgartiradi, shuning uchun proton hujayradan chiqarib yuboriladi
(3) kimyoviy potentsial protonning hujayraga qaytishiga olib keladi
(4) shunday qilib energiya ishlab chiqaradi
(5) shaklida adenozin trifosfat.[148]
Fototrofik metabolizm energiyani o'zgartiradigan uchta pigmentdan biriga asoslanadi: xlorofill, bakterioxlorofil va setchatka. Retinal bu xromofor ichida topilgan rodopsinlar. Xlorofillning yorug'lik energiyasini konvertatsiya qilishdagi ahamiyati o'nlab yillar davomida yozilgan, ammo retinal pigmentlarga asoslangan fototrofiya endigina o'rganilmoqda.[149]

2000 yilda boshchiligidagi mikrobiologlar jamoasi Edvard DeLong dengiz uglerod va energiya davrlarini tushunishda hal qiluvchi kashfiyot qildi. Ular bir necha turdagi bakteriyalarda genni topdilar[151][152] oqsil ishlab chiqarish uchun javobgardir rodopsin, ilgari bakteriyalarda eshitilmagan. Hujayra membranalarida mavjud bo'lgan bu oqsillar nur nurini biokimyoviy energiyaga aylantirishga qodir, chunki quyosh nurlari tushishi bilan rodopsin molekulasi konfiguratsiyasi o'zgaradi va proton ichkaridan va energiya ishlab chiqaradigan keyingi oqim.[153] Arxeoalga o'xshash rodopsinlar keyinchalik turli taksonlar, protistlar, shuningdek bakteriyalar va arxeylar orasida topilgan, ammo ular kompleksda kamdan-kam uchraydi. ko'p hujayrali organizmlar.[154][155][156]
2019 yildagi tadqiqotlar shuni ko'rsatadiki, bu "quyoshni yutib yuboradigan bakteriyalar" ilgari o'ylanganidan ko'ra kengroq tarqalgan va global isish ta'sirida okeanlar qanday ta'sir qilishi mumkin. "Topilmalar dengiz ekologiyasining darsliklarda keltirilgan an'anaviy talqinidan kelib chiqadi, unda okeandagi deyarli barcha quyosh nurlari xlorofill tomonidan suv o'tlarida olinadi. Buning o'rniga rodopsin bilan jihozlangan bakteriyalar gibrid avtomobillar kabi ishlaydi, mavjud bo'lganda organik moddalar bilan ishlaydi. aksariyat bakteriyalar - va ozuqa moddalari kam bo'lganida quyosh nurlari ta'sirida. "[157][149]
Bor astrobiologik deb nomlangan taxmin Binafsha Yer gipotezasi Yerdagi asl hayot shakllari xlorofillga emas, balki retinaga asoslangan bo'lib, bu Yerni yashil o'rniga binafsha rangga aylantiradi deb taxmin qilmoqda.[158][159]
Simbiyoz
Ba'zi dengiz organizmlarida a simbiyoz bakteriyalar yoki arxa bilan. Pompei qurtlari 80 ° S gacha bo'lgan haroratda gidrotermal shamollatish orqali katta chuqurlikda yashaydi. Ularda tukli ko'rinadigan narsa bor, ammo bu "sochlar" aslida bakteriyalar kabi koloniyalardir Nautilia profundicola, bu qurtni ma'lum darajada izolyatsiyalashga qodir deb o'ylashadi. Qurt orqasidagi bezlar bakteriyalar oziqlanadigan shilimshiqni chiqaradi, bu shakl simbiyoz.

"Tukli" orqa Pompei qurtlari simbiyotik bakteriyalar koloniyalaridir

Hesiocaeca methanicola chuqurlikda yashaydi metan muzi va bakteriyalar bilan simbiozda omon qoladi metabolizm The klatrat.[160]

Olavius algarvensis uning oziqlanishi uchun simbiyotik bakteriyalarning beshta turiga bog'liq


Endosimbiont bakteriyalar - bu organizm yoki boshqa organizm hujayralarida yashaydigan bakteriyalar. Siyanobakteriyalarning ayrim turlari endosimbiont siyanobakteriyalarda azot fiksatsiyasini o'tashga imkon beradigan genlarga ega ekanligi aniqlandi.[161]
Organizmlar odatda simbiyotik munosabatlarni o'zlarining yashash joylarida resurslarning cheklanganligi yoki oziq-ovqat manbalarining cheklanganligi sababli o'rnatadilar. Midiya bilan bog'liq bo'lgan kashf etilgan simbiyotik, xemosintetik bakteriyalar (Batimodiolus ) located near hydrothermal vents have a gene that enables them to utilize hydrogen as a source of energy, in preference to sulphur or methane as their energy source for production of energy.[162]
Olavius algarvensis is a worm which lives in coastal sediments in the Mediterranean and depends on symbiotic bacteria for its nutrition. It lives with five different species of bacteria located under its cuticle: two sulfide-oxidizing , ikkitasi sulfate-reducing va bitta spirochaete. The symbiotic bacteria also allow the worm to use vodorod va uglerod oksidi as energy sources, and to metabolise organic compounds like malate va atsetat.[163][164]
Roles in marine food webs
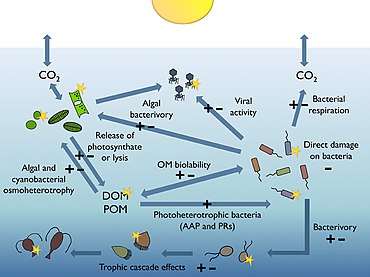
Most of the volume of the world ocean is in darkness. The processes occurring within the thin illuminated surface layer (the photic layer from the surface down to between 50 and 170 metres) are of major significance to the global biosphere. For example, the visible region of the solar spectrum (the so-called photosynthetically available radiation or PAR) reaching this sunlit layer fuels about half of the birlamchi mahsuldorlik of the planet, and is responsible for about half of the atmospheric oxygen necessary for most life on Earth.[166][167]
Geterotrofik bakterioplankton are main consumers of dissolved organic matter (DOM) in pelagic marine food webs, including the sunlit upper layers of the ocean. Their sensitivity to ultraviolet radiation (UVR), together with some recently discovered mechanisms bacteria have evolved to benefit from photosynthetically available radiation (PAR), suggest that natural sunlight plays a relevant, yet difficult to predict role in modulating bacterial biogeochemical functions in the oceans.[165]
Ocean surface habitats sit at the interface between the atmosphere and the ocean. The biofilm-like habitat at the surface of the ocean harbours surface-dwelling microorganisms, commonly referred to as neuston. This vast air–water interface sits at the intersection of major air–water exchange processes spanning more than 70% of the global surface area . Bacteria in the surface microlayer of the ocean, called bacterioneuston, are of interest due to practical applications such as air-sea gas exchange of greenhouse gases, production of climate-active marine aerosols, and remote sensing of the ocean.[168] Of specific interest is the production and degradation of sirt faol moddalar (surface active materials) via microbial biochemical processes. Major sources of surfactants in the open ocean include phytoplankton,[169] terrestrial runoff, and deposition from the atmosphere.[168]

Unlike coloured algal blooms, surfactant-associated bacteria may not be visible in ocean colour imagery. Having the ability to detect these "invisible" surfactant-associated bacteria using sintetik diafragma radar has immense benefits in all-weather conditions, regardless of cloud, fog, or daylight.[168] This is particularly important in very high winds, because these are the conditions when the most intense air-sea gas exchanges and marine aerosol production take place. Therefore, in addition to colour satellite imagery, SAR satellite imagery may provide additional insights into a global picture of biophysical processes at the boundary between the ocean and atmosphere, air-sea greenhouse gas exchanges and production of climate-active marine aerosols.[168]
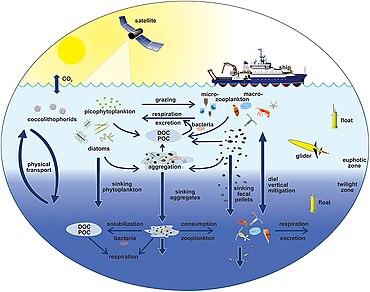
The diagram on the right shows links among the ocean's biologik nasos and the pelagic food web and the ability to sample these components remotely from ships, satellites, and autonomous vehicles. Light blue waters are the eyfotik zona, while the darker blue waters represent the alacakaranlık zonasi.[170]
Roles in biogeochemical cycling
Archaea recycle elements such as uglerod, azot va oltingugurt through their various habitats.[171] Archaea carry out many steps in the azot aylanishi. This includes both reactions that remove nitrogen from ecosystems (such as nitrat -based respiration and denitrifikatsiya ) as well as processes that introduce nitrogen (such as nitrate assimilation and azot fiksatsiyasi ).[172][173]
Researchers recently discovered archaeal involvement in ammiak oxidation reactions. These reactions are particularly important in the oceans.[174][175] In oltingugurt aylanishi, archaea that grow by oxidizing oltingugurt compounds release this element from rocks, making it available to other organisms, but the archaea that do this, such as Sulfolobus, mahsulot sulfat kislota as a waste product, and the growth of these organisms in abandoned mines can contribute to kislota konini drenajlash and other environmental damage.[176] In uglerod aylanishi, methanogen archaea remove hydrogen and play an important role in the decay of organic matter by the populations of microorganisms that act as parchalovchilar in anaerobic ecosystems, such as sediments and marshes.[177]
Shuningdek qarang
- Bioluminescent bacteria
- Pelagibakteralar – model organisms in streamlining theory
- Nazariyani soddalashtirish
Adabiyotlar
- ^ Youngson RM (2006). Collins Dictionary of Human Biology. Glazgo: HarperCollins. ISBN 978-0-00-722134-9.
