Qushlarning kelib chiqishi - Origin of birds

Hayvonlarning qaysi katta guruhi haqida ilmiy savol qushlar rivojlandi an'anaviy ravishda "qushlarning kelib chiqishi'. Hozirgi ilmiy konsensus shu qushlar guruhidir maniraptoran teropod dinozavrlar bu kelib chiqqan davomida Mezozoy erasi.
Qushlar va dinozavrlar o'rtasidagi yaqin munosabatlar birinchi marta XIX asrda ibtidoiy qush kashf etilganidan keyin taklif qilingan Arxeopteriks Germaniyada. Qushlar va yo'q bo'lib ketgan parranda bo'lmagan dinozavrlar ko'plab noyob skelet xususiyatlariga ega.[1] Bundan tashqari, parraklari saqlanib qolgan, qushlarga tegishli bo'lmagan dinozavrlarning o'ttizdan ortiq turlarining qoldiqlari to'plangan. Hatto juda kichik dinozavrlar ham bor Mikroraptor va Anchiornis uzoq vaqt bo'lgan, g'oyib bo'ldi qanot hosil qiluvchi qo'l va oyoq patlari. Yura davri bazali avialan Pedopenna bu uzun oyoq patlarini ham ko'rsatadi. Paleontolog Lourens Vitmer 2009 yilda ushbu dalillar qushlar evolyutsiyasi to'rt qanotli bosqichdan o'tganligini ko'rsatish uchun etarli degan xulosaga keldi.[2] Qoldiqlar, shuningdek, qushlar va dinozavrlarning bo'shliq, pnevmatik suyaklar, gastrolitlar ichida ovqat hazm qilish tizim, uyalarni qurish va tug'ma xatti-harakatlar.
Garchi qushlarning kelib chiqishi tarixan ichida munozarali mavzu bo'lgan evolyutsion biologiya, faqat bir nechta olimlar qushlarning dinozavr kelib chiqishi haqida bahslashmoqdalar va boshqa turlardan kelib chiqishini taxmin qilishmoqda arxosaurian sudralib yuruvchilar. Dinozavrlarning ajdodlarini qo'llab-quvvatlovchi kelishuv doirasida maniraptoran teropodlari tarkibidagi dastlabki qushlarni keltirib chiqargan evolyutsion hodisalarning aniq ketma-ketligi bahslidir. Kelib chiqishi qushlarning parvozi - bu alohida, lekin bir-biriga bog'liq bo'lgan savol, buning uchun bir nechta taklif qilingan javoblar mavjud.
Tadqiqot tarixi
Xaksli, Arxeopteriks va dastlabki tadqiqotlar

Qushlarning kelib chiqishi bo'yicha ilmiy tadqiqotlar 1859 yilda nashr etilganidan ko'p o'tmay boshlangan Charlz Darvin "s Turlarning kelib chiqishi to'g'risida.[3] 1860 yilda toshbo'ron qilingan pat topildi Germaniya "s Kech yura Solnhofen ohaktosh. Christian Erich Hermann fon Meyer bu tukni quyidagicha ta'rifladi Arxeopteriks litografikasi keyingi yil.[4] Richard Ouen 1863 yilda deyarli to'liq skeletni tasvirlab, uni eslatuvchi ko'plab xususiyatlarga qaramay uni qush deb tan oldi sudralib yuruvchilar, shu jumladan tirnoqli old oyoq va uzun suyakli dum.[5]
Biolog Tomas Genri Xaksli "Darvin Bulldogi" nomi bilan tanilgan, evolyutsiyaning yangi nazariyasini tabiiy selektsiya yordamida qat'iy qo'llab-quvvatlaganligi uchun Arxeopteriks kabi o'tish davri fotoalbomlari qushlar va sudralib yuruvchilar o'rtasida. 1868 yildan boshlab va ilgari berilgan takliflarga rioya qilgan holda Karl Gegenbaur,[6] va Edvard ichuvchisi,[7] Xaksli batafsil taqqoslashlarni amalga oshirdi Arxeopteriks turli xil tarixiy sudralib yuruvchilar bilan va uning dinozavrlarga o'xshashligini aniqladi Gipsilofodon va Kompsognatus.[8][9] 1870 yillarning oxirida kashfiyot "Berlin namunasi" ning Arxeopteriks, sudraluvchi tishlarning to'plami bilan to'ldirilgan, qo'shimcha dalillar keltirdi. Cope singari, Xaksli ham qushlar va dinozavrlar o'rtasidagi evolyutsion munosabatlarni taklif qildi. Garchi Xaksliga juda ta'sirli Ouen qarshi bo'lgan bo'lsa-da, uning xulosalari ko'plab biologlar tomonidan qabul qilindi, shu jumladan Baron Frants Nopcsa,[10] boshqalar, xususan Garri Sili,[11] o'xshashliklarga bog'liqligini ta'kidladi konvergent evolyutsiyasi.
Heilmann va teodont gipotezasi
Yigirmanchi asrning boshlarida yozilishlar bilan burilish yuz berdi Gerxard Heilmann ning Daniya. Ixtisos bo'yicha rassom Heilmann 1913 yildan 1916 yilgacha qushlarga ilmiy qiziqish ko'rsatgan va avvalgi asarlarini kengaytirgan Othenio Abel,[12] anatomiya bilan bog'liq bo'lgan tadqiqot natijalarini bir necha qismlarga ajratdi, embriologiya, qushlarning xulq-atvori, paleontologiya va evolyutsiyasi.[13] Dastlab yozilgan uning ishi Daniya kabi Vu Nuvaerende Viden om Fuglenes Afstamning, kompilyatsiya qilingan, ingliz tiliga tarjima qilingan va 1926 yilda nashr etilgan Qushlarning kelib chiqishi.

Xaksli singari, Heilmann ham taqqoslagan Arxeopteriks va boshqa qushlar tarixiy sudralib yuruvchilarning to'liq ro'yxatiga kiritilgan va shuningdek, terropod dinozavrlari yoqadi degan xulosaga kelishgan. Kompsognatus eng o'xshash edi. Biroq, Heilmann qushlar borligini ta'kidladi klavikula (yoqa suyaklari) suyak hosil qilish uchun birlashtirilgan furkula ("tilak suyagi") va klavikula ibtidoiy sudralib yuruvchilarda ma'lum bo'lgan bo'lsa-da, ular dinozavrlarda hali tan olinmagan edi. U qat'iy ishonganligi sababli Dolloning qonuni evolyutsiyasi qaytarilmasligini bildiruvchi Heilmann klavikulalar dinozavrlarda yo'qolganini va qushlarda qayta rivojlanganligini qabul qila olmadi. Shuning uchun u dinozavrlarni qush ajdodlari sifatida chiqarib tashlashga va ularning barcha o'xshashliklarini aytib berishga majbur bo'ldi konvergent evolyutsiyasi. Heilmann, qushlarning ajdodlari o'rniga ibtidoiy odamlar orasida topilishini aytdi "kodont "sudralib yuruvchilarning darajasi.[14] Heilmannning nihoyatda puxta yondashuvi uning kitobining bu sohada klassikaga aylanishini ta'minladi va uning qushlarning kelib chiqishi haqidagi xulosalari, boshqa ko'plab mavzular singari, keyingi to'rt yilliklar davomida ham barcha evolyutsion biologlar tomonidan qabul qilindi.[15]
Klavikula nisbatan nozik suyaklardir, shuning uchun ularni yo'q qilish yoki hech bo'lmaganda tanib bo'lmaydigan darajada zarar etkazish xavfi mavjud. Shunga qaramay, Heilmann o'z kitobini yozishdan oldin ba'zi toshbaqa teropod klavikulalari qazilgan, ammo ular noto'g'ri aniqlangan.[16]Dinozavrlarda klavikulalarning yo'qligi ibtidoiy teropodda klavikulalar topilganiga qaramay, pravoslav nuqtai nazarga aylandi Segisaurus 1936 yilda.[17] Dinozavrdagi klavikulalarning navbatdagi hisoboti 1983 yilda ruscha maqolada bo'lgan.[18]
Xeylmann ishonganidan farqli o'laroq, paleontologlar klavikulalar va aksariyat hollarda furkulalar nafaqat ekropodlarning, balki odatiy xususiyat ekanligini qabul qilishmoqda saurischian dinozavrlar. 2007 yil oxirigacha suyaklangan furkulalar (ya'ni suyakdan emas, suyakdan yasalgan) xaftaga ) eng asosiy bazallardan tashqari barcha tipopropodlarda uchraydi, Eoraptor va Herrerasaurus.[19] Ibtidoiy teropoddagi furkulaning asl hisoboti Segisaurus (1936) 2005 yildagi qayta ekspertiza bilan tasdiqlangan.[20] Birlashtirilgan, furkulaga o'xshash klavikulalar ham topilgan Massospondil, erta yura davri sauropodomorf.[21]
Ostrom, Deinonychus va dinozavrlarning qayta tiklanishi

Olam 1964 yilda yangi teropod dinozavr kashf etilgandan so'ng "kodon" gipotezasiga qarshi burila boshladi. Montana. 1969 yilda ushbu dinozavr tavsiflangan va nomlangan Deinonychus tomonidan Jon Ostrom ning Yel universiteti.[22] Keyingi yil Ostrom uning namunasini qayta ko'rib chiqdi Pterodaktil ichida Golland Teyler muzeyi ning yana bir skeleti sifatida Arxeopteriks.[23] Namuna asosan bitta qanotdan iborat bo'lib, uning tavsifi Ostromni bilaklari orasidagi o'xshashliklardan xabardor qildi Arxeopteriks va Deinonychus.[24]
1972 yilda, Inglizlar paleontolog Alik Uoker qushlarning "kodonts" dan emas, balki paydo bo'lganligi haqidagi faraz timsoh ajdodlar kabi Sphenosuchus.[25] Ostromning ham tropodlar, ham erta qushlar bilan ishlashi uni 70-yillarning o'rtalarida bir qator nashrlar bilan javob berishga undadi, unda u qushlar va terropod dinozavrlari o'rtasidagi ko'plab o'xshashliklarni bayon qilib, bir asr oldin Xaksli tomonidan ilgari surilgan g'oyalarni tiriltirdi.[26][27][28] Ostrom qushlarning dinozavr ajdodlarini tan olishi va dinozavrlar almashinuvi haqidagi boshqa yangi g'oyalar bilan bir qatorda[29] faollik darajasi va ota-ona g'amxo'rligi,[30] deb nomlanuvchi narsa boshlandi dinozavrlarning qayta tiklanishi, 1970-yillarda boshlangan va hozirgi kungacha davom etmoqda.
