Qushlarning ko'rinishi - Bird vision

Vizyon uchun eng muhim ma'no qushlar, chunki yaxshi ko'rish xavfsiz parvoz uchun juda muhimdir. Qushlarning ko'rish moslashuvchanligini boshqalarnikidan ustun qo'yadigan bir qator moslashuvi mavjud umurtqali hayvonlar guruhlar; kaptar "qanotli ikki ko'z" deb ta'riflangan.[1] Qushlarning ko'zlari a-ga o'xshaydi sudralib yuruvchi, bilan siliyer mushaklari shaklini o'zgartirishi mumkin ob'ektiv ga nisbatan tezroq va ko'proq darajada sutemizuvchilar. Qushlarning hayvonot dunyosidagi kattaligiga nisbatan eng katta ko'zlari bor va shuning uchun harakat ko'zning suyak uyasi ichida cheklangan.[1] Odatda umurtqali hayvonlarda uchraydigan ikkita ko'z qovog'idan tashqari, u uchinchi shaffof harakatlanuvchi membrana bilan himoyalangan. Ko'zning ichki anatomiyasi boshqa umurtqali hayvonlarnikiga o'xshash, ammo tuzilishga ega pekten okuli, qushlarga xos.
Ba'zi qushlar guruhlari o'zlarining hayot tarzlari bilan bog'liq bo'lgan vizual tizimida o'ziga xos modifikatsiyaga ega. Yirtqich qushlar retseptorlarning juda yuqori zichligi va ko'rish keskinligini maksimal darajada oshiradigan boshqa moslashuvlarga ega. Ko'zlarining joylashishi ularga masofani aniq aniqlashga imkon beradigan yaxshi binokulyar ko'rish imkonini beradi. Tungi turlarining ko'zlari naychali, rang detektorlari soni kam, ammo kam nurda yaxshi ishlaydigan tayoq hujayralarining zichligi yuqori. Terns, marralar va albatroslar orasida dengiz qushlari qizil yoki sariq rangga ega yog 'tomchilari masofaviy ko'rishni yaxshilash uchun, ayniqsa, xavfli sharoitda rangli retseptorlarda.
Ko'zdan tashqari anatomiya
The ko'z qushning sudralib yuruvchilarnikiga juda o'xshash. Dan farqli o'laroq sutemizuvchi Ko'z, u sferik emas va tekisroq shakli uning ko'proq ko'rish maydonini diqqat markazida bo'lishiga imkon beradi. Suyak plitalari doirasi, sklerotik halqa, ko'zni o'rab oladi va uni qattiq ushlab turadi, ammo sutemizuvchilarda ham uchraydigan sudralib yuruvchi ko'zning yaxshilanishi shundan iboratki, linzalar oldinga siljiydi va retinada tasvir hajmini oshiradi.[2]

Ko'pgina qushlarning ko'zlari katta, juda yumaloq emas va faqat orbitalarda cheklangan harakatga qodir,[3] odatda gorizontal ravishda 10-20 ° (lekin ba'zi passerinalarda> 80 °). Shuning uchun qushlardagi bosh harakatlari ko'z harakatlariga qaraganda katta rol o'ynaydi.[4] Ikki ko'z odatda mustaqil ravishda harakat qiladi,[4][5] va ba'zi turlarda ular qarama-qarshi yo'nalishlarda muvofiqlashtirilgan harakat qilishlari mumkin.[6]
Ko'zlari boshlari yon tomonida bo'lgan qushlarning kengligi bor ko'rish maydoni, yirtqichlarni aniqlash uchun foydalidir, boshlari oldida ko'zlari bo'lganlar, masalan, boyqushlarda binokulyar ko'rish va ov qilishda masofani taxmin qila oladi.[7][8] The Amerika o'rmon aravasi gorizontal tekislikda 360 ° va vertikal tekislikda 180 ° har qanday qushlarning eng katta ko'rish maydoniga ega.[9]
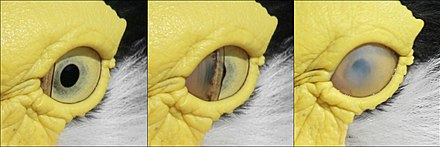
Qushlarning ko'z qovoqlari miltillashda ishlatilmaydi. Buning o'rniga ko'z moylanadi nikitatsiya qiluvchi membrana, ko'zni gorizontal ravishda shamol tozalagich singari siljigan uchinchi yashirin asr.[10] Nikitatsiya qiluvchi membrana shuningdek ko'zni qoplaydi va a rolini bajaradi kontakt linzalari suvda bo'lganida ko'plab suv qushlarida.[3] Uyqu paytida pastki qovoq ko'tariladi, aksariyat qushlarda ko'zni qoplash uchun shoxli boyqushlar bu erda yuqori ko'z qovog'i harakatchan.[11]
Ko'zni yoshdagi sekretsiya bilan ham tozalaydi lakrimal bez va yog'li moddalar bilan himoyalangan Harderian bezlari shox pardani qoplaydi va quruqlikni oldini oladi. Qushlarning ko'zi boshqa hayvon guruhlariga qaraganda hayvonning kattaligiga nisbatan kattaroqdir, ammo uning ko'p qismi bosh suyagida yashiringan. The tuyaqush quruqlikdagi umurtqali hayvonlarning eng katta ko'ziga ega, uning eksenel uzunligi 50 mm (2 dyuym), odamning ko'zidan ikki baravar ko'p.[1]
Qushlarning ko'zlari kattaligi tana massasi bilan bog'liq. Beshta buyruqni (to'tiqushlar, kaptarlar, petrellar, raptors va boyqushlar) o'rganish shuni ko'rsatdiki, ko'z massasi tana massasiga mutanosib, ammo ularning odatlari va vizual ekologiyasidan kutilganidek, raptors va boyo'g'li tanasining massasi uchun nisbatan katta ko'zlarga ega.[12]
Xulq-atvor tadqiqotlari shuni ko'rsatadiki, ko'plab parrandalar o'zlarining lateral va monokulyar ko'rish sohalari bilan uzoqroq ob'ektlarga e'tibor berishadi va qushlar vizual o'lchamlarini maksimal darajaga ko'tarish uchun o'zlarini yon tomonga yo'naltiradilar. Kabutar uchun rezolyutsiya yonma-yon monokulyar ko'rish bilan oldinga qarab durbinli ko'rishga qaraganda ikki baravar yaxshi, odamlar uchun esa bu teskari.[1]

Kam yorug'lik darajalarida ko'zning ishlashi linzalar va retinaning orasidagi masofaga bog'liq va kichik qushlar kunduzgi bo'lishga majbur bo'lishadi, chunki ularning ko'zlari etarli darajada tungi ko'rish uchun etarli emas. Ko'p turlarga qaramay ko'chib o'tish tunda ular ko'pincha dengiz chiroqlari yoki neft platformalari kabi yorqin yoritilgan narsalar bilan to'qnashadilar. Yirtqich qushlar kunlikdir, chunki ularning ko'zlari katta bo'lsa-da, ular yorug'lik yig'ilishidan ko'ra maksimal fazoviy rezolyusiyani berish uchun optimallashtirilgan, shuning uchun ham ular yomon nurda yaxshi ishlamaydi.[13] Ko'pgina qushlar ko'zning tuzilishidagi assimetriyaga ega, bu ularga ufqni va erning muhim qismini bir vaqtning o'zida diqqat markazida saqlashga imkon beradi. Ushbu moslashuvning qiymati shundaki, ular bor miyopi ularning ko'rish maydonining pastki qismida.[1]
Kabi tanasining massasi bilan solishtirganda nisbatan katta ko'zlari bo'lgan qushlar oddiy qizil yulduzlar va Evropa robinlari bir xil o'lchamdagi va tana massasi kichikroq qushlarga qaraganda erta tongda qo'shiq ayt. Ammo, agar qushlarning ko'zlari bir xil bo'lsa, lekin tana massalari har xil bo'lsa, katta turlar kichigidan kechroq kuylaydi. Buning sababi shundaki, kichkina qush bir kecha davomida vazn yo'qotishi sababli kunni ertaroq boshlashi kerak.[14] Kichkina qushlar uchun bir kechada vazn yo'qotish odatda 5-10% ni tashkil qiladi va qishning sovuq kechalarida 15% dan yuqori bo'lishi mumkin.[14] Bir tadqiqotda, robinlar kechasi sovuq bo'lganda, qorong'i ovqatlanishida ko'proq massa qo'yishdi.[15]
Tungi qushlarning ko'zlari sezuvchanligi uchun optimallashtirilgan bo'lib, ko'zning uzunligiga nisbatan katta shox pardalari bor, kunduzgi qushlar esa kornea diametriga nisbatan uzunroq ko'zlarga ega bo'lib, ko'rish qobiliyatini oshiradi. Yo'qolib ketgan turlarning faoliyati to'g'risida ma'lumotni sklerotik halqa va orbitaning chuqurligini o'lchashdan bilib olish mumkin. Oxirgi o'lchovni amalga oshirish uchun fotoalbom uch o'lchovli shaklini saqlab qolgan bo'lishi kerak, shuning uchun faollik naqshini tekislangan namunalardan ishonch bilan aniqlab bo'lmaydi. Arxeopteriks to'liq sklerotik halqaga ega, ammo orbitada chuqurlik o'lchovi yo'q.[16]
Ko'z anatomiyasi

Qush ko'zining asosiy tuzilmalari boshqasiga o'xshashdir umurtqali hayvonlar. Ko'zning tashqi qatlami shaffofdan iborat shox parda old tomonida va ikkita qatlam sklera - ko'zning qolgan qismini o'rab turgan va umuman ko'zni qo'llab-quvvatlaydigan va himoya qiladigan qattiq oq kollagen tolasi qatlami. Ko'z ichki tomonidan bo'linadi ob'ektiv ikkita asosiy segmentga: oldingi segment va orqa segment. Old qism suvli hazil deb nomlangan suvli suyuqlik bilan to'ldirilgan, orqa segment esa shisha jilosiga o'xshash moddadan iborat shishasimon hazilni o'z ichiga oladi.
