Dengiz oziq-ovqat tarmog'i - Marine food web - Wikipedia

Quruq muhit bilan taqqoslaganda dengiz muhiti mavjud biomassa piramidalari bazasida teskari bo'lgan. Xususan, iste'molchilar biomassasi (kopepodlar, krill, qisqichbaqalar, em-xashak baliqlari) biomassasidan kattaroqdir. asosiy ishlab chiqaruvchilar. Bu okeanning asosiy ishlab chiqaruvchilari kichik bo'lganligi sababli sodir bo'ladi fitoplankton tez o'sadigan va ko'payadigan, shuning uchun kichik massa tez tezlikka ega bo'lishi mumkin birlamchi ishlab chiqarish. Aksincha, ko'plab muhim quruqlikdagi asosiy ishlab chiqaruvchilar, masalan etuk o'rmonlar, sekin o'sib boradi va ko'payadi, shuning uchun birlamchi ishlab chiqarishning bir xil darajasiga erishish uchun juda katta massa kerak bo'ladi.
Ushbu inversiya tufayli, bu zooplankton dengiz hayvonlarining ko'p qismini tashkil etadi biomassa. Sifatida asosiy iste'molchilar, zooplankton birlamchi ishlab chiqaruvchilar (asosan fitoplankton) va qolgan dengiz oziq-ovqat tarmog'i (ikkilamchi iste'molchilar ).[1]
Agar fitoplankton eyishdan oldin o'lsa, u orqali tushadi eyfotik zona qismi sifatida dengiz qorlari va dengiz tubiga joylashadi. Shunday qilib, fitoplankton har yili okeanga qariyb 2 milliard tonna karbonat angidridni ajratadi, natijada okean barcha sekvestrlangan uglerodning taxminan 90 foizini ushlab turgan karbonat angidrid cho'kmasiga aylanadi.[2] Okean dunyodagi kislorodning taxminan yarmini ishlab chiqaradi va atmosferaga qaraganda 50 barobar ko'proq karbonat angidridni to'playdi.[3]
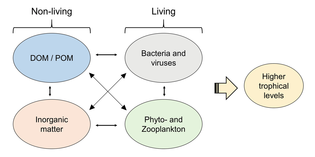
Ekotizimni, uning oziq-ovqat tarmog'i materiallar va energiya oqimini qanday belgilashini bilmasdan tushunib bo'lmaydi. Fitoplankton konvertatsiya qilish yo'li bilan biomassani avtotrofik tarzda hosil qiladi noorganik birikmalar ichiga organik bittasi. Shu tarzda fitoplankton okeandagi barcha boshqa hayotni qo'llab-quvvatlash orqali dengiz oziq-ovqat tarmog'ining asosi sifatida ishlaydi. Dengiz oziq-ovqat tarmog'idagi ikkinchi markaziy jarayon bu mikrobial tsikl. Ushbu pastadir yomonlashadi dengiz bakteriyalari va arxey, remineralises organik va noorganik moddalarni o'z ichiga oladi, so'ngra mahsulotni pelagik oziq-ovqat tarmog'ida yoki cho'kindi jinsi sifatida dengiz tubiga yotqizish orqali qayta ishlaydi.[4]
Oziq-ovqat zanjirlari va trofik darajalar
| dengiz oziq-ovqat zanjiri (odatiy) |
|---|
| ↓ |
| fitoplankton |
| ↓ |
| o'txo'r zooplankton |
| ↓ |
| yirtqich zooplankton |
| ↓ |
| ↓ |
Oziq-ovqat tarmoqlari dan qurilgan oziq-ovqat zanjirlari. Dengizdagi barcha hayot shakllari boshqa hayot shakli uchun oziq-ovqatga aylanish imkoniyatiga ega. Okeanda oziq-ovqat zanjiri odatda quyosh energiyasi bilan boshlanadi fitoplankton va quyidagi kursga amal qiladi:
fitoplankton → o‘txo‘r zooplankton → yirtqich zooplankton → filtri oziqlantiruvchi → yirtqich umurtqali hayvon
Fitoplankton oziq-ovqat uchun boshqa organizmlarning hojati yo'q, chunki ular o'zlarining oziq-ovqatlarini to'g'ridan-to'g'ri noorganik ugleroddan ishlab chiqarish qobiliyatiga ega va quyosh nurlaridan energiya manbai sifatida foydalanishlari mumkin. Ushbu jarayon deyiladi fotosintez va natijada fitoplankton tabiiy ravishda hosil bo'lgan uglerodni konversiyalashiga olib keladi protoplazma. Shu sababli fitoplankton deb aytilgan asosiy ishlab chiqaruvchilar dengiz oziq-ovqat zanjirining pastki qismida yoki birinchi darajasida. Ular birinchi darajadagi bo'lgani uchun ular a trofik daraja 1 dan (dan Yunoncha kubok ovqat degani). Keyin fitoplanktonni zooplankton deb nomlangan mikroskopik hayvonlar oziq-ovqat zanjiridagi keyingi trofik darajada iste'mol qiladilar.
Zooplankton oziq-ovqat zanjiridagi ikkinchi trofik darajani tashkil etadi va mikroskopik bir hujayrali organizmlarni o'z ichiga oladi protozoa shuningdek kichik qisqichbaqasimonlar, kabi kopepodlar va krill, va lichinka baliq, kalamar, lobster va qisqichbaqalar. Ushbu darajadagi organizmlarni quyidagicha tasavvur qilish mumkin asosiy iste'molchilar.
O'z navbatida, kichikroq o'txo'r zoplanktonni yirik yirtqich protozoa kabi yirik yirtqich hayvonot hayvonlari iste'mol qiladilar. krill va tomonidan em-xashak baliqlari kichik, maktabda o'qish, filtr bilan oziqlantirish baliq. Bu oziq-ovqat zanjiridagi uchinchi trofik darajani tashkil qiladi.
To'rtinchi trofik darajadan iborat yirtqich baliqlar, dengiz sutemizuvchilar va dengiz qushlari em-xashak baliqlarini iste'mol qiladiganlar. Misollar qilich-baliq, muhrlar va gannets.
Apex yirtqichlari, masalan orcas, muhrlarni iste'mol qilishi mumkin va shorto mako akulalari, qilich baliqlarini iste'mol qilishi mumkin, beshinchi trofik darajani tashkil qiladi. Balin kitlari to'g'ridan-to'g'ri zooplankton va krillni iste'mol qilishi mumkin, bu faqat uchta yoki to'rtta trofik darajadagi oziq-ovqat zanjiriga olib keladi.
Amalda trofik darajalar odatda oddiy tamsayılar emas chunki bir xil iste'mol turlari ko'pincha bir nechta trofik darajalarda oziqlanadi.[8][9] Masalan katta dengiz umurtqali hayvonlar kichikroq yirtqich baliqlarni iste'mol qilishi mumkin, lekin filtrli oziqlantiruvchilarni ham iste'mol qilishi mumkin; The nayza yeydi qisqichbaqasimonlar, lekin bolg'a ikkala qisqichbaqasimon va qaqshatqich hayvonlarni ham iste'mol qiladi. Hayvonlar ham bir-birlarini eyishi mumkin; The cod kichikroq baliqlarni ham iste'mol qiladi Qisqichbaqa, va kerevitlar treska lichinkalarini eyishadi. Voyaga etmagan hayvonni oziqlantirish odatlari va natijada uning trofik darajasi o'sishi bilan o'zgarishi mumkin.
Baliqchilik sohasidagi olim Daniel Pauly trofik darajalarning qiymatlarini birlamchi ishlab chiqaruvchilardan biriga o'rnatadi va detrit, ikkisi o'txo'r va detritivorlarda (birlamchi iste'molchilar), uchtasi ikkilamchi iste'molchilarda va boshqalar. Trofik darajaning ta'rifi, har qanday iste'mol turi uchun TL:[10]

qayerda yirtqichning fraksiyonel trofik darajasi jva ning qismini ifodalaydi j ning dietasida men. Dengiz ekotizimlarida ko'plab baliqlar va boshqa dengiz iste'molchilarining trofik darajasi 2,0 dan 5,0 gacha qiymatga ega. Yuqori qiymati 5.0, hatto katta baliqlar uchun ham g'ayrioddiy,[11] garchi bu dengiz sutemizuvchilarining cho'qqi ayiqlari va qotil kitlari kabi apeks yirtqichlarida uchraydi.[12] Qarama-qarshilik nuqtai nazaridan odamlarning o'rtacha trofik darajasi taxminan 2,21 ga teng, bu cho'chqa yoki hamsi bilan bir xil.[13][14]


Takson tomonidan
Birlamchi ishlab chiqaruvchilar

Okeanning oziqlanish tarmog'i asosida bir hujayrali suv o'tlari va boshqa o'simlikka o'xshash organizmlar ma'lum fitoplankton. Quruqlikdagi o'simliklar singari fitoplanktondan foydalanish xlorofill va boshqalar engil yig'adigan pigmentlar amalga oshirish fotosintez, yoqilg'i uchun shakar ishlab chiqarish uchun atmosferadagi karbonat angidridni yutadi. Suvdagi xlorofil suvning aks etishi va quyosh nurlarini yutish usulini o'zgartiradi, olimlarga fitoplankton miqdori va joylashishini xaritada ko'rsatishga imkon beradi. Ushbu o'lchovlar olimlarga okean atrof-muhitining salomatligi to'g'risida qimmatli tushunchalar beradi va olimlarga bularni o'rganishga yordam beradi okean uglerod tsikli.[15]
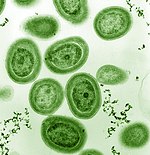
Fitoplankton orasida bakteriyalar filumidan a'zolar bor siyanobakteriyalar. Dengiz siyanobakteriyalariga ma'lum bo'lgan eng kichik fotosintetik organizmlar kiradi. Eng kichigi, Proxlorokokk, atigi 0,5 dan 0,8 mikrometrgacha.[16] Shaxsiy raqamlar bo'yicha, Proxlorokokk ehtimol Yerdagi eng mo'l tur: bir millilitr dengiz suvi 100000 hujayradan yoki undan ko'proq hujayradan iborat bo'lishi mumkin. Dunyo bo'ylab bir nechta bo'lishi taxmin qilinmoqda oktillion (1027) jismoniy shaxslar.[17] Proxlorokokk hamma joyda 40 ° N dan 40 ° S gacha bo'lgan va oligotrofik (ozuqaviy jihatdan kambag'al) okeanlarning mintaqalari.[18] Bakteriya Yer atmosferasidagi kislorodning taxminan 20% ni tashkil qiladi.[19]
- Fitoplankton okeandagi oziq-ovqat zanjirining asosini tashkil qiladi

Fitoplankton

Dinoflagellat

Diatomlar



Okeanlarda, ko'pchilik birlamchi ishlab chiqarish tomonidan amalga oshiriladi suv o'tlari. Bu asosiy ishlab chiqarishning aksariyati tomonidan amalga oshiriladigan quruqlikdan farqli o'laroq qon tomir o'simliklar. Yosunlar bitta suzuvchi hujayralardan biriktirilgangacha dengiz o'tlari, tomir o'simliklari esa okeanda kabi guruhlar bilan ifodalanadi dengiz o'tlari va mangrovlar. Kattaroq ishlab chiqaruvchilar, masalan dengiz o'tlari va dengiz o'tlari, asosan bilan cheklangan qirg'oq zonasi va sayoz suvlari, ular qaerda biriktirish asosiy substratga va hali ham fonik zona. Ammo suv o'tlari tomonidan ishlab chiqariladigan asosiy ishlab chiqarishning ko'p qismi fitoplankton tomonidan amalga oshiriladi.
Shunday qilib, okean muhitida birinchi pastki trofik sath asosan egallaydi fitoplankton, asosan mikroskopik driftli organizmlar bir hujayrali suv o'tlari, dengizda suzadi. Ko'pgina fitoplanktonlar juda kichik bo'lib, ularni alohida-alohida ko'rish mumkin emas yordamsiz ko'z. Ular etarlicha yuqori miqdordagi suv mavjud bo'lganda (ko'pincha yashil) rang o'zgarishi kabi ko'rinishi mumkin. Ular biomassasini asosan fotosintez orqali ko'paytirgani uchun ular quyosh nurli sirt qatlamida yashaydilar (eyfotik zona ) dengiz.
Fitoplanktonning eng muhim guruhlariga quyidagilar kiradi diatomlar va dinoflagellatlar. Diyatomlar okeanlarda juda muhimdir, chunki ba'zi taxminlarga ko'ra ular okeanning asosiy ishlab chiqarish hajmining 45 foizini tashkil etadi.[20] Diatomlar odatda mikroskopik, garchi ba'zi turlarning uzunligi 2 millimetrga etishi mumkin.
Birlamchi iste'molchilar
Ikkinchi trofik daraja (asosiy iste'molchilar ) egallaydi zooplankton qaysi oziqlantiradi fitoplankton. Ular fitoplankton bilan birgalikda dunyodagi eng katta baliq ovlanadigan joylarni qo'llab-quvvatlaydigan oziq-ovqat piramidasining asosini tashkil qiladi. Zooplankton - bu fitoplankton bilan topilgan mayda hayvonlar okeanik er usti suvlari va kichkinagina kiradi qisqichbaqasimonlar va baliq lichinkalari va qovurmoq (yaqinda chiqqan baliq). Zooplanktonlarning aksariyati filtrli oziqlantiruvchi vositalar va ular fitoplanktonni suvdagi filtrlash uchun qo'shimchalardan foydalanadilar. Ba'zi yirik zooplanktonlar kichikroq zooplanktonlar bilan ham oziqlanadi. Ba'zi zooplanktonlar yirtqichlardan saqlanish uchun biroz sakrashi mumkin, ammo ular suzishga qodir emaslar. Fitoplankton singari, ular o'rniga oqimlar, to'lqinlar va shamollar bilan suzishadi. Zooplanktonlar tezda ko'payishi mumkin, qulay sharoitda ularning populyatsiyasi kuniga o'ttiz foizgacha ko'payishi mumkin. Ko'pchilik qisqa va samarali hayot kechiradi va tezda etuklikka erishadi.
- Zooplankton okean oziq-ovqat zanjirida ikkinchi darajani hosil qiladi