Ostromning vahiylari, shuningdek, filogenetik sistematikaning tobora ko'payib borayotgan davriga to'g'ri keldi (kladistika ), 1960-yillarda ishi bilan boshlangan Villi Xenig.[31] Kladistika - bu turlarni qat'iyan ularning evolyutsion munosabatlariga asoslangan holda tashkil etishning aniq usuli, bu ularning anatomik xususiyatlarida eng kam o'zgarishlarni nazarda tutuvchi evolyutsion daraxtni aniqlash orqali hisoblanadi. 1980-yillarda birinchi marta dinozavrlar filogeniyasiga kladistik metodologiya qo'llanildi Jak Gotye va boshqalar, qushlarning teropod dinozavrlaridan kelib chiqqan guruh ekanligini aniq ko'rsatib berishdi.[32] Dastlabki tahlillar shuni ko'rsatdiki, dromaeosaurid theropodlar Deinonychus qushlar bilan chambarchas bog'liq edi, natijadan beri ko'p marta tasdiqlangan.[33][34]
Xitoyda tukli dinozavrlar

1990-yillarning boshlarida bir nechta ajoyib qush qoldiqlari topildi Erta bo'r geologik shakllanishlar Xitoyning shimoli-sharqidagi Liaoning viloyatida.[35][36] 1996 yilda xitoylik paleontologlar ta'rif berishdi Sinozauropteriks qushlarning yangi turi sifatida Yixian shakllanishi,[37] ammo bu hayvon tezda bazal teropod dinozavr bilan chambarchas bog'liq deb tanildi Kompsognatus. Ajablanarlisi shundaki, uning tanasi uzun filamentli tuzilmalar bilan qoplangan. Ular "protofeathers" deb nomlangan va ko'rib chiqilgan gomologik qushlarning yanada rivojlangan patlari bilan,[38] garchi ba'zi olimlar ushbu bahoga qo'shilmaydilar.[39] Xitoy va Shimoliy Amerika olimlar tasvirlangan Caudipteryx va Protarxeopteriks ko'p o'tmay. Suyak xususiyatlariga asoslanib, bu hayvonlar parranda bo'lmagan dinozavrlar bo'lgan, ammo ularning qoldiqlari qushlarnikiga o'xshash to'liq shakllangan patlarni olib yurgan.[40] "Archaeoraptor ", holda tasvirlangan taqriz ning 1999 yilgi sonida National Geographic,[41] kontrabandali qalbakilashtirish bo'lib chiqdi,[42] ammo qonuniy qoldiqlar qonuniy va noqonuniy ravishda Yixiandan oqib chiqishda davom etmoqda. Tuklar yoki "protofeathers" Yixianda turli xil terropodlardan topilgan,[43][44] va qushlarga o'xshamaydigan qushlarga o'xshamaydigan dinozavrlarning kashfiyotlari,[45] shuningdek, parranda bo'lmagan dinozavrga o'xshash ibtidoiy qushlar,[46] deyarli parranda bo'lmagan theropodlar va qushlar o'rtasidagi morfologik bo'shliqni deyarli butunlay yopib qo'ygan.
Raqamli homologiya
O'rtasida munozara mavjud embriologlar va paleontologlar qo'llari bo'lsin teropod dinozavrlar va qushlar asosan farq qiladi falangal hisoblar, qo'ldagi falanjlar (barmoqlar) sonini hisoblash. Bu muhim va qattiq muhokama qilinadigan tadqiqot sohasi, chunki uning natijalari qushlar dinozavrlar (avlodlari) ekanligi haqidagi kelishuvga qarshi chiqishi mumkin.
Embriologlar va parranda-dinozavrlar bog'lanishiga qarshi bo'lgan ba'zi paleontologlar tuxum rivojlanishini ko'p marta o'rganish asosida qushlarning raqamlarini uzoq vaqt davomida II-III-IV raqamlariga kiritishgan.[47] Bu ko'pchilikda ekanligiga asoslanadi amniotlar, 5 barmoqli qo'lda hosil bo'lgan birinchi raqam IV o'q bo'lib, u asosiy o'qni rivojlantiradi. Shuning uchun embriologlar qushlardagi asosiy o'qni IV raqam, qolgan tirik qolgan raqamlarni esa II-III-IV deb aniqladilar. Ilg'orlarning qoldiqlari teropod (Tetanuralar ) qo'llarda I-II-III raqamlari bor ko'rinadi (ichida ba'zi nasllar) Avetheropoda shuningdek, kamaytirilgan IV raqamga ega[48]). Agar bu to'g'ri bo'lsa, unda qushlardagi raqamlarning II-III-IV rivojlanishi terropod (dinozavr) ajdodlariga qarshi ko'rsatkichdir. Biroq, yo'q ontogen (rivojlantiruvchi) asos bo'lib, qaysi raqamlar ekropod qo'lida joylashganligini aniq belgilab beradi (chunki bugungi kunda hech qanday parranda bo'lmagan teropodlar o'sishi va rivojlanib borishi kuzatilishi mumkin emas), teropod qo'lining markirovkasi mutlaqo aniq emas.