Ob'ektiv tashqi qatlami yumshoqroq va ichki qatlami yumshoqroq bo'lgan shaffof qavariq yoki 'ob'ektiv' shaklidagi tanadir. U nurni retinaga qaratadi. Ob'ektiv shaklini zonular tolalar yordamida to'g'ridan-to'g'ri ob'ektiv kapsulasiga biriktirilgan siliyer mushaklari o'zgartirishi mumkin. Ushbu mushaklar bilan bir qatorda, ba'zi qushlarda shox parda shaklini o'zgartirishi mumkin bo'lgan ikkinchi to'plam - Krampton mushaklari ham mavjud, bu esa qushlarga ko'proq turar joy sutemizuvchilar uchun mumkin bo'lganidan. Ushbu turar joy sho'ng'in suvi qushlarida, masalan merganzerlarda tez bo'lishi mumkin. The ìrísí ko'zga tushadigan yorug'lik miqdorini boshqaradigan ob'ektiv oldida mushak orqali boshqariladigan rangli diafragma. Therísí markazida o'quvchi joylashgan bo'lib, u o'zgaruvchan dumaloq maydon orqali yorug'lik ko'zga o'tadi.[2][17]

The retina fotosensitivlikni o'z ichiga olgan nisbatan silliq kavisli ko'p qatlamli strukturadir novda va konusning hujayralari bog'liq neyronlar va qon tomirlari bilan. Fotoreseptorlarning zichligi maksimal ko'rish keskinligini aniqlashda juda muhimdir. Odamlarning har bir mm uchun 200000 ga yaqin retseptorlari bor2, lekin uy chumchuqi 400000 va oddiy shov-shuv 1 000 000. Fotoreseptorlar hammasi ham individual ravishda optik asab bilan bog'liq emas va ularning nisbati ganglionlar retseptorlarga rezolyutsiyani aniqlashda muhim ahamiyatga ega. Bu qushlar uchun juda baland; The oq dumaloq 100000 gangliyon hujayradan 120000 gacha fotoreseptorlarga ega.[2]
Tayoqchalar nurga nisbatan sezgirroq, ammo rang haqida ma'lumot bermaydilar, kam sezgir konuslar esa rangni ko'rish imkoniyatini beradi. Yilda kunduzgi qushlar, retseptorlarning 80% konus bo'lishi mumkin (ba'zilarida 90%) tezkorlar ) tungi boyqushlarda deyarli barcha tayoqchalar mavjud. Boshqa umurtqali hayvonlar singari plasental sutemizuvchilar, ba'zi konuslar bo'lishi mumkin er-xotin konuslar. Ular ba'zi turlardagi barcha konuslarning 50% ni tashkil qilishi mumkin.[18]
Retinaning markaziga qarab fovea (yoki kamroq ixtisoslashgan, markaziy markaz), bu retseptorlarning katta zichligiga ega va oldinga qarab ko'rish keskinligining maydoni, ya'ni ob'ektlarni eng aniq va aniq aniqlash. Qushlarning 54 foizida, shu jumladan yirtqich qushlar, qirg'oqchilar, kolbalar va qaldirg'ochlar, kengaytirilgan yon tomondan ko'rish uchun ikkinchi fovea mavjud. The optik asab bu ko'zdan miyaning tegishli qismlariga xabarlarni etkazib beradigan asab tolalari to'plami. Sutemizuvchilar singari, qushlarning ham kichigi bor ko'r nuqta optik diskda fotoreseptorlarsiz, uning ostida optik asab va qon tomirlari ko'zni birlashtiradi.[2]
The pekten bu to'r pardadan chiqadigan buklangan to'qimalardan tashkil topgan, yaxshi tushunilmagan tanadir. U qon tomirlari bilan yaxshi ta'minlangan va to'r pardasini ozuqaviy moddalar bilan ta'minlaydigan ko'rinadi,[1] shuningdek, ko'zning to'r pardasini ko'zni qamashtiradigan nurdan soya qilishi yoki harakatlanuvchi narsalarni aniqlashda yordam berishi mumkin.[2] Pekten okuli mo'l-ko'l melanin granulalari bilan to'ldirilgan bo'lib, ular fon parıltısını kamaytirish uchun parranda ko'ziga kiradigan yorug'likni yutish uchun taklif qilingan. Pekten metabolizmini kuchaytirish uchun melanin granulalari tomonidan nurni yutishi tufayli pekten okuli ozgina isishi taklif qilingan. Bu vitreus tanasida ozuqa moddalarining sekretsiyasini ko'paytirishga yordam beradi va oxir-oqibat ovqatlanishni yaxshilash uchun qushlarning avaskulyar retinasi tomonidan so'riladi.[19] Pekten okuli tarkibidagi ishqoriy fosfatazaning o'ta yuqori ferment faolligi retinaning ovqatlanishini to'ldirish uchun pektenning yuqori sekretor faolligini qo'llab-quvvatlash uchun taklif qilingan.[20]
The choroid retinaning orqasida joylashgan qatlam bo'lib, u ko'pgina mayda moddalarni o'z ichiga oladi arteriyalar va tomirlar. Ular retinani arterial qon bilan ta'minlaydi va venoz qonni to'kadi. Xoroid tarkibiga kiradi melanin, ichki ko'zga qorong'i rang beradigan va buzuvchi akslarning oldini olishga yordam beradigan pigment.
Yorug'lik hissi

Qushning ko'zida ikki xil yorug'lik retseptorlari mavjud, tayoqchalar va konuslar. Vizual pigmentni o'z ichiga olgan tayoqchalar rodopsin tungi ko'rish uchun yaxshiroqdir, chunki ular oz miqdordagi yorug'likka sezgir. Konuslar yorug'likning o'ziga xos ranglarini (yoki to'lqin uzunliklarini) aniqlaydi, shuning uchun ular qushlar kabi rangga yo'naltirilgan hayvonlar uchun muhimroqdir.[23] Ko'pincha qushlar tetrakromatik, ularning har biri o'ziga xos maksimal assimilyatsiya cho'qqisiga ega bo'lgan to'rt turdagi konus hujayralariga ega. Ba'zi qushlarda, eng qisqa to'lqin uzunligi uchun mas'ul bo'lgan konus hujayrasining maksimal singdirish cho'qqisi ultrabinafsha (UV) diapazoni, ularni ultrabinafsha nurlariga sezgir qiladi.[24] Bunga qo'shimcha ravishda, qushning to'r pardasidagi konuslar fazoviy taqsimotning o'ziga xos ko'rinishida joylashgan bo'lib, u giperuniform tarqalish deb nomlanadi, bu uning yorug'ligi va ranglarini yutishini maksimal darajada oshiradi. Fazoviy taqsimotning ushbu shakli faqat ba'zi bir optimallashtirish jarayoni natijasida kuzatiladi, bu holda qushlarning evolyutsion tarixi nuqtai nazaridan tavsiflanishi mumkin.[25]
To'rt spektral ravishda ajratilgan konus pigmentlari oqsildan olingan opsin deb nomlangan kichik molekulaga bog'langan setchatka bilan chambarchas bog'liq A vitamini. Pigment nurni yutganda, to'r pardasi shakli o'zgaradi va retinaning ganglion qatlamidagi neyronlarga ta'sir qiluvchi konus hujayrasining membrana potentsialini o'zgartiradi. Ganglion qatlamidagi har bir neyron bir nechta ma'lumotni qayta ishlashi mumkin fotoreseptor hujayralari, va o'z navbatida a ni keltirib chiqarishi mumkin asab impulsi miyadagi ixtisoslashgan vizual markazlarda qo'shimcha ishlov berish uchun optik asab bo'ylab ma'lumotlarni uzatish. Yorug'lik qanchalik kuchli bo'lsa, vizual pigmentlar shunchalik ko'p fotonlarni yutadi; har bir konusning qo'zg'alishi qanchalik katta bo'lsa va yorqinroq yorug'lik paydo bo'ladi.[23]
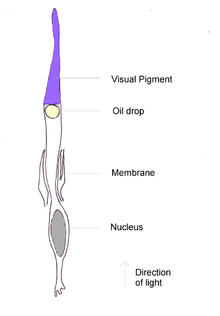
Ko'rib chiqilgan har bir qush turidagi eng ko'p konus pigmenti uzun to'lqin shaklidir yodopsin to'lqin uzunliklarida 570 nm ga yaqin yutadi. Bu taxminan primat retinada qizil va yashil rangga sezgir pigmentlar egallagan spektral mintaqa bo'lib, bu vizual pigment qushlarning rang sezgirligidan ustun turadi.[26] Yilda pingvinlar, bu pigment assimilyatsiya pikini 543 nm ga o'zgartirganga o'xshaydi, ehtimol bu ko'k suv muhitiga moslashish.[27]
