Nav1.8 - Nav1.8




Nav1.8 a natriy ionli kanal odamlarda kodlangan subtip SCN10A gen.[5][6][7][8]
Nav1.8 o'z ichiga olgan kanallar tetrodotoksin (TTX) - chidamli kuchlanishli kanallar. Nav1.8 maxsus ravishda ifodalangan dorsal ildiz ganglioni (DRG), mayinsiz, kichik diametrda sezgir neyronlar deb nomlangan C tolalari va ishtirok etadi nosiseptsiya.[9][10] C-tolalari zararli termik yoki mexanik stimullar yordamida faollashishi va shu bilan ularni tashiy olishi mumkin og'riq xabarlar.
Na ning o'ziga xos joylashishivDRG sezgir neyronlaridagi 1.8 uni yangi rivojlanish uchun asosiy terapevtik maqsadga aylantirishi mumkin og'riq qoldiruvchi vositalar[11] va davolash surunkali og'riq.[12]
Funktsiya
Natriy-ionli kuchlanish kanallari (VGSC) ishlab chiqarish va ko'paytirishda juda muhimdir harakat potentsiali. Tetrodotoksin, tarkibida toksin puferfish, ba'zi bir VGSC-larni blokirovka qilishga qodir va shuning uchun har xil subtiplarni ajratish uchun foydalaniladi. Uchta TTXga chidamli VGSC mavjud: Nav1.5, Nav1.8 va Nav1.9. Nav1.8 va Nav1.9 ikkalasi ham ifodalangan nosiseptorlar (zararni sezuvchi neyronlar). Nav1.7, Nav1.8 va Nav1.9 DRGda topilgan va surunkali yallig'lanish og'rig'iga yordam beradi.[13] Nav1.8 - har biri oltita transmembranli mintaqaga ega bo'lgan to'rtta gomologik domendan tashkil topgan a tipidagi kanal subbirligi, ulardan biri kuchlanish sensori.
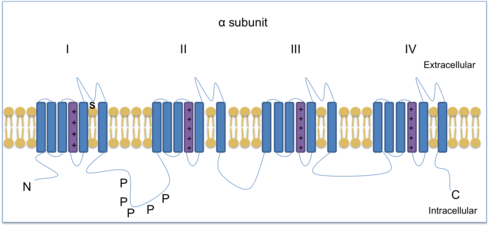
Kuchlanish qisqichi usullari Na ekanligini isbotladiV1.8, natriy kanallari orasida nisbatan depolarizatsiyalangan barqaror holatdagi inaktivatsiyani namoyish etishda noyobdir. Shunday qilib, NaVNeyronlar boshqa natriy kanallarini inaktivatsiya qiladigan darajalarga qadar depolarizatsiya qilinganida 1,8 ishlashga yaroqli bo'lib qoladi. DRG hujayralaridagi ta'sir potentsiallari TTXga chidamli natriy kanallari tomonidan qanday shakllanishini ko'rsatish uchun kuchlanish qisqichi ishlatilgan. Nav1.8 nosiseptiv sezgir neyronlarda takrorlanadigan yuqori chastotali potentsiallarning depolarizatsiya bosqichini ta'minlashga eng katta hissa qo'shadi, chunki u tez faollashadi va zararli stimul.[14][15] Shuning uchun, Nav1.8 hissa qo'shadi giperaljeziya (og'riqqa sezgirlikni oshirish) va allodiniya (odatda uni keltirib chiqarmaydigan stimullardan og'riq), bu surunkali og'riq elementlari.[16] Nav1.8 nokaut sichqonlar tadqiqotlar shuni ko'rsatdiki, kanal yallig'lanish va neyropatik og'riq bilan bog'liq.[9][17][18] Bundan tashqari, Nav1,8 sovuq og'riqda hal qiluvchi rol o'ynaydi.[19] Haroratni 30 ° C dan 10 ° C gacha kamaytirish VGSClarning faollashishini sekinlashtiradi va shu sababli oqim kamayadi. Biroq, Nav1.8 sovuqqa chidamli bo'lib, nosiseptorlardan to ma'lumotga etkazish uchun sovuqda ta'sir potentsialini yaratishga qodir markaziy asab tizimi (CNS). Bundan tashqari, Nav1,8 nol sichqonlar harakat potentsialini ishlab chiqara olmadi, bu Na ni ko'rsatmoqdav1.8 sovuq haroratda og'riqni his qilish uchun juda muhimdir.[19]
Na biofizikasi bo'yicha dastlabki tadqiqotlar bo'lsa-daV1,8 kanal kemiruvchilar kanallarida o'tkazildi, yaqinda o'tkazilgan tadqiqotlar inson Na ning xususiyatlarini o'rganib chiqdiV1,8 kanal. Ayniqsa, inson NaV1.8 kanallari kemiruvchilarnikiga qaraganda depolyarizatsiya qilingan inaktivatsiyaga bog'liq bo'lgan kuchlanishni namoyon qiladi va u ham doimiy doimiy oqimni namoyish etadi.[20] Shunday qilib, insonning Na ta'siriVSensor neyronlarning otilishidagi 1,8 kanal kemiruvchilar Na dan kattaroq bo'lishi mumkinV1,8 kanal.