Paleontologlar an'anaviy ravishda I-II-III parranda raqamlarini aniqladilar. Ularning fikriga ko'ra, parropod dinozavrlari singari I-II-III qushlarning raqamlari saqlanib qolgan falanj formulasi bo'yicha. Arxosavrlar uchun falanj soni 2-3-4-5-3; ko'plab arxhosaur nasllari kamaytirilgan raqamlarga ega, ammo bir xil phalangeal formula qolgan raqamlarda. Boshqacha qilib aytganda, paleontologlarning ta'kidlashicha, raqamlar yo'qolishi paytida tashqi nasldan ichkariga qarab har xil nasabdagi arxaavrlar bir xil raqamlarni yo'qotadi. Ning uchta raqami dromaeosaurs va Arxeopteriks I-II-III raqamlarining I-II-III raqamlari bilan bir xil falanj formulasiga ega bo'ling bazal arxhosaurs. Shuning uchun yo'qolgan raqamlar V va IV bo'ladi. Agar bu to'g'ri bo'lsa, unda zamonaviy qushlar ham I-II-III raqamlariga ega bo'lishadi.[47] Shuningdek, 1999 yildagi bir nashr qushlarga olib boruvchi (shunday qilib I raqamini II, II dan III gacha va hokazolarga aylantiradigan) tropod chizig'ining raqamlarini ramkaga o'tkazishni taklif qildi.[49][50]Biroq, bu kabi ramkalarning siljishi amniotlarda kam uchraydi va qushlarning kelib chiqishi bilan mos kelish uchun - orqa qavatlarda emas, balki faqat qushlar-theropod nasabining oldingi oyoqlarida bo'lishi kerak edi (bu holat biron bir hayvonda noma'lum).[51]Bu deyiladi Raqamni lateral qisqartirish (LDR) ga nisbatan Raqamni ikki tomonlama qisqartirish (BDR) (shuningdek qarang Limuzavr[52]
Qisqartma nomi bilan tanilgan kichik ozchilik BAND (Qushlar dinozavr emas)[53] shu jumladan ornitologlar Alan Feduchcia va Larri Martin, kabi qushlar oldingi sudralib yuruvchilar bilan chambarchas bog'liqligini ta'kidlamoqda Longisquama yoki Euparkeriya, dinozavrlarga qaraganda.[54][55] Embriologik qushlarni o'rganish rivojlanish biologiyasi qushlar va dinozavrlarning oldingi oyoqlarida raqamli gomologiya haqida savollar tug'dirdi.[56] Biroq, qiyosiy anatomiya va filogenetik tomonidan keltirilgan aniq dalillar hamda Xitoydan olingan dramatik tukli dinozavr qoldiqlari tufayli qushlar olingan Dastlab Xaksli, keyinroq Nopksa va Ostrom tomonidan kurash olib borilgan dinozavrlar bugungi paleontologlar tomonidan yakdil qo'llab-quvvatlanmoqda.[15]
Termogen mushaklarning gipotezasi
2011 yildagi nashr nashrni kengaytirish uchun tanlov taklif qildi skelet mushaklari, parvoz evolyutsiyasidan ko'ra, bu qoplamaning paydo bo'lishi uchun harakatlantiruvchi kuch edi.[57][58] Muskullar istiqbolli jihatdan kattalashdi endotermik sauriyaliklar, ushbu gipotezaga ko'ra, ning yo'qolishiga javob sifatida umurtqali hayvonlar mitoxondrial birlashtiruvchi oqsil, UCP1,[59] qaysi termogen. Yilda sutemizuvchilar, UCP1 ichida ishlaydi jigarrang yog 'to'qimasi yangi tug'ilgan chaqaloqlarni himoya qilish gipotermiya. Zamonaviy qushlarda skelet mushaklari xuddi shunday funktsiyani bajaradi va ularning ajdodlarida qilgan deb taxmin qilinadi. Shu nuqtai nazardan, ikki oyoqlilik va boshqa parrandalar skelet o'zgarishlar mushaklarning yon ta'siri edi giperplaziya, parvoz yoki suzish uchun moslashtirishlarni o'z ichiga olgan old oyoqlarning keyingi evolyutsion modifikatsiyalari bilan va qoldiq, ikki oyoqlilikning ikkinchi darajali oqibatlari.
Filogeniya
Arxeopteriks tarixiy ravishda birinchi qush deb hisoblanadi yoki Urvogel. Qadimgi qazilma kashfiyotlari theropodlar va ular orasidagi bo'shliqni to'ldirgan bo'lsa ham Arxeopteriks, shuningdek, orasidagi bo'shliq Arxeopteriks va zamonaviy qushlar, filogenetik taksonomistlar, an'anaga muvofiq, deyarli har doim foydalaning Arxeopteriks Aves-ni aniqlashga yordam beradigan aniqlovchi sifatida.[60][61] Aves kamdan-kam hollarda a deb ta'riflangan toj guruhi faqat zamonaviy qushlardan iborat.[32] Deyarli barcha paleontologlar qushlarni shunday deb hisoblashadi coelurosaurian teropod dinozavrlar.[15] Coelurosauria ichida bir nechta kladistik tahlillar a qo'llab-quvvatladi qoplama nomlangan Maniraptora iborat terizinosauroidlar, oviraptorozavrlar, troodontidlar, dromaeosauridlar va qushlar.[33][34][62] Ulardan dromaeosauridlar va troodontidlar odatda qoplamada birlashtirilgan Deinonixozauriya, bu a opa-singillar guruhi qushlarga (birgalikda tugun pardasini hosil qiladi) Eumaniraptora ) poyada Parafonlar.[33][63]
Boshqa tadqiqotlar muqobil filogeniyalarni taklif qildi, unda odatda parranda bo'lmagan deb hisoblanadigan dinozavrlarning ayrim guruhlari parranda ajdodlaridan kelib chiqqan bo'lishi mumkin. Masalan, 2002 yilgi tahlil natijasida oviraptorozavrlar bazal qushlar ekanligi aniqlandi.[64] Alvaressauridlar, dan ma'lum Osiyo va Amerika kabi turli xil tasniflangan bazal maniraptorans,[33][34][65][66] paravviyalar,[62] singlisi takson ornitomimozavrlar,[67] shuningdek, ixtisoslashgan erta qushlar.[68][69] Jins Rahonavis, dastlab erta qush deb ta'riflangan,[70] bir nechta tadqiqotlarda parranda bo'lmagan dromaeosaurid ekanligi aniqlandi.[63][71] Dromaeosauridlar va troodontidlarning o'zlari ham Avesning tashqarisida emas, balki uning ichida yotishlari tavsiya etilgan.[72][73]
Qushlar va dinozavrlarni bog'laydigan xususiyatlar
Ko'pchilik anatomik[74] xususiyatlari qushlar va terropod dinozavrlari tomonidan taqsimlanadi.
Tuklar
Arxeopteriks, "tukli dinozavr" ning birinchi yaxshi namunasi 1861 yilda kashf etilgan. Birinchi namunasi Solnhofen ohaktosh Germaniyaning janubiy qismida, bu a lagerstätte, noyob va ajoyib geologik ajoyib detallarga ega bo'lgan qoldiqlari bilan tanilgan. Arxeopteriks a o'tish davri fotoalbomlari, Qush bo'lmagan theropod dinozavrlari va ular orasida aniq oraliq xususiyatlarga ega qushlar. Darvinning seminalidan atigi ikki yil o'tgach topilgan Turlarning kelib chiqishi, uning kashfiyoti tarafdorlari o'rtasida paydo bo'lgan munozarani qo'zg'atdi evolyutsion biologiya va kreatsionizm. Ushbu erta qush shunchalik dinozavrga o'xshashki, atrofdagi toshda, hech bo'lmaganda bitta patda aniq bir taassurot qoldirmasdan namuna bilan adashgan Kompsognatus.[75]

1990-yillardan boshlab bir qator qo'shimcha tukli dinozavrlar dinozavrlar va zamonaviy qushlar o'rtasidagi yaqin aloqaning yanada kuchli dalillarini taqdim etgan holda topildi. Ulardan birinchisi dastlab oddiy filamentli deb ta'riflangan protofeatherskabi dinozavr nasablarida qayd etilgan kompsognatidlar va tirannosauroidlar.[76] Biroq, zamonaviy qushlarnikidan farq qilmaydigan patlarni tez orada avialan bo'lmagan dinozavrlarda ham topishdi.[40]
Kichik ozgina tadqiqotchilar oddiy filamentli "protofeather" tuzilmalari shunchaki dinozavrlar terisi ostidagi yoki ularning suyagi suyaklari tarkibidagi kollagen tolasining parchalanishining natijasidir va bu kabi shubhasiz tuklar bo'lgan turlar, masalan. oviraptorozavrlar va dromaeosaurs dinozavrlar emas, balki dinozavrlarga aloqasi bo'lmagan haqiqiy qushlardir.[77] Biroq, tadqiqotlarning aksariyati, tukli dinozavrlar aslida dinozavrlar ekanligi va shubhasiz teropodlarning oddiyroq iplari oddiy patlarni ifodalaydi degan xulosaga kelishdi. Ba'zi tadqiqotchilar rangni o'z ichiga olganligini ko'rsatdilar melanin tuzilmalarda - bu patlarda kutilgan, ammo kollagen tolalari emas.[78] Boshqalar zamonaviy parrandalarning parchalanishini o'rganish natijasida, ilgari tuklar ham toshbo'ron qilish paytida boshdan kechirayotgan ezish kuchlariga duch kelganida ipli bo'lib ko'rinishini va taxmin qilingan "protofeather" lar ilgari o'ylanganidan ko'ra murakkabroq bo'lganligini namoyish etishdi.[79] "Protofeathers" ning batafsil tekshiruvi Sinosauropteryx prima individual tuklar markaziy kvilingdan iborat ekanligini ko'rsatdi (raxis) ingichka bilan tikanlar undan tarvaqaylab ketgan, tuzilishi jihatidan zamonaviy qush patlariga qaraganda o'xshash, ammo ibtidoiy.[80]

Skelet
Tuklar ko'pincha qushlar bilan bog'langanligi sababli, tukli dinozavrlar ko'pincha sifatida tanilgan yo'qolgan havola qushlar va dinozavrlar o'rtasida. Shu bilan birga, ikkala guruh tomonidan taqsimlangan ko'plab skelet xususiyatlari, bu uchun muhimroq aloqani anglatadi paleontologlar. Bundan tashqari, qushlar va dinozavrlar o'rtasidagi munosabatlar va parvoz evolyutsiyasi ilgari amalga oshirilganidan ko'ra murakkabroq mavzular ekanligi tobora ravshanlashib bormoqda. Masalan, ilgari qushlar dinozavrlardan bitta chiziqli progressiyada rivojlangan deb ishonilgan bo'lsa, ba'zi olimlar, eng muhimi Gregori S. Pol kabi dinozavrlar degan xulosaga kelishdi dromaeosaurs qushlardan rivojlangan bo'lishi mumkin, tuklarini zamonaviyga o'xshash tarzda ushlab turish paytida parvoz kuchini yo'qotadi tuyaqush va boshqalar ratitlar.
