Kelp o'rmoni - Kelp forest
| Dengiz yashash joylari |
|---|
 Kelp o'rmoni |
Kelp o'rmonlari ning zichligi yuqori bo'lgan suv osti zonalari kelp, bu dunyodagi qirg'oqlarning katta qismini egallaydi. Ular eng samarali va dinamik biri sifatida tan olingan ekotizimlar Yerda.[1] Ankrajli laminariyaning kichik joylari deyiladi suv o'tlari yotoqlari.
– Charlz Darvin, 1834 yil 1-iyun, Tierra del Fuego, Chili[2]
Kelp o'rmonlari butun dunyo bo'ylab uchraydi mo''tadil va qutbli qirg'oq okeanlari.[1] 2007 yilda suv o'tlari o'rmonlari ham topilgan tropik yaqin suvlar Ekvador.[3]
Jismoniy ravishda jigarrang tomonidan hosil qilingan makroalglar, suv o'tlari o'rmonlari dengiz organizmlari uchun noyob yashash muhitini yaratadi[4] va ko'plab ekologik jarayonlarni tushunish uchun manba hisoblanadi. So'nggi asrda ular keng qamrovli tadqiqotlarning markaziga aylandilar, xususan trofik ekologiya va ushbu noyob ekotizimdan tashqari muhim g'oyalarni qo'zg'atishda davom eting. Masalan, suv o'tlari o'rmonlari qirg'oqqa ta'sir qilishi mumkin okeanografik naqshlar[5] va ko'pchilikni ta'minlaydi ekotizim xizmatlari.[6]
Biroq, odamlarning ta'siri ko'pincha suv o'tlariga yordam berdi o'rmon tanazzuli. Ta'sirlari ayniqsa tashvishlantiradi ortiqcha baliq ovlash bo'shatishi mumkin bo'lgan yaqin atrofdagi ekotizimlar o'txo'rlar ularning normal aholini tartibga solishidan kelib chiqadi va o'tlab ketish suv o'tlari va boshqa suv o'tlari.[7] Bu tez o'tishga olib kelishi mumkin bepusht manzaralar bu erda nisbatan oz sonli tur saqlanib qoladi.[8][9] Allaqachon tufayli birlashgan ta'siri ortiqcha baliq ovlash va Iqlim o'zgarishi, kelp o'rmonlari, ayniqsa, ko'pgina zaif joylarda yo'qolib ketgan, masalan Tasmaniya sharqiy qirg'oq va Shimoliy Kaliforniya.[10][11] Amalga oshirish dengiz muhofaza qilinadigan hududlari Bunday muammolarni hal qilish uchun foydali bo'lgan boshqaruv strategiyasidir, chunki u baliq ovlash ta'sirini cheklashi va ekotizimni boshqa ekologik stress omillarining qo'shimcha ta'siridan himoya qilishi mumkin.
Kelp
Atama kelp ga tegishli dengiz yosunlari ga tegishli buyurtma Laminariales (filum: Heterokontofit ). Taqsonomik jihatdan xilma-xil tartib deb qaralmasa ham, suv o'tlari tarkibiy va funktsional jihatdan juda xilma-xildir.[6] Eng taniqli turlar - bu ulkan suv o'tlari (Makrokistis kabi ko'plab boshqa nasllarga ega bo'lishiga qaramay Laminariya, Ekkloniya, Lessoniya, Alariya va Eyzenia tasvirlangan.
Dengiz hayotining keng doirasi suv o'tlari o'rmonlarini himoya qilish yoki oziq-ovqat, shu jumladan baliqlardan foydalanadi. Shimoliy Tinch okeanining kelp o'rmonlarida, ayniqsa tosh baliqlar va ko'p umurtqasizlar, kabi amfipodlar, mayda qisqichbaqa, dengiz salyangozlari, tukli qurtlar va mo'rt yulduzlar. Ko'plab dengiz sutemizuvchilar va qushlar, shu jumladan muhrlar, dengiz sherlari, kitlar, dengiz samurlari, gullalar, terns, qorli egretlar, katta ko'k bug'doylar va kormorantlar, shuningdek ba'zi qirg'oq qushlari.[12]
Tez-tez ko'rib chiqiladi ekotizim muhandisi, kelp suv o'tlari jamoalari uchun jismoniy substrat va yashash muhitini ta'minlaydi.[13] Yosunlarda (qirollik) Protista ), individual organizm tanasi a sifatida tanilgan talus o'simlik sifatida emas (shohlik) Plantae ). Lalp talusining morfologik tuzilishini uchta asosiy tarkibiy birlik belgilaydi:[8]
- The ushlanib turish; to'xtamoq Tallni dengiz tubiga bog'lab turadigan ildizga o'xshash massa, ammo haqiqiy ildizlardan farqli o'laroq u talumning qolgan qismiga ozuqa moddalarini singdirish va etkazib berish uchun javobgar emas.
- The stipe O'simliklar poyasiga o'xshaydi, mahkamlangandan vertikal ravishda uzayadi va boshqa morfologik xususiyatlarni qo'llab-quvvatlaydi.
- The jabhalar stipdan, ba'zan uning butun uzunligi bo'ylab cho'zilgan barg yoki pichoqqa o'xshash birikmalar bo'lib, ular ozuqa moddalarini qabul qilish va fotosintez qilish faolligi joylari.
Bundan tashqari, ko'plab suv o'tlari turlari mavjud pnevmatotsistalar, yoki odatda stip yaqinidagi jabhalar tagida joylashgan gaz bilan to'ldirilgan siydik pufagi. Ushbu tuzilmalar suv sathida tik holatini saqlab qolish uchun suv o'tlari uchun zarur bo'lgan suzishni ta'minlaydi.