- ^ Nelson DL, Cox MM (2005). Lehninger Biokimyo tamoyillari (4-nashr). Nyu-York: W.H. Freeman. ISBN 978-0-7167-4339-2.
- ^ Martin EA, ed. (1983). Macmillan Dictionary of Life Sciences (2-nashr). London: Macmillan Press. ISBN 978-0-333-34867-3.
- ^ Fuerst JA (2010). "Beyond Prokaryotes and Eukaryotes: Planctomycetes and Cell Organization". Tabiatni o'rganish. 3 (9): 44.
- ^ 2002 WHO mortality data Accessed 20 January 2007
- ^ University of Georgia (10 December 2015). "Functions of global ocean microbiome key to understanding environmental changes". www.sc tajribali.com. Olingan 11 dekabr 2015.
- ^ Bar-On YM, Phillips R, Milo R (2018). "The biomass distribution on Earth" (PDF). Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 115 (25): 6506–6511. Bibcode:1998 PNAS ... 95.6578W. doi:10.1073 / pnas.1711842115. PMC 6016768. PMID 29784790.
- ^ a b Doglioni C, Pignatti J, Coleman M (2016). "Why did life develop on the surface of the Earth in the Cambrian?". Geoscience Frontiers. 7 (6): 865–873. doi:10.1016/j.gsf.2016.02.001.
- ^ "Er yoshi". Amerika Qo'shma Shtatlarining Geologik xizmati. 2007 yil 9-iyul. Olingan 2015-05-31.
- ^ Dalrymple 2001, pp. 205–221
- ^ Manhesa G, Allègre CJ, Dupréa B, Hamelin B (May 1980). "Lead isotope study of basic-ultrabasic layered complexes: Speculations about the age of the earth and primitive mantle characteristics". Yer va sayyora fanlari xatlari. 47 (3): 370–382. Bibcode:1980E&PSL..47..370M. doi:10.1016/0012-821X(80)90024-2.
- ^ Schopf JW, Kudryavtsev AB, Czaja AD, Tripathi AB (5 October 2007). "Evidence of Archean life: Stromatolites and microfossils". Prekambriyen tadqiqotlari. 158 (3–4): 141–155. Bibcode:2007 yil PRER..158..141S. doi:10.1016 / j.precamres.2007.04.009.
- ^ Raven & Johnson 2002, p. 68
- ^ Baumgartner RJ, et al. (2019). "Nano−porous pyrite and organic matter in 3.5-billion-year-old stromatolites record primordial life". Geologiya. 47 (11): 1039–1043. Bibcode:2019Geo....47.1039B. doi:10.1130/G46365.1.
- ^ Earliest signs of life: Scientists find microbial remains in ancient rocks Phys.org. 26 sentyabr 2019 yil.
- ^ Jablonski D (25 June 1999). "The Future of the Fossil Record". Ilm-fan. 284 (5423): 2114–2116. doi:10.1126 / science.284.5423.2114. PMID 10381868.
- ^ a b Cavalier-Smith T (2006 yil 29 iyun). "Cell evolution and Earth history: stasis and revolution". Qirollik jamiyatining falsafiy operatsiyalari B: Biologiya fanlari. 361 (1470): 969–1006. doi:10.1098/rstb.2006.1842. PMC 1578732. PMID 16754610.
- ^ Schopf JW (2006 yil 29 iyun). "Fossil evidence of Archaean life". Qirollik jamiyatining falsafiy operatsiyalari B: Biologiya fanlari. 361 (1470): 869–885. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604.
- Altermann W, Kazmierczak J (November 2003). "Arxey mikrofosillalari: Yerdagi dastlabki hayotni qayta baholash". Mikrobiologiya bo'yicha tadqiqotlar. 154 (9): 611–617. doi:10.1016 / j.resmic.2003.08.006. PMID 14596897.
- ^ Schopf JW (19 July 1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 91 (15): 6735–6742. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. PMC 44277. PMID 8041691.
- ^ Poole AM, Penny D (January 2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. PMID 17187354.
- ^ a b Dyall SD, Brown MT, Johnson PJ (2004 yil 9 aprel). "Ancient Invasions: From Endosymbionts to Organelles". Ilm-fan. 304 (5668): 253–257. Bibcode:2004Sci...304..253D. doi:10.1126/science.1094884. PMID 15073369.
- ^ Martin W (October 2005). "The missing link between hydrogenosomes and mitochondria". Mikrobiologiya tendentsiyalari. 13 (10): 457–459. doi:10.1016/j.tim.2005.08.005. PMID 16109488.
- ^ Lang BF, Gray MW, Burger G (December 1999). "Mitochondrial genome evolution and the origin of eukaryotes". Genetika fanining yillik sharhi. 33: 351–397. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
- McFadden GI (1 December 1999). "Endosymbiosis and evolution of the plant cell". O'simliklar biologiyasidagi hozirgi fikr. 2 (6): 513–519. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659.
- ^ Ciccarelli FD, Doerks T, von Mering C, et al. (3 March 2006). "Toward Automatic Reconstruction of a Highly Resolved Tree of Life". Ilm-fan. 311 (5765): 1283–1287. Bibcode:2006 yil ... 311.1283C. CiteSeerX 10.1.1.381.9514. doi:10.1126 / science.1123061. PMID 16513982.
- ^ DeLong EF, Pace NR (2001 yil 1-avgust). "Environmental Diversity of Bacteria and Archaea". Tizimli biologiya. 50 (4): 470–478. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Kaiser D (2001 yil dekabr). "Building a multicellular organism". Genetika fanining yillik sharhi. 35: 103–123. doi:10.1146 / annurev.genet.35.102401.090145. PMID 11700279.
- ^ Zimmer C (2016 yil 7-yanvar). "Genetik flip organizmlarning bir hujayradan ko'plarga o'tishiga yordam berdi". The New York Times. Olingan 7 yanvar 2016.