Qushlar va dinozavrlar skeletlarini taqqoslash, shuningdek kladistik tahlil, bog'lanish uchun ishni kuchaytiradi, xususan, deb nomlangan theropodlarning filiali uchun maniraptors. Suyak o'xshashliklariga bo'yin, pubis, bilak (yarim telba) karpal ), qo'l va ko'krak kamari, elka pichog'i, klavikula va ko'krak suyagi.
Embrional, balog'at yoshiga etmagan va kattalardagi arxosavr bosh suyaklarini taqqoslash bo'yicha olib borilgan tadqiqotlar natijasida qushlarning bosh suyaklari teropod tomonidan dinozavrlar nasli, paedomorfning bir turi heteroxroniya Bu ota-bobolarining voyaga etmaganlik xususiyatlarini saqlab qolishga olib keldi.[81]
O'pka

Patrik M. O'Konnor boshchiligidagi tergovga ko'ra, yirik go'sht yeyuvchi dinozavrlar zamonaviy qushlarda uchraydigan havo xaltachalarining murakkab tizimiga ega edi. Ogayo universiteti. Teropod dinozavrlarida (ikki oyog'ida yurgan va qushlarga o'xshash oyoqlari bo'lgan yirtqichlar) egiluvchan yumshoq to'qimalarning havo xaltachalari qushlarda bo'lgani kabi qattiq o'pka orqali havo pompalagan. "Bir vaqtlar rasmiy ravishda qushlarga xos bo'lgan narsa qushlarning ajdodlarida qaysidir ma'noda mavjud edi", dedi O'Konnor.[82][83]
Yurak
Kompyuter tomografiyasi (CT) skanerlashi 2000 yilda o'tkazilgan namunaning ko'krak qafasi bo'shlig'i ornithopod Felsevra hozirgi sutemizuvchilar va qushlarda uchraydigan kabi to'rt kamerali murakkab yurakning aniq qoldiqlarini topdi.[84] Ushbu g'oya ilmiy jamoatchilik orasida munozarali bo'lib, yomon anatomik fan deb tanqid qilindi[85] yoki shunchaki xayolparastlik.[86]
2011 yilda nashr etilgan tadqiqotda ob'ektning identifikatori haqidagi savolga bir nechta so'rovlar qo'llanildi, shu jumladan yanada mukammal kompyuter tomografiyasi, gistologiya, Rentgen difraksiyasi, Rentgen fotoelektron spektroskopiyasi va skanerlash elektroni mikroskopiya. Ushbu usullardan mualliflar quyidagilarni aniqladilar: ob'ektning ichki tuzilishi kameralarni o'z ichiga olmaydi, lekin zichligi past bo'lgan materialning uchta bir-biriga bog'liq bo'lmagan maydonlaridan tashkil topgan va ularning tuzilishi bilan taqqoslanmaydi. tuyaqush yuragi; "devorlar" dan tashkil topgan cho'kindi biologik tizimlarda ishlab chiqarilishi ma'lum bo'lmagan minerallar, masalan, goetit, dala shpati minerallar, kvarts va gips, shuningdek ba'zi o'simlik parchalari; uglerod, azot va fosfor, kimyoviy elementlar hayot uchun muhim bo'lgan, ularning namunalarida etishmayotgan edi; va yurakning uyali tuzilmalari yo'q edi. Hayvonlarning uyali tuzilmalari bilan bitta mumkin bo'lgan yamoq mavjud edi. Mualliflar o'zlarining ma'lumotlarini to'qimalarning izolyatsiya qilingan joylarini saqlab qolish ehtimoli bilan yurakdan emas, balki ko'milgan muhitdan olingan qum konkretsiyasi sifatida aniqladilar.[87]
Ushbu topilma metabolizm tezligini va dinozavrlarning ichki anatomiyasini qanday aks ettiradi degan savol, ob'ektning o'ziga xos xususiyatidan qat'i nazar, juda muhimdir.[87] Ikkalasi ham zamonaviy timsohlar va qushlar, dinozavrlarning eng yaqin qarindoshlari, to'rt kamerali yuraklarga ega (timsohlarda o'zgartirilgan bo'lsa ham), shuning uchun dinozavrlar ham ularga ega bo'lishgan; tuzilish metabolizm tezligiga bog'liq emas.[88]
Uyqu holati
Qoldiqlar troodontlar Mei va Sinornitoidlar dinozavrlarning ba'zi zamonaviy qushlar kabi uxlab yotganliklarini, boshlarini qo'ltiqlariga tiqishganligini namoyish eting.[89] Boshni isitishga yordam bergan bo'lishi mumkin bo'lgan bu xatti-harakatlar zamonaviy qushlarga ham xosdir.
Reproduktiv biologiya
Tuxum qo'yishda urg'ochi qushlar oyoq-qo'llarida suyakning maxsus turini o'stiradilar. Bu medullariya suyak qattiq tashqi suyak ichida kaltsiyga boy qatlam bo'lib shakllanadi va kaltsiy manbai sifatida tuxum qobig'ini hosil qiladi. A qismlarining ichki ilik bo'shliqlarini qoplagan endostelyal suyak to'qimalarining mavjudligi Tyrannosaurus rex namunaning orqa a'zosi buni taxmin qildi T. rex shunga o'xshash reproduktiv strategiyalardan foydalangan va namuna ayol ekanligini aniqlagan.[90] Keyingi tadqiqotlar natijasida teropodda medullar suyagi topildi Allosaurus va ornithopod Tenontosaurus. Dinozavrlar qatorini o'z ichiga olganligi sababli Allosaurus va Tiranozavr olib kelgan chiziqdan ajralib chiqdi Tenontosaurus dinozavrlar evolyutsiyasida juda erta, bu shuni ko'rsatadiki, umuman dinozavrlar medullar to'qimasini ishlab chiqargan.[91]
Yoshlarni parvarish qilish va parvarish qilish

Bir nechta Citipati namunalari o'z uyasida tuxumlarning ustiga eng ko'p eslatadigan holatda yotganligi aniqlandi qoralash.[92]
Masalan, ko'plab dinozavr turlari Mayasaura, podalar orasida juda yosh va katta yoshdagi odamlarni aralashtirib, ular o'rtasida boy o'zaro ta'sirni ko'rsatmoqda.
Tishsiz dinozavr embrioni topildi, demak, yosh dinozavrni boqish uchun ba'zi ota-onalar g'amxo'rligi kerak bo'lishi mumkin, ehtimol kattalar dinozavri yosh dinozavrning og'ziga regurgitatsiya qilingan ovqat (qarang altrikial ). Ushbu xatti-harakatlar ko'plab qush turlarida uchraydi; ota-ona qushlari tuxumdan chiqqan og'ziga oziq-ovqatni qaytaradi.
Gizzard toshlari
Ham qushlar, ham dinozavrlar foydalanadilar g'ilof toshlar. Ushbu toshlar hayvonlar tomonidan oshqozonga kirgandan so'ng ovqat hazm qilish va oziq-ovqat va qattiq tolalarni parchalash uchun yutiladi. Bilan birgalikda topilganda fotoalbomlar, toshbaqa toshlari deyiladi gastrolitlar.[93]Gizzard toshlari ba'zi baliqlarda ham uchraydi (mollar, loy shad, va gillaroo, alabalık turi) va timsohlarda.