Laminariyaning omon qolishi uchun zarur bo'lgan atrof-muhit omillari orasida qattiq substrat (odatda tosh yoki qum), yuqori oziq moddalar (masalan, azot, fosfor) va yorug'lik (minimal yillik nurlanish dozasi> 50 E m−2[14]). Ayniqsa, samarali kelp o'rmonlari muhim okeanografik joylar bilan bog'liq ko'tarilish, ozuqa moddalariga boy salqin suvni chuqurlikdan okeanga etkazib beradigan jarayon aralash sirt qatlami.[14] Suv oqimi va turbulentlik suv ustunidagi suv o'tlari bo'ylab ozuqa moddalarining assimilyatsiyasini osonlashtiradi.[15] Suvning tiniqligi etarli yorug'lik uzatilishi mumkin bo'lgan chuqurlikka ta'sir qiladi. Ideal sharoitda ulkan kelp (Makrokistis spp.) kuniga vertikal ravishda 30-60 sm gacha o'sishi mumkin. Kabi ba'zi turlari Nereokistis, bor bir yillik kabi boshqalar esa Eyzenia bor ko'p yillik o'simliklar, 20 yildan ortiq yashaydi.[16] Ko'p yillik kelp o'rmonlarida maksimal o'sish sur'atlari ko'tarilish oylarida (odatda bahor va yoz oylarida) ro'y beradi va o'liklarning orqa qismi ozuqa moddalarining qisqarishiga, fotoperiodlarning qisqarishiga va bo'ron chastotasining ko'payishiga mos keladi.[8]
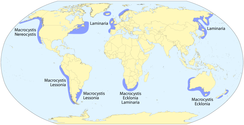
Kelps birinchi navbatda bog'liqdir mo''tadil va Arktika butun dunyo bo'ylab suvlar. Ko'proq hukmron avlodlardan, Laminariya asosan ikkala tomoni bilan bog’liq Atlantika okeani va qirg'oqlari Xitoy va Yaponiya; Ekkloniya topilgan Avstraliya, Yangi Zelandiya va Janubiy Afrika; va Makrokistis shimoliy-sharqiy va janubi-sharqiy bo'ylab sodir bo'ladi tinch okeani, Janubiy okean arxipelaglar va Avstraliya, Yangi Zelandiya va Janubiy Afrika atrofidagi yamoqlarda.[8] Kelplarning eng xilma-xilligi (> 20 tur) bo'lgan mintaqa Tinch okeanining shimoli-sharqidir, shimoldan San-Fransisko, Kaliforniya, uchun Aleut orollari, Alyaska.
Tropik er usti suvlarida kelp o'rmonlari noma'lum bo'lishiga qaramay, bir nechta turlari Laminariya faqat tropik chuqur suvlarda paydo bo'lishi ma'lum bo'lgan.[17][18] Tropikadan kelpning bu umumiy yo'qligi, asosan, iliqlik bilan bog'liq bo'lgan ozuqaviy darajalarning etarli emasligi, oligotrofik suvlar.[8] Yaqinda o'tkazilgan bir tadqiqot, suv o'tlari uchun zarur bo'lgan fizik parametrlarni o'rtacha okeanografik sharoitlar bilan qoplagan holda butun dunyo bo'ylab tropik mintaqalarda 200 m chuqurlikgacha er osti suv o'tlari mavjudligini taxmin qiladigan modelni ishlab chiqdi. Ichida joylashgan nuqta uchun Galapagos orollari, mahalliy model nozik ma'lumotlar bilan yaxshilandi va sinovdan o'tkazildi; tadqiqot guruhi namuna olingan sakkizta uchastkasida gullab-yashnayotgan kelp o'rmonlarini topdi, ularning barchasi model tomonidan taxmin qilingan va shu bilan ularning yondashuvlarini tasdiqladi. Bu shuni ko'rsatadiki, ularning global modeli haqiqatan ham aniq bo'lishi mumkin va agar shunday bo'lsa, kelp o'rmonlari butun dunyo bo'ylab tropik er osti suvlarida samarali bo'lishi mumkin.[3] Ushbu hissaning ahamiyati ilmiy jamoatchilikda tezda tan olindi va suv o'tlari o'rmonlarini tadqiq qilishning mutlaqo yangi traektoriyasini qo'zg'atdi, xususan, iqlim o'zgarishidan fazoviy boshpana olish imkoniyatini ta'kidlab, butun dunyo bo'ylab kelplarning evolyutsion naqshlariga izohlar berdi.[19]
Ekotizim arxitekturasi




Suv o'tlari ekotizimining arxitekturasi uning jismoniy tuzilishiga asoslanadi, bu esa uning jamoaviy tuzilishini belgilaydigan bog'langan turlarga ta'sir qiladi. Tarkibiy jihatdan ekotizimga uchta suv o'tlari gildiyasi va boshqa suv o'tlari egallagan ikkita gildiya kiradi:[8]
- Kanopi suv o'tlari eng katta turlarni o'z ichiga oladi va ko'pincha okean sathiga cho'zilgan suzuvchi soyabonlarni tashkil qiladi (masalan, Makrokistis va Alariya).
- Cho'chqalar, odatda dengiz tubidan bir necha metr balandlikda joylashgan va zich birikmalarda o'sishi mumkin (masalan, Eyzenia va Ekkloniya).
- Dajjol kelplari dengiz tubiga yaqin va bo'ylab yotadi (masalan, Laminariya).
- The bentik yig'ilish boshqa alg turlaridan (masalan, filamentli va folioz funktsional guruhlar, bo'g'inli korallinlar) va okean tubi bo'ylab o'tirgan organizmlardan iborat.
- To'g'ridan-to'g'ri va ko'pincha geologik substratni qamrab olgan korallin yosunlarini yopish.
Ko'pgina suv o'tlari turlari ko'pincha o'rmon ichida yashaydi; ostidagi soyabon atamasi nafaqa va sajda suvi o'tlarini anglatadi. Masalan, a Makrokistis soyabon dengiz sathidan okean sathiga qarab bir necha metrga cho'zilishi mumkin Eyzenia va Pterigofora faqat bir necha metr yuqoriga ko'tariladi. Ushbu suv o'tlari ostida folioz qizil suv o'tlarining bentik birikmasi paydo bo'lishi mumkin. Yuqori vertikal infratuzilma quyoshli soyabon mintaqasi, qisman soyali o'rtasi va qorong'i dengiz tubi bilan quruqlikdagi o'rmonda kuzatilgan mikroelementlar tizimini tashkil qiladi.[8] Har bir gildiyada yashash muhitiga bog'liqlik darajasi turlicha bo'lgan birlashtirilgan organizmlar mavjud va bu organizmlarning to'planishi kelp morfologiyalari bilan farq qilishi mumkin.[20][21][22] Masalan, Kaliforniyada, Macrocystis pyrifera o'rmonlar, yalang'och Melibe leonina va skelet qisqichbaqasi Caprella californica sirt soyabonlari bilan chambarchas bog'liq; kelp perch Brachyistius frenatus, tosh baliqlar Sebastes spp., va boshqa ko'plab baliqlar shartnoma ostidan topilgan; mo'rt yulduzlar va salla salyangozlari Tegula spp. kelp tutqich bilan chambarchas bog'langan, turli xil o'txo'rlar, masalan dengiz kirpi va abalone esa, prostata soyaboni ostida yashaydi; ko'plab dengiz yulduzlari, gidroidlar va bentik bentik birikmalar orasida baliqlar yashaydi; yakka marjonlar, har xil gastropodlar va echinodermalar korallin suv o'tlari ustida yashang.[20] Bunga qo'chimcha, pelagik baliqlar va dengiz sutemizuvchilar suv o'tlari bilan erkin bog'langan bo'lib, odatda ular yashovchi organizmlarni oziqlantirish uchun tashrif buyurganlarida qirralarning yaqinida o'zaro ta'sir o'tkazadilar.