Bitta konus orqali etkazilgan ma'lumot cheklangan: hujayra o'z-o'zidan miyaga yorug'likning qaysi to'lqin uzunligi qo'zg'atganligini aniqlay olmaydi. Vizual pigment ikkita to'lqin uzunligini teng ravishda o'zlashtirishi mumkin, ammo ularning fotonlari turli xil energiyaga ega bo'lishiga qaramay, konus ularni bir-biridan ajrata olmaydi, chunki ularning ikkalasi ham retinaning shakli o'zgarishiga olib keladi va shu bilan bir xil impulsni keltirib chiqaradi. Miya rangni ko'rish uchun turli xil ingl.Pigmentlarni o'z ichiga olgan ikki yoki undan ortiq konus sinflarining javoblarini taqqoslashi kerak, shuning uchun qushlardagi to'rtta pigment kamsitishni kuchaytiradi.[23]
Qushlarning yoki sudralib yuruvchilarning har bir konusida a rangli yog 'tomchisi; bular endi sutemizuvchilarda mavjud emas. Ning yuqori konsentratsiyasini o'z ichiga olgan tomchilar karotenoidlar, vizual pigmentga etib borishdan oldin yorug'lik ular orqali o'tadigan qilib joylashtirilgan. Ular filtr vazifasini bajaradi, ba'zi to'lqin uzunliklarini olib tashlaydi va pigmentlarning assimilyatsiya spektrlarini toraytiradi. Bu pigmentlar orasidagi javobning ustma-ust tushishini kamaytiradi va qush aniqlay oladigan ranglar sonini oshiradi.[23] Konus moyi tomchilarining olti turi aniqlandi; ulardan beshtasida karotenoid aralashmalari har xil to'lqin uzunliklarida va intensivligida so'riladi, oltinchi turda esa pigmentlar yo'q.[28] Eng past assimilyatsiya cho'qqisiga ega bo'lgan konus pigmentlari, shu jumladan ultrabinafsha nuriga sezgir bo'lganlar, spektral sozlash effektiga ega bo'lmagan "aniq" yoki "shaffof" yog 'tomchilariga ega.[29]
Retinal yog 'tomchilarining ranglari va tarqalishi turlar orasida sezilarli darajada farq qiladi va ishlatilgan ekologik maydonga (ovchi, baliqchi, o'txo'r) bog'liqdir. genetik munosabatlar. Masalan, kunduzgi ovchilar shunga o'xshash omborni yutish va yirtqich qushlarda rangli tomchilar kam, holbuki yer usti baliq ovi oddiy tern dorsal retinada qizil va sariq tomchilar ko'p. Dalillar shuni ko'rsatadiki, yog 'tomchilari javob berishadi tabiiy selektsiya konusning vizual pigmentlaridan tezroq.[26] Odamlarga ko'rinadigan to'lqin uzunliklari oralig'ida ham passerin qushlar odamlar ro'yxatdan o'tkazmaydigan rang farqlarini aniqlashi mumkin. Ushbu nozik diskriminatsiya, ultrabinafsha nurlarini ko'rish qobiliyati bilan birga, ko'plab turlar qushlarga ko'rinadigan, ammo odamlarga ko'rinmaydigan jinsiy dikromatizmni ko'rsatadi.[30]
Ko'chib yuruvchi qushlar o'zlarining migratsiya yo'nalishini aniqlash uchun Yerning magnit maydoni, yulduzlari, Quyosh va boshqa noma'lum signallardan foydalanadilar. Amerikalik tadqiqot shuni ko'rsatdiki, migratsiya Savanna chumchuqlari magnit navigatsiya tizimini quyosh chiqishida ham, quyosh botishida ham qayta kalibrlash uchun ufqqa yaqin osmon maydonidan qutblangan nur ishlatgan. Bu shuni ko'rsatadiki, osmon yoritgichi polarizatsiyasi naqshlari barcha ko'chib yuruvchi qushlar uchun asosiy kalibrlash moslamasi hisoblanadi.[31] Ammo, aftidan, qushlar qutblanish burchagi ikkilamchi ko'rsatkichlariga javob berishi mumkin va aslida bu ko'rsatmalar bo'lmaganda qutblanish yo'nalishini bevosita aniqlay olmasligi mumkin.[32]
Ultraviyole sezgirligi

Qushlarning ko'p turlari tetraxromatik bo'lib, yorug'lik spektrining ultrabinafsha va binafsha mintaqalarida to'lqin uzunligini sezish uchun ajratilgan konus hujayralari mavjud. Ushbu hujayralar tarkibiga qisqa to'lqinli sezgir (SWS1) opsinlar, SWS1 ga o'xshash opsinlar (SWS2) va uzoq to'lqinli filtrlovchi karotenoid pigmentlar kiradi.[33] 300 dan 400 nm gacha bo'lgan yorug'likni tanlab filtrlash va qabul qilish uchun. Qushlarda qisqa to'lqinli rang ko'rishning ikki turi mavjud: binafsha rang sezgir (VS) va ultrabinafsha sezgir (UVS).[34] SWS1 opsin ketma-ketligidagi bitta nukleotid o'rnini bosish opsin spektral sezuvchanligini binafsha rang sezgirdan (λmaksimal = 400) ultrabinafsha rangga sezgir (λmaksimal = 310-360).[35] Dastlab ultrabinafsha ko'rish vujudga kelgan evolyutsion mexanizm. Asosiy qoplamalar UVS ko'rish qobiliyatiga ega qushlar Paleognata (ratitlar va tinamous), Charadriiformes (qirg'oq qushlari, gullalar va alsidlar), Trogoniformes (trogonlar), Psittaciformes (to'tiqushlar) va Passeriformes (qushlarning sayg'oqlari, bu barcha parranda turlarining yarmidan ko'pini tashkil qiladi).[36]
UVS tuyulishi uchrashish uchun foydali bo'lishi mumkin. Ko'rinadigan to'lqin uzunliklarida jinsiy dikromatizmni ko'rsatmaydigan qushlar, ba'zida patlarida ultrabinafsha aks etuvchi parchalar borligi bilan ajralib turadi.[37][38] Erkak ko'k ko'krak ultrabinafsha aks ettiruvchi toj patchiga ega bo'lib, u o'zlarining patlarini ko'tarish va ko'tarish orqali uchrashishda namoyish etiladi.[39] Erkak ko'k grosbeaks shilliq qavatida eng yorqin va ultrafiolet nurlari o'zgargan ko'k rang kattaroq, mo'l-ko'l o'ljaga ega bo'lgan eng keng hududlarni o'z ichiga oladi va o'z avlodlarini boshqa erkaklarga qaraganda tez-tez boqadi.[23] O'rta er dengizi bo'ronlari ultrabinafsha naqshlarida jinsiy dimorfizmni ko'rsatmaydi, ammo ultrabinafsha nurlari va erkak tanasining holati o'rtasidagi bog'liqlik jinsiy tanlovda mumkin bo'lgan rolni ko'rsatadi.[40]
Xarajatlarni o'zaro ta'sirida qonunning ko'rinishi muhim ahamiyatga ega karapuz. Apelsin darajasi asosiy omil bo'lgan hududni ushlab turuvchi erkaklar o'rtasidagi o'zaro munosabatlarda ultrabinafsha nurlanish komponenti ahamiyatsiz bo'lib tuyulsa ham, ayol erkaklarga ultrafiolet nurlari yaxshi bo'lgan veksellar bilan javob beradi.[41]
UVS shuningdek, em-xashak vazifalarini bajarishi,[42] o'lja identifikatsiyasi,[43] va tejamkor. Trikromatik primatlarga o'xshash afzalliklar, tarkibidagi dikromatik primatlarga nisbatan tejamkor[44] odatda qushlarda mavjud deb hisoblanadi. Ko'plab mevalar va mevalarning mumsimon yuzalarida UVS nurlari aks etadi, ular UVS qushlariga ularning borligini reklama qiladi.[23] Oddiy kestrellar izlarini topishga qodir voles ko'rish bilan; bu kichkina kemiruvchilar siydik va najasning ultrafiolet nurlarini aks ettiruvchi xushbo'y yo'llarini yotqizib, ularni kestrellarga ko'rinadigan qiladi.[43] Biroq, bu fikr raptorlarda ultrabinafsha nurlanishining pastligi va sutemizuvchilar siydigining ultrabinafsha nurlarining zaif nurlanishini topishi bilan e'tiroz bildirilgan.[45]
Tetraxromatik ko'rish nafaqat qushlar uchun (hasharotlar, sudralib yuruvchilar va qisqichbaqasimonlar ham qisqa to'lqin uzunliklariga sezgir) ega bo'lishiga qaramay, UVS qushlarining ba'zi yirtqichlari ultrabinafsha nurlarini ko'ra olmaydilar. Bu ultrabinafsha ko'rish qushlarga maxsus signal bera oladigan kanalni berish imkoniyatini oshiradi va shu bilan yirtqichlar uchun sezilmaydigan bo'lib qoladi.[46] Biroq, so'nggi taxminlar ushbu farazni tasdiqlamaydi.[47]
Idrok
Kontrast sezgirligi