Segmentlangan qurt

Qisqichbaqasimon mayda qisqichbaqasimonlar

Voyaga etmagan planktonik kalmar

Zooplanktonning muhim guruhlari quyidagilardir kopepodlar va krill. Kopepodlar kichik guruhdir qisqichbaqasimonlar okean va chuchuk suv yashash joylari. Ular dengizdagi eng katta oqsil manbai,[21] va em-xashak baliqlari uchun muhim o'lja hisoblanadi. Krill oqsilning keyingi eng katta manbasini tashkil qiladi. Krill kichikroq zooplankton bilan oziqlanadigan yirik yirtqich zooplanktondir. Bu shuni anglatadiki, ular chindan ham em-xashak baliqlari bilan birga uchinchi trofik darajaga, ikkilamchi iste'molchilarga tegishli.

Birgalikda, fitoplankton va zooplankton ko'pini tashkil qiladi plankton dengizda. Plankton - bu har qanday kichik driftga nisbatan qo'llaniladigan atama organizmlar dengizda suzuvchi (Yunoncha planktos = sarson yoki drifter). Ta'rifga ko'ra, plankton deb tasniflangan organizmlar okean oqimlariga qarshi suzishga qodir emaslar; ular atrof-muhit oqimiga qarshi tura olmaydi va o'z pozitsiyalarini nazorat qila olmaydi. Okean muhitida dastlabki ikkita trofik sath asosan egallaydi plankton. Plankton ishlab chiqaruvchilar va iste'molchilarga bo'linadi. Ishlab chiqaruvchilar fitoplankton (yunoncha) fiton = o'simlik) va fitoplanktonni iste'mol qiladigan iste'molchilar zooplankton (yunoncha) zoon = hayvon).
Meduza sekin suzuvchilar va ko'pchilik turlari planktonning bir qismini tashkil qiladi. An'anaviy ravishda meduzalar trofik o'lik, dengiz oziq-ovqat tarmog'idagi kichik o'yinchilar, jelatinli organizmlar tana rejasi asosan suv kabi ozgina ozuqaviy qiymat yoki boshqa organizmlar uchun qiziqishni taklif qiladi, masalan okean quyosh baliqlari va teri toshbaqasi.[23][22] Yaqinda bu fikrga qarshi chiqdi. Meduza va umuman olganda jelatinli zooplankton o'z ichiga oladi qalampir va ktenoforlar, juda xilma-xil, qattiq qismlari bo'lmagan mo'rt, ko'rish va kuzatish qiyin, aholining tez o'zgarib turishi va ko'pincha qirg'oqdan uzoq yoki okean tubida noqulay yashashadi. Olimlar uchun meduzalarni yirtqichlarning ichaklaridan aniqlash va tahlil qilish qiyin, chunki ular eyish paytida mushga aylanib, tez hazm qilinadi.[23] Ammo meduza juda ko'p miqdorda gullaydi va ularning dietasida asosiy tarkibiy qismlarni tashkil etishi ko'rsatilgan orkinos, nayza baliq va qilich-baliq kabi turli xil qushlar va umurtqasizlar sakkizoyoq, dengiz bodringlari, Qisqichbaqa va amfipodlar.[24][22] "Energiya zichligi past bo'lishiga qaramay, meduzalarning yirtqichlarning energiya byudjetiga qo'shadigan hissasi tez hazm bo'lishi, qo'lga olish xarajatlari pastligi, mavjudligi va energiyaga boy tarkibiy qismlar bilan tanlab ovqatlanish sababli taxmin qilinganidan ancha katta bo'lishi mumkin. Meduzalar bilan oziqlanish plastmassalarni yutib yuboradigan dengiz yirtqichlari. "[22]
Yuqori darajadagi iste'molchilar

- Dengiz umurtqasizlar
- Baliq
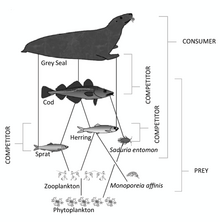
- Oziq-ovqat baliqlari: Yem-xashak baliqlari okeandagi markaziy pozitsiyalarni egallaydi oziq-ovqat tarmoqlari. U iste'mol qiladigan organizmlar pastroqda joylashgan trofik daraja va uni iste'mol qiladigan organizmlar yuqori trofik darajada. Oziq-ovqat baliqlari oziq-ovqat tarmog'ida o'rta darajalarni egallab, yuqori darajadagi baliqlar, dengiz qushlari va sutemizuvchilarga ustunlik qiladi.[25]
- Yirtqich baliq
- Tuproqdagi baliqlar
- Boshqa dengiz umurtqali hayvonlari
2010 yilda tadqiqotchilar kitlar ozuqa moddalarini okean tubidan suv yuzasiga yana o'zlari deb atagan jarayon yordamida olib borishini aniqladilar kit nasosi.[26] Kitlar okeandagi chuqurroq darajalarda oziqlanadi krill topildi, lekin nafas olish uchun muntazam ravishda yuzaga qaytib boring. U erda kitlar suyuqlikni defekatsiya qilish azot va temirga boy. Cho'kish o'rniga suyuqlik sirtda qoladi fitoplankton uni iste'mol qiling. Meyn ko'rfazida kit nasosi daryolarga qaraganda ko'proq azot beradi.[27]

Humpback kitlar pastdan o'pka em-xashak baliqlari bilan boqish
 Kit nasosi ozuqa aylanishi
Kit nasosi ozuqa aylanishi

Mikroorganizmlar

So'nggi yillarda tobora ko'proq tan olinmoqda dengiz mikroorganizmlari dengiz ekotizimlarida ilgari o'ylanganidan ancha katta rol o'ynaydi. Rivojlanishlar metagenomika tadqiqotchilarga mikroskopik hayotning ilgari yashirib kelingan xilma-xilligini ochib berish qobiliyatini beradi, mikroblar dunyosini ko'rish uchun jonli ob'ektiv va tirik dunyoni anglash uchun inqilob qilish imkoniyatini beradi.[29] Parhez tahlilini metabarkodlash taksonomik rezolyutsiyaning yuqori darajalarida oziq-ovqat tarmoqlarini rekonstruksiya qilish uchun texnikadan foydalanilmoqda va o'zaro ta'sirlar tarmog'idagi chuqurroq murakkabliklar aniqlanmoqda.[30]
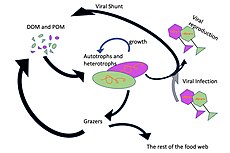
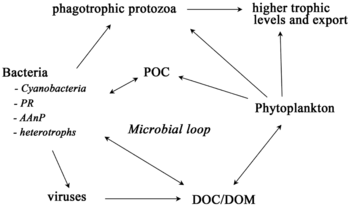
Mikroorganizmlar dengiz oziq-ovqat tarmoqlarida asosiy rollarni o'ynaydi. The virusli shunt yo'l dengiz mikroblarini oldini oluvchi mexanizmdir zarracha bo'lgan organik moddalar (POM) yuqoriga ko'chishdan trofik sathlar ularni qayta ishlash orqali erigan organik moddalar (DOM), bu mikroorganizmlar tomonidan osonlikcha olinishi mumkin.[31] Virusli manevr, mikroblar ekotizimida xilma-xillikni saqlashga yordam beradi, dengiz mikroblarining bir turini mikro muhitda hukmron bo'lishiga yo'l qo'ymaslik.[32] Virusli shunt yo'li bilan qayta ishlangan DOM dengiz DOM ning boshqa asosiy manbalari tomonidan ishlab chiqarilgan miqdor bilan taqqoslanadi.[33]
Umuman, erigan organik uglerod (DOC) okean muhitiga bakterial lizisdan, fitoplanktondan qattiq uglerodning chiqishi yoki ekssudatsiyasidan (masalan, shilimshiq ekzopolimerdan) kiradi. diatomlar ), hujayralarning to'satdan qarishi, zooplankton bilan beparvo oziqlanishi, suv hayvonlari tomonidan chiqindi mahsulotlarning chiqarilishi yoki er usti o'simliklari va tuproqlaridan organik zarralarning parchalanishi yoki erishi.[34] Bakteriyalar mikrobial tsikl o'sish uchun ushbu energiyaga boy moddadan foydalanish uchun bu zarracha detritini parchalash. Organik moddalarning 95% dan ortig'i dengiz ekotizimlarida polimer, yuqori molekulyar og'irlik (HMW) birikmalari (masalan, oqsil, polisaxaridlar, lipidlar), ularning umumiy miqdorining atigi oz qismi erigan organik moddalar (DOM) yuqori trofik darajadagi dengiz organizmlarining ko'pchiligida osonlikcha ishlatilishi mumkin. Demak, erigan organik uglerod to'g'ridan-to'g'ri dengiz organizmlarining ko'pchiligida mavjud emas; dengiz bakteriyalari ushbu organik uglerodni oziq-ovqat tarmog'iga kiritadi, natijada qo'shimcha energiya yuqori trofik darajalarga ega bo'ladi.
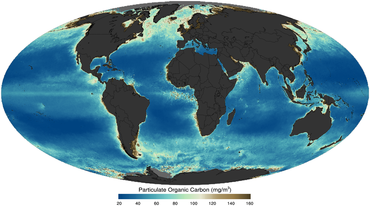
kabi sun'iy yo'ldosh orqali tasvirlangan 2011 yilda




Gigant dengiz virusi CroV yuqtiradi va o'limga olib keladi lizis dengiz zooflagellatining Roenbergensis kafeteryasi.[38] Bu qirg'oq ekologiyasiga ta'sir qiladi, chunki Roenbergensis kafeteryasi suvda uchraydigan bakteriyalar bilan oziqlanadi. Qachon kam sonlar Roenbergensis kafeteryasi keng CroV infektsiyalari tufayli bakterial populyatsiyalar tez o'sib boradi.[39] Ta'siri CroV ning tabiiy populyatsiyalari bo'yicha C. roenbergensis noma'lum bo'lib qolmoqda; ammo, virus juda mezbonga xos ekanligi aniqlandi va boshqa yaqin organizmlarni yuqtirmaydi.[40] Cafeteria roenbergensis, shuningdek, ikkinchi virus bilan yuqtiriladi Mavirus virusi, bu a sun'iy yo'ldosh virusi, demak u faqat boshqa o'ziga xos virus mavjud bo'lganda, bu holda CroV ishtirokida takrorlanishi mumkin.[41] Ushbu virus CroV replikatsiyasiga xalaqit beradi, bu esa omon qolishiga olib keladi C. roenbergensis hujayralar. Mavirus hujayralarining genomiga qo'shilishga qodir C. roenbergensisva shu bilan aholiga immunitet beradi.[42]
Qo'ziqorinlar