- ^ Valentine JW, Jablonski D, Erwin DH (1 March 1999). "Fossils, molecules and embryos: new perspectives on the Cambrian explosion". Rivojlanish. 126 (5): 851–859. PMID 9927587. Olingan 2014-12-30.
- ^ Ohno S (January 1997). "The reason for as well as the consequence of the Cambrian explosion in animal evolution". Molekulyar evolyutsiya jurnali. 44 (Suppl. 1): S23–S27. Bibcode:1997JMolE..44S..23O. doi:10.1007/PL00000055. PMID 9071008.
- Valentine JW, Jablonski D (2003). "Morphological and developmental macroevolution: a paleontological perspective". Rivojlanish biologiyasining xalqaro jurnali. 47 (7–8): 517–522. PMID 14756327. Olingan 2014-12-30.
- ^ Campbell, N. "Biology:Concepts & Connections". Pearson ta'limi. San Francisco: 2003.
- ^ Xarper, Duglas. "prokaryote". Onlayn etimologiya lug'ati.
- ^ Xarper, Duglas. "eukaryotic". Onlayn etimologiya lug'ati.
- ^ Stanier RY, Van Niel CB (1962). "Bakteriya to'g'risida tushuncha". Archiv für Mikrobiologie. 42: 17–35. doi:10.1007/BF00425185. PMID 13916221.
- ^ Woese CR, Fox GE (1977 yil noyabr). "Prokaryotik domenning filogenetik tuzilishi: asosiy shohliklar". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 74 (11): 5088–90. Bibcode:1977 PNAS ... 74.5088W. doi:10.1073 / pnas.74.11.5088. PMC 432104. PMID 270744.
- ^ Woese CR, Kandler O, Wheelis ML (1990 yil iyun). "Organizmlarning tabiiy tizimiga: Arxeya, Bakteriyalar va Evkarya domenlariga taklif". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 87 (12): 4576–9. Bibcode:1990PNAS ... 87.4576W. doi:10.1073 / pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ Ohtomo, Yoko; Kakegava, Takeshi; Ishida, Akizumi; Nagase, Toshiro; Rosing, Minik T. (January 2014). "Dastlabki Arxey Isuasi metasentiment jinslarida biogen grafitga dalillar". Tabiatshunoslik. 7 (1): 25–28. Bibcode:2014 yil NatGe ... 7 ... 25O. doi:10.1038 / ngeo2025.
- ^ Nutman, Allen P.; Bennett, Vickie C.; Friend, Clark R. L.; Kranendonk, Martin J. Van; Chivas, Allan R. (September 2016). "Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures". Tabiat. 537 (7621): 535–538. Bibcode:2016Natur.537..535N. doi:10.1038 / nature19355. PMID 27580034.
- ^ Borenshteyn, Set (19 oktyabr 2015). "Erning ilk qismida xarob bo'lgan deb hisoblangan hayot haqidagi maslahatlar". Ajoyib. Yonkers, NY: Mindspark interaktiv tarmog'i. Associated Press. Arxivlandi asl nusxasi 2015 yil 23 oktyabrda. Olingan 8 oktyabr 2018.
- ^ Bell, Elizabeth A.; Boehnike, Patrik; Xarrison, T. Mark; va boshq. (2015 yil 19 oktyabr). "4,1 milliard yillik tsirkonda saqlanib qolishi mumkin bo'lgan biogen uglerod". Proc. Natl. Akad. Ilmiy ish. AQSH. 112 (47): 14518–21. Bibcode:2015PNAS..11214518B. doi:10.1073 / pnas.1517557112. PMC 4664351. PMID 26483481.
- ^ Dodd, Metyu S.; Papinyo, Dominik; Grenne, Tor; sust, Jon F.; Rittner, Martin; Pirajno, Franko; O'Nil, Jonatan; Kichkina, Krispin T. S. (2017 yil 2 mart). "Yerning eng qadimgi gidrotermal shamollatishida erta hayot uchun dalillar cho'kindi" (PDF). Tabiat. 543 (7643): 60–64. Bibcode:2017 yil Noyabr 543 ... 60D. doi:10.1038 / tabiat21377. PMID 28252057.
- ^ Zimmer, Karl (2017 yil 1 mart). "Olimlar Kanada bakteriyalarining qoldiqlari Yerdagi eng qadimgi bo'lishi mumkin". The New York Times. Olingan 2 mart 2017.
- ^ Nisbet, Euan G.; Fowler, C. M. R. (1999 yil 7-dekabr). "Mikrobial paspaslarning arxe metabolik evolyutsiyasi". London Qirollik jamiyati materiallari B. 266 (1436): 2375–2382. doi:10.1098 / rspb.1999.0934. PMC 1690475.
- ^ Anbar, Ariel D.; Yun Duan; Lyons, Timothy W.; va boshq. (2007 yil 28 sentyabr). "A Whiff of Oxygen Before the Great Oxidation Event?". Ilm-fan. 317 (5846): 1903–1906. Bibcode:2007Sci...317.1903A. doi:10.1126/science.1140325. PMID 17901330.
- ^ Knoll, Endryu H.; Javaux, Emmanuel J.; Hewitt, David; Koen, Fib (2006 yil 29 iyun). "Proterozoy okeanidagi eukaryotik organizmlar". Qirollik jamiyatining falsafiy operatsiyalari B. 361 (1470): 1023–1038. doi:10.1098 / rstb.2006.1843. PMC 1578724. PMID 16754612.
- ^ Fedonkin, Mixail A. (2003 yil 31 mart). "Metazoa kelib chiqishi proterozoy davridagi qoldiq qoldiqlari asosida" (PDF). Paleontologik tadqiqotlar. 7 (1): 9–41. doi:10.2517 / prpsj.7.9. Arxivlandi asl nusxasi (PDF) 2009 yil 26 fevralda. Olingan 2008-09-02.
- ^ Bonner, Jon Tayler (1998). "Ko'p hujayralilikning kelib chiqishi". Integrativ biologiya. 1 (1): 27–36. doi:10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6.
- ^ May, R.M. (1988) "How many species are there on earth?". Ilm-fan, 241(4872): 1441–1449. doi:10.1126 / science.241.4872.1441.
- ^ Locey, K.J. and Lennon, J.T. (2016) "Scaling laws predict global microbial diversity". Milliy fanlar akademiyasi materiallari, 113(21): 5970–5975. doi:10.1073/pnas.1521291113.
- ^ Vitorino, L.C. and Bessa, L.A. (2018) "Microbial diversity: the gap between the estimated and the known". Turli xillik, 10(2): 46. doi:10.3390/d10020046.
 Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi. - ^ Tirik bakteriyalar Yerdagi havo oqimlarini boshqarmoqda Smithsonian jurnali, 11 January 2016.
- ^ Robbins, Jim (2018 yil 13-aprel). "Osmondan har kuni trillionlab viruslar tushadi". The New York Times. Olingan 14 aprel 2018.
- ^ Rexe, Izabel; D'Orta, Gaetano; Mladenov, Natali; Winget, Danielle M; Suttle, Kertis A (29-yanvar, 2018-yil). "Viruslar va bakteriyalarning atmosfera chegarasi qatlamidan yuqori bo'lish darajasi". ISME jurnali. 12 (4): 1154–1162. doi:10.1038 / s41396-017-0042-4. PMC 5864199. PMID 29379178.
- ^ Suttle, C.A. (2005). "Viruses in the Sea". Tabiat. 437 (9): 356–361. Bibcode:2005Natur.437..356S. doi:10.1038/nature04160. PMID 16163346.
- ^ Shors 2017, p. 5
- ^ Shors 2017, p. 593
- ^ Suttle CA. Marine viruses—major players in the global ecosystem. Tabiat sharhlari Mikrobiologiya. 2007;5(10):801–12. doi:10.1038/nrmicro1750. PMID 17853907.