Kontrast (yoki aniqroq Mixelson-kontrast) ning farqi sifatida aniqlanadi nashrida ikkitasining yorqinligi yig'indisiga bo'linadigan ikkita ogohlantiruvchi maydon o'rtasida. Kontrast sezgirligi - bu aniqlanishi mumkin bo'lgan eng kichik kontrastning teskari tomoni; 100 ga teng bo'lgan kontrastli sezgirlik aniqlanishi mumkin bo'lgan eng kichik kontrastning 1% ekanligini anglatadi. Qushlarning kontrast sezgirligi sutemizuvchilardan kam. Odamlar qarama-qarshiliklarni 0,5-1% gacha aniqlaydilar[48] sinovdan o'tgan qushlarning ko'pchiligini talab qiladi. Xulq-atvorga javob berish uchun 10% kontrast.[49][50][51] Qarama-qarshi sezgirlik funktsiyasi hayvonning turli fazoviy chastotalardagi panjara naqshlarining kontrastini aniqlash qobiliyatini tavsiflaydi (ya'ni har xil detal). Statsionar ko'rish tajribalari uchun kontrastli sezgirlik o'rtacha fazoviy chastotada eng yuqori, yuqori va pastki fazoviy chastotalarda past bo'ladi.[52]
Harakat

Qushlar tezkor harakatlarni odamlarga qaraganda yaxshiroq hal qila olishadi, ular uchun soniyada 50 zarba pulsidan kattaroq miltillovchi doimiy harakat bo'lib ko'rinadi. Shuning uchun odamlar lyuminestsent lampochkaning soniyasiga 60 yorug'lik pulsining tsiklida tebranayotgan individual nurlarini ajrata olmaydilar, ammo budgerigars va tovuqlar soniyada miltillovchi yoki engil puls tsikllari soniyada 100 dan ortiq yorug'lik pulslari davrlariga ega.[iqtibos kerak ] A Kuperning kalxati chaqqon o'ljani o'rmonzor orqali ta'qib qilishi va yuqori tezlikda shoxlardan va boshqa narsalardan qochishi mumkin; odamlar uchun bunday ta'qib xiralashgan bo'lib ko'rinadi.[9]
Qushlar sekin harakatlanadigan narsalarni ham aniqlay olishadi. Quyosh va yulduz turkumlarining osmon bo'ylab harakatlanishi odamlarga sezilmaydi, lekin qushlar tomonidan aniqlanadi. Ushbu harakatlarni aniqlash qobiliyati ko'chib yuruvchi qushlarga o'zlarini to'g'ri yo'naltirishga imkon beradi.[9]
Parvoz qilayotganda yoki chayqalayotgan novdada o'tirganda barqaror tasvirlarni olish uchun qushlar kompensatsion reflekslar bilan boshni iloji boricha barqaror ushlab turadilar. Doimiy tasvirni saqlash, ayniqsa, yirtqich qushlar uchun dolzarbdir.[9] Tasvir bir vaqtning o'zida faqat bitta ko'zning chuqur fovasida joylashgan bo'lishi mumkinligi sababli, sho'ng'in paytida aksariyat lochinlar maqsadli shaxsga qulflangandan keyin o'ljalariga yaqinlashish uchun spiral yo'ldan foydalanadilar. Yaxshi ko'rinish uchun boshni burish alternativasi sho'ng'in tezligini oshirib, spiral tezlikni sezilarli darajada kamaytirmaydi.[53][54]
Yon va shakllar
Ob'ekt boshqasi tomonidan qisman to'sib qo'yilganda, odamlar ongsiz ravishda uni to'ldirishga va shakllarni to'ldirishga intilishadi (Qarang) Amodal idrok ). Ammo kaptarlarning tiqilib qolgan shakllarni to'ldirmasligi isbotlangan.[55] Fondan boshqacha rangga ega bo'lgan perchning kulrang darajasini o'zgartirishga asoslangan tadqiqot shuni ko'rsatdi budgerigars ranglarga asoslangan qirralarni aniqlamang.[56]
Magnit maydonlari
The magnit maydonlarni idrok etish Ko'chib yuruvchi qushlar tomonidan nurga bog'liq bo'lish taklif qilingan.[57] Magnit maydonning yo'nalishini aniqlash uchun qushlar boshlarini siljitadi,[58] va asab yo'llari bo'yicha olib borilgan tadqiqotlar shuni ko'rsatdiki, qushlar magnit maydonlarini "ko'rishlari" mumkin.[59] Ko'chib yuruvchi qushning o'ng ko'zida fotoreseptiv oqsillar mavjud kriptoxromlar. Yorug'lik bu molekulalarni qo'zg'atib, erning magnit maydoni bilan o'zaro aloqada bo'ladigan juft bo'lmagan elektronlarni hosil qiladi va shu bilan yo'naltirilgan ma'lumot beradi.[60][61]
Qushlar guruhlari bo'yicha farqlar
Kundalik yirtqich qushlar

Yirtqich qushlarning ko'rish qobiliyati afsonaviy bo'lib, ularning ko'rish qobiliyati turli xil omillarga bog'liq. Raptorlarning kattaligi katta ko'zlar, xuddi shu vazndagi qushlarning o'rtacha ko'rsatkichidan 1,4 baravar katta,[12] va ko'z naycha shaklida bo'lib, kattaroq to'r pardasi tasvirini hosil qiladi. Ko'zning rezolyutsiya kuchi ham optikaga bog'liq, katta teshiklari bo'lgan katta ko'zlar difraksiyadan kamroq aziyat chekadi va uzoq fokus masofasi tufayli retinaning kattaroq tasvirlariga ega bo'lishi mumkin va retseptorlar oralig'ining zichligiga bog'liq. Retinada kvadrat millimetrda juda ko'p sonli retseptorlar mavjud bo'lib, ular ko'rish keskinligining darajasini aniqlaydi. Hayvon qancha ko'p retseptorlarga ega bo'lsa, uning alohida ob'ektlarni masofadan ajratib olish qobiliyati shunchalik yuqori bo'ladi, ayniqsa, xuddi raptorlarda bo'lgani kabi, har bir retseptor odatda bitta ganglionga biriktirilgan.[1] Ko'plab raptorlarda fovealar odam foveasidan (65000 / mm) ancha ko'p novda va konuslarga ega2 Amerika kestrelida, odamlarda 38000 ta) va bu ushbu qushlarni uzoq masofadan turib ko'rish imkoniyatini beradi.[iqtibos kerak ] Raptorlarning chuqur markaziy fovasining shakli telefoto optik tizimni yaratishi mumkin,[62] foveadagi retinal tasvir hajmini oshirish va shu bilan mekansal o'lchamlarini oshirish. Xulq-atvor tadqiqotlari shuni ko'rsatadiki, ba'zi katta ko'zli raptorlar (takozli burgut, qadimgi dunyo qushqo'nmaslari) fazoviy rezolyusiyasiga nisbatan odamlarga nisbatan 2 baravar yuqori, ammo ko'plab o'rta va kichik o'lchamdagi raptorlarning fazoviy o'lchamlari taqqoslanadigan yoki pastroq.[63][64][65][66][67][68]
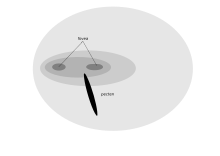
Yirtqich qushning oldinga qaragan ko'zlari durbinli ko'rishni beradi, unga er-xotin fovea yordam beradi.[2] Optimal vizual aniqlik uchun raptorning moslashuvi (an Amerika karahindidi 18 mm daraxtning tepasidan 2 mm hasharotni ko'rish mumkin) kamchiliklari bor, chunki uning yorug'ligi past darajadagi ko'rish qobiliyati zaif va u tunda uxlashi kerak.[1] Raptorslar ko'rish maydonining pastki qismida mobil o'lja bilan shug'ullanishlari kerak va shuning uchun ko'plab boshqa qushlar tomonidan ko'rsatiladigan pastki miyopi moslashuvi mavjud emas.[1] Qushlarni tozalash tulporlar bunday o'tkir ko'rish kerak emas, shuning uchun a kondor taxminan 35000 retseptorlari mm bo'lgan bitta fovaga ega2. Vultures, ammo ko'plab muhim fermentlarning uzoqdagi aniqligiga mos keladigan yuqori fiziologik faollikka ega.[70][iqtibos kerak ] Janubiy Karakara Shuningdek, faqat bitta fovea mavjud, chunki bu tur karrion va hasharotlar uchun erga yem beradi. Biroq, ular boshqa lochinlarga qaraganda durbin bilan bir-birining ustiga o'ralashish darajasiga ega, bu esa, ehtimol, qorakaraga toshlar kabi narsalarni boshqarish uchun imkon beradi.[71]
Tergov qilinayotgan boshqa qushlar singari, raptorlarning ham konuslarida rangli yog 'tomchilari bor.[65][66][72] Ushbu guruhning odatda jigarrang, kulrang va oq tuklari va uchrashishda rangli displeylarning yo'qligi, bu qushlar uchun rangning ahamiyatsizligini ko'rsatmoqda.[2]