Molekulyar dalillar
Bir necha marotaba mezozoy dinozavrlari qoldiqlaridan DNK va oqsillarni qazib olish da'vo qilingan, bu qushlar bilan taqqoslashga imkon beradi. Bir nechta oqsillar taxminlarga ko'ra dinozavr qoldiqlarida aniqlangan,[94] shu jumladan gemoglobin.[95]
2005 yil mart sonida Ilm-fan, Doktor Meri Xigbi Shvaytser va uning jamoasi 68 million yoshli odamning ichidagi haqiqiy yumshoq to'qimalarga o'xshash egiluvchan material topilganligini e'lon qildi Tyrannosaurus rex oyoq suyak MOR 1125 namunasi Hell Creek Formation yilda Montana. Etti kollagen suyak bo'laklaridan olingan turlari, tirik qushlarning kollagen ma'lumotlari bilan solishtirganda (xususan, a tovuq ), keksa teropodlar va qushlar bir-biri bilan chambarchas bog'liq.[96] Yumshoq to'qima molekulyar taqqoslashga imkon berdi uyali anatomiya va oqsillarni ketma-ketligi ning kollagen 2007 yilda nashr etilgan to'qima, ikkalasi ham shuni ko'rsatdi T. rex va qushlar bir-birlari bilan ikkalasidan ham ko'proq bog'liqdir Alligator.[97][98] Ikkinchi molekulyar tadqiqotlar qushlarning dinozavrlarga bo'lgan munosabatini qat'iy qo'llab-quvvatladi, ammo kutilganidek Theropoda qushlarni joylashtirmadi. Ushbu tadqiqotda "mumiyalangan" femur suyagidan olingan sakkizta qo'shimcha kollagen sekanslari ishlatilgan. Brachylophosaurus canadensis MOR 2598 namunasi, a hadrosaur.[99] Biroq, bu natijalar juda tortishuvlarga sabab bo'ldi. Boshqa yo'q peptidlar mezozoy erasi haqida xabar berilgan. 2008 yilda taxmin qilingan yumshoq to'qimalar aslida bakterial mikrofilm ekanligi taxmin qilingan.[100] Bunga javoban, aynan shu mikrofilmlar yumshoq to'qimalarni himoya qiladi, deb ta'kidladilar.[101] Boshqa bir e'tiroz natijalar ifloslanish tufayli yuzaga kelishi mumkin edi.[102] 2015 yilda ifloslanishdan saqlanadigan ko'proq nazorat ostida sharoitlarda peptidlar aniqlandi.[103] 2017 yilda o'tkazilgan tadqiqotlar shuni ko'rsatdiki, zamonaviy tuyaqush suyagida peptid mavjud bo'lib, Tiranozavr va Brakilofozavr namunalar, o'zaro ifloslanish xavfini ta'kidlaydi.[104]
Qadimgi DNKning dinozavr qoldiqlaridan muvaffaqiyatli ekstraktsiyasi haqida ikki alohida holatda xabar berilgan, ammo keyingi tekshirish va taqriz, ushbu xabarlarning ikkalasi ham tasdiqlanmadi.[105]
Qushlarning uchishining kelib chiqishi
Qushlarning uchishining kelib chiqishi haqidagi bahs-munozaralar qushlar paydo bo'lgan g'oyadan deyarli qadimgi dinozavrlar, topilganidan ko'p o'tmay paydo bo'lgan Arxeopteriks 1862 yilda. O'shandan beri munozaralarning aksariyat qismida ikkita nazariya hukmronlik qilmoqda: kursor ("yerdan") nazariyasi qushlarning er yuzida yugurib yuradigan kichik, tezkor yirtqichlardan rivojlanishini taklif qiladi; Arboreal ("daraxtlardan pastga") nazariyasi shuni ko'rsatadiki, quvvatli parvoz daraxtsiz (daraxtga ko'tarilgan) hayvonlar tomonidan kuchsiz sirpanishdan kelib chiqqan. So'nggi nazariya, "qanotlarga yordam beradigan moyillik" (WAIR), bu kursor nazariyaning bir variantidir va qanotlar o'zlarining rivojlanishini taklif qiladi aerodinamik kichik tukli dinozavrlarning yirtqichlardan qochishiga yordam beradigan daraxtlar kabi juda baland qiyaliklarda tezlik bilan yugurish zarurati natijasida.
2018 yil mart oyida olimlar bu haqda xabar berishdi Arxeopteriks ehtimol qodir edi parvoz, lekin undan farqli ravishda farq qiladi zamonaviy qushlar.[106][107]
Kursorial ("yerdan") nazariyasi

Parvozning kelib chiqishiga oid kursor nazariyani birinchi marta taklif qilgan Samuel Wendell Williston va tomonidan ishlab chiqilgan Baron Nopcsa. Ushbu gipoteza shuni ko'rsatadiki, uzun dumli tez yuradigan ba'zi hayvonlar yugurish paytida muvozanatni saqlash uchun qo'llarini ishlatgan. Ushbu nazariyaning zamonaviy versiyalari Williston-Nopcsa versiyasidan ko'p tafsilotlar bilan farq qiladi, asosan Nopksa davridan beri kashfiyotlar natijasida.
Nopcsa, uzatilgan qo'llarning sirtini ko'paytirish, kichkina yirtqich hayvonlarning muvozanatini saqlashga yordam berishi mumkinligi va bilaklar tarozi cho'zilib ketganligi haqida nazariyani ilgari surdi. rivojlanayotgan tuklar ichiga. Tuklar hasharotlarni yoki boshqa o'ljalarni ushlash uchun ham ishlatilishi mumkin edi. Bora-bora hayvonlar rivojlanayotgan qanotlari yordamida uzoqroq masofalarga sakrab o'tdilar. Nopcsa parvoz evolyutsiyasining uch bosqichini ham taklif qildi. Birinchidan, hayvonlar passiv parvozni rivojlantirdilar, unda rivojlanayotgan qanot tuzilmalari bir xil vazifani bajargan parashyut. Ikkinchidan, ular qanot qoqib faol parvozga erishdilar. U foydalangan Arxeopteriks ushbu ikkinchi bosqichga misol sifatida. Nihoyat, qushlar uchish qobiliyatiga ega bo'lishdi.[108]

Hozirgi fikr shundan iboratki, tuklar tarozidan rivojlanmagan, chunki patlar har xil oqsillar.[109] Keyinchalik jiddiyroq bo'lsa, Nopcsa nazariyasi tuklar parvoz evolyutsiyasining bir qismi sifatida rivojlangan deb taxmin qiladi va so'nggi kashfiyotlar taxminning yolg'on ekanligini isbotlaydi.
Tuklar ichida juda keng tarqalgan coelurosaurian dinozavrlari (shu jumladan erta tirannosauroid Dilong ).[110] Zamonaviy qushlar deyarli barcha paleontologlar tomonidan koelurozavrlar deb tasniflanadi,[111] ozgina bo'lsa ham ornitologlar.[112][113][114] "Yerdan" gipotezasining zamonaviy versiyasida qushlarning ajdodlari kichik bo'lgan, tukli, quruqlikdagi yirtqich dinozavrlar (aksincha) yo'l egalari ularning ov uslubida[115]) o'ljani ta'qib qilishda muvozanat uchun old oyoqlarini ishlatgan va keyinchalik oyoq parraklari parvozni ta'minlaydigan va keyinchalik parvozni ta'minlaydigan tarzda rivojlangan. Tuklarning eng keng tarqalgan o'ziga xos funktsiyalari orasida zamonaviy qushlardagi kabi issiqlik izolyatsiyasi va raqobatbardosh displeylar mavjud.[116][117]
Hammasi Arxeopteriks qoldiqlari dengiz cho'kmalaridan kelib chiqadi va qanotlar qushlarga Iso Masih Lizard (oddiy reyhan ).[118]
So'nggi paytlarda "yerdan turib" gipotezasining rad etilishi zamonaviy versiyada qushlarning o'zgargan celurosaurian dinozavrlari haqidagi taxminlarini rad etishga urinishdir. Eng kuchli hujumlarga asoslangan embriologik tahlillar qushlarning qanotlari 2, 3 va 4 raqamlaridan hosil bo'ladi (odamlarda ko'rsatkich, o'rta va halqa barmoqlariga to'g'ri keladi. Qushlarning uchta raqamlaridan birinchisi alula, ulardan qochish uchun foydalanadilar to'xtash past tezlikda uchishda - masalan, qo'nish paytida). Kelurozavrlarning qo'llari esa 1, 2 va 3 (odamlarda bosh barmog'i va dastlabki ikki barmog'i) bilan hosil bo'ladi.[119] Shu bilan birga, ushbu embriologik tahlillar darhol "qo'l" ko'pincha boshqacha rivojlanib boradigan embriologik asoslarga qarshi chiqdi qoplamalar evolyutsiyasi jarayonida ba'zi raqamlarni yo'qotgan va qushlarning "qo'llari" 1, 2 va 3 raqamlaridan rivojlanadi.[120][121][122] Ushbu bahs juda murakkab va hal qilinmagan - qarang "Raqamli homologiya ".