- ^ Durham, Bryndan P.; Grote, Jana; Whittaker, Kerry A.; Bender, Sara J.; Luo, Haiwei; Grim, Sharon L.; Braun, Julia M.; Casey, John R.; Dron, Antony; Florez-Leiva, Lennin; Krupke, Andreas; Luria, Catherine M.; Mine, Aric H.; Nigro, Olivia D.; Pather, Santhiska; Talarmin, Agathe; Wear, Emma K.; Weber, Thomas S.; Wilson, Jesse M.; Church, Matthew J.; Delong, Edvard F.; Karl, David M.; Steward, Grieg F.; Eppley, John M.; Kirpides, Nikos S.; Schuster, Stephan; Rappé, Michael S. (2014). "Draft genome sequence of marine alphaproteobacterial strain HIMB11, the first cultivated representative of a unique lineage within the Roseobacter clade possessing an unusually small genome". Genomik fanlarning standartlari. 9 (3): 632–645. doi:10.4056/sigs.4998989. PMC 4148974. PMID 25197450.
- ^ Fredrickson JK, Zachara JM, Balkwill DL, Kennedy D, Li SM, Kostandarithes HM, Daly MJ, Romine MF, Brockman FJ (2004). "Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the Hanford site, Washington state". Amaliy va atrof-muhit mikrobiologiyasi. 70 (7): 4230–41. doi:10.1128/AEM.70.7.4230-4241.2004. PMC 444790. PMID 15240306.
- ^ Woese CR, Kandler O, Wheelis ML (1990). "Organizmlarning tabiiy tizimiga: Arxeya, Bakteriyalar va Evkarya domenlariga taklif". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 87 (12): 4576–9. Bibcode:1990PNAS ... 87.4576W. doi:10.1073 / pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ Schopf JW (1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 91 (15): 6735–42. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. PMC 44277. PMID 8041691.
- ^ DeLong EF, Pace NR (2001). "Environmental diversity of bacteria and archaea". Tizimli biologiya. 50 (4): 470–8. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Brown JR, Doolittle WF (1997). "Archaea and the prokaryote-to-eukaryote transition". Mikrobiologiya va molekulyar biologiya sharhlari. 61 (4): 456–502. doi:10.1128/.61.4.456-502.1997. PMC 232621. PMID 9409149.
- ^ Poole AM, Penny D (2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. PMID 17187354.
- ^ Lang BF, Gray MW, Burger G (1999). "Mitochondrial genome evolution and the origin of eukaryotes". Genetika fanining yillik sharhi. 33: 351–97. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
- ^ McFadden GI (1999). "Endosymbiosis and evolution of the plant cell". O'simliklar biologiyasidagi hozirgi fikr. 2 (6): 513–9. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659.
- ^ Koch AL (2002). "Control of the bacterial cell cycle by cytoplasmic growth". Mikrobiologiyadagi tanqidiy sharhlar. 28 (1): 61–77. doi:10.1080/1040-840291046696. PMID 12003041.
- ^ Eagon RG (April 1962). "Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes". Bakteriologiya jurnali. 83 (4): 736–37. doi:10.1128/jb.83.4.736-737.1962. PMC 279347. PMID 13888946.
- ^ "Candidatus Pelagibacter Ubique." Evropa bioinformatika instituti. European Bioinformatics Institute, 2011. Web. 08 Jan. 2012. http://www.ebi.ac.uk/2can/genomes/bacteria/Candidatus_Pelagibacter_ubique.html Arxivlandi 2008 yil 1-dekabr, soat Orqaga qaytish mashinasi
- ^ "Flea market: A newly discovered virus may be the most abundant organism on the planet". Iqtisodchi. 2013 yil 16-fevral. Olingan 16 fevral 2013.
- ^ Chjao, Y .; Temperton, B.; Thrash, J. C .; Shvalbax, M. S .; Vergin, K. L .; Landry, Z. C.; Ellisman, M.; Deerinck, T.; Sullivan, M. B.; Giovannoni, S. J. (2013). "Abundant SAR11 viruses in the ocean". Tabiat. 494 (7437): 357–360. Bibcode:2013Natur.494..357Z. doi:10.1038/nature11921. PMID 23407494.
- ^ Bentzon-Tilia M, Gram L (2017). Biologik qidiruv. Topics in Biodiversity and Conservation. Springer, Xam. 137–166 betlar. doi:10.1007/978-3-319-47935-4_7. ISBN 978-3-319-47933-0.
- ^ NCBI Taxonomy Browser: Roseobacter Milliy Biotexnologiya Axborot Markazi. Accessed: 8 May 2020.
- ^ "The largest Bacterium: Scientist discovers new bacterial life form off the African coast", Maks Plank nomidagi dengiz mikrobiologiyasi instituti, 8 April 1999, archived from asl nusxasi 2010 yil 20 yanvarda
- ^ List of Prokaryotic names with Standing in Nomenclature - Genus Thiomargarita
- ^ Changes in oxygen concentrations in our ocean can disrupt fundamental biological cycles Phys.org, 25 November 2019.
- ^ Walsh PJ, Smith S, Fleming L, Solo-Gabriele H, Gerwick WH, eds. (2011 yil 2 sentyabr). "Cyanobacteria and cyanobacterial toxins". Oceans and Human Health: Risks and Remedies from the Seas. Akademik matbuot. 271–296 betlar. ISBN 978-0-08-087782-2.
- ^ "The Rise of Oxygen - Astrobiology Magazine". Astrobiologiya jurnali. 2003 yil 30-iyul. Olingan 2016-04-06.
- ^ Flannery, D. T.; R.M. Walter (2012). "Archean tufted microbial mats and the Great Oxidation Event: new insights into an ancient problem". Avstraliya Yer fanlari jurnali. 59 (1): 1–11. Bibcode:2012AuJES..59....1F. doi:10.1080/08120099.2011.607849.
- ^ Rotshild, Lin (2003 yil sentyabr). "Hayotning evolyutsion mexanizmlari va atrof-muhit chegaralarini tushunib oling". NASA. Arxivlandi asl nusxasi 2012 yil 11 martda. Olingan 13 iyul 2009.
- ^ Patrick J. Keeling (2004). "Diversity and evolutionary history of plastids and their hosts". Amerika botanika jurnali. 91 (10): 1481–1493. doi:10.3732 / ajb.91.10.1481. PMID 21652304.
- ^ Nadis S (December 2003). "The cells that rule the seas" (PDF). Ilmiy Amerika. 289 (6): 52–3. Bibcode:2003SciAm.289f..52N. doi:10.1038/scientificamerican1203-52. PMID 14631732. Arxivlandi asl nusxasi (PDF) 2014 yil 19 aprelda. Olingan 2 iyun 2019.
- ^ "The Most Important Microbe You've Never Heard Of". npr.org.
- ^ Flombaum, P.; Gallegos, J. L.; Gordillo, R. A.; Rincon, J.; Zabala, L. L.; Jiao, N.; Karl, D. M .; Li, W. K. W.; Lomas, M. V.; Veneziano, D.; Vera, C. S.; Vrugt, J. A.; Martiny, A. C. (2013). "Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus". Milliy fanlar akademiyasi materiallari. 110 (24): 9824–9829. Bibcode:2013PNAS..110.9824F. doi:10.1073/pnas.1307701110. PMC 3683724. PMID 23703908.
- ^ Nabors, Murray W. (2004). Botanika faniga kirish. San Francisco, CA: Pearson Education, Inc. ISBN 978-0-8053-4416-5.
- ^ Allaby, M., ed. (1992). "Algae". The Concise Dictionary of Botany. Oksford: Oksford universiteti matbuoti.
- ^ Bang C, Schmitz RA (2015). "Archaea associated with human surfaces: not to be underestimated". FEMS Mikrobiologiya sharhlari. 39 (5): 631–48. doi:10.1093/femsre/fuv010. PMID 25907112.