Na ning funktsional yutuqlariV1.8, og'riqli periferik neyropatiyalar bilan og'rigan bemorlarda aniqlangan DRG neyronlarini giper qo'zg'aluvchanligi aniqlandi va shu bilan og'riq sabablari hisoblanadi.[21][22] Garchi NaV1.8 odatda serebellum ichida ifodalanmaydi, uning ekspressioni MS (Multiple Sclerosis) hayvon modellarida va odam MS da serebellar Purkinje hujayralarida yuqori darajada tartibga solinadi.[23] Na ning mavjudligiVOdatda bu bo'lmagan serebellar neyronlar ichidagi 1,8 kanal, ularning qo'zg'aluvchanligini oshiradi va otishni o'rganish tartibini o'zgartiradi. in vitro,[24] eksperimental otoimmun ensefalomiyeliti bo'lgan kemiruvchilarda, MS modeli.[25] Xulq-atvor darajasida, Na ning ektopik ifodasiVSerebellar Purkinje neyronlari tarkibidagi 1,8 ning transgenik modeldagi vosita ish faoliyatini susaytirishi isbotlangan.[26]
Klinik ahamiyati
Og'riq signalizatsiya yo'llari
Nosiseptorlar boshqa sezgir neyronlardan farq qiladi, chunki ular faollashish chegarasi past va natijada ularning doimiy stimulga bo'lgan ta'sirini oshiradi. Shuning uchun nosiseptorlar kabi vositalar tomonidan osongina sezgir bo'ladi bradikinin va asab o'sishi omili, ular to'qimalarning shikastlanish joyida ajralib chiqadi va natijada ion kanallarining o'tkazuvchanligini o'zgartiradi. VGSClarning asab shikastlangandan keyin zichligi oshgani isbotlangan.[27] Shuning uchun VGSC-larni asab shikastlangandan so'ng ajralib chiqadigan turli xil giperaljezik moddalar modulyatsiya qilishi mumkin. Boshqa misollarga quyidagilar kiradi prostaglandin E2 (PGE2), serotonin va adenozin, ularning barchasi Na orqali oqimni oshirishga harakat qiladiv1.8.[28]
PGE kabi prostaglandinlar2 nosiseptorlarni termal, kimyoviy va mexanik ogohlantirishlarga sezgir qilishi va DRG sezgir neyronlarining qo'zg'aluvchanligini oshirishi mumkin. Bu PGE tufayli sodir bo'ladi2 Na savdosini modulyatsiya qiladivG-oqsil bilan bog'langan holda 1.8 EP2 retseptorlari, bu esa o'z navbatida faollashadi oqsil kinazasi A.[29][30] Protein kinaz A fosforillanadiv1.8 hujayra ichidagi joylarda, natijada natriy ionlarining oqimlari ko'payadi. PGE o'rtasidagi bog'liqlik uchun dalillar2 va giperaljeziya antiseptik deoksinukleotidni Na-ning urishidan kelib chiqadivSichqonlarning DRG-da 1.8.[31] Na ning yana bir modulyatoriv1.8 ning ε izoformidir PKC. Ushbu izoform yallig'lanish mediatori bradikinin va fosforilatlar Na bilan faollashadiv1.8, bu mexanik giperaljeziyani kuchaytiradigan sezgir neyronlarda natriy oqimining ko'payishiga olib keladi.[32]
Brugada sindromi
Mutatsiyalar SCN10A bilan bog'liq Brugada sindromi.[33]
Membrana savdosi