Trofik ekologiya



Suv o'tlari ekologiyasidagi klassik tadqiqotlar asosan trofik o'zaro ta'sirga (organizmlar va ularning o'zaro aloqalariga) qaratilgan oziq-ovqat tarmoqlari ), ayniqsa tushunish va yuqoridan pastga trofik jarayonlar. Pastdan yuqoriga ko'tarilish jarayonlari odatda birlamchi ishlab chiqaruvchilar o'sishi uchun zarur bo'lgan abiotik sharoitlar, masalan, yorug'lik va ozuqa moddalarining mavjudligi va keyinchalik energiyani yuqori trofik darajada iste'molchilarga uzatishi bilan bog'liq. Masalan, kelpning paydo bo'lishi ko'pincha atrof muhitni ozuqa moddalarining g'ayrioddiy yuqori konsentratsiyasini ta'minlaydigan okeanografik ko'tarilish zonalari bilan o'zaro bog'liq.[23][24] Bu suv o'tlari o'sishi va keyinchalik o'txo'r hayvonlarni qo'llab-quvvatlashi mumkin, bu esa iste'molchilarni yuqori darajada qo'llab-quvvatlaydi trofik sathlar.[25] Aksincha, yuqoridan pastga tushadigan jarayonlarda yirtqichlar iste'mol qilish orqali trofik darajadagi turlarning biomassasini cheklaydi. Yirtqich hayvon bo'lmagan taqdirda, ushbu quyi darajadagi turlar rivojlanadi, chunki ularning energetik ehtiyojlarini ta'minlaydigan resurslar cheklanmaydi. Alyaska kelp o'rmonlaridan yaxshi o'rganilgan misolda,[26] dengiz samurlari (Enhidra lutris) o'txo'rlarning populyatsiyasini nazorat qilish dengiz kirpi yirtqichlik orqali. Dengiz quyruqlari ekotizimdan chiqarilganda (masalan, odam ekspluatatsiyasi bilan), urchin populyatsiyasi yirtqich nazoratdan ozod qilinadi va keskin o'sadi. Bu mahalliy suv o'tlari o'simliklariga o't o'simliklari bosimining oshishiga olib keladi. Laminariya buzilishining o'zi jismoniy ekotizim tuzilishini yo'qotishiga va keyinchalik bu yashash joyi bilan bog'liq boshqa turlarning yo'qolishiga olib keladi. Alaskan suv o'tlari ekotizimlarida dengiz samurlari hisoblanadi asosiy tosh turlari bunga vositachilik qiladi trofik kaskad. Kaliforniyaning janubiy qismida suv o'tlari o'rmonlari dengiz quyruqlarisiz davom etadi va o'txo'r urchinlarni boshqarish vositasida yirtqichlar to'plami, jumladan lobsterlar va yirik baliqlar, masalan, Kaliforniya qo'y boshi. Ushbu tizimdagi bitta yirtqich turni olib tashlashning ta'siri Alyaskadan farq qiladi, chunki ortiqcha narsa trofik darajada mavjud va boshqa yirtqich turlar kirpiklarni boshqarishda davom etishi mumkin.[21] Shu bilan birga, ko'plab yirtqichlarni yo'q qilish urchinlarni yirtqichlarning bosimidan samarali ravishda xalos qilishi va tizimga suv o'tlari degradatsiyasi tomon traektoriyalarni kuzatishi mumkin.[27] Shunga o'xshash misollar mavjud Yangi Shotlandiya,[28] Janubiy Afrika,[29] Avstraliya[30] va Chili.[31] Suv o'tlari ekotizimida tepadan pastga va pastdan yuqoriga qarab nisbiy ahamiyati va trofik o'zaro ta'sirlarning kuchli tomonlari muhim ilmiy tadqiqot mavzusi bo'lib qolmoqda.[32][33][34]
Makroalgaldan (ya'ni kelp o'rmonidan) dengiz kirpi (yoki "urchin barrens") hukmron bo'lgan denudlangan landshaftlarga o'tish keng tarqalgan hodisa,[6][35][36][37] ko'pincha yuqorida tavsiflangan kabi trofik kaskadlardan kelib chiqadi; ikki faza ekotizimning muqobil barqaror holati sifatida qaraladi.[38][39] Kelp o'rmonlarining bepusht holatidan tiklanishi, urchin kasalligi yoki issiqlik sharoitida katta siljishlar kabi dramatik bezovtaliklardan so'ng hujjatlashtirilgan.[27][40][41] Buzilishning oraliq holatlaridan tiklanish kamroq taxmin qilinadi va har bir holatda abiotik omillar va biotik o'zaro ta'sirlarning kombinatsiyasiga bog'liq.