Yashash joyi bo'yicha
Pelagik to'rlar
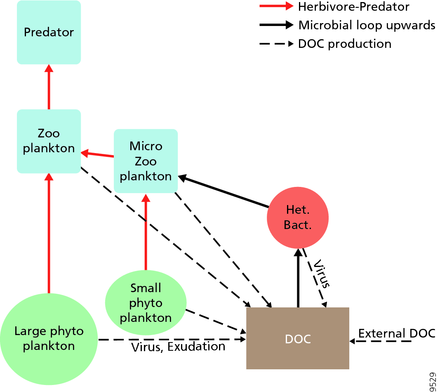
Legendre va Rassoulzadagan pelagik ekotizimlari uchun 1995 yilda o'txo'r oziq-ovqat zanjiri bilan trofik yo'llarning davomiyligini va mikrobial tsikl oziq-ovqat veb-a'zolari sifatida.[47] Klassik chiziqli oziq-ovqat zanjiri so'nggi a'zosi zooplankton tomonidan katta fitoplanktonda o'tlashni va undan keyin zooplanktonda yoki undan kattaroq zooplanktonda yoki boshqa yirtqich hayvonlarda o'tlashni o'z ichiga oladi. Bunday chiziqli oziq-ovqat zanjirida yirtqich yuqori fitoplanktonga olib kelishi mumkin biomassa (fitoplankton, o'txo'r va yirtqich hayvon bo'lgan tizimda) yoki kamaytirilgan fitoplankton biomassasi (to'rt darajali tizimda). Yirtqichlarning ko'payishidagi o'zgarishlar, shunga olib kelishi mumkin trofik kaskadlar.[48] Mikrobial tsiklning so'nggi a'zosi bazal resurs sifatida nafaqat fitoplanktonni, balki uni ham o'z ichiga oladi erigan organik uglerod.[49] Eritilgan organik uglerodni o'sishi uchun geterotrof bakteriyalar foydalanadilar, undan kattaroq zooplankton. Natijada, erigan organik uglerod bakterial-mikrozooplankton tsikli orqali zooplanktonga aylanadi. Ushbu ikkita uglerodni qayta ishlash yo'llari bir necha darajalarda bog'langan. Kichik fitoplanktonni to'g'ridan-to'g'ri mikrozooplankton iste'mol qilishi mumkin.[46]
O'ngdagi diagrammada ko'rsatilgandek, erigan organik uglerod turli xil va turli xil organizmlar tomonidan ishlab chiqariladi, ham organik uglerodning asosiy ishlab chiqaruvchilari, ham iste'molchilari tomonidan. Birlamchi ishlab chiqaruvchilar tomonidan DOC chiqishi passiv ravishda oqish bilan va ozuqaviy moddalarni cheklash paytida muvozanatsiz o'sish paytida sodir bo'ladi.[50][51] Fitoplanktondan erigan organik hovuzga yana bir to'g'ridan-to'g'ri yo'l kiradi virusli lizis.[52] Dengiz viruslari okeandagi fitoplankton o'limining asosiy sababidir, ayniqsa iliq va past kenglikdagi suvlarda. Yalang'och ovqatlanish o'txo'rlar tomonidan va iste'molchilar tomonidan o'ljani to'liq hazm bo'lmasligi erigan organik uglerodning boshqa manbalari hisoblanadi. Geterotrof mikroblar eritish uchun hujayradan tashqari fermentlardan foydalanadilar zarracha bo'lgan organik uglerod va shu va boshqa erigan organik uglerod resurslaridan o'sishi va saqlanishi uchun foydalaning. Mikrobial geterotrofik ishlab chiqarishning bir qismi mikrozooplankton tomonidan qo'llaniladi; heterotrofik hamjamiyatning yana bir qismi kuchli virusli lizisga uchraydi va bu yana erigan organik uglerodning chiqishini keltirib chiqaradi. Mikrobial tsiklning samaradorligi bir qancha omillarga bog'liq, xususan, geterotrof mikroblarning o'limiga yirtqichlik va virusli lizisning nisbiy ahamiyati.[46]
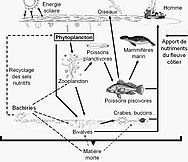
- Pelagik oziq-ovqat tarmog'i

Pelagik oziq-ovqat tarmog'i va biologik nasos. Okeanning biologik nasosi va pelagik oziq-ovqat tarmog'i va ushbu komponentlarni kemalardan, sun'iy yo'ldoshlardan va avtonom transport vositalaridan uzoqdan namuna olish qobiliyati o'rtasidagi aloqalar. Ochiq ko'k suvlar eyfotik zona, quyuqroq ko'k suvlar esa alacakaranlık zonasi.[53]

- Mesopelagik oziq-ovqat tarmog'i

Mesopelagik turlarning global uglerod byudjetiga ta'siri[54]
DVM = diel vertikal migratsiya NM = migratsiya emas


Mesopelagik bristlemutlar sayyoradagi eng ko'p tarqalgan umurtqali hayvonlar bo'lishi mumkin, ammo ular haqida kam ma'lumot mavjud.[55]

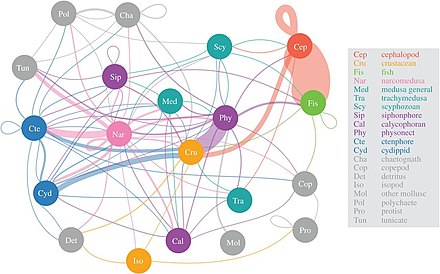
Jelatinli yirtqichlar shunga o'xshash narcomedusan mezopelagik o'ljaning eng xilma-xilligini iste'mol qilish


Olimlar asosan noma'lum alacakaranlık zonasini batafsil o'rganishga kirishmoqdalar mezopelagik, 200 dan 1000 metrgacha chuqurlikda. Ushbu qatlam har yili atmosferadan qariyb 4 milliard tonna karbonat angidridni olib tashlash uchun javobgardir. Mezopelagik qatlamda dengiz baliqlarining aksariyati yashaydi biomassa.[55]
2017 yilgi tadqiqotlarga ko'ra, narcomedusae mezopelagik o'ljaning eng xilma-xilligini iste'mol qiling, keyin fitonekt sifonoforlar, ktenoforlar va sefalopodlar. "Jelly web" ning ahamiyati endi tushunila boshlandi, ammo medusa, ktenofor va sifonoforlar yirtqich baliq va kalamarga o'xshash ekologik ta'sirga ega bo'lgan chuqur pelagik oziq-ovqat tarmoqlarida asosiy yirtqichlar bo'lishi mumkin. An'anaviy ravishda jelatinli yirtqichlar dengiz trofik yo'llarining samarasiz provayderlari deb hisoblangan, ammo ular chuqur pelagik oziq-ovqat tarmoqlarida muhim va ajralmas rollarga ega.[56] Diel vertikal migratsiyasi, muhim faol transport mexanizmi, imkon beradi mezozooplankton atmosferadagi karbonat angidrid gazini ajratish va boshqa mezopelagik organizmlar uchun uglerod ehtiyojini ta'minlash.[57]
2020 yilgi tadqiqotlar shuni ko'rsatdiki, 2050 yilga kelib global isish, hatto issiqxona gazlari chiqarilishi kamaygan taqdirda ham, okean tubida hozirgi darajadan etti baravar tezroq tarqalishi mumkin. Isitish mezopelagik va chuqur qatlamlar chuqur okean oziq-ovqat tarmog'i uchun katta oqibatlarga olib kelishi mumkin, chunki okean turlari omon qolish haroratida turish uchun harakatlanishi kerak.[58][59]
- Alacakaranlıkta baliqlar okean ekotizimiga yangi nur sochdi Suhbat, 2014 yil 10-fevral.
- Trillionlarda okean sirlari The New York Times, 2015 yil 29-iyun.
- Mesopelagik baliqlar - Malaspina atrofini aylanib chiqish ekspeditsiyasi 2010 yil.[60][61]
Okean sathida

Okean sathidagi yashash joylari okean va atmosfera oralig'ida joylashgan. The biofilmga o'xshash okean sathidagi yashash joylari odatda yashaydigan mikroorganizmlarni yashaydi neuston. Ushbu ulkan havo-suv interfeysi global sirt maydonining 70 foizidan ko'prog'ini o'z ichiga olgan asosiy havo-suv almashinuvi jarayonlari chorrahasida joylashgan. Okean yuzaki mikro qatlamidagi bakteriyalar, deyiladi bakterionuston, parnik gazlarining havo-dengiz gazlari almashinuvi, iqlim-faol dengiz aerozollarini ishlab chiqarish va okeanni masofadan zondlash kabi amaliy qo'llanmalar tufayli qiziqish uyg'otmoqda.[63] Ishlab chiqarish va degradatsiyasi o'ziga xos qiziqish uyg'otadi sirt faol moddalar (sirt faol materiallar) mikrobial biokimyoviy jarayonlar orqali. Okeanning sirt faol moddalarining asosiy manbalariga fitoplankton,[64] er usti oqimi va atmosferadan cho'kish.[63]
Rangli alg gullaridan farqli o'laroq, sirt faol moddalar bilan bog'liq bakteriyalar okean rangidagi tasvirlarda ko'rinmasligi mumkin. Ushbu "ko'rinmas" sirt faol moddalar bilan bog'liq bakteriyalarni aniqlash qobiliyatiga ega bo'lish sintetik diafragma radar ob-havo sharoitida, bulut, tuman yoki kunduzgi yorug'likdan qat'i nazar, juda katta foyda keltiradi.[63] Bu juda kuchli shamollarda juda muhimdir, chunki bu eng kuchli havo-dengiz gaz almashinuvi va dengiz aerozollarini ishlab chiqarish sharoitida. Shu sababli, sun'iy yo'ldoshli tasvirlardan tashqari, SAR sun'iy yo'ldosh tasvirlari okean va atmosfera chegarasida biofizik jarayonlarning global surati, havo-dengiz parnik gazlari almashinuvi va iqlim-faol dengiz aerozollarini ishlab chiqarish to'g'risida qo'shimcha ma'lumot berishi mumkin.[63]
Okean tubida
Okean tubi (bentik ) yashash joylari okean va erning ichki qismi o'rtasida joylashgan.
- Oqish va shamollatish
Sohil tarmoqlari

Sohil suvlari ichiga suvlar kiradi daryolar va ustidan kontinental javonlar. Ular umumiy okean maydonining taxminan 8 foizini egallaydi[70] va butun okean unumdorligining taxminan yarmini tashkil etadi.
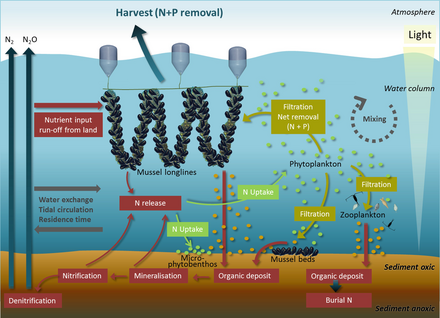
Asosiy oziq moddalar evrofikatsiya qirg'oq suvlarida azot va ko'llarda fosfor. Ikkalasi ham yuqori konsentratsiyalarda uchraydi guano (dengiz qushi najasi), u atrofdagi okean yoki unga qo'shni ko'l uchun o'g'it vazifasini bajaradi. Siydik kislotasi dominant azot birikmasi bo'lib, uning minerallashuvi jarayonida har xil azot shakllari hosil bo'ladi. O'ngdagi diagrammada: (1) ammonifikatsiya NH3 va NH4 + ni hosil qiladi va (2) nitrifikatsiya NO3− ni NH4 + oksidlanish bilan hosil qiladi. Dengiz qushi najasiga xos bo'lgan ishqoriy sharoitda NH3 tez o'zgarib turadi (3) va NH4 + ga aylanadi, u koloniyadan tashqariga chiqarib yuboriladi va evtrofize qilingan uzoq ekotizimlarga nam-cho'kma orqali eksport qilinadi (4). Fosfor tsikli oddiyroq va harakatchanlikni pasaytiradi. Ushbu element dengiz qushi najas materialida bir qator kimyoviy shakllarda uchraydi, ammo eng harakatchan va biologik mavjud ortofosfat bo'lishi mumkin yuvilgan er osti yoki yuzaki suvlar bilan (5).[69]

qit'a tokchasida
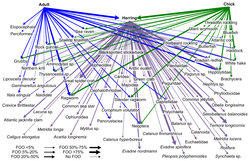
DNKning shtrix-kodi veb-tugunlarida taksonomik aniqligi bilan oziq-ovqat veb-tuzilmalarini qurish uchun ishlatilishi mumkin. Bu aniqroq turlarni aniqlash va kim kimni yeyishi haqida aniqroq ma'lumot beradi. "DNK shtrix-kodlari va DNK ma'lumotlari o'zaro ta'sirlashadigan yirik tarmoqlarni qurishda yangi yondashuvlarni amalga oshirishi va namuna hajmini olish uchun ba'zi to'siqlarni engib chiqishi mumkin"[30]
Turlarni identifikatsiyalash uchun yangi qo'llaniladigan usul DNK metabarkodlashdir. Orqali turlarni aniqlash morfologiya nisbatan qiyin va ko'p vaqt va tajribani talab qiladi.[79][80] Yuqori samaradorlikni ketma-ketligi DNKning metabarkodlashi taksonomik tayinlashga imkon beradi va shuning uchun guruhga xos bo'lgan to'liq namunani aniqlaydi astarlar oldingi DNK uchun tanlangan kuchaytirish.