Yallig'langan yoki shikastlangan to'qimalarda asab o'sishining darajasi oshib, og'riq sezuvchanligini oshiradi (giperaljeziya).[34] Nerv o'sish omilining darajasi oshdi va o'sma nekrozi omil-a (TNF-a) Na ning regulyatsiyasini keltirib chiqaradivQo'shimcha oqsil orqali sezgir neyronlarda 1.8 p11 (II ilova zanjiri). Yordamida ko'rsatildi xamirturush-ikkita duragay p11 ning Na-ning N terminalidagi 28-aminokislota bo'lagiga bog'langan skrining usuliv1.8 va uni targ'ib qiladi translokatsiya uchun plazma membranasi. Bu og'riq paytida sezgir neyronlarning gipereksitilligiga yordam beradi.[35] yordamida yaratilgan sichqonlarda p11-null nosiseptiv sezgir neyronlar Cre-loxP rekombinaza tizimi, Na ning pasayishini ko'rsatingv1.8 plazma membranasidagi ifoda.[36] Shuning uchun p11 va Na o'rtasidagi o'zaro ta'sirlarni buzishv1.8 og'riqni kamaytirish uchun yaxshi terapevtik maqsad bo'lishi mumkin.
Yilda miyelinlangan tolalar, VGSClar joylashgan Ranvier tugunlari; ammo, miyelinsiz tolalarda VGSClarning aniq joylashuvi aniqlanmagan. NavBilan bog'liq klasterlarda 1,8 mielinatsiz tolalar topilgan lipidli raftlar ikkala DRG tolasi bo'ylab in vitro va jonli ravishda.[37] Lipidli raflar odam savdosi va ionlashtiruvchi kanallarni o'z ichiga olgan hujayra membranasini tashkil qiladi. Membranadagi lipidli raftlarni olib tashlash MβCD, bu kamayadi xolesterin plazma membranasidan, Na siljishiga olib keladiv1.8 membrananing sal bo'lmaydigan qismiga, ta'sirlanish potentsialining pasayishi va tarqalishiga olib keladi.[37]
Og'riqli periferik neyropatiyalar
Alamli periferik neyropatiyalar yoki kichik tolali neyropatiyalar bu neyropatik og'riqni keltirib chiqaradigan miyelinsiz nosisitseptiv S-tolalarining buzilishi; ba'zi hollarda ma'lum bir sabab yo'q.[38] Ushbu idiopatik neyropatiyalar bilan og'rigan bemorlarni genetik skrining tekshiruvida mutatsiyalar aniqlandi SCN9A gen, tegishli kanalni kodlovchi Nav1.7. A funktsiyadan kelib chiqadigan mutatsiya Na-davDRG sezgir neyronlarida joylashgan 1.7 bemorlarning 30 foizida topilgan.[39] Funktsiyaning ortib borishi bilan bog'liq bo'lgan bu mutatsiya DRG sezgir neyronlarining qo'zg'aluvchanligini (gipereksititatsiyasini) kuchayishiga va shu bilan og'riqning kuchayishiga olib keladi. Nav1.7 shu tariqa inson dardi bilan bog'liqligi isbotlangan; Nav1.8, aksincha, faqat yaqinda hayvonlarni o'rganishdagi og'riq bilan bog'liq edi. Na-da funktsional yutuq mutatsiyasi aniqlandiv1.8-kodlash SCN10A og'riqli periferik neyropati bo'lgan bemorlarda gen.[21] 104 nafar bemorni o'rganish idyopatik mutatsiyaga ega bo'lmagan periferik neyropatiyalar SCN9A ishlatilgan kuchlanish qisqichi va joriy qisqich bashorat qilish bilan birga usullari algoritmlar, va funktsiyalari bo'yicha ikkita mutatsiyani hosil qildi SCN10A uchta bemorda. Ikkala mutatsiya ham DRG sezgir neyronlarida qo'zg'aluvchanlikni kuchayishiga olib keladi va shuning uchun og'riq paydo bo'lishiga yordam beradi, ammo ularni amalga oshirish mexanizmi tushunilmaydi.