Odatda urchinlar dominant o'txo'rlar bo'lishiga qaramay, o'zaro ta'sir kuchiga ega bo'lgan boshqalar kiradi dengiz yulduzlari, izopodlar, kelp Qisqichbaqa va o'txo'r baliqlar.[8][32] Ko'pgina hollarda, bu organizmlar substratdan ajralgan suv o'tlari va okean tubining yaqinida suzib yurib, ozuqa berish uchun butun tallini qidirish uchun sarflamaydilar. Dreyf suv o'tlari etarli bo'lganda, o'txo'r o'tlovchilar biriktirilgan o'simliklarga bosim o'tkazmaydi; drift dotatsiyalari mavjud bo'lmaganda, o'tlovchilar ekotizimning fizikaviy tuzilishiga bevosita ta'sir qiladi.[42][43] Kaliforniyadagi Janubiy Kaliforniyada o'tkazilgan ko'plab tadqiqotlar shuni ko'rsatdiki, suv o'tlari suv o'tqazishining mavjudligi dengiz urchinlarining ozuqaviy xatti-harakatlariga ta'sir qiladi.[44][45] Drift kelp va kelpdan olingan zarrachalar, shuningdek, qumli plyajlar va toshloq intertidal kabi qo'shni yashash joylarini subsidiyalashda muhim ahamiyatga ega.[46][47][48]
Yamoqlarning dinamikasi
Kelp o'rmonlarini tadqiq qilishning yana bir muhim yo'nalishi suv o'tlari yamoqlarining fazoviy-vaqtinchalik naqshlarini tushunishga qaratilgan. Bunday dinamikalar nafaqat jismoniy landshaftga ta'sir qilmaydi, balki ular kelp bilan boshpana yoki em-xashak faoliyati bilan bog'liq turlarga ham ta'sir qiladi.[20][25] Keng miqyosdagi atrof-muhit buzilishi mexanizmlar va ekotizim haqida muhim tushunchalarni taqdim etdi chidamlilik. Ekologik buzilishlarga quyidagilar kiradi:
- O'tkir va surunkali ifloslanish hodisa Kaliforniyaning janubiy kelp o'rmonlariga ta'sir ko'rsatdi, ammo ta'sir intensivligi ifloslantiruvchi moddalarning tabiati va ta'sir qilish muddatiga bog'liq.[49][50][51][52][53] Ifloslanish tarkibiga cho'kindi yotqizish va evrofikatsiya kanalizatsiya, sanoat mahsuloti va shunga o'xshash ifloslantiruvchi moddalardan Tenglikni va og'ir metallar (masalan, mis, rux), oqimi organofosfatlar portlar va marinalarda ishlatiladigan qishloq xo'jaligi hududlaridan, ifloslanishga qarshi kimyoviy vositalar (masalan, TBT va kreozot ) va shunga o'xshash quruqlik patogenlari najas koliform bakteriyalari.
- Katastrofik bo'ronlar to'lqin faolligi orqali sirt suv o'tlari soyabonlarini olib tashlashi mumkin, lekin odatda pastki suv o'tlarini buzilmasdan qoldiradi; ozgina fazoviy boshpana mavjud bo'lganda, ular kirpiklarni olib tashlashlari mumkin.[38][43] Aralashgan soyabonlarni tozalash dengiz manzarasi mozaikasini yaratadi, u erda quyosh nuri suv o'tlari o'rmoniga chuqurroq kirib boradi va odatda ostki qismida yorug'lik bilan cheklangan turlar gullab-yashnashi mumkin. Xuddi shu tarzda, suv o'tlari tutqunlaridan tozalangan substrat boshqa o'tiradigan turlar uchun o'zlarini o'rnatishi va dengiz sathini egallashi uchun joy ajratib berishi mumkin, ba'zida balog'at yosh suv o'tlari bilan to'g'ridan-to'g'ri raqobatlashadi va hatto ularning joylashishini to'xtatadi.[54]
- El-Nino-Janubiy tebranish (ENSO) hodisalari okeanografik termoklinlarning depressiyasini, ozuqa moddalarining kamayishini va bo'ron shaklidagi o'zgarishlarni o'z ichiga oladi.[38][55] Iliq suv tufayli stress va ozuqa moddalarining etishmasligi kelpning bo'ron zararlanishiga va o'txo'r yaylovlarga ta'sirchanligini oshirishi mumkin, ba'zida esa urchinlar ustun bo'lgan landshaftlarga bosqichma-bosqich o'tishi ham mumkin.[41][44][56] Umuman olganda, okeanografik sharoitlar (ya'ni suv harorati, oqimlari) suv o'tlari va uning raqobatchilarini ishga qabul qilish muvaffaqiyatiga ta'sir qiladi, bu keyingi turlarning o'zaro ta'siriga va suv o'tlari dinamikasiga aniq ta'sir qiladi.[38][57]
- O'simliklar populyatsiyasini tabiiy ravishda tartibga soluvchi yuqori trofik darajalarni ortiqcha ovlash ham suv o'tlari o'rmonlarida muhim stress omil sifatida tan olingan.[7][34][58] Oldingi bobda aytib o'tilganidek, trofik kaskadlarning haydovchilari va natijalari suv o'tlari o'rmonlarining fazoviy-vaqtinchalik shakllarini tushunish uchun muhimdir.[26][27][32]
Bunday buzilishlardan oldin, paytida va undan keyin suv o'tlari o'rmonlarini ekologik monitoringidan tashqari, olimlar eksperimental manipulyatsiya yordamida suv o'tlari o'rmonlari dinamikasining nozik tomonlarini chalg'itishga harakat qilishadi. Kichikroq kosmik-vaqtinchalik tarozilar ustida ishlash orqali ular operatsion mexanizmlarni kashf qilish uchun o'ziga xos biotik va abiotik omillarning mavjudligini yoki yo'qligini nazorat qilishlari mumkin. Masalan, Avstraliyaning janubiy qismida suv o'tlari soyabon turlari bilan manipulyatsiya nisbiy miqdori ekanligini ko'rsatdi Ekkloniya radiati soyabonda pastki turlarning birikmalarini taxmin qilish uchun foydalanish mumkin; Binobarin, ning nisbati E. radiata atrof muhitda uchraydigan boshqa turlarning ko'rsatkichi sifatida ishlatilishi mumkin.[59]
Insondan foydalanish
Kelp o'rmonlari insoniyat uchun ming yillar davomida muhim ahamiyatga ega.[60] Darhaqiqat, hozirgi kunda ko'pchilik Amerikaning birinchi mustamlakasi so'nggi muzlik davrida Tinch okeanining suv o'tlari o'rmonlaridan keyin baliq ovlash jamoalari tufayli yuzaga kelgan deb taxmin qilishmoqda. Bir nazariya, Osiyodagi shimoli-sharqdan Amerikaning Tinch okeanining qirg'og'igacha cho'zilgan suv o'tlari o'rmonlari qadimgi qayiqchilarga ko'p foyda keltirishi mumkin edi. Suv o'tlari o'rmonlari ko'plab oziq-ovqat imkoniyatlarini taqdim etishi bilan bir qatorda, qo'pol suvdan bufer turi sifatida ham ish yuritishi mumkin edi. Ushbu afzalliklardan tashqari, tadqiqotchilar kelp o'rmonlari "qayiq magistrali" ning bir turi sifatida harakat qilib, erta qayiqchilarning suzishiga yordam bergan bo'lishi mumkin deb hisoblashadi. Nazariyotchilar, shuningdek, suv o'tlari o'rmonlari bu qadimiy kolonistlarga barqaror hayot tarzini ta'minlab, yangi ekotizimlarga moslashish va minglab chaqirim yo'l bosib o'tishda ham yangi omon qolish usullarini ishlab chiqishlariga yo'l qo'ymaslik orqali yordam bergan bo'lar edi.[61] Zamonaviy iqtisodiyot asoslanadi baliqchilik kabi kelp bilan bog'liq turlarning katta dengiz qisqichbagasi va tosh baliqlar. Odamlar, shuningdek, akvakultura turlarini boqish uchun to'g'ridan-to'g'ri kelpni yig'ib olishlari mumkin oyoq osti va birikmani ajratib olish uchun algin kislotasi, tish pastasi va antasidlar kabi mahsulotlarda ishlatiladi.[62][63] Kelp o'rmonlari SCUBA sho'ng'in va baydarka kabi dam olish tadbirlari uchun qadrlanadi; ushbu sport turlarini qo'llab-quvvatlovchi sohalar ekotizim bilan bog'liq bir foydani anglatadi va ushbu faoliyatdan olingan lazzatlanish boshqasini anglatadi. Bularning barchasi misollar ekotizim xizmatlari kelp o'rmonlari tomonidan maxsus taqdim etilgan.