Ko'pgina raptorlarda taniqli ko'z tizmasi va uning patlari ko'zning yuqorisida va oldida cho'zilgan. Ushbu "qosh" yirtqich qushlarga o'ziga xos qarashlarini beradi. Tog'lar jismonan ko'zni shamol, chang va qoldiqlardan himoya qiladi va uni haddan tashqari porlashdan saqlaydi. The osprey bu tizmaga etishmaydi, garchi uning ko'zlari ustidagi patlarning joylashishi shunga o'xshash vazifani bajarsa; Shuningdek, u ko'z oldida qorong'u tuklarga ega bo'lib, ular qush baliqning asosiy parhezi uchun ov qilayotganda, suv sathidan porlashni kamaytirishga xizmat qiladi.[9]
Tungi qushlar

Boyqushlarning kattaligi uchun juda katta ko'zlari bor, xuddi shu vazndagi qushlarning o'rtacha ko'rsatkichidan 2,2 baravar katta,[12] va boshning old qismida joylashgan. Ko'zlar maydonni 50-70% gacha qoplaydi va yaxshiroq beradi binokulyar ko'rish kunlik yirtqich qushlarga qaraganda (30-50% qoplanadi).[73] Tovuq boyo'g'li retina taxminan 56000 yorug'likka sezgir tayoqchalar kvadrat milimetr uchun (kvadrat dyuym uchun 36 million); ilgari u buni ko'rishi mumkinligini da'vo qilgan bo'lsa-da infraqizil qismi spektr ishdan bo'shatilgan.[74]

Kecha ko'rishga moslashish ko'zning katta hajmini, uning naychasimon shaklini, bir-biriga chambarchas o'ralgan ko'plab retinal tayoqchalarni va konusning yo'qligini o'z ichiga oladi, chunki konus hujayralari kam fotonli tungi muhit uchun sezgir emas. Yorug'lik intensivligini pasaytiradigan bir nechta rangli yog 'tomchilari mavjud, ammo retinada aks etuvchi qatlam mavjud tapetum lucidum. Bu nurga sezgir bo'lgan har bir hujayraning qabul qiladigan yorug'lik miqdorini oshiradi va kam yorug'lik sharoitida qushni yaxshiroq ko'rish imkonini beradi.[2] Boyqushlarda odatda bitta fovea bor, va u kabi rivojlangan kunduzgi ovchilardan tashqari, kam rivojlangan kalta quloqli boyqush.[73]
Boyqushlardan tashqari, kaltakesak, qurbaqalar va tungi mashinalar shuningdek, tungi yaxshi ko'rishni namoyish eting. Ba'zi qush turlari ko'rish uchun juda qorong'i bo'lgan g'or tizimlariga chuqur joylashadi va oddiy shakli bilan uyasiga yo'l topadi. echolokatsiya. The yog 'qushi echolokatsiya qiladigan yagona tungi qush,[75] lekin bir nechta Aerodramus tezyurar sportchilar ham ushbu usuldan foydalanadilar, bir tur bilan, Atiu tezkor, shuningdek, uning g'orlari tashqarisidagi ekolokatsiya yordamida.[76][77]
Suv qushlari

Kabi dengiz qushlari terns va marralar yuzasida oziqlanadigan yoki oziq-ovqat uchun sho'ng'adigan yog'ning qizil tomchilari bor konuslar ularning retinalar. Bu kontrastni yaxshilaydi va masofani ko'rishni keskinlashtiradi, ayniqsa xavfli sharoitda.[2] Havo / suv interfeysidan o'tishi kerak bo'lgan qushlar chuqurroq rangga ega karotenoid pigmentlar yog 'tomchilarida boshqa turlarga qaraganda.[26]
Bu ularga baliq shoalsini topishga yordam beradi, garchi ular ko'rayotganlari aniq emas fitoplankton baliqlar yoki boshqa oziqlanadigan qushlar boqadigan narsadir.[78]
Suvdan yashirincha baliq tutadigan qushlar, ayniqsa baliqlar burchak ostida kuzatilganda sinishi uchun tuzatishga to'g'ri keladi. Rif burchlari va kichik egretlar baliqlarni ovlashda zarur bo'lgan tuzatishlarni amalga oshira oladigan ko'rinadi va o'tkir burchak ostida zarbalar berilganda baliqlarni ushlashda yanada muvaffaqiyatli bo'ladi va bu yuqori yutuq baliqlarning o'z yirtqichlarini aniqlay olmasligi bilan bog'liq bo'lishi mumkin.[79] Boshqa tadqiqotlar shuni ko'rsatadiki, egretlar zarba berishning afzal burchagi doirasida ishlaydi va burchak vertikaldan juda uzoqlashganda, aniq va haqiqiy o'lja chuqurligi orasidagi farqni kuchayishiga olib kelganda yo'qolish ehtimoli ortadi.[80]
Baliqni suv ostida ta'qib qiladigan qushlar auks va g'avvoslar qizil moy tomchilari juda kam,[2] ammo ular maxsus egiluvchan linzalarga ega va nikitatsiya qiluvchi membranani qo'shimcha ob'ektiv sifatida ishlatadilar. Bu havo va suvda yaxshi ko'rish uchun ko'proq optik joylashishni ta'minlaydi.[3] Kormorantlarning ingl turar joy, 50 yoshda dioptrlar, boshqa qushlarga qaraganda, ammo qirg'oqchilar eng yaxshi har tomonlama (havo va suv) ko'rishga ega deb hisoblanadi.[2]
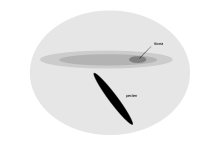
Tubenoz Dengiz qirg'oqlari, faqat nasl berish uchun va umrining katta qismini okean yuzasida adashib o'tkazish uchun keladi, retinada ingl.[1] Ushbu mintaqa giganto cellularis maydoni, topilgan Manx qirqish suvi, Kerguelen petrel, katta suv oqimi, keng pionli prion va umumiy sho'ng'in-petrel. U muntazam ravishda massivlangan va retinaning qolgan qismida joylashgan kattaroq ganglion hujayralarining mavjudligi bilan ajralib turadi va morfologik jihatdan retinaning hujayralariga o'xshaydi mushuklar. Ushbu yangi hududning joylashuvi va uyali morfologiyasi hisob-kitobning pastki qismida va atrofida proektsiyalangan kichik durbinli maydonda narsalarni aniqlashda qanday funktsiyani taklif qiladi. Bu, avvalambor, yuqori fazoviy rezolyutsiya bilan bog'liq emas, balki parranda pastdan uchib o'tganligi sababli, dengiz sathidan o'ljani topishda yordam berishi mumkin.[81]
The Manx qirqish suvi, boshqa ko'plab dengiz qushlari singari, tungi vaqtda naslchilik koloniyalariga tashrif buyurib, havo yirtqichlari tomonidan hujum qilish ehtimolini kamaytiradi. Uning optik tuzilishining ikki jihati shuni ko'rsatadiki, ushbu turning ko'zlari tunda ko'rishga moslashgan. Kesish suvi ko'zlarida linzalar nurning egilishining ko'p qismini to'r pardasida yo'naltirilgan tasvir hosil qilish uchun bajaradi. Shox parda, ko'zning tashqi qoplamasi nisbatan tekis va past darajada sinishi kuch. Kabutar kabi kunduzgi qushda teskari haqiqat; shox parda juda kavisli va asosiy refraktsion komponent hisoblanadi. Ob'ektivning sinishi bilan shox pardaning nisbati qirqma suv uchun 1,6, kaptar uchun 0,4; qirqma suvining ko'rsatkichi tungi qushlar va sutemizuvchilar qatoriga to'g'ri keladi.[82]
Qirqish suvi ko'zlarining fokus masofasi kaptarlarga qaraganda kichikroq, ammo yorqinroq tasvirni beradi, shuning uchun ikkinchisi kunduzgi ko'rishni yanada aniqroq qiladi. Manx qirqish suvi tunda ko'rish uchun moslashtirilgan bo'lsa-da, lekin ta'siri unchalik katta emas va ehtimol bu qushlar ham uyalarini topish uchun hid va eshitish vositalaridan foydalanadilar.[82]
Ilgari shunday deb o'ylaganlar pingvinlar quruqlikda uzoqni ko'ra bilishgan. Shox parda tekis va suv ostida suzishga moslashgan bo'lsa-da, ob'ektiv juda kuchli va suvsiz qolganda shox pardaning pasayishini qoplashi mumkin.[73] Deyarli qarama-qarshi echim qalpoqli merganser suv ostiga qo'yilganda ob'ektivning bir qismini ìrísí orqali shishirishi mumkin.[73]
Shuningdek qarang
Izohlar
- ^ a b v d e f g h men j Güntürkün, Onur, "Ko'zning tuzilishi va funktsiyalari" (Sturkie) (1998) 1-18
- ^ a b v d e f g h men j k Sinkler (1985) 88-100
- ^ a b v Gill, Frank (1995). Ornitologiya. Nyu-York: WH Freeman and Co. ISBN 978-0-7167-2415-5. OCLC 30354617.