Qanot yordamida nishab yugurish
The qanotli nishab yugurish (WAIR) gipotezasiga yoshlarni kuzatish sabab bo'ldi chukar jo'jalar va ularning qanotlari rivojlanganligini taklif qiladi aerodinamik daraxtlar tanasi kabi juda tik qiyaliklarda tez yugurish zarurati natijasida, masalan yirtqichlardan qochish.[123] Bu uni kursor ("yerdan") nazariyasining ixtisoslashgan turiga aylantiradi. Ushbu stsenariyda qushlar kerakligiga e'tibor bering downforce oyoqlarini kuchaytirishi uchun.[124][125] Ammo erta qushlar, shu jumladan Arxeopteriks, etishmadi elka mexanizmi zamonaviy qushlarning qanotlari tez va kuchli zarbalarni hosil qiladi. WAIR pastga tushish kuchi tepish natijasida hosil bo'lganligi sababli, erta qushlar WAIRga qodir emas edilar.[126] WAIR - bu osteologik ixtisoslashuvga ega bo'lmagan xulq-atvor xususiyati bo'lgani uchun parvoz zarbasining filogenetik joylashuvi Neornithes, mavjud bo'lgan barcha qushlarni o'z ichiga olgan guruh, WAIRning parranda parvozi uchun ajdod ekanligini yoki undan kelib chiqqanligini aniqlashga imkon bermaydi.[127]
Arboreal ("daraxtlardan pastga") nazariyasi

Arboreal gipotezaning aksariyat versiyalarida ta'kidlanishicha, qushlarning ajdodlari shoxdan shoxga otilib, daraxtlarda yashagan juda kichik dinozavrlardir. Ushbu kichik dinozavrda allaqachon tuklar bor edi, ular evolyutsiyada birgalikda tanlangan, aerodinamikada foydali bo'lgan va oxir-oqibat qanotlarni ishlab chiqaradigan uzoqroq, qattiqroq shakllarni ishlab chiqarishgan. Keyinchalik qanotlar rivojlanib, tobora takomillashib borayotgan bo'lar edi, ular pog'onani ko'proq boshqarish, parashyut bilan sakrash, sirpanish va pog'onali uchish imkoniyatini beradigan qurilmalar sifatida. Arboreal gipotezada shuni ta'kidlash kerakki, daraxt hayvonlari uchun aerodinamika ancha tejamkor bo'ladi, chunki bunday hayvonlar shunchaki minimal sirpanish tezligiga erishish uchun tushadi.[128][129]
Yura yoki erta bo'r davridagi bir nechta kichik dinozavrlar, ularning hammasi patlar bilan, ehtimol daraxt va / yoki aerodinamik moslashuvlarga ega deb talqin qilingan. Bunga quyidagilar kiradi Sansorioteriks, Epidexipteryx, Mikroraptor, Pedopenna va Anchiornis. Anchiornis bu mavzu uchun juda muhimdir, chunki u Yura davrining boshlarida, ancha oldin yashagan Arxeopteriks.[130]
Eng ibtidoiy qushlarning oyoq suyaklari nisbatlarini tahlil qilish Arxeopteriks va Konfutsiyornis, tirik turlar bilan taqqoslaganda, dastlabki turlar ham yerda, ham daraxtlarda yashagan bo'lishi mumkin.[131]
Bir tadqiqot shuni ko'rsatadiki, eng qadimgi qushlar va ularning yaqin ajdodlari daraxtlarga chiqmaslikgan. Ushbu tadqiqot dastlabki qushlarning oyoq tirnoqlari egriligi miqdori qushlarning qushlariga qaraganda zamonaviy zamin bilan oziqlanadigan qushlarda kuzatilganiga o'xshashligini aniqladi.[132]
Ning ahamiyati pasaygan Arxeopteriks
Arxeopteriks birinchi va uzoq vaqt davomida ma'lum bo'lgan yagona tuklar edi Mezozoy hayvon. Natijada, qushlarning rivojlanishi va qushlarning uchishi masalalari muhokama qilindi Arxeopteriks hech bo'lmaganda 1990-yillarning o'rtalariga qadar.
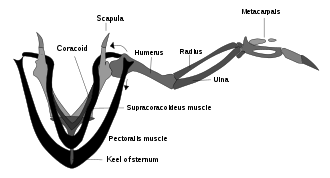
Bo'lishi haqida munozaralar bo'lib o'tdi Arxeopteriks chindan ham uchishi mumkin edi. Ko'rinib turibdiki Arxeopteriks qushlarning parvozlarini boshqarish uchun foydalanadigan miya tuzilmalari va quloq ichidagi muvozanat sezgichlari mavjud edi.[133] Arxeopteriks shuningdek, zamonaviy qushlar singari qanotli patlarni tartibga solgan va qanotlari va dumida xuddi shunday assimetrik uchish patlari bo'lgan. Ammo Arxeopteriks etishmadi elka mexanizmi zamonaviy qushlarning qanotlari tez va kuchli tepishlarni hosil qiladi (yuqoridagi suprakorakoid kasnagi diagrammasiga qarang); bu uning va boshqa qushlarning uchib yurishga qodir emasligini va faqat sirpanib tushishini anglatishi mumkin.[126]
Ammo 1990-yillarning boshlaridan beri kashfiyot ko'pchilik tukli dinozavrlar shuni anglatadiki Arxeopteriks endi qushlarning parvozi evolyutsiyasining asosiy ko'rsatkichi emas. Dan boshqa mayda tukli koleurozavrlar Bo'r va kech Yura davri qush parvozining mumkin bo'lgan prekursorlarini ko'rsatish. Bunga quyidagilar kiradi Rahonavis, bilan erga yuguruvchi Velociraptor - ikkinchi barmoq ustidagi ko'tarilgan o'roq tirnoqqa o'xshab, ba'zi paleontologlar parvozga qaraganda yaxshiroq moslashgan deb o'ylashadi Arxeopteriks,[134] Sansorioteriks, "daraxtlardan" nazariyasini qo'llab-quvvatlaydigan daraxtli dinozavr,[135] va Mikroraptor, arboreal dinozavr, ehtimol parvozga qodir, ammo agar shunday bo'lsa, ko'proq o'xshash ikki qanotli, chunki oyoqlarida yaxshi rivojlangan patlar bor edi.[136] 1915 yildayoq ba'zi olimlar qushlarning parvozi evolyutsiyasi to'rt qanotli (yoki) o'tgan bo'lishi mumkin deb ta'kidlashdi tetrapteryx) bosqich.[137][138]
Dinozavrlarda ikkilamchi parvozsizlik
| Koelurozavrlar |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Odatda qushlar deb qaraladigan guruhlar qalin turga kiradi.[73]
Hisobga olingan gipoteza Gregori Pol va kitoblarida keng tarqalgan Dunyoning yirtqich dinozavrlari (1988) va Havoning dinozavrlari (2002), parvoz qilmaydigan yirtqich dinozavrlarning ayrim guruhlari - ayniqsa deinonixosaurlar, lekin ehtimol boshqalar kabi oviraptorozavrlar, trizinozavrlar, alvaressauridlar va ornitomimozavrlar - aslida qushlardan tushadi. Pavlus, shuningdek, ushbu guruhlarning qush ajdodlari parvozga moslashish jihatidan ancha rivojlangan deb taklif qildi Arxeopteriks. Gipoteza shuni anglatadiki Arxeopteriks mavjud dinozavrlarga qaraganda kamroq mavjud qushlar bilan chambarchas bog'liq.[139]
Mayr paytida Polning gipotezasi qo'shimcha qo'llab-quvvatlandi va boshq. (2005) yangi, o'ninchi namunasini tahlil qildi Arxeopteriksva shunday xulosaga keldi Arxeopteriks singil Deinonixosauriyaga yopishgan edi, ammo bu rivojlangan qush Konfutsiyornis Dromaeosauridae ichida bo'lgan. Ushbu natija Polning gipotezasini qo'llab-quvvatlaydi, Deinonixosauriya va Troodontidae Avesning bir qismi, qushlarning nasabiga to'g'ri keladi va ikkinchidan uchib bo'lmaydi.[140] Shu bilan birga, ushbu hujjat boshqa barcha qushlarni hisobga olmagan va shu bilan ularning belgilar taqsimotini tanlamagan. The paper was criticized by Corfe and Butler (2006) who found the authors could not support their conclusions statistically. May va boshq. agreed that the statistical support was weak, but added that it is also weak for the alternative scenarios.[141]
Joriy kladistik tahlillar do not support Paul's hypothesis about the position of Arxeopteriks. Instead, they indicate that Arxeopteriks is closer to birds, within the clade Avialae, than it is to deinonychosaurs or oviraptorosaurs. However, some fossils support the version of this theory that holds that some non-flying carnivorous dinosaurs may have had flying ancestors. Jumladan, Mikroraptor, Pedopenna va Anchiornis all have winged feet, share many features, and lie close to the base of the clade Parafonlar. This suggests that the ancestral paravian was a four-winged glider, and that larger Deinonychosaurs secondarily lost the ability to glide, while the bird lineage increased in aerodynamic ability as it progressed.[2] Deinonychus may also display partial volancy, with the young being capable of flight or gliding and the adults being flightless.[142] In 2018, a study concluded that the last common ancestor of the Pennaraptora had joint surfaces on the fingers, and between the metatarsus and the wrist, that were optimised to stabilise the hand in flight. This was seen as an indication for secondary flightlessness in heavy basal members of that group.[143]
Yilda Euornithes, the earliest unequivocal example of secondary flightlessness is Patagopteryx.[144]
Shuningdek qarang
![]() Dinozavrlar portali
Dinozavrlar portali
Izohlar
- ^ Chiappe, Luis M. (2009). "Downsized Dinosaurs: The Evolutionary Transition to Modern Birds". Evolyutsiya: Ta'lim va targ'ibot. 2 (2): 248–256. doi:10.1007/s12052-009-0133-4.