Adabiyotlar
- ^ a b v GRCh38: Ensembl relizi 89: ENSG00000185313 - Ansambl, 2017 yil may
- ^ a b v GRCm38: Ensembl relizi 89: ENSMUSG00000034533 - Ansambl, 2017 yil may
- ^ "Human PubMed ma'lumotnomasi:". Milliy Biotexnologiya Axborot Markazi, AQSh Milliy Tibbiyot Kutubxonasi.
- ^ "Sichqoncha PubMed ma'lumotnomasi:". Milliy Biotexnologiya Axborot Markazi, AQSh Milliy Tibbiyot Kutubxonasi.
- ^ "Entrez Gen: natriy kanal".
- ^ Rabert DK, Koch BD, Ilnicka M, Obernolte RA, Naylor SL, Herman RC, Eglen RM, Hunter JC, Sangameswaran L (1998 yil noyabr). "Odamning dorsal ildizi ganglionlaridan tetrodotoksinga chidamli kuchlanishli natriy kanali, hPN3 / SCN10A". Og'riq. 78 (2): 107–14. doi:10.1016 / S0304-3959 (98) 00120-1. PMID 9839820. S2CID 45480324.
- ^ Plummer NW, Meisler MH (1999 yil aprel). "Sutemizuvchilar natriy kanal genlarining rivojlanishi va xilma-xilligi". Genomika. 57 (2): 323–31. doi:10.1006 / geno.1998.5735. PMID 10198179.
- ^ Catterall WA, Goldin AL, Waxman SG (dekabr 2005). "Xalqaro farmakologiya ittifoqi. XLVII. Voltajli natriy kanallarining nomenklaturasi va tuzilishi-funktsiya aloqalari". Farmakologik sharhlar. 57 (4): 397–409. doi:10.1124 / pr.57.4.4. PMID 16382098. S2CID 7332624.
- ^ a b Akopian AN, Souslova V, Angliya S, Okuse K, Ogata N, Ure J, Smit A, Kerr BJ, McMahon SB, Boyce S, Hill R, Stanfa LC, Dikenson AH, Wood JN (iyun 1999). "Tetrodotoksinga chidamli natriy kanal SNS og'riq yo'llarida maxsus funktsiyaga ega". Tabiat nevrologiyasi. 2 (6): 541–8. doi:10.1038/9195. PMID 10448219. S2CID 17487906.
- ^ Akopian AN, Sivilotti L, Wood JN (yanvar 1996). "Sensorli neyronlar tomonidan ifodalangan tetrodotoksinga chidamli voltajli natriy kanali". Tabiat. 379 (6562): 257–62. Bibcode:1996 yil Natur.379..257A. doi:10.1038 / 379257a0. PMID 8538791. S2CID 4360775.
- ^ Cummins TR, Sheets PL, Waxman SG (2007 yil oktyabr). "Nozitseptsiyadagi natriy kanallarining roli: og'riq mexanizmlariga ta'siri". Og'riq. 131 (3): 243–57. doi:10.1016 / j.pain.2007.07.026. PMC 2055547. PMID 17766042.
- ^ a b Swanwick RS, Pristerá A, Okuse K (2010 yil dekabr). "Na (V) 1.8 savdosi". Nevrologiya xatlari. 486 (2): 78–83. doi:10.1016 / j.neulet.2010.08.074. PMC 2977848. PMID 20816723.
- ^ Strickland IT, Martindale JC, Woodhams PL, Reeve AJ, Chessell IP, McQueen DS (iyul 2008). "Surunkali yallig'lanishli og'riyotgan og'rig'i modelida kalamush tizza qo'shimchasini innervatsiya qiladigan dorsal ildiz ganglionlarining alohida populyatsiyasida NaV1.7, NaV1.8 va NaV1.9 ekspressionidagi o'zgarishlar". Evropa og'rig'i jurnali. 12 (5): 564–72. doi:10.1016 / j.ejpain.2007.09.001. PMID 17950013. S2CID 24952010.