- ^ a b Land, M. F. (2014). "Umurtqali hayvonlarning ko'z harakatlari va ularning ko'z shakli va faoliyati bilan aloqasi". Qiyosiy fiziologiya jurnali A. 201 (2): 195–214. doi:10.1007 / s00359-014-0964-5. PMID 25398576. S2CID 15836436.
- ^ Martin G. R. (2007). "Vizual maydonlar va ularning qushlardagi vazifalari". Ornitologiya jurnali. 148: 547–562. doi:10.1007 / s10336-007-0213-6. S2CID 24678783.
- ^ Voss J., Bischof H.-J. (2009). "Yanal ko'zli qushlarning ko'z harakatlari mustaqil emas" (PDF). Eksperimental biologiya jurnali. 212 (10): 1568–1575. doi:10.1242 / jeb.024950. PMID 19411551. S2CID 931601.
- ^ Martin, Grem R. Katzir, G (1999). "Qisqa barmoqli burgutlarda ingl. Maydonlar, Circaetus gallicus (Accipitridae), va qushlardagi durbinlik funktsiyasi ". Miya, o'zini tutish va evolyutsiyasi. 53 (2): 55–66. doi:10.1159/000006582. PMID 9933782. S2CID 44351032.
- ^ Tyrrell L. P., Fernández-Yuricic E. (2017). "Qushlarning durbinli ko'rinishi: Bu nafaqat qushlar ko'rishi mumkin, balki ular nimani ko'ra olmasligi haqida ham". PLOS ONE. 12 (3): e0173235. Bibcode:2017PLoSO..1273235T. doi:10.1371 / journal.pone.0173235. PMC 5371358. PMID 28355250.
- ^ a b v d e Jons, Maykl P; Pirs Jr, Kennet E.; Ward, Daniel (2007 yil aprel). "Qushlarning ko'rinishi: yirtqich qushlarni hisobga olgan holda shakl va funktsiyalarni ko'rib chiqish" (PDF). Ekzotik uy hayvonlari tibbiyoti jurnali. 16 (2): 69–87. doi:10.1053 / j.jepm.2007.03.012. Arxivlandi asl nusxasi (PDF) 2008-12-17 kunlari.
- ^ Uilyams, Devid L.; Flach, E (2003 yil mart). "Symblepharon qorli boyqushda nikitatsiya qiluvchi membrananing aberrant chiqishi bilan (Nyctea scandiaca)" (PDF). Veterinariya oftalmologiyasi. 6 (1): 11–13. doi:10.1046 / j.1463-5224.2003.00250.x. PMID 12641836. Arxivlandi asl nusxasi (PDF) 2008-12-17 kunlari.
- ^ Beebe, C. William (1906). Qush: uning shakli va vazifasi. Genri Xolt va Ko, Nyu-York. pp.214.
- ^ a b v Bruk, M. de L.; Xenli, S .; Laughlin, S. B. (1999 yil fevral). "Qushlarda tana massasi bilan ko'zning kattalashishi". Qirollik jamiyati materiallari B. 266 (1417): 405–412. doi:10.1098 / rspb.1999.0652. PMC 1689681.
- ^ Martin, Grem. Ziegler & Bischof (1993) 5–24 da "Tasvirni ishlab chiqarish"
- ^ a b Tomas, Robert J.; Suzuki, M; Seyto, S; Tanda, S; Nyusson, Styuart E.; Frayling, Tim D.; Wallis, Paul D. (2002). "Eye size in birds and the timing of song at dawn". Qirollik jamiyati materiallari B. 269 (1493): 831–837. doi:10.1098/rspb.2001.1941. PMC 1690967. PMID 11958715.
- ^ THOMAS, ROBERT (2002). "Body mass regulation and the daily singing routines of European robins". Hayvonlar harakati. 63 (2): 285–295. doi:10.1006/anbe.2001.1926. S2CID 54376020. Arxivlandi asl nusxasidan 2015 yil 1 yanvarda. Olingan 1 yanvar 2015.
- ^ Hall, Margaret I. (June 2008). "The anatomical relationships between the avian eye, orbit and sclerotic ring: implications for inferring activity patterns in extinct birds". Anatomiya jurnali. 212 (6): 781–794. doi:10.1111/j.1469-7580.2008.00897.x. PMC 2423400. PMID 18510506.
- ^ Sivak, Jacob G. (2004). "Through the Lens Clearly: Phylogeny and Development". Investitsiya. Oftalmol. Vis. Ilmiy ish. 45 (3): 740–747. doi:10.1167/iovs.03-0466. PMID 14985284.
- ^ Nalbach Hans-Ortwin; Wolf-Oberhollenzer, Friedericke; Remy Monika. "Exploring the image" in Ziegler & Bischof (1993) 26–28
- ^ Bawa, S.R.; YashRoy, R. (1974). "Structure and function of vulture pecten". Acta Anatomica. 89 (3): 473–480. doi:10.1159/000144308. PMID 4428954. Arxivlandi from the original on 2015-07-14.
- ^ Bawa, S.R.; YashRoy, R. (1972). "Effect of dark and light adaptation on the retina and pecten of chicken". Ko'zlarni eksperimental tadqiq qilish. 13 (1): 92–97. doi:10.1016/0014-4835(72)90129-7. PMID 5060117. Arxivlandi asl nusxasidan 2014-10-09.
- ^ Xart, NS; Partridge, J.C.; Bennett, A.T.D.; Cuthill, Innes C. (2000). "Visual pigments, cone oil droplets and ocular media in four species of estrildid finch" (PDF). Qiyosiy fiziologiya jurnali A. 186 (7–8): 681–694. doi:10.1007/s003590000121. PMID 11016784. S2CID 19458550. Arxivlandi asl nusxasi (PDF) on February 20, 2005.
- ^ The effect of the coloured oil droplets is to narrow and shift the absorption peak for each pigment. The absorption peaks without the oil droplets would be broader and less peaked, but these are not shown here.
- ^ a b v d e f Goldsmith, Timothy H. (July 2006). "What birds see" (PDF). Ilmiy Amerika: 69–75. Arxivlandi asl nusxasi (PDF) 2011 yil 10 aprelda.
- ^ Wilkie, Susan E.; Vissers, PM; Das, D; Degrip, VJ; Bowmaker, JK; Hunt, DM (1998). "The molecular basis for UV vision in birds: spectral characteristics, cDNA sequence and retinal localization of the UV-sensitive visual pigment of the budgerigar (Melopsittacus undulatus)". Biokimyoviy jurnal. 330 (Pt 1): 541–47. doi:10.1042/bj3300541. PMC 1219171. PMID 9461554.
- ^ "Hyperuniformity Found in Birds, Math and Physics - Quanta Magazine". kvantamagazine.org. Arxivlandi asl nusxasidan 2017 yil 9 aprelda. Olingan 6 may 2018.
- ^ a b v Varela, F. J .; Palasios, A. G.; Goldsmith T. M. "Color vision of birds" in Ziegler & Bischof (1993) 77–94
- ^ Bowmaker, J. K .; Martin, G. R. (January 1985). "Visual pigments and oil droplets in the penguin, Spheniscus humbolti". Qiyosiy fiziologiya jurnali. 156 (1): 71–77. doi:10.1007/BF00610668. S2CID 29312337.
- ^ Goldsmith, T. H.; Kollinz, JS; Licht, S (1984). "The cone oil droplets of avian retinas". Vizyon tadqiqotlari. 24 (11): 1661–1671. doi:10.1016/0042-6989(84)90324-9. PMID 6533991. S2CID 13730240.
- ^ Vorobyev, M.; Osorio, D.; Bennett, A. T. D.; Marshall, N. J .; Cuthill, I. C. (3 July 1998). "Tetrachromacy, oil droplets and bird plumage colours" (PDF). Qiyosiy fiziologiya jurnali A. 183 (5): 621–633. doi:10.1007/s003590050286. PMID 9839454. S2CID 372159. Arxivlandi asl nusxasi (PDF) 2012 yil 25 aprelda.
- ^ Eaton, Muir D. (August 2005). "Human vision fails to distinguish widespread sexual dichromatism among sexually "monochromatic" birds". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 102 (31): 10942–10946. Bibcode:2005PNAS..10210942E. doi:10.1073/pnas.0501891102. PMC 1182419. PMID 16033870. Arxivlandi asl nusxasi 2012-12-23 kunlari.