- ^ Arxeya Onlayn etimologiya lug'ati. Qabul qilingan 17 avgust 2016 yil.
- ^ Pace NR (May 2006). "Time for a change". Tabiat. 441 (7091): 289. Bibcode:2006 yil natur.441..289P. doi:10.1038 / 441289a. PMID 16710401.
- ^ Stoeckenius W (1 October 1981). "Walsby's square bacterium: fine structure of an orthogonal procaryote". Bakteriologiya jurnali. 148 (1): 352–60. doi:10.1128/JB.148.1.352-360.1981. PMC 216199. PMID 7287626.
- ^ Madigan M, Martinko J, nashr. (2005). Mikroorganizmlarning Brok biologiyasi (11-nashr). Prentice Hall. ISBN 978-0-13-144329-7.
- ^ Xuber, Xarald; va boshq. (2002). "Arxeyaning yangi filomi nanozlangan gipertermofil simbiont bilan ifodalangan". Tabiat. 417 (6884): 63–67. Bibcode:2002 yil Tabiat 417 ... 63H. doi:10.1038 / 417063a. PMID 11986665.
- ^ Brochier, Celine; Gribaldo, S; Zivanovich, Y; Konfalonieri, F; va boshq. (2005). "Nanoarxeya: yangi arxeologik filim vakillari yoki termokokkalarga aloqador tez rivojlanayotgan evarxeal naslmi?". Genom biologiyasi. 6 (5): R42. doi:10.1186 / gb-2005-6-5-r42. PMC 1175954. PMID 15892870.
- ^ Orellana, Luis H.; Ben Francis, T.; Krüger, Karen; Teeling, Hanno; Müller, Marie-Caroline; Fuchs, Bernhard M.; Konstantinidis, Konstantinos T.; Amann, Rudolf I. (2019). "Niche differentiation among annually recurrent coastal Marine Group II Euryarchaeota". ISME jurnali. 13 (12): 3024–3036. doi:10.1038/s41396-019-0491-z. PMC 6864105. PMID 31447484.
- ^ See especially Fig. 4 in Nishimura, Yosuke; Watai, Hiroyasu; Honda, Takashi; Mihara, Tomoko; Omae, Kimiho; Rou, Simon; Blan-Matye, Romain; Yamamoto, Keigo; Hingamp, Pascal; Sako, Yoshihiko; Sullivan, Matthew B.; Goto, Susumu; Ogata, Xiroyuki; Yoshida, Takashi (2017). "Environmental Viral Genomes Shed New Light on Virus-Host Interactions in the Ocean". mSphere. 2 (2). doi:10.1128/mSphere.00359-16. PMC 5332604. PMID 28261669.
- ^ Philosof, Alon; Yutin, Natalya; Flores-Uribe, José; Sharon, Itai; Koonin, Evgeniy V.; Béjà, Oded (2017). "Novel Abundant Oceanic Viruses of Uncultured Marine Group II Euryarchaeota". Hozirgi biologiya. 27 (9): 1362–1368. doi:10.1016/j.cub.2017.03.052. PMC 5434244. PMID 28457865.
- ^ Xia, Xiaomin; Guo, Wang; Liu, Hongbin (2017). "Basin Scale Variation on the Composition and Diversity of Archaea in the Pacific Ocean". Mikrobiologiya chegaralari. 8: 2057. doi:10.3389/fmicb.2017.02057. PMC 5660102. PMID 29109713.
- ^ Martin-Cuadrado, Ana-Belen; Garcia-Heredia, Inmaculada; Moltó, Aitor Gonzaga; López-Úbeda, Rebeca; Kimes, Nikole; López-García, Purificación; Moreira, Devid; Rodriguez-Valera, Francisco (2015). "A new class of marine Euryarchaeota group II from the mediterranean deep chlorophyll maximum". ISME jurnali. 9 (7): 1619–1634. doi:10.1038/ismej.2014.249. PMC 4478702. PMID 25535935.
- ^ NCBI: Candidatus Poseidoniales (buyurtma)
- ^ NCBI: Marine Group III
- ^ NCBI: Marine Group IV
- ^ Zillig W (December 1991). "Comparative biochemistry of Archaea and Bacteria". Genetika va rivojlanish sohasidagi dolzarb fikrlar. 1 (4): 544–51. doi:10.1016/S0959-437X(05)80206-0. PMID 1822288.
- ^ Slonczewski JL, Foster JW. Microbiology: An Evolving Science (3 nashr). WW Norton & Company. pp. 491–44.
- ^ Hogan CM (2010). "Extremophile". In Monosson E, Cleveland C (eds.). Yer entsiklopediyasi. National Council of Science & the Environment.
- ^ Yurkov, V. V.; Beatty, J. T. (1998). "Aerobic anoxygenic phototrophic bacteria". Mikrobiologiya va molekulyar biologiya sharhlari. 62 (3): 695–724. doi:10.1128/MMBR.62.3.695-724.1998. PMC 98932. PMID 9729607.
- ^ Lami, R.; Cottrell, M. T.; Ras, J.; Ulloa, O.; Obernosterer, I.; Claustre, H.; Kirchman, D. L.; Lebaron, P. (2007). "High Abundances of Aerobic Anoxygenic Photosynthetic Bacteria in the South Pacific Ocean". Amaliy va atrof-muhit mikrobiologiyasi. 73 (13): 4198–205. doi:10.1128/AEM.02652-06. PMC 1932784. PMID 17496136.
- ^ Life science weekly. (2012). Bacteria; Reports from Spanish National Research Council (CSIC) Describe Recent Advances in Bacteria. ISSN 1552-2466. P.4582.
- ^ Jiao, Nianzhi; Chjan, Yao; Zeng, Yonghui; Hong, Ning; Liu, Rulong; Chen, Feng; Wang, Pinxian (2007). "Distinct distribution pattern of abundance and diversity of aerobic anoxygenic phototrophic bacteria in the global ocean". Atrof-muhit mikrobiologiyasi. 9 (12): 3091–3099. doi:10.1111/j.1462-2920.2007.01419.x. PMID 17991036.
- ^ Lami, Raphaël; Cottrell, Matthew T.; Ras, JoséPhine; Ulloa, Osvaldo; Obernosterer, Ingrid; Klaustr, Erve; Kirchman, David L.; Lebaron, Philippe (2007). "High Abundances of Aerobic Anoxygenic Photosynthetic Bacteria in the South Pacific Ocean". Amaliy va atrof-muhit mikrobiologiyasi. 73 (13): 4198–4205. doi:10.1128/AEM.02652-06. PMID 17496136.
- ^ Kolber, Z. S. (2001). "Contribution of Aerobic Photoheterotrophic Bacteria to the Carbon Cycle in the Ocean". Ilm-fan. 292 (5526): 2492–2495. doi:10.1126/science.1059707. PMID 11431568.
- ^ Aragno M, Schlegel HG (1981). "The Hydrogen-Oxidizing Bacteria". In Starr MP, Stolp H, Trüper HG, Balows A, Schlegel HG (eds.). The Prokaryotes. Berlin, Geydelberg: Springer. pp. 865–893. doi:10.1007/978-3-662-13187-9_70. ISBN 978-3-662-13187-9.
- ^ a b Albers SV, Jarrell KF (2015 yil 27-yanvar). "Archaellum: Arxeya qanday suzadi". Mikrobiologiya chegaralari. 6: 23. doi:10.3389 / fmicb.2015.00023. PMC 4307647. PMID 25699024.
- ^ Silverman M, Simon M (May 1974). "Flagellar rotation and the mechanism of bacterial motility". Tabiat. 249 (452): 73–4. Bibcode:1974Natur.249...73S. doi:10.1038/249073a0. PMID 4598030.
- ^ Meister GL, Berg HC (1987). "Rapid rotation of flagellar bundles in swimming bacteria". Tabiat. 325 (6105): 637–640. Bibcode:1987Natur.325..637L. doi:10.1038/325637a0.