- ^ Bler NT, Bean BP (2002). "Tetrodotoksin (TTX) ta'sirchan Na + oqimi, TTXga chidamli Na + oqimi va Ca2 + oqimi nosiseptiv sezgir neyronlarning ta'sir potentsialida". Neuroscience jurnali. 22 (23): 10277–10290. doi:10.1523 / JNEUROSCI.22-23-10277.2002. PMC 6758735. PMID 12451128.
- ^ Renganatan M, Cummins TR & Waxman SG (2001). "DR1 neyronlarida potentsial elektrogenezga Nav1.8 natriy kanallarining hissasi". Neyrofiziologiya jurnali. 86 (2): 629–640. doi:10.1152 / jn.2001.86.2.629. PMID 11495938.
- ^ Millan MJ (1999). "Og'riqni induktsiya qilish: integral tekshiruv". Neyrobiologiyada taraqqiyot. 57 (1): 1–164. doi:10.1016 / S0301-0082 (98) 00048-3. PMID 9987804. S2CID 206054345.
- ^ Matthews EA, Wood JN, Dickenson AH (2006 yil fevral). "Na (v) 1,8 null sichqonlar o'murtqa neyron faoliyatida stimulga bog'liq defitsitni ko'rsatmoqda". Molekulyar og'riq. 2: 1744-8069–2-5. doi:10.1186/1744-8069-2-5. PMC 1403745. PMID 16478543.
- ^ Jarvis MF, Honore P, Shieh CC, Chapman M, Joshi S, Zhang XF, Kort M, Carroll V, Marron B, Atkinson R, Tomas J, Liu D, Krambis M, Liu Y, McGaraughty S, Chu K, Roeloffs R , Zhong C, Mikusa JP, Hernandez G, Gauvin D, Wade C, Zhu C, Pai M, Scanio M, Shi L, Drizin I, Gregg R, Matulenko M, Hakeem A, Gross M, Jonson M, Marsh K, Vagoner. PK, Sallivan JP, Faltynek CR, Krafte DS (may 2007). "A-803467, kuchli va selektiv Nav1.8 natriy kanal blokerlari, kalamushdagi neyropatik va yallig'lanish og'riqlarini susaytiradi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 104 (20): 8520–5. doi:10.1073 / pnas.0611364104. PMC 1895982. PMID 17483457.
- ^ a b Zimmermann K, Leffler A, Babes A, Cendan CM, Carr RW, Kobayashi J, Nau C, Wood JN, Reeh PW (iyun 2007). "Sensorli neyronli natriy kanal Nav1.8 past haroratlarda og'riq uchun juda muhimdir". Tabiat. 447 (7146): 855–8. Bibcode:2007 yil natur.447..856Z. doi:10.1038 / nature05880. PMID 17568746. S2CID 4391511.
- ^ Xan S, Estacion M, Xuang J, Vasylyev D, Chjao P, Dib-Xaj SD, Vaksman SG (may, 2015). "Human Na (v) 1.8: kuchaytirilgan doimiy va rampa oqimlari inson DRG neyronlarining aniq otish xususiyatlariga yordam beradi". Neyrofiziologiya jurnali. 113 (9): 3172–85. doi:10.1152 / jn.00113.2015. PMC 4432682. PMID 25787950.
- ^ Huang J, Yang Y, Zhao P, Gerrits MM, Hoeijmakers JG, Bekelaar K, Merkies IS, Faber CG, Dib-Hajj SD, Waxman SG (2013 yil avgust). "Kichik tolali neyropatiya Nav1.8 mutatsiyasi aktivatsiyani giperpolarizatsiyalangan potentsialga o'tkazadi va dorsal ganglion neyronlarining qo'zg'aluvchanligini oshiradi". Neuroscience jurnali. 33 (35): 14087–97. doi:10.1523 / JNEUROSCI.2710-13.2013. PMC 6618513. PMID 23986244.