Tahdidlar va boshqarish

Suv o'tlari o'rmonlarining murakkabligini hisobga olgan holda - ularning o'zgaruvchan tuzilishi, geografiyasi va o'zaro ta'siri - ular atrof-muhit menejerlari uchun katta qiyinchiliklar tug'diradi. Kelajakka yaxshi o'rganilgan tendentsiyalarni ekstrapolyatsiya qilish juda qiyin, chunki ekotizimdagi o'zaro ta'sirlar o'zgaruvchan sharoitlarda o'zgarib turadi, ekotizimdagi barcha munosabatlar tushunilmaydi va o'tish uchun chiziqli bo'lmagan chegaralar hali tan olinmagan.[64] Kelp o'rmonlariga nisbatan, dengizning ifloslanishi va suv sifati, kelp yig'ish va baliq ovlash, invaziv turlar,[6] va Iqlim o'zgarishi.[65] Suv o'tlari muhofazasi uchun eng dolzarb tahdid qirg'oq ekotizimlarining ortiqcha ovlanishi bo'lishi mumkin, bu esa yuqori trofik sathlarni olib tashlash bilan ularning urchin bepushtlarini depauperatsiya qilishga o'tishini osonlashtiradi.[7] Ta'minlash biologik xilma-xillik funktsional kompensatsiya va begona turlarning ishg'ol etilishining pasayishi kabi mexanizmlar orqali ekotizimlarni va ularning xizmatlarini umuman barqarorlashtirish usuli sifatida tan olingan.[66][67][68][69]
Ko'p joylarda menejerlar suv o'tlari hosilini tartibga solishni tanladilar[24][70] va / yoki baliq ovlari bilan suv o'tlari o'rmon turlarini olish.[6][58] Garchi ular bir ma'noda samarali bo'lishi mumkin bo'lsa-da, ular ekotizimni to'liq himoya qilishi shart emas. Dengiz muhofazalangan hududlari (MPAs) nafaqat yig'ish uchun maqsadli turlarni, balki ular atrofidagi o'zaro ta'sirlarni va umuman mahalliy muhitni qamrab oladigan noyob echimni taklif etadi.[71][72] MPA-larning baliqchilikka to'g'ridan-to'g'ri foydalari (masalan, parchalanish effektlari) butun dunyoda yaxshi hujjatlashtirilgan.[7][73][74][75] Markaziy Kaliforniyadagi dengiz osti baliqlari va baliqlar kabi turlar orasida bir nechta holatlar uchun bilvosita foyda ko'rsatildi.[76][77] Eng muhimi, MPA mavjud suv o'tlari ekotizimlarini himoya qilishda samarali bo'lishi va ta'sirlanganlarni qayta tiklashga imkon berishi mumkin.[38][78][79]
Adabiyotlar
- ^ a b Mann, K.H. 1973. Dengiz o'simliklari: ularning unumdorligi va o'sish strategiyasi. Ilmiy 182: 975-981.
- ^ Darvin, C. 1909 yil. Beaglening sayohati. Garvard klassiklari jildi 29. Nyu-York, AQSh: P.F. Collier & Son kompaniyasi.
- ^ a b Grem, M.H., B.P. Kinlan, L.D. Druehl, L.E. Garske va S. Banks. 2007. Tropik dengiz xilma-xilligi va unumdorligining potentsial qaynoq nuqtalari sifatida chuqur suvli qayiq refugiyasi. Milliy Fanlar Akademiyasi materiallari 104: 16576-16580.
- ^ Christie, H., Jorgensen, N.M., Norderhaug, KM, Waage-Nielsen, E., 2003. Norvegiya qirg'oqlari bo'ylab kelp (Laminaria hyperborea) bilan bog'liq hayvonot dunyosining turlari tarqalishi va yashash joylarini ekspluatatsiya qilish. Buyuk Britaniyaning Dengiz biologik assotsiatsiyasi jurnali 83, 687-699.
- ^ Jekson, G.A. va C.D. G'olib. 1983. Suv o'tlari o'rmonining qirg'oq oqimlariga ta'siri. Kontinental tokcha hisoboti 2: 75-80.
- ^ a b v d e Stenek, R.S., M.H. Grem, BJ Bork, D. Korbet, J.M.Erlandson, J.A. Estlar va M.J. Tegner. 2002. Kelp o'rmon ekotizimlari: bioxilma-xillik, barqarorlik, barqarorlik va kelajak. Atrof muhitni muhofaza qilish 29: 436-459.
- ^ a b v d Sala, E., CF. Bourdouresque va M. Harmelin-Vivien. 1998. Baliq ovlash, trofik kaskadlar va suv o'tlari birikmalarining tuzilishi: eski, ammo tekshirilmagan paradigmani baholash. Oikos 82: 425-439.
- ^ a b v d e f g h Deyton, P.K. 1985a. Suv o'tlari jamoalari ekologiyasi. Ekologiya va sistematikaning yillik sharhi 16: 215-245.
- ^ Norderxaug, KM, Kristi, H., 2009. Atlantika okeanida dengiz kirpini boqish va suv o'tlari o'simliklari. Dengiz biologiyasi tadqiqotlari 5, 515-528
- ^ Morton, Odam; Kordell, Marni; Fanner, Devid; To'p, Endi; Evershed, Nik. "O'lik dengiz: Tasmaniyaning suv osti o'rmonlari bizning hayotimizda yo'q bo'lib ketmoqda". Guardian. Olingan 2020-10-22.