- Mikrobial DNKning shtrix-kodi
- Yosunlarning DNK-shtrix-kodi
- Baliqning DNK shtrix-kodi
- Diyetani baholashda DNKning shtrix-kodi
- Kelp o'rmonlari
- Byrnes, JE, Reynolds, P.L. va Stachowicz, J.J. (2007) "Bosqinlar va yo'q bo'lib ketish dengiz sohilidagi oziq-ovqat tarmoqlarini qayta shakllantiradi". PLOS ONE, 2(3): e295. doi:10.1371 / journal.pone.0000295
Qutbiy tarmoqlar
okeanlar bilan o'ralgan
quruqlik bilan o'ralgan
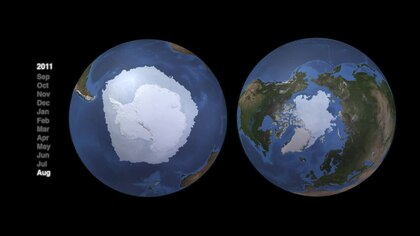
Arktika va Antarktida dengiz tizimlari juda boshqacha topografik tuzilmalar va natijada turli xil oziq-ovqat veb-tuzilmalari mavjud.[82] Arktika va Antarktika pelagik oziq-ovqat tarmoqlari asosan bir nechta asosiy turlar tomonidan boshqariladigan xarakterli energiya oqimlariga ega. Ammo ikkalasi uchun ham bitta umumiy veb-sayt mavjud emas. Muqobil yo'llar chidamlilik va energiya oqimlarini saqlash uchun muhimdir. Biroq, ushbu murakkab alternativalar yuqori trofik darajadagi turlarga kamroq energiya oqimini ta'minlaydi. "Oziq-ovqat veb-tuzilishi turli mintaqalarda o'xshash bo'lishi mumkin, ammo o'rtacha trofik darajalarda hukmronlik qiladigan alohida turlar qutb mintaqalarida farq qiladi".[83]



- Arktika
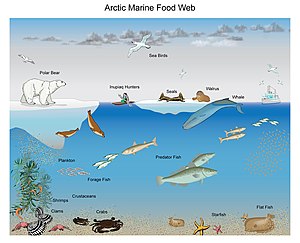
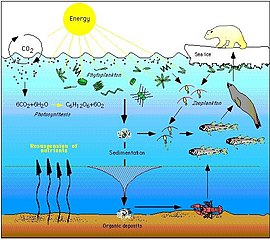
Arktikadagi oziq-ovqat tarmog'i murakkab. Dengiz muzining yo'qolishi oxir-oqibat suv o'tlari va planktonlardan baliqlarga, sutemizuvchilargacha bo'lgan oziq-ovqat tarmog'iga ta'sir qilishi mumkin. Iqlim o'zgarishining ma'lum bir turga ta'siri oziq-ovqat tarmog'i orqali o'tishi va boshqa ko'plab boshqa organizmlarga ta'sir qilishi mumkin ... Dengiz muzlarining pasayishi nafaqat oqsil ayiqlarining populyatsiyasini ularning yashash muhitini kamaytirish orqali, balki ularga veb-effektlar orqali salbiy ta'sir ko'rsatmoqda. Arktikada dengiz muzining davomiyligi va darajasining pasayishi muzdagi ozuqaviy moddalarga boy cho'ntaklarda yaxshi rivojlanadigan muz suv o'tlari ko'pligining pasayishiga olib keladi. Ushbu suv o'tlari zooplankton tomonidan iste'mol qilinadi, ular esa o'z navbatida ko'plab dengiz sutemizuvchilari, shu jumladan muhrlar uchun muhim oziq-ovqat manbai bo'lgan Arktika cod tomonidan iste'mol qilinadi. Muhrlarni oq ayiqlar yeydi. Demak, muz yosunlarining kamayishi oq ayiqlar populyatsiyasining kamayishiga yordam beradi.[84]
2020 yilda tadqiqotchilar so'nggi 20 yil ichida o'lchovlar o'tkazilganligi haqida xabar berishdi birlamchi ishlab chiqarish ichida Shimoliy Muz okeani ning yuqori konsentratsiyasi tufayli qariyb 60% ga o'sishini ko'rsatadi fitoplankton. Ular boshqa okeanlardan yangi ozuqa moddalari oqayotgani haqida taxmin qilishadi va bu Shimoliy Muz okeanini qo'llab-quvvatlashi mumkin degan ma'noni anglatadi. trofik darajadagi yuqori ishlab chiqarish va qo'shimcha uglerod birikmasi kelajakda.[85][86]


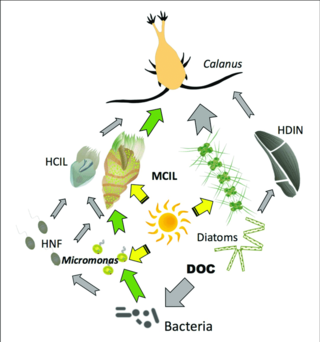
Kulrang o'qlar: uglerodning geterotroflarga oqishi
Yashil o'qlar: uglerod oqimining mikotroflarga tushadigan yoki o'tadigan asosiy yo'llari
HCIL: geterotrofik kirpiklar; MCIL: mikotrofik siliyatlar; HNF: geterotrofik nanoflagellatlar; DOC: erigan organik uglerod; HDIN: heterotrofik dinoflagellatlar[87]

- Antarktika

Antarktika meduzasi Diplulmaris antarktida muz ostida

Yosun koloniyalari Feokistis antarktida Ross dengizining muhim fitoplanteri, erta mavsumda hukmronlik qiladi, dengiz muzlari chekingandan keyin gullaydi va muhim uglerod eksport qiladi.[89]

The qalamcha diatom Fragilariopsis kerguelensis, Antarktika sirkumpolyar oqimi bo'ylab topilgan, global miqyosdagi asosiy harakatlantiruvchi vositadir silikat nasosi.[90]

Bir guruh qotil kitlar ko'chirishga urinish a Qisqichbaqa muhri bo'yicha muz parchasi




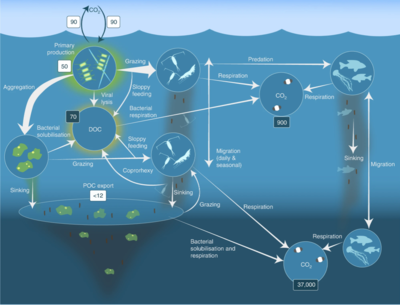
va qorong'i qutilarda uglerod massalari (Gt C)


Poydevor va asosiy tosh turlari



Poydevor turlari are species that have a dominant role structuring an ecological community, shaping its environment and defining its ecosystem. Such ecosystems are often named after the foundation species, such as seagrass meadows, oyster beds, coral reefs, kelp forests and mangrove forests.[97] Masalan, qizil mangrov is a common foundation species in mangrove forests. The mangrove’s root provides nursery grounds for young fish, such as qistirmoq.[98] A foundation species can occupy any trophic level in a food web but tend to be a producer.[99]
The term was coined in 1972 by Paul K. Dayton,[100] who applied it to certain members of marine umurtqasizlar va suv o'tlari jamoalar. It was clear from studies in several locations that there were a small handful of species whose activities had a disproportionate effect on the rest of the marine community and they were therefore key to the resilience of the community. Dayton’s view was that focusing on foundation species would allow for a simplified approach to more rapidly understand how a community as a whole would react to disturbances, such as pollution, instead of attempting the extremely difficult task of tracking the responses of all community members simultaneously.


Keystone turlari are species that have large effects, disproportionate to their numbers, within ecosystem food webs.[101] An ecosystem may experience a dramatic shift if a keystone species is removed, even though that species was a small part of the ecosystem by measures of biomassa yoki hosildorlik.[102]
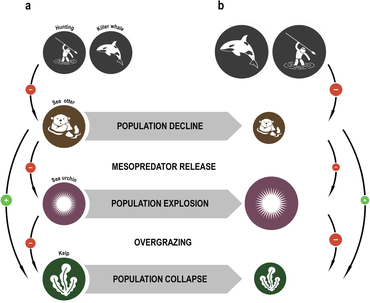
The concept of the keystone species was introduced in 1969 by the zoologist Robert T. Peyn.[103][104] Paine developed the concept to explain his observations and experiments on the relationships between dengiz umurtqasizlari ning intertidal zona (between the high and low tide lines), including dengiz yulduzi va Midiya. Biroz dengiz yulduzlari prey on dengiz kirpi, Midiya va boshqalar qisqichbaqalar that have no other natural predators. If the sea star is removed from the ecosystem, the mussel population explodes uncontrollably, driving out most other species.[105]
Dengiz otasi limit the damage sea urchins inflict on suv o'tlari o'rmonlari. When the sea otters of the North American west coast were hunted commercially for their fur, their numbers fell to such low levels that they were unable to control the sea urchin population. The urchins in turn grazed the ushlab turish ning kelp so heavily that the kelp forests largely disappeared, along with all the species that depended on them. Reintroducing the sea otters has enabled the kelp ecosystem to be restored.[106][107]
Cryptic interactions

Cryptic interactions, interactions which are "hidden in plain sight", occur throughout the marine planktonic foodweb but are currently largely overlooked by established methods, which mean large‐scale data collection for these interactions is limited. Despite this, current evidence suggests some of these interactions may have perceptible impacts on foodweb dynamics and model results. Incorporation of cryptic interactions into models is especially important for those interactions involving the transport of nutrients or energy.[108]
Simplifications such as “zooplankton consume phytoplankton,” “phytoplankton take up inorganic nutrients,” “gross primary production determines the amount of carbon available to the foodweb,” etc. have helped scientists explain and model general interactions in the aquatic environment. Traditional methods have focused on quantifying and qualifying these generalizations, but rapid advancements in genomics, sensor detection limits, experimental methods, and other technologies in recent years have shown that generalization of interactions within the plankton community may be too simple. These enhancements in technology have exposed a number of interactions which appear as cryptic because bulk sampling efforts and experimental methods are biased against them.[108]
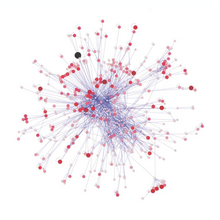
Murakkablik va barqarorlik
Oziq-ovqat tarmoqlari provide a framework within which a complex network of predator–prey interactions can be organised. A food web model is a network of oziq-ovqat zanjirlari. Each food chain starts with a primary producer yoki avtotrof, an organism, such as an alga or a plant, which is able to manufacture its own food. Next in the chain is an organism that feeds on the primary producer, and the chain continues in this way as a string of successive predators. The organisms in each chain are grouped into trofik sathlar, based on how many links they are removed from the primary producers. The length of the chain, or trophic level, is a measure of the number of species encountered as energy or nutrients move from plants to top predators.[111] Oziq-ovqat energiyasi flows from one organism to the next and to the next and so on, with some energy being lost at each level. At a given trophic level there may be one species or a group of species with the same predators and prey.[112]
1927 yilda, Charlz Elton published an influential synthesis on the use of food webs, which resulted in them becoming a central concept in ecology.[113] In 1966, interest in food webs increased after Robert Paine's experimental and descriptive study of intertidal shores, suggesting that food web complexity was key to maintaining species diversity and ecological stability.[114] Many theoretical ecologists, including Robert May va Styuart Pimm, were prompted by this discovery and others to examine the mathematical properties of food webs. According to their analyses, complex food webs should be less stable than simple food webs.[115]:75–77[116]:64 The apparent paradox between the complexity of food webs observed in nature and the mathematical fragility of food web models is currently an area of intensive study and debate. The paradox may be due partially to conceptual differences between persistence of a food web and equilibrial barqarorlik of a food web.[115][116]
A trofik kaskad can occur in a food web if a trophic level in the web is suppressed.
For example, a top-down cascade can occur if predators are effective enough in predation to reduce the abundance, or alter the behavior, of their o'lja, thereby releasing the next lower trophic level from predation. A top-down cascade is a trophic cascade where the top consumer/predator controls the primary consumer aholi. In turn, the primary producer population thrives. The removal of the top predator can alter the food web dynamics. In this case, the primary consumers would overpopulate and exploit the primary producers. Eventually there would not be enough primary producers to sustain the consumer population. Top-down food web stability depends on competition and predation in the higher trophic levels. Invasive species can also alter this cascade by removing or becoming a top predator. This interaction may not always be negative. Studies have shown that certain invasive species have begun to shift cascades; and as a consequence, ecosystem degradation has been repaired.[117][118] An example of a cascade in a complex, open-ocean ecosystem occurred in the northwest Atlantika 1980 va 1990 yillar davomida. Olib tashlash Atlantika cod (Gadus morhua) and other ground fishes by sustained ortiqcha baliq ovlash resulted in increases in the abundance of the prey species for these ground fishes, particularly smaller em-xashak baliqlari and invertebrates such as the northern qor qisqichbaqasi (Chionoecetes opilio) va shimoliy mayda qisqichbaqa (Pandalus borealis). The increased abundance of these prey species altered the community of zooplankton that serve as food for smaller fishes and invertebrates as an indirect effect.[119] Top-down cascades can be important for understanding the knock-on effects of removing top predators from food webs, as humans have done in many places through ov qilish va baliq ovlash.
In a bottom-up cascade, the population of primary producers will always control the increase/decrease of the energy in the higher trophic levels. Primary producers are plants, phytoplankton and zooplankton that require photosynthesis. Although light is important, primary producer populations are altered by the amount of nutrients in the system. This food web relies on the availability and limitation of resources. All populations will experience growth if there is initially a large amount of nutrients.[120][121]
Terrestrial comparisons