- ^ Muheim, Rachel; Phillips, JB; Akesson, S (August 2006). "Polarized light cues underlie compass calibration in migratory songbirds" (PDF). Ilm-fan. 313 (5788): 837–839. Bibcode:2006Sci...313..837M. doi:10.1126/science.1129709. PMID 16902138. S2CID 22251614. Arxivlandi asl nusxasi (PDF) 2008-12-17 kunlari.
- ^ Greenwood, Verity J.; Smith, EL; Church, SC; Partridge, JC (2003). "Behavioural investigation of polarisation sensitivity in the Japanese quail (Coturnix coturnix japonica) and the European starling (Sturnus vulgaris)". Eksperimental biologiya jurnali. 206 (Pt 18): 3201–3210. doi:10.1242/jeb.00537. PMID 12909701.
- ^ Toomey, Matthew B.; Kollinz, Aaron M.; Frederiksen, Rikard; Cornwall, M. Carter; Timlin, Jerilyn A.; Corbo, Joseph C. (2015-10-06). "A complex carotenoid palette tunes avian colour vision". Qirollik jamiyati interfeysi jurnali. 12 (111): 20150563. doi:10.1098/rsif.2015.0563. ISSN 1742-5689. PMC 4614492. PMID 26446559.
- ^ Odin, Anders; Håstad, Olle (11 February 2013). "The phylogenetic distribution of ultraviolet sensitivity in birds". BMC evolyutsion biologiyasi. 13: 36. doi:10.1186/1471-2148-13-36. PMC 3637589. PMID 23394614.
- ^ Yokoyama, Shozo; Radlwimmer, F. Bernhard; Blow, Nathan S. (2000-06-20). "Ultraviolet pigments in birds evolved from violet pigments by a single amino acid change". Milliy fanlar akademiyasi materiallari. 97 (13): 7366–7371. Bibcode:2000PNAS...97.7366Y. doi:10.1073/pnas.97.13.7366. ISSN 0027-8424. PMC 16551. PMID 10861005.
- ^ Ödeen, Anders; Håstad, Olle; Alström, Per (2011-10-24). "Evolution of ultraviolet vision in the largest avian radiation - the passerines". BMC evolyutsion biologiyasi. 11: 313. doi:10.1186/1471-2148-11-313. ISSN 1471-2148. PMC 3225180. PMID 22024316.
- ^ Hunt, Sarah; Bennett, Andrew T. D.; Cuthill, Innes C.; Griffiths, Richard (1998-03-22). "Moviy tits ultrabinafsha tits". London B Qirollik jamiyati materiallari: Biologiya fanlari. 265 (1395): 451–455. doi:10.1098 / rspb.1998.0316. ISSN 0962-8452. PMC 1688906.
- ^ Barreira, A. S.; Lagorio, M. G.; Lijtmaer, D. A.; Lougheed, S. C .; Tubaro, P. L. (2012-10-01). "Fluorescent and ultraviolet sexual dichromatism in the blue-winged parrotlet". Zoologiya jurnali. 288 (2): 135–142. doi:10.1111/j.1469-7998.2012.00931.x. ISSN 1469-7998.
- ^ Andersson, S .; J. Ornborg; M. Andersson (1998). "Ko'k ko'krakdagi ultrafiolet jinsiy dimorfizm va assortimentli juftlashish". Qirollik jamiyati materiallari B. 265 (1395): 445–50. doi:10.1098 / rspb.1998.0315. PMC 1688915. Arxivlandi from the original on 2008-10-11.
- ^ Soravia, Camilla; Aguado‐Giménez, Felipe; Avilés, Jesús M. (2020). "The reliability of achromatic displays is island-dependent in nocturnal Storm Petrels". Ibis. 162 (3): 655–672. doi:10.1111/ibi.12772. hdl:10261/202994. ISSN 1474-919X.
- ^ Bright, Ashleigh; Waas, Joseph R. (August 2002). "Effects of bill pigmentation and UV reflectance during territory establishment in blackbirds" (PDF). Hayvonlar harakati. 64 (2): 207–213. doi:10.1006/anbe.2002.3042. S2CID 51833485. Arxivlandi asl nusxasi (PDF) 2011-09-29 kunlari. Olingan 2008-07-21.
- ^ O'Daniels, Sean T.; Kesler, Dylan C.; Mihail, Jeanne D.; Webb, Elisabeth B.; Werner, Scott J. (2017). "Functional visual sensitivity to ultraviolet wavelengths in the Pileated Woodpecker ( Dryocopus pileatus ), and its influence on foraging substrate selection". Fiziologiya va o'zini tutish. 174: 144–154. doi:10.1016/j.physbeh.2017.02.041. PMID 28259805.
- ^ a b Viitala, Jussi; Korplmaki, Erkki; Palokangas, Palvl; Koivula, Minna (1995). "Kestrellarni ultrabinafsha nurlarda ko'rinadigan vole belgilariga jalb qilish". Tabiat. 373 (6513): 425–27. Bibcode:1995Natur.373..425V. doi:10.1038 / 373425a0. S2CID 4356193.
- ^ Osorio, D.; Vorobyev, M. (1996-05-22). "Rangni ko'rish primatlardagi tejamkorlikka moslashish sifatida". Proc. R. Soc. London. B. 263 (1370): 593–599. Bibcode:1996RSPSB.263..593O. doi:10.1098 / rspb.1996.0089. ISSN 0962-8452. PMID 8677259. S2CID 21822879.
- ^ Lind, Olle; Mitkus, Mindaugas; Olsson, Peter; Kelber, Almut (2013-05-15). "Ultraviolet sensitivity and colour vision in raptor foraging". Eksperimental biologiya jurnali. 216 (10): 1819–1826. doi:10.1242/jeb.082834. ISSN 0022-0949. PMID 23785106.
- ^ Hastad, Olle; Visctorsson, Jonas; Odeen, Anders (March 28, 2005). "Differences in colour vision make passerines less conspicuous in the eyes of their predators". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 102 (18): 6391–6394. Bibcode:2005PNAS..102.6391H. doi:10.1073/pnas.0409228102. PMC 1088363. PMID 15851662.
- ^ Stevens, Martin; Cuthill, Innes (2007). "Hidden Messages: Are Ultraviolet Signals a Special Channel in Avian Communication?". BioScience. 57 (6): 501–507. doi:10.1641/b570607.
- ^ De Valois, Russel L; Morgan, Herman; Snodderly, Max D (1974). "Psychophysical studies of monkey vision - III. Spatial luminance contrast sensitivity tests of macaque and human observers". Vizyon tadqiqotlari. 14 (1): 75–81. doi:10.1016/0042-6989(74)90118-7. PMID 4204839.
- ^ Harmening, Wolf M; Nikolay, P; Orlowski, J; Wagner, Hermann J (2009). "Spatial contrast sensitivity and grating acuity of barn owls". Vizyon jurnali. 9 (7): 13. doi:10.1167/9.7.13. PMID 19761328.
- ^ Jarvis, John R; Abeyesinghe, Siobhan M; McMahon, Claire E; Wathes, Christopher M (2009). "Measuring and modelling the spatial contrast sensitivity of the chicken (Gallus g. domesticus)". Vizyon tadqiqotlari. 49 (11): 1448–1454. doi:10.1016/j.visres.2009.02.019. PMID 19272401. S2CID 15267506.
- ^ Ghim, Mimi M; Hodos, William (2006). "Spatial contrast sensitivity of birds" (PDF). Qiyosiy fiziologiya jurnali A. 192 (5): 523–534. doi:10.1007/s00359-005-0090-5. hdl:1903/65. PMID 16404602. S2CID 1020197.
- ^ Uhlrich, Daniel J; Essock, Edward A; Lehmkuhle, Stephen (1981). "Cross-species correspondence of spatial contrast sensitivity functions". Xulq-atvorni o'rganish. 2 (3): 291–299. doi:10.1016/0166-4328(81)90013-9. PMID 6784738. S2CID 3996329.
- ^ Tucker, V. A.; Tucker, A. E.; Akers, K.; Enderson, J. H. (December 2000). "Curved flight paths and sideways vision in peregrine falcons (Falco peregrinus)". Eksperimental biologiya jurnali. 203 (Pt 24): 3755–3763. ISSN 0022-0949. PMID 11076739.
- ^ Tucker, V. A. (December 2000). "The deep fovea, sideways vision and spiral flight paths in raptors". Eksperimental biologiya jurnali. 203 (Pt 24): 3745–3754. ISSN 0022-0949. PMID 11076738.
- ^ Sekuler AB, Lee JA, Shettleworth SJ (1996). "Pigeons do not complete partly occluded figures". Idrok. 25 (9): 1109–1120. doi:10.1068/p251109. PMID 8983050. S2CID 31085585.
- ^ Bhagavatula P, Claudianos C, Ibbotson M, Srinivasan M (2009). Warrant E (ed.). "Edge Detection in Landing Budgerigars (Melopsittacus undulatus)". PLOS ONE. 4 (10): e7301. Bibcode:2009PLoSO...4.7301B. doi:10.1371/journal.pone.0007301. PMC 2752810. PMID 19809500.