- ^ Black JA, Dib-Hajj S, Baker D, Newcombe J, Cuzner ML, Waxman SG (oktyabr 2000). "Sensorli neyronlarga xos natriy kanal SNS eksperimental allergik ensefalomiyelit bilan kasallangan sichqonlarning va sklerozli odamlarning miyasida g'ayritabiiy tarzda namoyon bo'ladi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 97 (21): 11598–602. Bibcode:2000PNAS ... 9711598B. doi:10.1073 / pnas.97.21.11598. PMC 17246. PMID 11027357.
- ^ Renganatan M, Gelderblom M, Black JA, Waxman SG (2003 yil yanvar). "Nav1.8 natriy kanallarining ifodasi serebellar Purkinje hujayralarining otish tartibini buzadi". Miya tadqiqotlari. 959 (2): 235–42. doi:10.1016 / s0006-8993 (02) 03750-2. PMID 12493611. S2CID 34784900.
- ^ Saab CY, Craner MJ, Kataoka Y, Waxman SG (sentyabr 2004). "Eksperimental allergik ensefalomiyelitda in Vivo jonli ravishda Purkinje hujayralarining anormal faolligi". Eksperimental miya tadqiqotlari. 158 (1): 1–8. doi:10.1007 / s00221-004-1867-4. PMID 15118796. S2CID 34656521.
- ^ Shields SD, Cheng X, Gasser A, Saab CY, Tyrrell L, Eastman EM, Iwata M, Zvinger PJ, Black JA, Dib-Hajj SD, Waxman SG (fevral, 2012). "Chanellopatiya ko'p skleroz modelida serebellar disfunktsiyaga yordam beradi". Nevrologiya yilnomalari. 71 (2): 186–94. doi:10.1002 / ana.22665. PMID 22367990. S2CID 25128887.
- ^ Devor M; Govrin-Lippmann R & Anjelides (1993). "Periferik sutemizuvchilar aksonlarida Na + kanal lmmunolokalizatsiyasi va asab shikastlanishi va neyromaning paydo bo'lishidan keyingi o'zgarishlar". Neuroscience jurnali. 13 (5): 1976–1992. doi:10.1523 / JNEUROSCI.13-05-01976.1993. PMC 6576562. PMID 7683047.
- ^ Gold MS, Reichling DB, Shuster MJ, Levine JD (fevral, 1996). "Giperalgezik vositalar nosiseptorlarda tetrodotoksinga chidamli Na + tokini oshiradi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 93 (3): 1108–12. Bibcode:1996 yil PNAS ... 93.1108G. doi:10.1073 / pnas.93.3.1108. PMC 40039. PMID 8577723.
- ^ Hector TH (1975 yil yanvar). "Shaffof asetat varag'i yordamida xromatografik yozuvlarni yaratishning oddiy usuli". Fiziologiya jurnali. 32 (1): 31–2. doi:10.1113 / jphysiol.1996.sp021604. PMC 1160802. PMID 8887754.
- ^ Liu S, Li Q, Su Y, Bao L (2010 yil mart). "Prostaglandin E2 oqsil kinaz A yo'li orqali hujayra ichidagi RRR motifi orqali Na1.8 savdosiga yordam beradi". Yo'l harakati. 11 (3): 405–17. doi:10.1111 / j.1600-0854.2009.01027.x. PMID 20028484. S2CID 997800.
- ^ Xasar SG, Gold MS va Levine JD (1998). "Tetrodotoksinga chidamli natriy oqimi kalamushdagi yallig'lanish og'rig'iga vositachilik qiladi". Nevrologiya xatlari. 256 (1): 17–20. doi:10.1016 / s0304-3940 (98) 00738-1. PMID 9832206. S2CID 5614913.
- ^ Vu DF, Chandra D, MakMaxon T, Vang D, Dadgar J, Xaraziya VN, Liang YJ, Vaksman SG, Dib-Xaj SD, Messing RO (aprel 2012). "NaV1.8 natriy kanalining PKCε fosforillanishi kanal funktsiyasini oshiradi va sichqonlarda mexanik giperaljeziya hosil qiladi". Klinik tadqiqotlar jurnali. 122 (4): 1306–15. doi:10.1172 / JCI61934. PMC 3315445. PMID 22426212.