Compared to terrestrial biomass pyramids, aquatic pyramids are generally inverted at the base


Marine environments can have inversions in their biomass pyramids. In particular, the biomass of consumers (copepods, krill, shrimp, forage fish) is generally larger than the biomass of primary producers. This happens because the ocean's primary producers are mostly tiny phytoplankton which have r-strategist traits of growing and reproducing rapidly, so a small mass can have a fast rate of primary production. In contrast, many terrestrial primary producers, such as mature forests, have K-strategist traits of growing and reproducing slowly, so a much larger mass is needed to achieve the same rate of primary production. The rate of production divided by the average amount of biomass that achieves it is known as an organism's Production/Biomass (P/B) ratio.[123] Production is measured in terms of the amount of movement of mass or energy per area per unit of time. In contrast, the biomass measurement is in units of mass per unit area or volume. The P/B ratio utilizes inverse time units (example: 1/month). This ratio allows for an estimate of the amount of energy flow compared to the amount of biomass at a given trophic level, allowing for demarcations to be made between trophic levels. The P/B ratio most commonly decreases as trophic level and organismal size increases, with small, ephemeral organisms containing a higher P/B ratio than large, long-lasting ones.
Misollar: The bristlecone qarag'ay can live for thousands of years, and has a very low production/biomass ratio. Siyanobakteriya Proxlorokokk lives for about 24 hours, and has a very high production/biomass ratio.


In oceans, most birlamchi ishlab chiqarish tomonidan amalga oshiriladi suv o'tlari. This is a contrast to on land, where most primary production is performed by qon tomir o'simliklar.
Planktonik suv o'tlari yoki suv o'simliklari kabi suv ishlab chiqaruvchilarida katta miqdordagi birikmalar mavjud emas ikkilamchi o'sish that exists in the woody trees of terrestrial ecosystems. Biroq, ular o'tlovchilarning katta biomassasini qo'llab-quvvatlash uchun etarlicha tez ko'payishga qodir. Bu piramidani teskari yo'naltiradi. Birlamchi iste'molchilar umr ko'rishlari uzoqroq va o'sish sur'atlari sekinroq, ular iste'mol qilayotgan ishlab chiqaruvchilarga qaraganda ko'proq biomassani to'playdi. Fitoplankton atigi bir necha kun yashaydi, fitoplanktonni iste'mol qiladigan zooplankton bir necha hafta yashaydi va zooplanktonni iste'mol qiladigan baliqlar ketma-ket bir necha yil yashaydi.[125] Suv yirtqichlari, shuningdek, kichik iste'molchilarga qaraganda past o'lim ko'rsatkichiga ega, bu esa teskari piramidal naqshga yordam beradi. Aholining tarkibi, migratsiya darajasi va o'lja uchun ekologik boshpana biomassa teskari o'girilgan piramidalarning boshqa sabablari hisoblanadi. Energy pyramids, however, will always have an upright pyramid shape if all sources of food energy are included, since this is dictated by the termodinamikaning ikkinchi qonuni."[126][127]
Comparison of productivity in marine and terrestrial ecosystems[128] | |||
|---|---|---|---|
| Ekotizim | Sof birlamchi mahsuldorlik billion tonnes per year | Total plant biomass billion tonnes | Aylanma vaqt yil |
Dengiz | 45–55 | 1–2 | 0.02–0.06 |
Quruqlik | 55–70 | 600–1000 | 9–20 |
Anthropogenic effects

- Haddan tashqari baliq ovlash
- Acidification
Pteropods va mo'rt yulduzlar together form the base of the Arctic oziq-ovqat tarmoqlari and both are seriously damaged by acidification. Pteropods shells dissolve with increasing acidification and brittle stars lose muscle mass when re-growing appendages.[130] Bundan tashqari, mo'rt yulduzning tuxumlari Arktikani kislotalash natijasida yuzaga keladigan kutilgan sharoitga duch kelganida bir necha kun ichida nobud bo'ladi.[131] Kislota Arktikadagi oziq-ovqat tarmoqlarini tagidan yuqoriga ko'tarish bilan tahdid qilmoqda. Arctic waters are changing rapidly and are advanced in the process of becoming undersaturated with aragonite.[132] Arktikadagi oziq-ovqat tarmoqlari oddiy deb hisoblanadi, ya'ni ozuqaviy zanjirda kichik organizmlardan kattaroq yirtqichlarga qadar bosqichlar mavjud. Masalan, pteropodlar "bir qancha baland yirtqichlarning asosiy o'ljasi - yirik planktonlar, baliqlar, dengiz qushlari, kitlar" dir.[133]