- ^ Shtaynbauer, Jeyms. "Kelp o'rmonini qaytarish uchun nima qilish kerak? -" Bay Nature "jurnali". Bay tabiati. Olingan 2020-10-22.
- ^ Kelp o'rmonlari turli xil umurtqasizlar, baliqlar, dengiz sutemizuvchilari va qushlarning yashash muhitini ta'minlaydi NOAA. 2013 yil 11 yanvarda yangilangan. 2014 yil 15 yanvarda olingan.
- ^ Jons, KG, J. H. Lauton va M. Shakak. 1997. Organiklarning fizik ekotizim muhandislari sifatida ijobiy va salbiy ta'siri. Ekologiya 78: 1946-1957.
- ^ a b Druehl, L.D. 1981. Laminarialesning Tinch okeanining shimoliy qismida tarqalishi atrof-muhit ta'siriga qarab. Tizimli evolyutsiya va biologiya bo'yicha Xalqaro Kongress materiallari 2: 248-256.
- ^ Wheeler, W.N. 1980. Chegara qatlam transportining uglerodni ulkan suv o'tlari bilan biriktirishga ta'siri Macrocystis pyrifera. Dengiz biologiyasi 56: 103-110.
- ^ Stenek, R.S. va M.N. Deti. 1994. Alglar ustun bo'lgan jamoalar tuzilishiga funktsional guruh yondashuvi. Oikos 69: 476-498.
- ^ Joly, A.B. va E.C. Oliveira Filho. 1967. Ikki braziliyalik Laminariyalar. Pesquisas da Marinya Instituti 4: 1-7.
- ^ Petrov, J.E., M.V. Suchovejeva va G.V. Avdejev. 1973. Jinsning yangi turlari Laminariya Filippin dengizidan. Nov tizim. Nijj. Tez. 10: 59-61.
- ^ Santelices, B. 2007. Tropik mintaqalarning chuqur suvli yashash joylarida suv o'tlari o'rmonlarining ochilishi. NationalAwan Riak Fanlar Akademiyasi materiallari 104: 19163-19164.
- ^ a b v Foster, M.S. va D.R. Schiel. 1985. Kaliforniyadagi ulkan kelp o'rmonlari ekologiyasi: jamoat profilidir. AQSh baliq va yovvoyi tabiatni muhofaza qilish xizmati to'g'risidagi hisobot 85: 1-152.
- ^ a b Grem, M.X. 2004. Mahalliy o'rmonlarni kesishning Janubiy Kaliforniyadagi yirik suv o'tlari o'rmonlari to'rlarining xilma-xilligi va tuzilishiga ta'siri. Ekotizimlar 7: 341-357.
- ^ Fowler-Walker, MJ, B. M. Gillanders, S.D. Konnell va A.D. Irving. 2005. Mo''tadil Avstraliya bo'ylab soyabon-morfologiya va ostki birikmalar o'rtasidagi bog'liqlik naqshlari. Estuariniya, qirg'oq va tokchali fan 63: 133-141.
- ^ Jekson, G.A. 1977. Oziq moddalar va ulkan suv o'tlari ishlab chiqarish, Macrocystis pyrifera, Kaliforniya janubidan tashqarida. Limnologiya va okeanografiya 22: 979-995.
- ^ a b Deyton, P.K. M.J. Tegner, P.B. Edvards va K.L. Riser. 1999. Suv o'tlari demografiyasining vaqtinchalik va fazoviy o'lchovlari: okeanografik iqlimning roli. Ekologik monografiyalar 69: 219-250.
- ^ a b Karr, M.H. 1994. Makroalgal dinamikasining mo''tadil rif baliqlarini jalb qilishga ta'siri. Ekologiya 75: 1320-1333.
- ^ a b Estes, J.A. va D.O. Duggins. 1995. Alyaskadagi dengiz samurotlari va suv o'tlari o'rmonlari: jamoaviy ekologik paradigmaning umumiyligi va o'zgarishi. Ekologik monografiyalar 65: 75-100.
- ^ a b v Pirs, J.S. va A.H.Hines. 1987. Dengiz kirpilarining ommaviy o'limidan so'ng Kaliforniyaning markaziy kelp o'rmonining kengayishi. Dengiz biologiyasi 51: 83-91.
- ^ Shaybiling, R.E. va A.V. Xenigar. 1997. Dengiz kirpiklarida kasallikning takroriy tarqalishi Strongilotsentrotus droebachiensis Yangi Shotlandiyada: keng ko'lamli meteorik mantiq va okeanografik hodisalar bilan bog'lanish uchun dalillar. Dengiz ekologiyasi taraqqiyoti seriyasi 152: 155-165.
- ^ Velimirov, B., J.G. Maydon, C.L. Griffits va P. Zoutendik. 1977. Benguela ko'tarilish tizimidagi suv o'tlari qatlamlari ekologiyasi. Helgoland dengiz tadqiqotlari 30: 495-518.
- ^ Endryu, N.L. 1993. Mo''tadil Avstraliyadagi riflardagi fazoviy heterojenlik, dengiz kirpiklarini boqish va yashash muhitining tuzilishi. Ekologiya 74: 292-302.
- ^ Deyton, P.K. 1985b. Ba'zi Janubiy Amerika suv o'tlari jamoalarining tuzilishi va tartibga solinishi. Ekologik monografiyalar 55: 447-468.
- ^ a b v Sala, E. va M.H. Grem. 2002. Kelp o'rmonlarida yirtqichlar va yirtqichlarning o'zaro ta'sir kuchini jamoat miqyosida taqsimlash. Milliy Fanlar Akademiyasi materiallari 99: 3678-3683.
- ^ Byrnes, J., JJ Stachowicz, K.M. Xultgren, A.R. Xyuz, S.V. Olyarnik va C.S.Thornber. 2006. Yirtqich hayvonlarning xilma-xilligi o'txo'rlarning xatti-harakatlarini o'zgartirish orqali kelp o'rmonlaridagi trofik kaskadlarni kuchaytiradi. Ekologiya Xatlari 9: 61-71.
- ^ a b Halpern, B.S., K. Kotteni va B.R. Broitman. 2006. Janubiy Kaliforniyadagi suv o'tlari ekotizimlarida tepadan pastga kuchli nazorat. Ilmiy 312: 1230-1232.