- ^ Mouritsen, Henrik; Gesa Feenders, Miriam Liedvogel, Kazuhiro Wada & Erich D. Jarvis (2005). "Night-vision brain area in migratory songbirds". PNAS. 102 (23): 8339–8344. Bibcode:2005PNAS..102.8339M. doi:10.1073/pnas.0409575102. PMC 1149410. PMID 15928090.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Mouritsen, H.; Feenders, G; Liedvogel, M; Kropp, W (2004). "Migratory birds use head scans to detect the direction of the Earth's magnetic field" (PDF). Hozirgi biologiya. 14 (21): 1946–1949. doi:10.1016/j.cub.2004.10.025. hdl:11858/00-001M-0000-0028-7BC5-A. PMID 15530397. S2CID 11012106.[doimiy o'lik havola ]
- ^ Heyers D, Manns M, Luksch H, Güntürkün O, Mouritsen H (2007). Iwaniuk A (ed.). "A Visual Pathway Links Brain Structures Active during Magnetic Compass Orientation in Migratory Birds". PLOS ONE. 2 (9): e937. Bibcode:2007PLoSO...2..937H. doi:10.1371/journal.pone.0000937. PMC 1976598. PMID 17895978.
- ^ Shanor, Karen; Kanwal, Jagmeet (2009). Bats sing, mice giggle: revealing the secret lives of animals. Icon Books. p. 25. ISBN 978-1-84831-071-1. (Despite its title, this is written by professional scientists with many references)
- ^ Heyers, Dominik; Manns, M; Luksch, H; Güntürkün, O; Mouritsen, H; Iwaniuk, Andrew (September 2007). Ivaniuk, Endryu (tahrir). "A Visual Pathway Links Brain Structures Active during Magnetic Compass Orientation in Migratory Birds". PLOS ONE. 2 (9): e937. Bibcode:2007PLoSO...2..937H. doi:10.1371/journal.pone.0000937. PMC 1976598. PMID 17895978.
- ^ Snyder, Allan W.; Miller, William H. (1978). "Telephoto lens system of falconiform eyes". Tabiat. 275 (5676): 127–129. Bibcode:1978Natur.275..127S. doi:10.1038/275127a0. PMID 692679. S2CID 4204025.
- ^ Poitier, Simone; Bonadonna, Franchesko; Kelber, Almut; Duriez, Olivier (2016). "Visual acuity in an opportunistic raptor, the chimango caracara (Milvago chimango)". Fiziologiya va o'zini tutish. 157: 125–128. doi:10.1016/j.physbeh.2016.01.032. PMID 26821187. S2CID 36207899.
- ^ Gaffney, Matthew F; Hodos, William (2003). "The visual acuity and refractive state of the American kestrel (Falco sparverius)". Vizyon tadqiqotlari. 43 (19): 2053–2059. doi:10.1016/S0042-6989(03)00304-3. PMID 12842158. S2CID 17008291.
- ^ a b Reymond, Liz (1985). "Spatial visual acuity of the eagle Aquila audax: a behavioural, optical and anatomical investigation". Vizyon tadqiqotlari. 25 (10): 1477–1491. doi:10.1016/0042-6989(85)90226-3. PMID 4090282. S2CID 20680520.
- ^ a b Reymond, Liz (1987). "Spatial visual acuity of the falcon, Falco berigora: A behavioural, optical and anatomical investigation". Vizyon tadqiqotlari. 27 (10): 1859–1874. doi:10.1016/0042-6989(87)90114-3. PMID 3445475. S2CID 1526462.
- ^ Fischer, Anna Barbara (1969). "Laboruntersuchungen und freilandbeobachtungen zum sehvermögen under verhalten von altweltgeiern". Zoologische Jahrbüche Abteilung für Systematik (in German) (96): 81–132.
- ^ Martin, Graham (1986). "Shortcomings of an eagle's eye". Tabiat. 319 (6052): 357. doi:10.1038/319357a0. PMID 3945316. S2CID 4233018.
- ^ a b v Schematic diagram of retina of right eye, loosely based on Sturkie (1998) 6
- ^ Bawa, S.R.; YashRoy, R. "Vulture retina enzyme distribution and function". Neyrobiologiya. 2: 162–168. PMID 4218304. Arxivlandi from the original on 2015-11-26.
- ^ Potier, S.; Bonadonna, F.; Martin, G. R .; Isard, P.F.; Dulaurent, T.; Mentek, M.; Duriez, O. (2018). "Visual configuration of two species of Falconidae with different foraging ecologies". Ibis. 160 (1): 54–61. doi:10.1111/ibi.12528.
- ^ Sillman, A. J.; Bolnik, D. A .; Haynes, L. W.; Walter, A. E.; Loew, Ellis R. (1981). "Microspectrophotometry of the photoreceptors of palaegnathous birds - the emu and the tinamou". Qiyosiy fiziologiya jurnali A. 144 (3): 271–276. doi:10.1007/BF00612558. S2CID 7182768.
- ^ a b v d Burton (1985) 44–48
- ^ Hecht, Selig; Pirenne, MH (1940). "The sensibility of the nocturnal Long-Eared Owl in the spectrum". Journal of General Physiology. 23 (6): 709–717. doi:10.1085/jgp.23.6.709. PMC 2237955. PMID 19873186.
- ^ Kler, Nayjel; Nurney, David (1998). Nightjars: Dunyodagi Nightjars, Frogmouths, Potoos, Oilbird va Owlet-Nightjars uchun qo'llanma.. Pika / Kristofer Helm. p. 7. ISBN 978-1-873403-48-8. OCLC 39882046.
- ^ Fullard, J. H.; Barclay; Thomas (1993). "Echolocation in free-flying Atiu Swiftlets (Aerodramus sawtelli)" (PDF). Biotropika. 25 (3): 334–339. doi:10.2307/2388791. JSTOR 2388791. Arxivlandi asl nusxasi (PDF) 2008 yil 17-dekabrda. Olingan 12 iyul 2008.
- ^ Konishi, M .; Knudsen, EI (April 1979). "The oilbird: hearing and echolocation". Ilm-fan. 204 (4391): 425–427. Bibcode:1979Sci...204..425K. doi:10.1126/science.441731. PMID 441731.
- ^ Lythgoe, J. N. (1979). Vizyon ekologiyasi. Oksford: Clarendon Press. pp. 180–183. ISBN 978-0-19-854529-3. OCLC 4804801.
- ^ Lotem A; Schechtman E & G Katzir (1991). "Capture of submerged prey by little egrets, Egretta garzetta garzetta: strike depth, strike angle and the problem of light refraction" (PDF). Anim. Behav. 42 (3): 341–346. doi:10.1016/S0003-3472(05)80033-8. S2CID 37797506. Arxivlandi (PDF) asl nusxasidan 2011-06-04.
- ^ Katzir, Gadi; Lotem, Arnon; Intrator, Nathan (1989). "Stationary underwater prey missed by reef herons, Egretta gularis: head position and light refraction at the moment of strike" (PDF). Qiyosiy fiziologiya jurnali A. 165 (4): 573–576. doi:10.1007/BF00611243. S2CID 6191096. Arxivlandi asl nusxasi (PDF) 2016-03-04 da.
- ^ Hayes, Brian; Martin, Grem R. Brooke, Michael de L. (1991). "Procellariiform dengiz qushlarining retinasida binokulyar ko'rishni ta'minlovchi yangi hudud". Miya, o'zini tutish va evolyutsiyasi. 37 (2): 79–84. doi:10.1159/000114348. PMID 2054586.
- ^ a b Martin, Grem R. Brooke, M. de L. (1991). "The Eye of a Procellariiform Seabird, the Manx Shearwater, Puffinus puffinus: Visual Fields and Optical Structure". Miya, o'zini tutish va evolyutsiyasi. 37 (2): 65–78. doi:10.1159/000114347. PMID 2054585.
Adabiyotlar
- Burton, Robert (1985). Qushlarning harakati. London: Granada nashriyoti. ISBN 978-0-246-12440-1.
- Sinkler, Sandra (1985). Hayvonlar qanday ko'rishadi: Bizning dunyomizning boshqa qarashlari. Bekxem, Kent: Croom Helm. ISBN 978-0-7099-3336-6.
- Sturkie, P. D. (1998). Sturki qushlar fiziologiyasi. 5-nashr. Academic Press, San-Diego. ISBN 978-0-12-747605-6. OCLC 162128712.
- Zigler, Xarris Filipp; Bischof, Hans-Joachim, eds. (1993). Qushlardagi ko'rish, miya va o'zini tutish: qiyosiy tahlil. MIT Press. ISBN 978-0-262-24036-9. OCLC 27727176.
Tashqi havolalar
- Robert G. Cook, ed. (2001). Qushlarning vizual bilimi (cyberbook). Tufts universiteti; in cooperation with Comparative Cognition Press.
Vision in animals | ||
|---|---|---|
| Vizyon |  | |
| Ko'zlar | ||
| Evolyutsiya | ||
| Rang berish | ||
| Tegishli mavzular | ||