- Iqlim o'zgarishi
"Our results show how future climate change can potentially weaken marine food webs through reduced energy flow to higher trophic levels and a shift towards a more detritus-based system, leading to food web simplification and altered producer–consumer dynamics, both of which have important implications for the structuring of benthic communities."[134][135]
"...increased temperatures reduce the vital flow of energy from the primary food producers at the bottom (e.g. algae), to intermediate consumers (herbivores), to predators at the top of marine food webs. Such disturbances in energy transfer can potentially lead to a decrease in food availability for top predators, which in turn, can lead to negative impacts for many marine species within these food webs... "Whilst climate change increased the productivity of plants, this was mainly due to an expansion of cyanobacteria (small blue-green algae)," said Mr Ullah. "This increased primary productivity does not support food webs, however, because these cyanobacteria are largely unpalatable and they are not consumed by herbivores. Understanding how ecosystems function under the effects of global warming is a challenge in ecological research. Most research on ocean warming involves simplified, short-term experiments based on only one or a few species."[135]
Shuningdek qarang
Adabiyotlar
- ^ U S Department of Energy (2008) Carbon Cycling and Biosequestration page 81, Workshop report DOE/SC-108, U.S. Department of Energy Office of Science.
- ^ Kempbell, Mayk (2011 yil 22-iyun). "Uglerod sekvestratsiyasida dengiz planktonining roli". EarthTimes. Olingan 22 avgust 2014.
- ^ Why should we care about the ocean? NOAA: National Ocean Service. Updated: 7 January 2020. Retrieved 1 March 2020.
- ^ Heinrichs, M.E., Mori, C. and Dlugosch, L. (2020) "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". In: Jungblut S., Liebich V., Bode-Dalby M. (Eds) YOUMARES 9-Okeanlar: bizning tadqiqotlarimiz, bizning kelajagimiz , 279–297 betlar. Springer. doi:10.1007/978-3-030-20389-4_15.
- ^ Dunne, J.A., Williams, R.J. and Martinez, N.D. (2002) "Food-web structure and network theory: the role of connectance and size". Milliy fanlar akademiyasi materiallari, 99(20): 12917–12922. doi:10.1073 / pnas.192407699.
- ^ Dunne, J.A. (2006) "The network structure of food webs". In: M Pascual and J. A. Dunne (Eds.) Ecological networks: linking structure to dynamics in food webs, pages 27–86. ISBN 9780199775057.
- ^ Karlson, AM, Goroxova, E., Gardmark, A., Pekcan-Hekim, Z., Kasini, M., Albertsson, J., Sundelin, B., Karlsson, O. va Bergström, L. (2020). "Iste'molchilarning fiziologik holatini oziq-ovqat mahsulotlarining veb-tuzilishi va Boltiq dengizidagi o'lja oziq-ovqat qiymati bilan bog'lash". Ambio, 49(2): 391–406. doi:10.1007 / s13280-019-01201-1
- ^ Odum, W. E.; Heald, E. J. (1975) "The detritus-based food web of an estuarine mangrove community". Pages 265–286 in L. E. Cronin, ed. Estuarine research. Vol. 1. Academic Press, New York.
- ^ Pimm, S. L.; Lawton, J. H. (1978). "Bir nechta trofik darajada ovqatlanish to'g'risida". Tabiat. 275 (5680): 542–544. doi:10.1038 / 275542a0. S2CID 4161183.
- ^ Pauly, D.; Palomares, M. L. (2005). "Dengizdagi oziq-ovqat tarmoqlarini baliq ovlash: bu biz o'ylagandan ancha keng tarqalgan" (PDF). Dengizchilik fanlari byulleteni. 76 (2): 197–211. Arxivlandi asl nusxasi (PDF) 2013-05-14.
- ^ Cortés, E. (1999). "Standardized diet compositions and trophic levels of sharks". ICES J. Mar. Sci. 56 (5): 707–717. doi:10.1006/jmsc.1999.0489.
- ^ Pauly, D.; Trites, A.; Capuli, E.; Christensen, V. (1998). "Diet composition and trophic levels of marine mammals". ICES J. Mar. Sci. 55 (3): 467–481. doi:10.1006/jmsc.1997.0280.
- ^ Researchers calculate human trophic level for first time Phys.org . 2013 yil 3-dekabr.
- ^ Bonhommeau, S., Dubroca, L., Le Pape, O., Barde, J., Kaplan, D.M., Chassot, E. and Nieblas, A.E. (2013) "Eating up the world’s food web and the human trophic level". Milliy fanlar akademiyasi materiallari, 110(51): 20617–20620. doi:10.1073 / pnas.1305827110.
- ^ a b Xlorofil NASA Yer Observatoriyasi. Kirish 30 Noyabr 2019.
- ^ Kettler GC, Martiny AC, Huang K, Zucker J, Coleman ML, Rodrigue S, Chen F, Lapidus A, Ferriera S, Johnson J, Steglich C, Church GM, Richardson P, Chisholm SW (December 2007). "Proxlorococcus evolyutsiyasida genlarning ko'payishi va yo'qotilishi naqshlari va oqibatlari". PLOS Genetika. 3 (12): e231. doi:10.1371 / journal.pgen.0030231. PMC 2151091. PMID 18159947.
- ^ Nemiroff, R .; Bonnell, J., nashr. (27 September 2006). "Earth from Saturn". Astronomiya kunining surati. NASA.
- ^ Partensky F, Hess WR, Vaulot D (March 1999). "Proxlorokokk, global ahamiyatga ega dengiz fotosintetik prokaryoti". Mikrobiologiya va molekulyar biologiya sharhlari. 63 (1): 106–27. doi:10.1128/MMBR.63.1.106-127.1999. PMC 98958. PMID 10066832.
- ^ "Siz hech qachon eshitmagan eng muhim mikrob". npr.org.
- ^ Mann, D. G. (1999). "The species concept in diatoms". Fikologiya. 38 (6): 437–495. doi:10.2216/i0031-8884-38-6-437.1.
- ^ Kopepodlar biologiyasi Arxivlandi 2009-01-01 da Orqaga qaytish mashinasi da Oldenburgdagi Karl fon Ossiyetskiy universiteti
- ^ a b v d Hays, G.C., Doyle, T.K. and Houghton, J.D. (2018) "A paradigm shift in the trophic importance of jellyfish?" Trends in ecology & evolution, 33(11): 874-884. doi:10.1016 / j.tree.2018.09.001
- ^ a b Xemilton, G. (2016) "Meduzalarning maxfiy hayoti: uzoq vaqt okean ekologiyasining kichik ishtirokchilari deb hisoblangan meduzalar aslida dengiz oziq-ovqat tarmog'ining muhim qismidir". Tabiat, 531(7595): 432-435. doi:10.1038 / 531432a
- ^ Cardona, L., De Quevedo, I.Á., Borrell, A. va Agilar, A. (2012) "O'rta er dengizi cho'qqisi yirtqichlari tomonidan jelatinli planktonni katta miqdorda iste'mol qilish". PLOS ONE, 7(3): e31329. doi:10.1371 / journal.pone.0031329
- ^ Tiny Forage Fish At Bottom Of Marine Food Web Get New Protections Milliy jamoat radiosi, 2016 yil 7 aprel.
- ^ Roman, J. va Makkarti, JJ (2010). "Kit nasosi: dengiz sutemizuvchilari qirg'oq havzasida birlamchi mahsuldorlikni oshiradi". PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007. e13255.CS1 maint: mualliflar parametridan foydalanadi (havola)
- ^ Brown, Joshua E. (2010 yil 12-oktabr). "Whale poop pumps up ocean health". Science Daily. Olingan 18 avgust 2014.
- ^ Raina, J.B. (2018) "The life aquatic at the microscale". m tizimlari, 3(2): e00150-17. doi:10.1128/mSystems.00150-17.
 Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi. - ^ Marco, D, ed. (2011). Metagenomics: Current Innovations and Future Trends. Caister Academic Press. ISBN 978-1-904455-87-5.
- ^ a b v Roslin, T. and Majaneva, S. (2016) "The use of DNA barcodes in food web construction—terrestrial and aquatic ecologists unite!". Genom, 59(9): 603–628. doi:10.1139/gen-2015-0229.
- ^ Wilhelm, Steven W.; Suttle, Curtis A. (1999). "Viruses and Nutrient Cycles in the Sea". BioScience. 49 (10): 781–788. doi:10.2307/1313569. ISSN 1525-3244. JSTOR 1313569.
- ^ Weinbauer, Markus G., et al. "Synergistic and antagonistic effects of viral lysis and protistan grazing on bacterial biomass, production and diversity." Atrof-muhit mikrobiologiyasi 9.3 (2007): 777-788.
- ^ Robinson, Carol, and Nagappa Ramaiah. "Microbial heterotrophic metabolic rates constrain the microbial carbon pump." The American Association for the Advancement of Science, 2011.
- ^ Van den Meersche, K., Middelburg, J.J., Soetaert, K., Van Rijswijk, P., Boschker, H.T. and Heip, C.H. (2004) "Carbon‐nitrogen coupling and algal‐bacterial interactions during an experimental bloom: Modeling a 13C tracer experiment". Limnologiya va okeanografiya, 49(3): 862–878. doi:10.4319 / lo.2004.49.3.0862.
- ^ Käse L, Geuer JK. (2018) "Phytoplankton Responses to Marine Climate Change – An Introduction". In Jungblut S., Liebich V., Bode M. (Eds) YOUMARES 8–Oceans Across Boundaries: Learning from each other, pages 55–72, Springer. doi:10.1007/978-3-319-93284-2_5. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Heinrichs, ME, Mori, C. and Dlugosch, L. (2020) "Suvda yashovchi organizmlar va ularning turli xil nuqtai nazardan yoritilgan kimyoviy muhiti o'rtasidagi murakkab o'zaro ta'sirlar". In: YOUMARES 9-Okeanlar: bizning tadqiqotlarimiz, bizning kelajagimiz , 279–297 betlar. Springer. doi:10.1007/978-3-030-20389-4_15.
- ^ Duponchel, S. and Fischer, M.G. (2019) "Viva lavidaviruslari! Gigant DNK viruslarini parazit qiladigan virusofaglarning beshta xususiyati". PLoS patogenlari, 15(3). doi:10.1371 / journal.ppat.1007592. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Fischer, M. G.; Allen, M. J .; Uilson, V. X.; Suttle, C. A. (2010). "Gendagi ajoyib komplektli ulkan virus dengiz zooplanktonini yuqtiradi" (PDF). Milliy fanlar akademiyasi materiallari. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073 / pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Matthias G. Fischer; Maykl J. Allen; Uilyam H. Uilson; Kurtis A. Suttle (2010). "Gendagi ajoyib komplektli ulkan virus dengiz zooplanktonini yuqtiradi" (PDF). Milliy fanlar akademiyasi materiallari. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073 / pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Massana, Ramon; Javier Del Campo; Christian Dinter; Ruben Sommaruga (2007). "Crash of a population of the marine heterotrophic flagellate Cafeteria roenbergensis by viral infection". Atrof-muhit mikrobiologiyasi. 9 (11): 2660–2669. doi:10.1111/j.1462-2920.2007.01378.x. PMID 17922751. S2CID 30191542.
- ^ Fischer MG, Suttle CA (aprel 2011). "Katta DNK transpozonlari kelib chiqishidagi virusofag". Ilm-fan. 332 (6026): 231–4. Bibcode:2011 yil ... 332..231F. doi:10.1126 / science.1199412. PMID 21385722. S2CID 206530677.
- ^ Fischer MG, Hackl (2016 yil dekabr). "Xost genomining integratsiyasi va virusofag mavirusining ulkan virus ta'sirida reaktivatsiyasi". Tabiat. 540 (7632): 288–91. Bibcode:2016 yil natur.540..288F. doi:10.1038 / tabiat20593. PMID 27929021. S2CID 4458402.
- ^ Kagami, M., Miki, T. and Takimoto, G. (2014) "Mycoloop: chytrids in aquatic food webs". Frontiers in microbiology, 5: 166. doi:10.3389/fmicb.2014.00166. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 3.0 International License.
- ^ Amend, A., Burgaud, G., Cunliffe, M., Edgcomb, V.P., Ettinger, C.L., Gutiérrez, M.H., Heitman, J., Hom, E.F., Ianiri, G., Jones, A.C. and Kagami, M. (2019) "Fungi in the marine environment: Open questions and unsolved problems". MBio, 10(2): e01189-18. doi:10.1128/mBio.01189-18. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Gutierrez MH, Jara AM, Pantoja S (2016) "Fungal parasites infect marine diatoms in the upwelling ecosystem of the Humboldt current system off central Chile". Environ Microbiol, 18(5): 1646–1653. doi:10.1111/1462-2920.13257.