- ^ Berg HC, Anderson RA (October 1973). "Bacteria swim by rotating their flagellar filaments". Tabiat. 245 (5425): 380–2. Bibcode:1973Natur.245..380B. doi:10.1038/245380a0. PMID 4593496.
- ^ Jahn TL, Bovee EC (1965). "Movement and locomotion of microorganisms". Mikrobiologiyaning yillik sharhi. 19: 21–58. doi:10.1146/annurev.mi.19.100165.000321. PMID 5318439.
- ^ Harshey RM (2003). "Bacterial motility on a surface: many ways to a common goal". Mikrobiologiyaning yillik sharhi. 57: 249–73. doi:10.1146 / annurev.micro.57.030502.091014. PMID 14527279.
- ^ Jarrell K (2009). "Archaeal Flagella and Pili". Pili and Flagella: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-48-6.
- ^ Brady, Richard M. (1993). "Torque and switching in the bacterial flagellar motor. An electrostatic model". Biofizika jurnali. 64 (4): 961–973. Bibcode:1993BpJ....64..961B. doi:10.1016/S0006-3495(93)81462-0. PMC 1262414. PMID 7684268.
- ^ Streif S, Staudinger WF, Marwan W, Oesterhelt D (2008). "Flagellar rotation in the archaeon Halobacterium salinarum depends on ATP". Molekulyar biologiya jurnali. 384 (1): 1–8. doi:10.1016/j.jmb.2008.08.057. PMID 18786541.
- ^ a b Skerker, J. M.; Berg, H. C. (2001-06-05). "Direct observation of extension and retraction of type IV pili". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 98 (12): 6901–6904. Bibcode:2001PNAS...98.6901S. doi:10.1073/pnas.121171698. ISSN 0027-8424. PMC 34450. PMID 11381130.
- ^ Mattick, John S. (2002). "IV turdagi qoziq va tebranish harakati". Mikrobiologiyaning yillik sharhi. 56: 289–314. doi:10.1146 / annurev.micro.56.012302.160938. ISSN 0066-4227. PMID 12142488.
- ^ Merz, A. J.; So, M.; Sheetz, M. P. (2000-09-07). "Pilus retraction powers bacterial twitching motility". Tabiat. 407 (6800): 98–102. Bibcode:2000Natur.407...98M. doi:10.1038/35024105. ISSN 0028-0836. PMID 10993081.
- ^ Henrichsen, J. (December 1972). "Bakterial sirt translokatsiyasi: tadqiqot va tasnif". Bakteriologik sharhlar. 36 (4): 478–503. ISSN 0005-3678. PMC 408329. PMID 4631369.
- ^ Nan, Beyyan (2017 yil fevral). "Bakteriyalarning sirpanish harakati: konsensus modelini chiqarish". Hozirgi biologiya. 27 (4): R154-R156. doi:10.1016 / j.cub.2016.12.035. PMID 28222296.
- ^ Nan, Beyyan; Makbrayd, Mark J.; Chen, Jing; Zusman, Devid R.; Oster, Jorj (fevral 2014). "Vertikal yo'llar bilan sirg'aladigan bakteriyalar". Hozirgi biologiya. 24 (4): 169–174. doi:10.1016 / j.cub.2013.12.034. PMC 3964879. PMID 24556443.
- ^ Sibley, L. Devid; Xaksonson, Sebastyan; Carruthers, Vern B (1998-01-01). "Gliding motility: hujayralarga kirib borishning samarali mexanizmi". Hozirgi biologiya. 8 (1): R12-R14. doi:10.1016/S0960-9822(98)70008-9. PMID 9427622.
- ^ Sibley, LDI (Oct 2010). "Apikomplexan parazitlari hujayralarga qanday va qanday qilib chiqib ketadi". Curr Opin Biotexnol. 21 (5): 592–8. doi:10.1016/j.copbio.2010.05.009. PMC 2947570. PMID 20580218.
- ^ Harshey, Rasika M. (2003-01-01). "Sirtdagi bakteriyalar harakatchanligi: umumiy maqsadga erishish uchun ko'plab usullar". Mikrobiologiyaning yillik sharhi. 57 (1): 249–73. doi:10.1146 / annurev.micro.57.030502.091014. PMID 14527279.
- ^ Henrichsen, J (1972). "Bakterial sirt translokatsiyasi: tadqiqot va tasnif" (PDF). Bakteriologik sharhlar. 36 (4): 478–503. PMC 408329. PMID 4631369.
- ^ "BIOL 230 Lab Manual: Nonmotile Bacteria in Motility Medium". faculty.ccbcmd.edu.
- ^ Walsby AE (1969). "The Permeability of Blue-Green Algal Gas-Vacuole Membranes to Gas". London Qirollik jamiyati materiallari. B seriyasi, Biologiya fanlari. 173 (1031): 235–255. Bibcode:1969RSPSB.173..235W. doi:10.1098/rspb.1969.0049. JSTOR 75817. OCLC 479422015.
- ^ Kalanetra KM, Huston SL, Nelson DC (December 2004). "Novel, attached, sulfur-oxidizing bacteria at shallow hydrothermal vents possess vacuoles not involved in respiratory nitrate accumulation". Amaliy va atrof-muhit mikrobiologiyasi. 70 (12): 7487–96. doi:10.1128/AEM.70.12.7487-7496.2004. PMC 535177. PMID 15574952.
- ^ Schulz-Vogt HN (2006). "Vacuoles". Inclusions in Prokaryotes. Mikrobiologiya monografiyalari. 1. 295-298 betlar. doi:10.1007/3-540-33774-1_10. ISBN 978-3-540-26205-3.
- ^ Montánchez, Itxaso; Ogayar, Elixabet; Plágaro, Ander Hernández; Esteve-Codina, Anna; Gómez-Garrido, Jèssica; Orruño, Maite; Arana, Inés; Kaberdin, Vladimir R. (2019). "Analysis of Vibrio harveyi adaptation in sea water microcosms at elevated temperature provides insights into the putative mechanisms of its persistence and spread in the time of global warming". Ilmiy ma'ruzalar. 9. doi:10.1038/s41598-018-36483-0. PMID 30670759. S2CID 58950215.
- ^ McFall-Ngai, Margaret; Xit-Xekman, Elizabeth A.C.; Gillette, Amani A.; Peyer, Suzanna M.; Harvi, Elizabeth A. (2012). "Hamjihat simbiozlarning maxfiy tillari: Evropimna skolopalaridan tushunchalar - Vibrio fischeri simbiozi". Immunologiya bo'yicha seminarlar. 24 (1): 3–8. doi:10.1016 / j.smim.2011.11.006. PMC 3288948. PMID 22154556.
- ^ Uoterlar, Kristofer M.; Bassler, Bonni L. (2005-10-07). "KUORUM SENSING: Bakteriyalarda hujayradan hujayraga aloqa". Hujayra va rivojlanish biologiyasining yillik sharhi. 21 (1): 319–346. doi:10.1146 / annurev.cellbio.21.012704.131001. PMID 16212498.
- ^ Yosh, R .; Roper, C. (1976). "O'rta suv hayvonlarida biolyuminestsent hisoblagichlar: tirik kalamar dalillari". Ilm-fan. 191 (4231): 1046–1048. Bibcode:1976 yil ... 191.1046Y. doi:10.1126 / science.1251214. PMID 1251214.
- ^ Ouens, Ley; Busiko-Salcedo, Nensi (2006). "Vibrio harveyi: Jannatdagi go'zal muammolar (19-bob)". Tompsonda, Fabiano; Ostin, Brayan; Swings, Jean (tahrir). Vibrios biologiyasi. ASM Press.