- ^ a b Witmer, LM (2009). "Palaeontology: Feathered dinosaurs in a tangle". Tabiat. 461 (7264): 601–2. Bibcode:2009Natur.461..601W. doi:10.1038/461601a. PMID 19794481. S2CID 205049989.
- ^ Darwin, Charles R. (1859). Tabiat seleksiyasi vositasida turlarning kelib chiqishi yoki hayot uchun kurashda qulay irqlarni saqlab qolish to'g'risida. London: Jon Myurrey. p. 502pp. ISBN 978-1-4353-9386-8.
- ^ von Meyer, C.E. Hermann. (1861). "Arxeopteriks litografikasi (Vogel-Feder) und Pterodaktil von Solnhofen". Neues Jahrbuch für Mineralogie, Geologie und Paläontologie (nemis tilida). 1861: 678–679.
- ^ Ouen, Richard. (1863). "Ustida Arxeopteriks [sp] of von Meyer, with a description of the fossil remains of a long-tailed species, from the lithographic stone of Solenhofen [sp]". London Qirollik Jamiyatining falsafiy operatsiyalari. 153: 33–47. doi:10.1098/rstl.1863.0003.
- ^ Gegenbaur, K. (1863). "Vergleichend-anatomische Bemerkungen über das Fußskelet der Vögel". Archiv für Anatomie, Physiologie und Wissenschaftliche Medicin. 1863: 450–472.
- ^ Cope, E.D. (1867). "An account of the extinct reptiles which approached the birds". Filadelfiya Tabiiy fanlar akademiyasi materiallari. 19: 234–235.havola
- ^ Xaksli, T.H. (1868). "Qushlar va sudralib yuruvchilar o'rtasida deyarli oraliq bo'lgan hayvonlarga". Tabiiy tarix yilnomalari va jurnali. 4-chi. 2: 66–75.
- ^ Huxley, Thomas H. (1870). "Further evidence of the affinity between the dinosaurian reptiles and birds". London Geologiya Jamiyatining har choraklik jurnali. 26 (1–2): 12–31. doi:10.1144/GSL.JGS.1870.026.01-02.08. S2CID 129125487.
- ^ Nopcsa, Franz. (1907). "Ideas on the origin of flight". London zoologik jamiyati materiallari: 223–238.
- ^ Seeley, Harry G. (1901). Dragons of the Air: An Account of Extinct Flying Reptiles. London: Methuen & Co. p. 239 pp.
- ^ Abel, O (1911). "Die Vorfahren der Vögel und ihre Lebensweise". Viyandagi Verhandlungen der Zoologisch-Botanischen Gesellschaft. 61: 144–191.
- ^ Nieuwland, Ilja J.J. (2004). "Gerhard Heilmann and the artist's eye in science, 1912-1927" (PDF). PalArchning umurtqali hayvonlar paleontologiyasi jurnali. 3 (2).
- ^ Heilmann, Gerhard (1926). Qushlarning kelib chiqishi. London: Witherby. p.208 pp. ISBN 978-0-486-22784-9.
- ^ a b v Padian, Kevin. (2004). "Bazal Avialae". Yilda Vayshampel, Devid B.; Dodson, Piter; Osmolska, Xalska (tahr.) Dinozavrlar (Ikkinchi nashr). Berkli: Kaliforniya universiteti matbuoti. 210-231 betlar. ISBN 978-0-520-24209-8.
- ^ For example in 1923, three years before Heilmans's book, Roy Chapman Endryus found a good Oviraptor qazib olish Mo'g'uliston, lekin Genri Feyrfild Osborn, who analyzed the fossil in 1924, misidentified the furcula as an interklavikula; tasvirlangan Pol, G.S. (2002). Havoning dinozavrlari: evolyutsiyasi va dinozavrlar va qushlarda parvoz yo'qolishi. JHU Press. ISBN 978-0-8018-6763-7.
- ^ Camp, Charles L. (1936). "A new type of small theropod dinosaur from the Navajo Sandstone of Arizona". Bulletin of the University of California Department of Geological Sciences. 24: 39–65.
- ^ In Oviraptor: Barsbold, R. (1983). "Carnivorous dinosaurs from the Cretaceous of Mongolia". Trudy Soumestnaya Sovetsko-Mongol'skaya Paleontogicheskaya Ekspeditsiya (rus tilida). 19: 1–117. See the summary and pictures at "A wish for Koelofiz". Arxivlandi asl nusxasi 2008 yil 3 fevralda.
- ^ Lipkin, C., Sereno, P.C., and Horner, J.R. (November 2007). "The Furcula In Suchomimus Tenerensis And Tyrannosaurus Rex (Dinosauria: Theropoda: Tetanurae)". Paleontologiya jurnali. 81 (6): 1523–1527. doi:10.1666/06-024.1. S2CID 86234363.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola) - full text currently online at "The Furcula in Suchomimus Tenerensis va Tyrannosaurus rex". 2007-11-11. Arxivlandi asl nusxasi 2011-05-22. Olingan 2008-04-17. This lists a large number of theropods in which furculae have been found, as well as describing those of Suchomimus Tenerensis va Tyrannosaurus rex.
- ^ Carrano, M.R., Hutchinson, J.R., and Sampson, S.D. (2005 yil dekabr). "Yangi ma'lumotlar Segisaurus halli, a small theropod dinosaur from the Early Jurassic of Arizona" (PDF). Umurtqali hayvonlar paleontologiyasi jurnali. 25 (4): 835–849. doi:10.1671/0272-4634(2005)025[0835:NIOSHA]2.0.CO;2. ISSN 0272-4634. Arxivlandi asl nusxasi (PDF) 2006-05-12 kunlari.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Yeyts, Adam M.; and Vasconcelos, Cecilio C. (2005). "Furcula-like clavicles in the prosauropod dinosaur Massospondil". Umurtqali hayvonlar paleontologiyasi jurnali. 25 (2): 466–468. doi:10.1671/0272-4634(2005)025[0466:FCITPD]2.0.CO;2. ISSN 0272-4634.
- ^ Ostrom, Jon H. (1969). "Osteologiyasi Deinonychus antirrhopus, Montananing quyi bo'ridan g'ayrioddiy teropod ". Peabody Tabiat tarixi muzeyi xabarnomasi. 30: 1–165.
- ^ Ostrom, Jon H. (1970). "Arxeopteriks: Notice of a "new" specimen". Ilm-fan. 170 (3957): 537–8. Bibcode:1970Sci...170..537O. doi:10.1126/science.170.3957.537. PMID 17799709. S2CID 46682559.
- ^ Palatalar, Pol (2002). Bones of Contention: The Archaeopteryx Scandals. London: John Murray Ltd. pp. 183–184. ISBN 978-0-7195-6054-5.
- ^ Walker, Alick D. (1972). "New light on the origin of birds and crocodiles". Tabiat. 237 (5353): 257–263. Bibcode:1972Natur.237..257W. doi:10.1038/237257a0. S2CID 4174690.
- ^ Ostrom, Jon H. (1973). "Qushlarning ajdodi". Tabiat. 242 (5393): 136. Bibcode:1973NPhS..242..136O. doi:10.1038 / 242136a0. S2CID 29873831.
- ^ Ostrom, Jon H. (1975). The origin of birds. Yer va sayyora fanlari bo'yicha yillik sharh. 3. pp. 55–77. Bibcode:1975AREPS...3...55O. doi:10.1146/annurev.ea.03.050175.000415. ISBN 978-0-912532-57-8.
- ^ Ostrom, Jon H. (1976). "Arxeopteriks and the origin of birds". Linnean Jamiyatining Biologik jurnali. 8 (2): 91–182. doi:10.1111 / j.1095-8312.1976.tb00244.x.
- ^ Bakker, Robert T. (1972). "Anatomical and ecological evidence of endothermy in dinosaurs". Tabiat. 238 (5359): 81–85. Bibcode:1972Natur.238...81B. doi:10.1038/238081a0. S2CID 4176132.
- ^ Horner, Jon R.; Makela, Robert (1979). "Voyaga etmaganlar uyasi dinozavrlar orasida oila tuzilishini tasdiqlaydi". Tabiat. 282 (5736): 296–298. Bibcode:1979 yil 28-iyun ... 296H. doi:10.1038 / 282296a0. S2CID 4370793.
- ^ Hennig, E.H. Villi (1966). Filogenetik sistematikasi. translated by Davis, D. Dwight; & Zangerl, Rainer. Urbana: Illinoys universiteti matbuoti. ISBN 978-0-252-06814-0.
- ^ a b Gauthier, Jacques. (1986). "Saurischian monofilligi va qushlarning kelib chiqishi". In Padian, Kevin (ed.). Qushlarning kelib chiqishi va parvoz evolyutsiyasi. Kaliforniya Fanlar akademiyasining xotiralari 8. 1-55 betlar.