- ^ a b v Middelburg J.J. (2019) "The Return from Organic to Inorganic Carbon". In: Marine Carbon Biogeochemistry: A Primer for Earth System Scientists, pages 37–56, Springer. doi:10.1007/978-3-030-10822-9_3. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Legendre L, Rassoulzadegan F (1995) "Plankton and nutrient dynamics in marine waters". Ofeliya, 41:153–172.
- ^ Pace ML, Cole JJ, Carpenter SR, Kitchell JF (1999) "Trophic cascades revealed in diverse ecosystems". Trends Ecol Evol, 14: 483–488.
- ^ Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F (1983) "The ecological role of water-column microbes in the sea". Mar Ecol-Prog Ser, 10: 257–263.
- ^ Anderson TR and LeB Williams PJ (1998) "Modelling the seasonal cycle of dissolved organic carbonat station E1 in the English channel". Estuar Coast Shelf Sci, 46: 93–109.
- ^ Van den Meersche K, Middelburg JJ, Soetaert K, van Rijswijk P, Boschker HTS, Heip CHR (2004) "Carbon–nitrogen coupling and algal–bacterial interactions during an experimental bloom: modeling a 13C tracer experiment". Limnol Oceanogr, 49: 862–878.
- ^ Suttle CA (2005) "Viruses in the sea". Tabiat, 437: 356–361.
- ^ Siegel, David A.; Buesseler, Ken O.; Behrenfeld, Michael J.; Benitez-Nelson, Claudia R.; Boss, Emmanuel; Brzezinski, Mark A.; Burd, Adrian; Carlson, Craig A.; d'Asaro, Eric A.; Doney, Skott S.; Perry, Mary J.; Stanley, Rachel H. R.; Steinberg, Deborah K. (2016). "Prediction of the Export and Fate of Global Ocean Net Primary Production: The EXPORTS Science Plan". Dengiz fanidagi chegara. 3. doi:10.3389/fmars.2016.00022. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Wang, F., Wu, Y., Chen, Z., Zhang, G., Zhang, J., Zheng, S. and Kattner, G. (2019) "Trophic interactions of mesopelagic fishes in the South China Sea illustrated by stable isotopes and fatty acids". Dengiz fanidagi chegara, 5: 522. doi:10.3389/fmars.2018.00522.
- ^ a b Tollefson, Jeff (27 February 2020) Enter the twilight zone: scientists dive into the oceans’ mysterious middle Tabiat yangiliklari. doi:10.1038/d41586-020-00520-8.
- ^ a b Choy, CA, Haddok, S.H. va Robison, B.H. (2017) "tomonidan aniqlangan chuqur pelagik oziq-ovqat veb-tuzilishi joyida oziqlantirish bo'yicha kuzatuvlar ". Qirollik jamiyati materiallari B: Biologiya fanlari, 284(1868): 20172116. doi:10.1098 / rspb.2017.2116. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Kelly, T.B., Davison, P.C., Goericke, R., Landry, M.R., Ohman, M. and Stukel, M.R. (2019) "The importance of mesozooplankton diel vertical migration for sustaining a mesopelagic food web". Dengiz fanidagi chegara, 6: 508. doi:10.3389/fmars.2019.00508.
- ^ Climate change in deep oceans could be seven times faster by middle of century, report says Guardian, 25 may 2020 yil.
- ^ Brito-Morales, I., Schoeman, D.S., Molinos, J.G., Burrows, M.T., Klein, C.J., Arafeh-Dalmau, N., Kaschner, K., Garilao, C., Kesner-Reyes, K. and Richardson, A.J. (2020) "Climate velocity reveals increasing exposure of deep-ocean biodiversity to future warming". Tabiat iqlimining o'zgarishi, pp.1-6. doi:10.5281/zenodo.3596584.
- ^ Irigoien, X., Klevjer, T.A., Røstad, A., Martinez, U., Boyra, G., Acuña, J.L., Bode, A., Echevarria, F., Gonzalez-Gordillo, J.I., Hernandez-Leon, S. and Agusti, S. (2014) "Large mesopelagic fishes biomass and trophic efficiency in the open ocean". Tabiat aloqalari, 5: 3271. doi:10.1038 / ncomms4271
- ^ Fish biomass in the ocean is 10 times higher than estimated EurekAlert, 7 February 2014.
- ^ Choy, C.A., Wabnitz, C.C., Weijerman, M., Woodworth-Jefcoats, P.A. and Polovina, J.J. (2016) "Finding the way to the top: how the composition of oceanic mid-trophic micronekton groups determines apex predator biomass in the central North Pacific". Dengiz ekologiyasi taraqqiyoti seriyasi, 549: 9–25. doi:10.3354/meps11680.
- ^ a b v d e Kurata, N., Vella, K., Hamilton, B., Shivji, M., Soloviev, A., Matt, S., Tartar, A. and Perrie, W. (2016) "Surfactant-associated bacteria in the near-surface layer of the ocean". Nature: Scientific Reports, 6(1): 1–8. doi:10.1038/srep19123. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Ẑutić, V., Ćosović, B., Marčenko, E., Bihari, N. and Kršinić, F. (1981) "Surfactant production by marine phytoplankton". Dengiz kimyosi, 10(6): 505–520. doi:10.1016/0304-4203(81)90004-9.
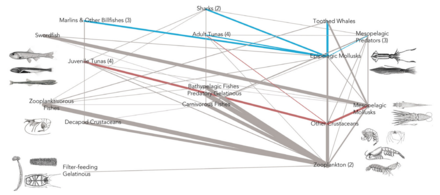
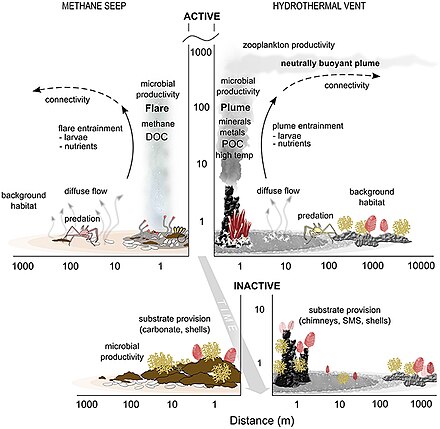
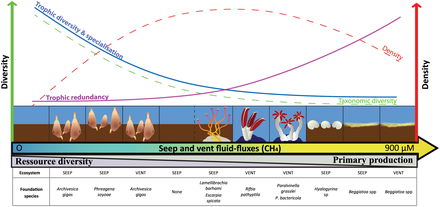
- ^ Levin LA, Baco AR, Bowden DA, Colaco A, Cordes EE, Cunha MR, Demopoulos AWJ, Gobin J, Grupe BM, Le J, Metaxas A, Netburn AN, Rouse GW, Thurber AR, Tunnicliffe V, Van Dover CL, Vanreusel A and Watling L (2016). "Hydrothermal Vents and Methane Seeps: Rethinking the Sphere of Influence". Old. Mar Sci. 3:72. doi:10.3389/fmars.2016.00072
- ^ Portail, M., Olu, K., Dubois, S.F., Escobar-Briones, E., Gelinas, Y., Menot, L. and Sarrazin, J. (2016). "Food-web complexity in Guaymas Basin hydrothermal vents and cold seeps". PLOS ONE, 11(9): p.e0162263. doi:10.1371/journal.pone.0162263.
- ^ Bernardino AF, Levin LA, Thurber AR and Smith CR (2012). "Comparative Composition, Diversity and Trophic Ecology of Sediment Macrofauna at Vents, Seeps and Organic Falls". PLOS ONE, 7(4): e33515. pmid:22496753. doi:10.1371/journal.pone.0033515.
- ^ Portail M, Olu K, Escobar-Briones E, Caprais JC, Menot L, Waeles M, et al. (2015). "Comparative study of vent and seep macrofaunal communities in the Guaymas Basin". Biogeoscience. 12(18): 5455–79. doi:10.5194/bg-12-5455-2015.
- ^ a b Otero, X.L., De La Peña-Lastra, S., Pérez-Alberti, A., Ferreira, T.O. and Huerta-Diaz, M.A. (2018) "Seabird colonies as important global drivers in the nitrogen and phosphorus cycles". Tabiat aloqalari, 9(1): 1–8. doi:10.1038/s41467-017-02446-8. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Xarris, P.T .; Makmillan-Lawler, M.; Rupp, J .; Beyker, E.K. (2014). "Okeanlar geomorfologiyasi". Dengiz geologiyasi. 352: 4–24. doi:10.1016 / j.margeo.2014.01.011.
- ^ Heymans, J.J., Coll, M., Libralato, S., Morissette, L. and Christensen, V. (2014). "Global patterns in ecological indicators of marine food webs: a modelling approach". PLOS ONE, 9(4). doi:10.1371/journal.pone.0095845.
- ^ Pranovi, F., Libralato, S., Raicevich, S., Granzotto, A., Pastres, R. and Giovanardi, O. (2003). "Mechanical clam dredging in Venice lagoon: ecosystem effects evaluated with a trophic mass-balance model". Dengiz biologiyasi, 143(2): 393–403. doi:10.1007/s00227-003-1072-1.
- ^ Petersen, J.K., Holmer, M., Termansen, M. and Hasler, B. (2019) "Nutrient extraction through bivalves". In: Smaal A., Ferreira J., Grant J., Petersen J., Strand Ø. (tahrir) Dengiz ikkilamchi mollari va xizmatlari, pages 179–208. Springer. doi:10.1007/978-3-319-96776-9_10. ISBN 9783319967769
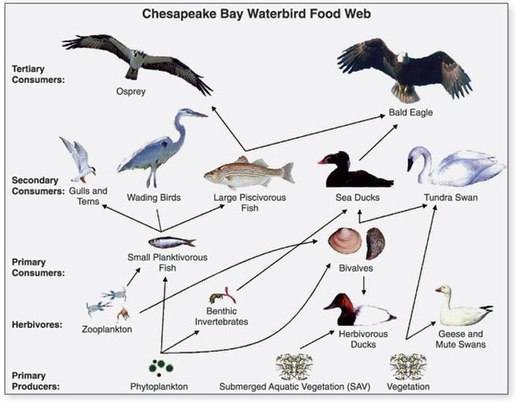
- ^ AQSh Geologik xizmati (USGS). "Chapter 14: Changes in Food and Habitats of Waterbirds." Figure 14.1. Synthesis of U.S. Geological Survey Science for the Chesapeake Bay Ecosystem and Implications for Environmental Management. USGS Circular 1316.
 Ushbu maqola ushbu manbadagi matnni o'z ichiga oladi jamoat mulki.
Ushbu maqola ushbu manbadagi matnni o'z ichiga oladi jamoat mulki. - ^ Perry, M.C., Osenton, P.C., Wells-Berlin, A.M., and Kidwell, D.M., 2005, Food selection among Atlantic Coast sea ducks in relation to historic food habits, [abs.] in Perry, M.C., Second North American Sea Duck Conference, November 7–11, 2005, Annapolis, Maryland, Program and Abstracts, USGS Patuxent Wildlife Research Center, Maryland, 123 p. (p. 105).
- ^ Bowser, A.K., Diamond, A.W. and Addison, J.A. (2013) "From puffins to plankton: a DNA-based analysis of a seabird food chain in the northern Gulf of Maine". PLOS ONE, 8(12): e83152. doi:10.1371/journal.pone.0083152
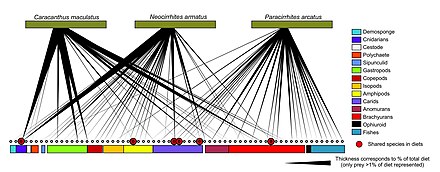
- ^ Leray M, Meyer CP, Mills SC. (2015) "Marjonda yashovchi yirtqich baliqlarning parhez tahlilini metabarkodlash koral mutalististlarining juda bo'linib ketgan, umumiy ovqatlanishiga ozgina qo'shganligini namoyish etadi". PeerJ, 3: e1047. doi:10.7717 / peerj.1047.
- ^ Stat, M., Huggett, MJ, Bernasconi, R., DiBattista, JD, Berri, TE, Nyuman, SJ, Harvey, E.S. va Bunce, M. (2017) "eDNA bilan ekotizim biomonitoringi: tropik dengiz muhitida hayot daraxti bo'ylab metabarkodlash". Ilmiy ma'ruzalar, 7(1): 1–11. doi:10.1038 / s41598-017-12501-5.
- ^ Lobo, Eduardo A.; Geynrix, Karla Giselda; Shuch, Mariliya; Vetsel, Karlos Eduardo; Ektor, Lyuk (2016), Necchi JR, Orlando (tahr.), "Diyatomlar daryolardagi bioindikatorlar sifatida", Daryo suv o'tlari, Springer International Publishing, 245–271 betlar, doi:10.1007/978-3-319-31984-1_11, ISBN 9783319319834
- ^ Stivenson, R. Jan; Pan, Yangdong; van Dam, Xerman (2010), Smol, Jon P.; Stoermer, Eugene F. (tahr.), "Diyotlar va daryolardagi atrof-muhit sharoitlarini diatom bilan baholash", Diatomlar (2-nashr), Kembrij universiteti matbuoti, 57–85-betlar, doi:10.1017 / cbo9780511763175.005, ISBN 9780511763175
- ^ Coll, M., Shmidt, A., Romanuk, T. va Lotze, H.K. (2011). "Turli xil fazoviy o'lchovlar va inson ta'sirlari bo'yicha dengiz o'tlari jamoalarining oziq-ovqat veb-tuzilishi". PLOS ONE, 6(7): e22591. doi:10.1371 / journal.pone.0022591. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Makkarti, JJ, Kanziani, O.F., Leary, N.A., Dokken, DJ va Oq, K.S. (Nashrlar) (2001) Iqlim o'zgarishi 2001 yil: ta'sirlar, moslashish va zaiflik: II ishchi guruhning iqlim o'zgarishi bo'yicha hukumatlararo panelning uchinchi baholash hisobotiga qo'shgan hissasi 807-bet, Kembrij universiteti matbuoti. ISBN 9780521015004
- ^ Murphy, EJ, Cavanagh, RD, Drinkwater, KF, Grant, SM, Heymans, JJ, Hofmann, EE, Hunt Jr, GL va Johnston, NM (2016) "Kutupli pelagik ekotizimlarning tuzilishini va ishlashini tushunish ta'sirini bashorat qilish. o'zgartirish ". Qirollik jamiyati materiallari B: Biologiya fanlari, 283(1844): 20161646. doi:10.1098 / rspb.2016.1646.
- ^ Iqlimning ekotizimlarga ta'siri: Internetdagi oziq-ovqat mahsulotlarining uzilishi EPA. Kirish 11 fevral 2020. Ushbu maqola ushbu manbadagi matnni o'z ichiga oladi jamoat mulki.
- ^ "Shimoliy Muz okeanida" rejim o'zgarishi "sodir bo'lmoqda, deydi olimlar". phys.org. Olingan 16 avgust 2020.
- ^ Lyuis K. M.; Dijken, G. L. van; Arrigo, K. R. (2020 yil 10-iyul). "Fitoplankton kontsentratsiyasining o'zgarishi endi Shimoliy Muz okeanining asosiy ishlab chiqarish hajmini oshirmoqda". Ilm-fan. 369 (6500): 198–202. doi:10.1126 / science.aay8380. ISSN 0036-8075. PMID 32647002. S2CID 220433818. Olingan 16 avgust 2020.
- ^ Stoekker, D.K. va Lavrentyev, PJ (2018). "Qutbiy dengizlarda mikotrofik plankton: panarktik obzor". Dengiz fanidagi chegara, 5: 292. doi:10.3389 / fmars.2018.00292