- ^ Lourens, JM 1975. Dengiz o'simliklari va dengiz kirpiklari o'rtasidagi munosabatlar to'g'risida. Okeanografiya va dengiz biologiyasi, yillik sharh. 13: 213-286.
- ^ Xyuz, T.P. 1994. Karib dengizi marjon rifining falokatlari, o'zgarishlar siljishi va keng ko'lamli degradatsiyasi. Ilmiy 265: 1547-1551.
- ^ Siversten, K. 2006. Norvegiya qirg'oqlari bo'ylab suv o'tlari yotoqlarining haddan tashqari o'tlab ketishi. Amaliy Fikologiya jurnali 18: 599-610.
- ^ a b v d e Dayton, P.K., MJ Tegner, P.E. Parnell va P.B. Edvards. 1992. Suv o'tlari o'rmonlari jamoasida vaqtinchalik va fazoviy bezovtalik va tiklanish usullari. Ekologik monografiyalar 62: 421-445.
- ^ Pirs, J.S. 2006. Binafsha dengiz kirpiklarining ekologik roli. Ilmiy 314: 940-941.
- ^ Lafferti, K.D. 2004. Lobsterlarni baliq ovlash bilvosita dengiz kirpini epidemiyasini ko'paytiradi. Ekologik dasturlar 14: 1566-1573.
- ^ a b Vaskes, JA, JM Alonso Vega va AH Buschmann. 2006. Chilining shimolidagi suv o'tlari jamoalari tarkibidagi uzoq muddatli o'zgaruvchanlik va 1997-98 yillardagi ENSO. Amaliy Fikologiya jurnali 18: 505-519.
- ^ Koven, R.K. 1983. qo'y qo'ylarining ta'siri (Semisossifus pulcherqizil dengiz urchinidagi yirtqich hayvon (Strongylocentrotus franciscanus) populyatsiyalar: eksperimental tahlil. Ekologiya 58: 249-255.
- ^ a b Ebeling, A.W., D.R. Laur va R.J. Rouli. 1985. Kaliforniya shtatining janubiy kelp o'rmonida kuchli bo'ron buzilishi va jamoat tuzilishini o'zgartirish. Dengiz biologiyasi 84: 287-294.
- ^ a b Deyton, P.K. va M.J. Tegner. 1984. Kaliforniya shtatining janubiy kelp jamoasida katastrofik bo'ronlar, El-Nino va yamoq barqarorligi. Ilmiy 224: 283-285.
- ^ Harrold, C. va DC Reed. 1985. Oziq-ovqat mahsulotlarining mavjudligi, dengiz kirpiklarini boqish va suv o'tlari jamoalarining tuzilishi. Ekologiya 66: 1160-1169.
- ^ Koop, K., RC Nyuell va M.I. Lukas. 1982. biologik parchalanish va suv o'tlari asosida uglerod oqimi (Ekkloniya maxima) qumli plyaj mikrokosmosidagi qoldiqlar. Dengiz ekologiyasining rivojlanishi 7-seriya: 315-326.
- ^ Bustamante, RH, G.M. Filial va S. Eekhout. 1995. Janubiy Afrikada g'ayritabiiy grazer biomassasini saqlash: subtidal suv o'tlari bilan subsidiya. Ekologiya 76: 2314-2329.
- ^ Kaehler, S., E.A. Paxomov, R.M. Kalin va S. Devis. 2006. Suv o'tqazadigan sub-Antarktika tizimidagi kelpdan olingan to'xtatilgan zarrachalarning trofik ahamiyati. Dengiz ekologiyasi taraqqiyoti seriyasi 316: 17-22.
- ^ Grigg, RW va R.S. Kivala. 1970. Chiqarilgan chiqindilarning dengiz hayotiga ba'zi ekologik ta'siri. Kaliforniya Baliq va o'yin bo'limi 56: 145-155.
- ^ Stull, J.K. 1989. Katta dengiz qirg'og'i yaqinidagi cho'kindilardagi ifloslantiruvchi moddalar: tarixi, ta'siri va kelajagi. OCEANS '89 2-nashr: 481-484.
- ^ Shimoliy, VJ, D.E. Jeyms va L.G. Jons. 1993. suv o'tlari yotoqlari tarixi (Makrokistis) Orange va San-Diego shtatlarida, Kaliforniya. Gidrobiologiya 260/261: 277-283.
- ^ Tegner, MJ, P.K. Dayton, PB. Edvards, K.L. Riser, D.B. Chadvik, T.A. Dekan va L. Deysher. 1995. Katta kanalizatsiya to'kilib ketadigan suv o'tlari jamoasiga ta'siri: falokatmi yoki bezovtalikmi? Dengiz atrof-muhit tadqiqotlari 40: 181-224.
- ^ Duradgor, S.R., R.F. Karako, D.F. Kornell, R. Xovart, A.N. Sharpley va V.N. Smit. 1998. Er usti suvlarining fosfor va azot bilan noaniq ifloslanishi. Ekologik dasturlar 8: 559-568.
- ^ Kennelli, S.J. 1987. Avstraliya suv o'tlari jamoasida jismoniy buzilishlar. I. Vaqtinchalik ta'sir. Dengiz ekologiyasi taraqqiyoti seriyasi 40: 145-153.
- ^ McPhaden, MJ 1999. 1997-1998 yillarda El-Ninoning genezisi va evolyutsiyasi. Ilmiy 283: 950-954.
- ^ Edvards, M.S. va G. Ernandes-Karmona. 2005. El Ninodan keyin Shimoliy Tinch okeanining janubiy oralig'idagi chegara yaqinida ulkan suv o'tlarining tiklanishi kechiktirildi. Dengiz biologiyasi 147: 273-279.
- ^ Duggins, DO, J.E. Ekman va A.T. Syuell. 1990. Suv o'tlari atrof-muhit ekologiyasi. II. Suv o'tlarining bentik umurtqasizlarni yollashga ta'siri. Eksperimental dengiz biologiyasi va ekologiyasi jurnali 143: 27-45.
- ^ a b Jekson, JB, M.X. Kirby, W.H. Berger, K.A. Byordal, L.V. Botsford, BJ Bourke, RH Bredberi, R. Kuk, J. Erlandson, J.A. Estlar, T.P. Xyuz, S. Kidvell, KB Lange, X.S. Lenihan, JM Pandolfi, C.H. Peterson, R.S. Stenek, MJ Tegner va RR Uorner. 2002. Tarixiy ortiqcha baliq ovi va yaqinda qirg'oq ekotizimlarining qulashi. Ilmiy 293: 629-638.