- ^ Posfai, M., Lefevre, C., Trubitsyn, D., Bazylinski, D.A. va Frankel, R. (2013) "Magnetosoma minerallarining tarkibi va kristal morfologiyasining filogenetik ahamiyati". Mikrobiologiyada chegara, 4: 344. doi:10.3389 / fmicb.2013.00344. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 3.0 xalqaro litsenziyasi.
- ^ Lin, Vey; Chjan, Vensi; Chjao, Sian; Roberts, Endryu; Paterson, Greig; Bazylinski, Dennis; Pan, Yongxin (2018 yil mart). "Magnetotaktik bakteriyalarning genomik kengayishi naslga xos evolyutsiyasi bilan magnetotaksisning erta umumiy kelib chiqishini ochib beradi". ISME jurnali.
- ^ a b Dyuzenberi, Devid B. (2009). Mikro miqyosda yashash: kichik bo'lishning kutilmagan fizikasi. Kembrij, Mass.: Garvard universiteti matbuoti. 100-101 betlar. ISBN 978-0-674-03116-6.
- ^ a b Chjan, VJ va Vu, LF, 2020. Dengiz magnetotaktik bakteriyalarining flagella va suzish harakati. Biyomolekulalar, 10 (3), s.460. doi:10.3390 / biom10030460. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Yuguruvchi, C .; Vanner, G.; Kolinko, S .; Nibler M.; Amann, R .; Petersen, N .; Kube, M .; Reyxardt, R .; Schuler, D. (2010). "Chuqur tarvaqaylab ketgan Nitrospira phylumining ishlov berilmagan a'zosida proteobakterial magnetosoma genlari va tuzilmalarining saqlanishi". Milliy fanlar akademiyasi materiallari. 108 (3): 1134–1139. Bibcode:2011PNAS..108.1134J. doi:10.1073 / pnas.1012694108. PMC 3024689. PMID 21191098.
- ^ Monteil, Kerolin L.; Vallenet, Devid; Menguy, Nikolas; Benzerara, Karim; Barbe, Valeri; Fouto, Stefani; Kruod, Korin; Floriani, Magali; Viollier, Erik; Adryanczyk, Jeraldin; Leonhardt, Natali (iyul 2019). "Dengiz protistida magnetoreseptsiya kelib chiqadigan ektosimbiotik bakteriyalar". Tabiat mikrobiologiyasi. 4 (7): 1088–1095. doi:10.1038 / s41564-019-0432-7. ISSN 2058-5276. PMC 6697534. PMID 31036911.
- ^ Natan, Eviatar; Fitak, Robert Rojers; Verber, Yuval; Vortman, Yoni (2020-09-28). "Simbiotik magnit zondlash: dalillarni oshirish va boshqalar". Qirollik jamiyatining falsafiy operatsiyalari B: Biologiya fanlari. 375 (1808): 20190595. doi:10.1098 / rstb.2019.0595.
- ^ Kopp, R. E. va Kirschvink, J. L. (2007). "Fotoalbom magnetotaktik bakteriyalarni aniqlash va biogeokimyoviy talqini" (PDF). Earth-Science sharhlari. 86 (1–4): 42–61. Bibcode:2008ESRv ... 86 ... 42K. doi:10.1016 / j.earscirev.2007.08.001.
- ^ Chang, S. R. & J. L. Kirschvink (1989). "Magnetofosillalar, cho'kindilarning magnitlanishi va magnetit biomineralizatsiya evolyutsiyasi". Yer va sayyora fanlari bo'yicha yillik sharh. 17: 169–195. Bibcode:1989AREPS..17..169C. doi:10.1146 / annurev.ea.17.050189.001125.
- ^ DeLong, E.F .; Beja, O. (2010). "Yorug'lik bilan ishlaydigan proton nasosi proteorhodopsin qiyin paytlarda bakteriyalarning yashashini kuchaytiradi". PLOS biologiyasi. 8 (4): e1000359. doi:10.1371 / journal.pbio.1000359. PMC 2860490. PMID 20436957. e1000359.
- ^ a b Gomes-Konsarnau, L .; Raven, J.A .; Levine, N.M .; To'sar, L.S .; Vang, D.; Segerlar, B .; Aristegi, J .; Fuhrman, J.A .; Gasol, JM .; Sañudo-Wilhelmy, SA (2019). "Mikrobial rodopsinlar dengizda qo'lga kiritilgan quyosh energiyasining asosiy hissasi hisoblanadi". Ilmiy yutuqlar. 5 (8): eaaw8855. Bibcode:2019SciA .... 5.8855G. doi:10.1126 / sciadv.aaw8855. PMC 6685716. PMID 31457093.
- ^ Oren, Aaron (2002). "Haddan tashqari halofil arxey va bakteriyalarning molekulyar ekologiyasi". FEMS Mikrobiologiya Ekologiyasi. 39 (1): 1–7. doi:10.1111 / j.1574-6941.2002.tb00900.x. PMID 19709178.
- ^ Beja, O .; Aravind, L .; Koonin, E.V .; Suzuki, M.T .; Xadd, A .; Nguyen, L.P.; Jovanovich, S.B .; Geyts, CM; Feldman, R.A .; Spudich, J.L .; Spudich, E.N. (2000). "Bakterial rodopsin: dengizdagi fototrofiyaning yangi turiga dalil". Ilm-fan. 289 (5486): 1902–1906. Bibcode:2000Sci ... 289.1902B. doi:10.1126 / science.289.5486.1902. PMID 10988064.
- ^ "Fursatdoshlar bilan suhbatlar: Ed Delong". Amerika Mikrobiologiya Akademiyasi. Arxivlandi asl nusxasi 2016 yil 7-avgustda. Olingan 2 iyul 2016.
- ^ Batareyali bakteriyalar, Ommabop ilm, 2001 yil yanvar, 55-bet.
- ^ Beja, O .; Aravind, L .; Koonin, E.V .; Suzuki, M.T .; Xadd, A .; Nguyen, L.P.; Jovanovich, S.B .; Geyts, CM; Feldman, R.A .; Spudich, J.L .; Spudich, E.N. (2000). "Bakterial rodopsin: dengizdagi fototrofiyaning yangi turiga dalil". Ilm-fan. 289 (5486): 1902–1906. Bibcode:2000Sci ... 289.1902B. doi:10.1126 / science.289.5486.1902. PMID 10988064.
- ^ Boeuf, Dominik; Audic, Stefan; Brillet-Gyeguen, Lorayn; Karon, Kristof; Jeanthon, Christian (2015). "MicRhoDE: mikroblarning rodopsin xilma-xilligi va evolyutsiyasini tahlil qilish uchun mo'ljallangan ma'lumotlar bazasi". Ma'lumotlar bazasi. 2015: bav080. doi:10.1093 / ma'lumotlar bazasi / bav080. PMC 4539915. PMID 26286928.
- ^ Yawo, Xiromu; Kandori, Xideki; Koizumi, Amane (2015 yil 5-iyun). Optogenetika: yorug'likni sezuvchi oqsillar va ularning qo'llanilishi. Springer. 3-4 bet. ISBN 978-4-431-55516-2. Olingan 30 sentyabr 2015.
- ^ Kichkina dengiz mikroblari iqlim o'zgarishida katta rol o'ynashi mumkin Janubiy Kaliforniya universiteti, Matbuot xonasi, 2019 yil 8-avgust.
- ^ DasSarma, Shiladitya; Shviterman, Edvard V. (11 oktyabr 2018). "Yerdagi binafsha retinal pigmentlarning dastlabki evolyutsiyasi va ekzoplaneta biosignaturalariga ta'siri". Xalqaro Astrobiologiya jurnali: 1–10. arXiv:1810.05150. doi:10.1017 / S1473550418000423.