- ^ Kilias, Estel S.; Jungs, Leandro; Shupraha, Luka; Leonard, Gay; Metfies, Katja; Richards, Tomas A. (2020). "Xitrid zamburug'larining tarqalishi va diatomlar bilan birgalikda uchrashi Shimoliy Muz okeanidagi dengiz muzlarining erishi bilan bog'liq". Aloqa biologiyasi. 3 (1): 183. doi:10.1038 / s42003-020-0891-7. PMC 7174370. PMID 32317738. S2CID 216033140. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ Bender, SJ, Moran, DM, McIlvin, MR, Zheng, H., McCrow, JP, Badger, J., DiTullio, GR, Allen, AE va Saito, MA (2018) "Koloniyaning shakllanishi Feokistis antarktida: molekulyar mexanizmlarni temir biogeokimyosi bilan bog'lash ". Biogeoscience, 15(16): 4923–4942. doi:10.5194 / bg-15-4923-2018.
- ^ Pinkernell, S. va Beszteri, B. (2014) "Iqlim o'zgarishining Janubiy okeanning asosiy silikat cho'ktiruvchisi tarqalish doirasiga ta'siri". Ekologiya va evolyutsiya, 4(16): 3147–3161. doi:10.1002 / ece3.1138
- ^ Cavan, EL, Belcher, A., Atkinson, A., Hill, SL, Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. and Steinberg, DK (2019) "Antarktika krillining biogeokimyoviy davrlarda ahamiyati". Tabiat aloqalari, 10(1): 1–13. doi:10.1038 / s41467-019-12668-7. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
- ^ a b Cordone, G., Marina, TI, Salinas, V., Doyle, SR, Saravia, LA va Momo, FR (2018). "Antarktida dengizidagi oziq-ovqat tarmog'idagi makroalglarni yo'qotish ta'siri: oziq-ovqat veb-tadqiqotida yo'q bo'lish chegaralarini qo'llash". PeerJ, 6: e5531. doi:10.7717 / peerj.5531
- ^ Marina, T.I., Salinas, V., Cordone, G., Campana, G., Moreira, E., Deregibus, D., Torre, L., Sahade, R., Tatian, M., Oro, E.B. va De Troch, M. (2018). "Potter Kovining oziq-ovqat tarmog'i (Antarktida): murakkabligi, tuzilishi va vazifasi". Estuarine, Coastal and Shelf Science, 200: 141-151. doi: 10.1016 / j.ecss.2017.10.015.
- ^ Koh, E.Y., Martin, AR, MakMinn, A. va Rayan, K.G. (2012) "Antarktika dengiz muzidagi mikrobial fototrofiyaning so'nggi yutuqlari va istiqbollari". Biologiya, 1(3): 542-556. doi:10.3390 / biologiya1030542.
- ^ Bu erda mikrobial tsikl qayta chizilgan va qisqartirilgan:
- Azam, F., Fenchel, T., Field, JG, Grey, JS, Meyer-Reil, LA va Thingstad, F. (1983) "Suv ustunli mikroblarning dengizdagi ekologik roli". Dengiz ekologiyasining rivojlanish seriyalari, 10(3): 257–263.
- Fenchel, T. (2008) "Mikrobial tsikl - 25 yildan keyin". Eksperimental dengiz biologiyasi va ekologiyasi jurnali, 366(1-2): 99-103. doi:10.1016 / j.jembe.2008.07.013.
- ^ Lamy, T., Koenigs, C., Holbrook, SJ, Miller, RJ, Stier, AC va Reed, DC (2020) "Jamg'arma turlari ulkan kelp o'rmonida xilma-xillikni oshirish orqali jamoat barqarorligini rivojlantiradi". Ekologiya, e02987. doi:10.1002 / ecy.2987.
- ^ Gigant kelp Janubiy Kaliforniyadagi dengiz ekotizimlariga mustahkam poydevor beradi, Milliy Ilmiy Jamg'arma, 2020 yil 4-fevral.
- ^ Anjelini, Kristin; Altieri, Endryu X.; va boshq. (Oktyabr 2011). "Jamg'arma turlari o'rtasidagi o'zaro munosabatlar va ularning jamoat tashkilotlari, biologik xilma-xillik va tabiatni muhofaza qilish uchun oqibatlari". BioScience. 61 (10): 782–789. doi:10.1525 / bio.2011.61.10.8.
- ^ Ellison, Aaron M.; Bank, Maykl S.; va boshq. (2005 yil noyabr). "Poydevor turlarini yo'qotish: o'rmonli ekotizimlarning tuzilishi va dinamikasi uchun oqibatlar". Ekologiya va atrof-muhit chegaralari. 3 (9): 479–486. doi:10.1890 / 1540-9295 (2005) 003 [0479: LOFSCF] 2.0.CO; 2.
- ^ Dayton, P. K. 1972 yil. Antarktidaning McMurdo Sound-dagi bentoslarga boyitishning potentsial ta'siri va jamoatchilikning barqarorligi to'g'risida tushuncha.. 81-96 betlar. Tabiatni muhofaza qilish muammolari bo'yicha kollokvium materiallari Allen Press, Lourens, Kanzas.
- ^ Paine, R. T. (1995). "Keystone turlari tushunchasini takomillashtirish bo'yicha suhbat". Tabiatni muhofaza qilish biologiyasi. 9 (4): 962–964. doi:10.1046 / j.1523-1739.1995.09040962.x.
- ^ Davic, Robert D. (2003). "Keystone turlari va funktsional guruhlarini bog'lash: Keystone turlari turlari kontseptsiyasining yangi operatsion ta'rifi". Tabiatni muhofaza qilish ekologiyasi. Olingan 2011-02-03.
- ^ Peyn, R. T. (1969). "Trofik murakkablik va jamoat barqarorligi to'g'risida eslatma". Amerikalik tabiatshunos. 103 (929): 91–93. doi:10.1086/282586. JSTOR 2459472. S2CID 83780992.
- ^ "Keystone turlari gipotezasi". Vashington universiteti. Arxivlandi asl nusxasi 2011-01-10. Olingan 2011-02-03.
- ^ Peyn, R. T. (1966). "Oziq-ovqat tarmoqlarining murakkabligi va turlarining xilma-xilligi". Amerikalik tabiatshunos. 100 (910): 65–75. doi:10.1086/282400. JSTOR 2459379. S2CID 85265656.
- ^ Szpak, Pol; Orchard, Trevor J.; Salomon, Anne K.; Grokke, Darren R. (2013). "Mintaqadagi ekologik o'zgaruvchanlik va dengizdagi mo'yna savdosining janubiy Xayday Gvayi (Britaniya Kolumbiyasi, Kanada) dagi qirg'oq ekotizimlariga ta'siri: tosh baliqlarining (Sebastes spp.) Suyak kollagenining izotoplari barqaror tahlilidan dalillar". Arxeologik va antropologik fanlar. 5 (2): 159–182. doi:10.1007 / s12520-013-0122-y. S2CID 84866250.
- ^ Kon, J. P. (1998). "Dengiz samurotlarini tushunish". BioScience. 48 (3): 151–155. doi:10.2307/1313259. JSTOR 1313259.
- ^ a b v Millette, NC, Grosse, J., Jonson, VM, Jungblut, MJ va Suter, E.A. (2018). "Oddiy ko'rinishda yashiringan: dengiz planktonidagi sirli o'zaro ta'sirlarning ahamiyati". Limnologiya va okeanografiya xatlari, 3(4): 341–356. doi:10.1002 / lol2.10084. Ushbu manbadan nusxa ko'chirilgan, u ostida mavjud Creative Commons Attribution 4.0 xalqaro litsenziyasi.
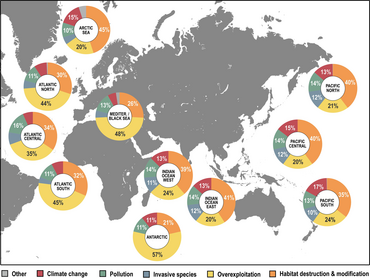
- ^ a b v Luypaert, T., Xagan, JG, Makkarti, ML. va Poti, M. (2020) "Antropotsendagi dengiz biologik xilma-xilligi holati". In: YOUMARES 9-Okeanlar: bizning tadqiqotlarimiz, bizning kelajagimiz, 57-82 betlar, Springer. doi:10.1007/978-3-030-20389-4_4.
- ^ Estes JA, Tinker MT, Uilyams TM va boshq (1998) "Okean va qirg'oq ekotizimlarini bog'laydigan dengiz otterlarida qotil kit o'ljasi". Ilm-fan, 282: 473–476. doi:10.1126 / science.282.5388.473.
- ^ Post, D. M. (1993). "Oziq-ovqat zanjiri uzunligining uzun va kalta". Ekologiya va evolyutsiya tendentsiyalari. 17 (6): 269–277. doi:10.1016 / S0169-5347 (02) 02455-2.
- ^ Jerri Bobrou, tibbiyot fanlari doktori; Stiven Fisher (2009). CliffsNotes CSET: bir nechta mavzular (2-nashr). John Wiley va Sons. p. 283. ISBN 978-0-470-45546-3.
- ^ Elton CS (1927) Hayvonlar ekologiyasi. Qayta nashr etilgan 2001. Chikago universiteti matbuoti.
- ^ Paine RT (1966). "Oziq-ovqat tarmoqlarining murakkabligi va turlarining xilma-xilligi". Amerikalik tabiatshunos. 100 (910): 65–75. doi:10.1086/282400. S2CID 85265656.
- ^ a b May RM (2001) Model ekotizimlarda barqarorlik va murakkablik Prinston universiteti matbuoti, 1973 yil nashr etilgan yangi so'z bilan qayta nashr etildi. ISBN 978-0-691-08861-7.
- ^ a b Pimm SL (2002) Oziq-ovqat veb-saytlari Chikago universiteti matbuoti, 1982 yilgi yangi so'zboshi bilan qayta nashr etilgan. ISBN 978-0-226-66832-1.
- ^ Kotta, J .; Vernberg, T .; Jeyn, X .; Kotta, I .; Nurse K.; Parnoja, M.; Orav-Kotta, H. (2018). "Qisqichbaqa yirtqichi dengiz ekotizimi rejimining o'zgarishiga olib keladi". Ilmiy ma'ruzalar. 8 (1): 4956. doi:10.1038 / s41598-018-23282-w. PMC 5897427. PMID 29651152.
- ^ Megrey, Bernard va Verner, Frantsisko. "Topdown va pastki ekotizim regulyatsiyasining rolini modellashtirish nuqtai nazaridan baholash" (PDF).CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Frank, K. T .; Petri, B.; Choi, J. S .; Leggett, V.S (2005). "Ilgari Cod-ustun bo'lgan ekotizimdagi trofik kaskadlar". Ilm-fan. 308 (5728): 1621–1623. doi:10.1126 / science.1113075. ISSN 0036-8075. PMID 15947186. S2CID 45088691.
- ^ Matsuzaki, Shin-Ichiro S.; Suzuki, Kenta; Kadoya, Taku; Nakagava, Megumi; Takamura, Noriko (2018). "Birlamchi ishlab chiqarish, zooplankton va sayoz, giperutrofik ko'ldagi baliqlar o'rtasidagi pastdan bog'lanishlar". Ekologiya. 99 (9): 2025–2036. doi:10.1002 / ecy.2414. PMID 29884987.
- ^ Linam, Kristofer Filipp; Llop, Markos; Molman, nasroniy; Helaouet, Per; Bayliss-Braun, Jorjiya shtati Anne; Stenset, Nils S (fevral 2017). "Shimoliy dengizdagi trofik va atrof-muhit nazorati". Milliy fanlar akademiyasi materiallari. 114 (8): 1952–1957. doi:10.1073 / pnas.1621037114. PMC 5338359. PMID 28167770.
- ^ "Oldlist". Rokki tog 'daraxtining halqasini tadqiq qilish. Olingan 8 yanvar, 2013.
- ^ Vetsel, Robert (2001). Limnologiya: ko'l va daryo ekotizimlari. San-Diego: Akademik matbuot. ISBN 9780127447605.
- ^ Bar-On, YM, Fillips, R. va Milo, R. (2018) "Yerdagi biomassaning tarqalishi". Milliy fanlar akademiyasi materiallari, 115(25): 6506–6511. doi:10.1073 / pnas.1711842115.
- ^ Spellman, Frank R. (2008). Suv haqidagi fan: tushuncha va qo'llanmalar. CRC Press. p. 167. ISBN 978-1-4200-5544-3.
- ^ Odum, E. P.; Barrett, G. V. (2005). Ekologiya asoslari (5-nashr). Brooks / Cole, Cengage Learning-ning bir qismi. ISBN 978-0-534-42066-6. Arxivlandi asl nusxasi 2011-08-20.
- ^ Vang, X.; Morrison, V.; Singx, A .; Vayss, H. (2009). "Ekotizimlarda teskari biomassa piramidalari va qochqinlarni modellashtirish" (PDF). Ekologik modellashtirish. 220 (11): 1376–1382. doi:10.1016 / j.ecolmodel.2009.03.005. Arxivlandi asl nusxasi (PDF) 2011-10-07 kunlari.
- ^ Field, KB, Behrenfeld, MJ, Randerson, J.T. va Falkovskiy, P. (1998) "Biosferani birlamchi ishlab chiqarish: quruqlik va okean komponentlarini birlashtirish". Ilm-fan, 281(5374): 237–240. doi:10.1126 / science.281.5374.237.
- ^ Maureaud, A., Gassel, D., Koller, M., Palomares, ML, Du Pontavits, H., Pauli, D. va Cheung, VW. (2017) "Dengiz oziq-ovqat tarmoqlarining trofik faoliyatidagi global o'zgarish". PLOS ONE, 12(8): e0182826. doi:10.1371 / journal.pone.0182826
- ^ "Okean kislotasini dengiz turlari va ekotizimlariga ta'siri". Hisobot. Okeaniya. Olingan 13 oktyabr 2013.
- ^ "Shimoliy Muz okeanining kislotalanishini kompleks o'rganish". O'qish. CICERO. Arxivlandi asl nusxasi 2013 yil 10-dekabrda. Olingan 14 noyabr 2013.
- ^ Lischka, S .; Büdenbender J .; Boxhammer T.; Riebesell U. (2011 yil 15 aprel). "Okeanning kislotaliligi va yuqori haroratning qutbli qobiqli limacina helicina pteropodining erta o'spirinlariga ta'siri: o'lim, qobiq tanazzuli va qobiq o'sishi" (PDF). Hisobot. Biogeoscience. 919-932 betlar. Olingan 14 noyabr 2013.
- ^ "Antarktida dengiz yovvoyi hayoti tahdid ostida, tadqiqot topilmalari". BBC tabiati. Olingan 13 oktyabr 2013.
- ^ Ullah, H., Nagelkerken, I., Goldenberg, S.U. va Fordham, D.A. (2018) "Iqlim o'zgarishi o'zgargan trofik oqimlar va siyanobakterial ko'payish orqali dengiz oziq-ovqat mahsulotlarining veb-kollapsini qo'zg'atishi mumkin". PLoS biologiyasi, 16(1): e2003446. doi:10.1371 / journal.pbio.2003446
- ^ a b Iqlim o'zgarishi dengiz oziq-ovqat tarmoqlarida qulab tushadi ScienceDaily. 9 yanvar 2018 yil.
- ^ IUCN (2018) IUCN xavf ostida bo'lgan turlarining Qizil ro'yxati: 2018-1 versiyasi