- ^ Irving, AD va S.D. Konnell. 2006. Kanopi borligi va tarkibidan pastki tuzilishni taxmin qilish: dengiz yosunlari uchun yig'ilish qoidasi. Ekologiya 148: 491-502.
- ^ Simenstad, Kaliforniya, J.A. Estlar va K.V. Kenyon. 1978. Aleutlar, dengiz samurlari va muqobil barqaror davlat jamoalari. Ilmiy 200: 403-411.
- ^ Pringle Odamlar qayiq bilan dunyoni mustamlaka qildimi?
- ^ Gutierrez, A., T. Korrea, V. Myunoz, A. Santibanez, R. Markos, C. Kaseres va A.X.Buschmann. 2006. Gigant laminariyaning etishtirish Macrocystis pyrifera yangi oziq-ovqat mahsulotlarini ishlab chiqarish uchun janubiy Chilida. Amaliy Fikologiya jurnali 18: 259-267.
- ^ Ortiz, M. va V. Stots. 2007. Abaloneni joriy etishning ekologik va ekologik ijtimoiy modellari Haliotis discus hannai Shimoliy-markaziy Chilining bentik tizimlariga: barqarorlikni baholash. Suvda tabiatni muhofaza qilish: dengiz va chuchuk suv ekotizimlari 17: 89-105.
- ^ Scheffer, M., S. Carpenter, JA. Fuli, C. Folke va B. Valter. 2001. Ekotizimdagi katastrofik siljishlar. Tabiat 413: 591-596.
- ^ Makdonald, Lyusi (2019-02-06). "Tasmaniyaning ulkan kelpidan 95 dona yo'q bo'lib ketdi, olimlar qolganini saqlab qolish uchun poygada". ABC News. Olingan 2020-02-09.
- ^ Frost, T.M., S.R. Duradgor, A.R. Ives va T.K. Kratz. 1995. "Turlarning kompensatsiyasi va ekotizim funktsiyalarida bir-birini to'ldirish". Yilda: C. Jons va J. Lauton, muharrirlar. Turlar va ekotizimlarni bog'lash. Chapman va Xoll, London. 387 pp.
- ^ Tilman, D., KL, Lehman va miloddan avvalgi Bristov. 1998. Turli xillik va barqarorlik munosabatlari: statistik muqarrarlikmi yoki ekologik oqibatmi? Amerikalik tabiatshunos 151: 277-282.
- ^ Stachowicz, JJ, RB Whitlatch va R.W. Usmon. 1999. Turlarning xilma-xilligi va dengiz ekotizimidagi bosqinlarga qarshilik. Ilmiy 286: 1577-1579.
- ^ Elmqvist, T., C. Folke, M. Nystrom, G. Peterson, J. Bengtsson, B. Uolker va J. Norberg. 2003. Javobning xilma-xilligi, ekotizimning o'zgarishi va barqarorligi. Ekologiya va atrof-muhit chegaralari 1: 488-494.
- ^ Stekoll, M.S., L.E. Deysher va M. Xess. 2006. O'rim-yig'im qilinadigan suv o'tlari biomassasini baholashga masofadan zondlash usuli. Amaliy Fikologiya jurnali 18: 323-334.
- ^ Allison, GA, J. Lubchenko va M.H. Karr. 1998. Dengiz zahiralari zarur, ammo dengizni saqlash uchun etarli emas. Ekologik dasturlar 8: S79-S92.
- ^ Airamé, S., JE Dugan, K.D. Lafferti, X. Lesli, D.A. MacArdle va R.R. Warner. 2003. Dengiz zahiralarini loyihalash uchun ekologik mezonlarni qo'llash: Kaliforniya Kanal orollaridan olingan misol. Ekologik dasturlar 13: S170-S184.
- ^ Bohnsack, J.A. 1998. Rif baliqchilikni boshqarish uchun dengiz zahiralarini qo'llash. Avstraliya ekologiya jurnali 23: 298-304.
- ^ Gell, F.R. va C.M. Roberts. 2003. Chegaradan tashqari foyda: dengiz zahiralarining baliq ovlash ta'siri. Ekologiya va evolyutsiya tendentsiyalari 18: 448-455.
- ^ Uillis, TJ, RB Millar va R.K. Babkok. 2003. Mo''tadil mintaqalarda ekspluatatsiya qilingan baliqlarni himoya qilish: yuqori zichlik va baliq ovining biomassasi Pagrus auratus (Sparidae) Yangi Zelandiya shimolidagi dengiz zahiralarida. Amaliy ekologiya jurnali 40: 214-227.
- ^ Paddak, MJ va J.A. Estlar. 2000. Markaziy Kaliforniyaning dengiz zaxiralari va unga qo'shni ekspluatatsiya qilingan hududlarida baliq suvi balig'i populyatsiyasi. Ekologik dasturlar 10: 855-870.
- ^ Rojers-Bennet, L. va J.S. Nok. 2001. Balog'atga etmagan bolalar uchun dengiz qo'riqlanadigan hududlarining bilvosita foydalari. Saqlash biologiyasi 15: 642-647.
- ^ Babkok, RC, S. Kelly, N.T. Qaychi, J.W. Uoker va T.J. Uillis. 1999. Mo''tadil dengiz zaxiralarida jamoat tarkibidagi o'zgarishlar. Dengiz ekologiyasi taraqqiyoti seriyasi 189: 125-134.
- ^ Halpern, B.S. va R.R.Uorner. 2002. Dengiz zahiralari tez va doimiy ta'sirga ega. Ekologiya xatlari 5: 361-366.
Tashqi havolalar
- "Kelp Forest va Rokki subtidal yashash joylari". noaa.gov. Arxivlandi asl nusxasi 2007-03-22.
- "Kelp tomoshasi". tas.gov.au. Tasmaniya, Avstraliya: Birlamchi sanoat, suv va atrof-muhitni muhofaza qilish bo'limi. Arxivlandi asl nusxasi 2004-12-04. Excellent general information on kelp forests, as well as specific information on Tasmanian kelp forests.
- "Monterey Bay Aquarium Kelp Cam". mbayaq.org. Monterey ko'rfazidagi akvarium. Arxivlandi asl nusxasi 1999-11-28 kunlari. Watch a live feed from the kelp forest exhibit.


| Vakolat nazorati |
|---|