- ^ Sparks, Uilyam B.; DasSarma, S .; Reid, I. N. (2006 yil dekabr). "Ibtidoiy fotosintez tizimlari o'rtasidagi evolyutsion raqobat: erta binafsha Yerning mavjudligi?". Amerika Astronomiya Jamiyati Uchrashuvining Referatlari. 38: 901. Bibcode:2006AAS ... 209.0605S.
- ^ Dane Konop (1997 yil 29-iyul). "Olimlar Meksika ko'rfazidagi dengiz tubida metan muzli qurtlarni topdilar". Milliy okean va atmosfera boshqarmasi. Arxivlandi asl nusxasi 2010 yil 9 iyunda. Olingan 22 yanvar, 2010.
- ^ Lema, KA, Uillis, B.L. va Bourne, D.G. (2012) "Marjonlar simbiyotik azotni biriktiruvchi bakteriyalar bilan xarakterli birlashmalar hosil qiladi". Amaliy va atrof-muhit mikrobiologiyasi, 78(9): 3136-3144. doi:10.1128 / AEM.07800-11.
- ^ Petersen, Jillian M.; Frank U. Zielinski; Tomas Peyp; Richard Zayfert; Kristina Moraru; va boshq. (2011-08-11). "Vodorod gidrotermal ventilyatsiya simbiozlari uchun energiya manbai". Tabiat. 476 (7359): 176–180. Bibcode:2011 yil natur.476..176P. doi:10.1038 / tabiat10325. PMID 21833083.

- ^ Klayner, Manuel; Wentrup, Cecilia; Lott, xristian; Tishlash, Hanno; Vetsel, Silke; Yosh, Jak; Chang, Yun-Xuan; Shoh, Manesh; VerBerkmoes, Natan C. (2012-05-08). "Ichaksiz dengiz qurtining metaproteomikasi va uning simbiotik mikroblar jamiyati uglerod va energiyadan foydalanish uchun odatiy bo'lmagan yo'llarni ochib beradi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 109 (19): E1173-E1182. doi:10.1073 / pnas.1121198109. PMC 3358896. PMID 22517752.
- ^ Voyk, Tanja; Tishlash, Hanno; Ivanova, Natalya N.; Xuntemann, Marsel; Rixter, Maykl; Gloekner, Frank Oliver; Boffelli, Dario; Anderson, Ieyn J.; Barri, Kerri V. (2006-10-26). "Mikrobial konsortsiumni metagenomik tahlil qilish orqali simbioz haqidagi tushunchalar" (PDF). Tabiat. 443 (7114): 950–955. Bibcode:2006 yil natur.443..950W. doi:10.1038 / nature05192. PMID 16980956.
- ^ a b Ruiz-Gonsales, S, Simo, R., Sommaruga, R. va Gasol, JM (2013) "Zulmatdan uzoqroq: quyosh nurlanishining heterotrofik bakterioplankton faolligiga ta'siri". Mikrobiologiyada chegara, 4: 131. doi:10.3389 / fmicb.2013.00131. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 3.0 xalqaro litsenziyasi.
- ^ Walker, J. C. G. (1980). Tabiiy muhitdagi kislorod tsikli va biogeokimyoviy tsikllar. Berlin: Springer-Verlag.
- ^ Longxurst, A., Sathyendranath, S., Platt, T. va Caverhill, C. (1995). Sun'iy yo'ldosh radiometr ma'lumotlaridan ummonda global birlamchi ishlab chiqarishni taxmin qilish. J. Plankton Res. 17, 1245–1271.
- ^ a b v d e Kurata, N., Vella, K., Xemilton, B., Shivji, M., Soloviev, A., Mett, S., Tartar, A. va Perri, W. (2016) "Yaqin atrofdagi sirt faol moddalar bilan bog'liq bakteriyalar. - okeanning sirt qatlami ". Tabiat: Ilmiy ma'ruzalar, 6(1): 1–8. doi:10.1038 / srep19123. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Chutić, V., Josovich, B., Marčenko, E., Bihari, N. va Krshinich, F. (1981) "Dengiz fitoplanktonlari tomonidan sirt faol moddalarini ishlab chiqarish". Dengiz kimyosi, 10(6): 505–520. doi:10.1016/0304-4203(81)90004-9.
- ^ a b Zigel, Devid A.; Buesseler, Ken O.; Behrenfeld, Maykl J.; Benites-Nelson, Klaudiya R.; Boss, Emmanuel; Bjezinski, Mark A.; Burd, Adrian; Karlson, Kreyg A.; d'Asaro, Erik A.; Doney, Skott S.; Perri, Meri J.; Stenli, Reychel H. R.; Steinberg, Deborah K. (2016). "Global Ocean Net boshlang'ich ishlab chiqarish eksporti va taqdiri bashorati: EXPORTS ilmiy rejasi". Dengiz fanidagi chegara. 3. doi:10.3389 / fmars.2016.00022. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Liu X, Pan J, Liu Y, Li M, Gu JD (oktyabr 2018). "Arxeylarning global estuariy ekotizimlarida xilma-xilligi va tarqalishi". Umumiy muhit haqida fan. 637–638: 349–358. Bibcode:2018ScTEn.637..349L. doi:10.1016 / j.scitotenv.2018.05.016. PMID 29753224.
- ^ Cabello P, Roldan MD, Moreno-Vivian C (2004 yil noyabr). "Arxeyadagi nitratlarning kamayishi va azot aylanishi". Mikrobiologiya. 150 (Pt 11): 3527-46. doi:10.1099 / mic.0.27303-0. PMID 15528644.
- ^ Mehta MP, Baross JA (2006 yil dekabr). "Gidrotermal vent arxeon tomonidan azotni 92 daraja S darajasida fiksatsiya qilish". Ilm-fan. 314 (5806): 1783–86. Bibcode:2006 yil ... 314.1783M. doi:10.1126 / science.1134772. PMID 17170307.
- ^ Frensis CA, Beman JM, Kuypers MM (may 2007). "Azot tsiklining yangi jarayonlari va ishtirokchilari: anaerob va arxaeal ammiak oksidlanishining mikrob ekologiyasi". ISME jurnali. 1 (1): 19–27. doi:10.1038 / ismej.2007.8. PMID 18043610.
- ^ Coolen MJ, Abbas B, van Bleijswijk J, Hopmans EC, Kuypers MM, Wakeham SG, Sinninghe Damsté JS va boshq. (2007 yil aprel). "Qora dengizning suboksik suvlaridagi ammiakni oksidlovchi Crenarchaeota: 16S ribosomal va funktsional genlar va membrana lipidlari yordamida havza bo'ylab ekologik tadqiqotlar". Atrof-muhit mikrobiologiyasi. 9 (4): 1001–16. doi:10.1111 / j.1462-2920.2006.01227.x. hdl:1912/2034. PMID 17359272.
- ^ Beyker BJ, Banfild JF (2003 yil may). "Kislota konlarini drenajlashda mikroblar jamoalari". FEMS Mikrobiologiya Ekologiyasi. 44 (2): 139–52. doi:10.1016 / S0168-6496 (03) 00028-X. PMID 19719632.
- ^ Schimel J (2004 yil avgust). "Metan tsiklida tarozi o'ynash: mikrobial ekologiyadan dunyoga". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 101 (34): 12400–01. Bibcode:2004 yil PNAS..10112400S. doi:10.1073 / pnas.0405075101. PMC 515073. PMID 15314221.
Bibliografiya
- Dalrimple, G. Brent (2001). "Yigirmanchi asrdagi Yerning yoshi: muammo (asosan) hal qilindi". Maxsus nashrlar, London Geologiya Jamiyati. 190 (1): 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144 / GSL.SP.2001.190.01.14.
- Raven, Piter Xemilton; Jonson, Jorj Bruks (2002). Biologiya. McGraw-Hill Education. ISBN 978-0-07-112261-0.
- Shors, Teri (2017). Viruslarni tushunish. Jons va Bartlett nashriyotlari. ISBN 978-1284025927.