- ^ Xu D, Barajas-Martines X, Pfeiffer R, Dezi F, Pfeiffer J, Buch T, Betzenxauzer MJ, Belardinelli L, Kahlig KM, Rajamani S, DeAntonio HJ, Myerburg RJ, Ito H, Deshmux P, Marieb M, Nam GB, Bhatia A, Hasdemir C, Hayssaguerre M, Veltmann C, Shimpf R, Borggrefe M, Viskin S, Antzelevitch C (2014 yil iyul). "SCN10A mutatsiyalari Brugada sindromi holatlarining katta qismi uchun javobgardir". Amerika kardiologiya kolleji jurnali. 64 (1): 66–79. doi:10.1016 / j.jacc.2014.04.032. PMC 4116276. PMID 24998131.
- ^ McMahon SB (mart 1996). "NGF yallig'lanish og'rig'i vositachisi sifatida". London Qirollik Jamiyatining falsafiy operatsiyalari. B seriyasi, Biologiya fanlari. 351 (1338): 431–40. Bibcode:1996RSPTB.351..431M. doi:10.1098 / rstb.1996.0039. PMID 8730782.
- ^ Okuse K, Malik-Hall M, Beyker MD, Poon WY, Kong H, Chao MV, Wood JN (iyun 2002). "Annexin II yorug'lik zanjiri sezgir neyronlarga xos natriy kanal ekspressionini boshqaradi". Tabiat. 417 (6889): 653–6. Bibcode:2002 yil Natur.417..653O. doi:10.1038 / nature00781. PMID 12050667. S2CID 4423351.
- ^ Fulkes T, Nassar MA, Leyn T, Metyus EA, Beyker MD, Gerke V, Okuse K, Dikenson AH, Vud JN (oktyabr 2006). "Nosiseptorlarda Annexin 2 nurli zanjiri p11 ni yo'q qilish somatosensor kodlashda nuqsonlarni keltirib chiqaradi va og'riqli harakatlarga olib keladi" (PDF). Neuroscience jurnali. 26 (41): 10499–507. doi:10.1523 / JNEUROSCI.1997-06.2006. PMC 6674704. PMID 17035534.
- ^ a b Pristerà A, Baker MD, Okuse K (2012). "Tetrodotoksinga chidamli kanallar va lipidli raftlar o'rtasidagi bog'liqlik sezgir neyronlarning qo'zg'aluvchanligini tartibga soladi". PLOS ONE. 7 (8): e40079. Bibcode:2012PLoSO ... 740079P. doi:10.1371 / journal.pone.0040079. PMC 3411591. PMID 22870192.
- ^ Hoeijmakers JG, Faber CG, Lauria G, Merkies IS, Waxman SG (2012 yil may). "Kichik tolali neyropatiyalar - diagnostika, patofiziologiya va boshqarish sohasidagi yutuqlar". Tabiat sharhlari. Nevrologiya. 8 (7): 369–79. doi:10.1038 / nrneurol.2012.97. PMID 22641108. S2CID 8804151.
- ^ Faber CG, Hoeijmakers JG, Ahn HS, Cheng X, Xan C, Choi JS, Estacion M, Lauria G, Vanhoutte EK, Gerrits MM, Dib-Hajj S, Drenth JP, Waxman SG, Merkies IS (yanvar 2012). "Idiopatik mayda tolali neyropatiyada Naν1.7 mutatsiyalar funktsiyasini olish". Nevrologiya yilnomalari. 71 (1): 26–39. doi:10.1002 / ana.22485. PMID 21698661. S2CID 11711575.
Qo'shimcha o'qish
- Okuse K (2007). "Og'riq signalizatsiya yo'llari: sitokinlardan ion kanallariga". Xalqaro biokimyo va hujayra biologiyasi jurnali. 39 (3): 490–6. doi:10.1016 / j.biocel.2006.11.016. PMID 17194618.
- Waxman SG (2013 yil iyul). "Og'riqli Na-channelopathies: kengayib borayotgan koinot". Molekulyar tibbiyot tendentsiyalari. 19 (7): 406–9. doi:10.1016 / j.molmed.2013.04.003. PMID 23664154.
- Lay J, Porreca F, Hunter JC, Gold MS (2004). "Voltajli natriy kanallari va giperaljeziya". Farmakologiya va toksikologiyaning yillik sharhi. 44: 371–97. doi:10.1146 / annurev.pharmtox.44.101802.121627. PMID 14744251.
- Wood JN, Boorman JP, Okuse K, Baker MD (oktyabr 2004). "Voltajli natriy kanallari va og'riq yo'llari". Neurobiology jurnali. 61 (1): 55–71. doi:10.1002 / neu.20094. PMID 15362153.
- Malik-Xoll M, Poon VY, Beyker MD, Vud JN, Okus K (Fevral 2003). "Sensor neyron oqsillari NaV1.8 natriy kanalining hujayra ichidagi domenlari bilan o'zaro ta'sir qiladi". Miya tadqiqotlari. Molekulyar miya tadqiqotlari. 110 (2): 298–304. doi:10.1016 / S0169-328X (02) 00661-7. PMID 12591166.
- Yamaoka K, Inoue M, Miyazaki K, Xirama M, Kondo C, Kinoshita E, Miyoshi H, Seyama I (mart 2009). "Sintetik ciguatoksinlar HEK293 hujayralarida ifoda etilgan Nav1.8 dan kelib chiqqan ximerik natriy kanallarini tanlab faollashtiradi". Biologik kimyo jurnali. 284 (12): 7597–605. doi:10.1074 / jbc.M806481200. PMC 2658054. PMID 19164297.
- Choi JS, Xudmon A, Vaksman SG, Dib-Haj SD (iyul 2006). "Kalmodulin DRG neyronlarida Nav1.8 natriy kanalining oqim zichligi va chastotaga bog'liq inhibisyonini tartibga soladi". Neyrofiziologiya jurnali. 96 (1): 97–108. doi:10.1152 / jn.00854.2005. PMID 16598065.
- Liu CJ, Priest BT, Bugianesi RM, Dulski PM, Feliks JP, Dik IE, Brochu RM, Knaus HG, Middleton RE, Kaczorowski GJ, Slaughter RS, Garcia ML, Köhler MG (fevral 2006). "NaV1.8 kanallari uchun yuqori quvvatli membrana potentsiali FRET asosidagi tahlil". Sinov va dori vositalarini rivojlantirish texnologiyalari. 4 (1): 37–48. doi:10.1089 / adt.2006.4.37. PMID 16506887.
- Browne LE, Blaney FE, Yusaf SP, Clare JJ, Wray D (Aprel 2009). "Nav1.8 kanalida ta'sir qiluvchi dorilarning strukturaviy determinantlari". Biologik kimyo jurnali. 284 (16): 10523–36. doi:10.1074 / jbc.M807569200. PMC 2667739. PMID 19233853.
- Rabert DK, Koch BD, Ilnicka M, Obernolte RA, Naylor SL, Herman RC, Eglen RM, Hunter JC, Sangameswaran L (1998 yil noyabr). "Odamning dorsal ildizi ganglionlaridan tetrodotoksinga chidamli kuchlanishli natriy kanali, hPN3 / SCN10A". Og'riq. 78 (2): 107–14. doi:10.1016 / S0304-3959 (98) 00120-1. PMID 9839820. S2CID 45480324.
- Plummer NW, Meisler MH (1999 yil aprel). "Sutemizuvchilar natriy kanal genlarining rivojlanishi va xilma-xilligi". Genomika. 57 (2): 323–31. doi:10.1006 / geno.1998.5735. PMID 10198179.
- Catterall WA, Goldin AL, Waxman SG (dekabr 2005). "Xalqaro farmakologiya ittifoqi. XLVII. Voltajli natriy kanallarining nomenklaturasi va tuzilishi-funktsiya aloqalari". Farmakologik sharhlar. 57 (4): 397–409. doi:10.1124 / pr.57.4.4. PMID 16382098. S2CID 7332624.