- ^ a b v d Senter, Fil (2007). "Coelurosauria (Dinosauria: Theropoda) filogeniyasiga yangi ko'rinish". Tizimli paleontologiya jurnali. 5 (4): 429–463. doi:10.1017 / S1477201907002143. S2CID 83726237.
- ^ a b v Tyorner, Alan X.; Hwang, Sunny; & Norell, Mark A. (2007). "A small derived theropod from Öösh, Early Cretaceous, Baykhangor, Mongolia". Amerika muzeyi Novitates. 3557 (1): 1–27. doi:10.1206 / 0003-0082 (2007) 3557 [1: ASDTFS] 2.0.CO; 2. hdl:2246/5845. ISSN 0003-0082.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Sereno, Paul C.; Rao Chenggang (1992). "Early evolution of avian flight and perching: new evidence from the Lower Cretaceous of China". Ilm-fan. 255 (5046): 845–8. Bibcode:1992Sci...255..845S. doi:10.1126 / science.255.5046.845. PMID 17756432. S2CID 41106677.
- ^ Hou Lian-Hai, Lian-hai; Chjou Chxun; Martin, Larry D.; & Feduccia, Alan (1995). "A beaked bird from the Jurassic of China". Tabiat. 377 (6550): 616–618. Bibcode:1995Natur.377..616H. doi:10.1038/377616a0. S2CID 4357707.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Ji Qiang; Ji Shu-an (1996). "On the discovery of the earliest bird fossil in China and the origin of birds" (PDF). Xitoy geologiyasi. 233: 30–33.
- ^ Chen Pei-ji, Pei-ji; Dong Zhiming; & Zhen Shuo-nan. (1998). "An exceptionally preserved theropod dinosaur from the Yixian Formation of China". Tabiat. 391 (6663): 147–152. Bibcode:1998Natur.391..147C. doi:10.1038/34356. S2CID 4430927.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Lingham-Soliar, Theagarten; Feduccia, Alan; & Wang Xiaolin. (2007). "A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres". Qirollik jamiyati materiallari B. 274 (1620): 1823–9. doi:10.1098/rspb.2007.0352. PMC 2270928. PMID 17521978.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ a b Ji Qiang, Philip J.; Currie, Filipp J.; Norell, Mark A .; & Ji Shu-an. (1998). "Two feathered dinosaurs from northeastern China" (PDF). Tabiat. 393 (6687): 753–761. Bibcode:1998Natur.393..753Q. doi:10.1038/31635. S2CID 205001388.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Sloan, Christopher P. (1999). "Feathers for T. rex?". National Geographic. 196 (5): 98–107. Arxivlandi asl nusxasi 2012-10-11.
- ^ Monastersky, Richard (2000). "All mixed up over birds and dinosaurs". Fan yangiliklari. 157 (3): 38. doi:10.2307/4012298. JSTOR 4012298. Arxivlandi asl nusxasi 2008-10-03 kunlari. Olingan 2008-07-04.
- ^ Xu Xing, Xing; Tang Zhi-lu; & Wang Xiaolin. (1999). "A therizinosaurid dinosaur with integumentary structures from China". Tabiat. 399 (6734): 350–354. Bibcode:1999 natur.399..350X. doi:10.1038/20670. S2CID 204993327.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Xu Xing, X; Norell, Mark A .; Kuang Xuewen; Wang Xiaolin; Zhao Qi; & Jia Chengkai. (2004). "Xitoydan kelgan bazal tirannosauroidlar va tirannosauroidlardagi protofeatherlarga dalillar". Tabiat. 431 (7009): 680–4. Bibcode:2004 yil natur.431..680X. doi:10.1038 / tabiat02855. PMID 15470426. S2CID 4381777.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Xu Xing, X; Chjou Chxun; Wang Xiaolin; Kuang Xuewen; Chjan Fucheng; & Du Xiangke (2003). "Xitoydan kelgan to'rt qanotli dinozavrlar". Tabiat. 421 (6921): 335–40. Bibcode:2003Natur.421..335X. doi:10.1038 / tabiat01342. PMID 12540892. S2CID 1160118.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Chjou, Chxun; Chjan Fucheng (2002). "Xitoyning erta bo'ridan uzun dumli, urug 'yeyuvchi qush". Tabiat. 418 (6896): 405–9. Bibcode:2002Natur.418..405Z. doi:10.1038 / nature00930. PMID 12140555. S2CID 4423299.
- ^ a b Chatterjee, Sankar (17 April 1998). "Counting the Fingers of Birds and Dinosaurs". Ilm-fan. 280 (5362): 355a–355. Bibcode:1998Sci...280..355C. doi:10.1126/science.280.5362.355a.
- ^ Merilend universiteti geologiya bo'limi bosh sahifasi, "Theropoda I" yoqilgan Avetheropoda, 2006 yil 14-iyul.
- ^ Vagner, G. P.; Gautthier, J. A. (1999). "1,2,3 = 2,3,4: A solution to the problem of the homology of the digits in the avian hand". Proc. Natl. Akad. Ilmiy ish. AQSH. 96 (9): 5111–6. Bibcode:1999 PNAS ... 96.5111W. doi:10.1073 / pnas.96.9.5111. PMC 21825. PMID 10220427.
- ^ Scienceblogs: Limuzavr is awesome Arxivlandi 2009-06-22 da Orqaga qaytish mashinasi.
- ^ Developmental Biology 8e Online. Chapter 16: Did Birds Evolve From the Dinosaurs? Arxivlandi 2008-01-18 da Orqaga qaytish mashinasi
- ^ Vargas AO, Wagner GP and Gauthier, JA. 2009. Limusaurus and bird digit identity. Available from Nature Precedings [1]
- ^ Havstad, Joyce C.; Smith, N. Adam (2019). "Fossils with Feathers and Philosophy of Science". Tizimli biologiya. 68 (5): 840–851. doi:10.1093/sysbio/syz010. ISSN 1076-836X. PMC 6701454. PMID 30753719.
- ^ Martin, Larry D. (2004). "A basal archosaurian origin for birds". Acta Zoologica Sinica. 50 (6): 978–990.
- ^ Feduccia, Alan; Lingham-Soliar, Theagarten; & Hincliffe, J. Richard. (2005). "Tukli dinozavrlar mavjudmi? Gipotezani neontologik va paleontologik dalillarda tekshirish". Morfologiya jurnali. 266 (2): 125–66. doi:10.1002 / jmor.10382. PMID 16217748. S2CID 15079072.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Burke, Ann C.; Feduccia, Alan (1997). "Rivojlanish naqshlari va qushlar qo'lidagi homologiyalarni aniqlash". Ilm-fan. 278 (5338): 666–668. Bibcode:1997 yil ... 278..666B. doi:10.1126/science.278.5338.666.
- ^ Newman SA (2011). "Thermogenesis, muscle hyperplasia, and the origin of birds". BioEssays. 33 (9): 653–656. doi:10.1002/bies.201100061. PMID 21695679. S2CID 42012034.
- ^ Newman SA, Mezentseva NV, Badyaev AV (2013). "Gene loss, thermogenesis, and the origin of birds". Nyu-York Fanlar akademiyasining yilnomalari. 1289 (1): 36–47. Bibcode:2013NYASA1289...36N. doi:10.1111/nyas.12090. PMID 23550607.
- ^ Mezentseva NV, Kumaratilake JS, Newman SA (2008). "The brown adipocyte differentiation pathway in birds: An evolutionary road not taken". BMC biologiyasi. 6 (1): 17. doi:10.1186/1741-7007-6-17. PMC 2375860. PMID 18426587.
- ^ Sereno, Paul C. (1997). "Dinozavrlarning kelib chiqishi va evolyutsiyasi". Yer va sayyora fanlari bo'yicha yillik sharh. 25 (1): 435–489. Bibcode:1997AREPS..25..435S. doi:10.1146/annurev.earth.25.1.435.
- ^ Chiappe, Luis M. (1997). "Aves". Yilda Currie, Filipp J.; Padian, Kevin (eds.). Dinozavrlar entsiklopediyasi. San-Diego: Akademik matbuot. pp. 45–50. ISBN 978-0-12-226810-6.
- ^ a b Holtz, Thomas R.; Osmólska, Halszka. (2004). "Saurischia". Yilda Vayshampel, Devid B.; Dodson, Piter; Osmolska, Xalska (tahr.) Dinozavrlar (Ikkinchi nashr). Berkli: Kaliforniya universiteti matbuoti. pp. 21–24. ISBN 978-0-520-24209-8.
- ^ a b Tyorner, Alan X.; Pol, Diego; Klark, Julia A .; Erikson, Gregori M.; & Norell, Mark A. (2007). "A basal dromaeosaurid and size evolution preceding avian flight". Ilm-fan. 317 (5843): 1378–81. Bibcode:2007Sci...317.1378T. doi:10.1126/science.1144066. PMID 17823350.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Osmólska, Halszka; Maryańska, Teresa; & Wolsan, Mieczysław. (2002). "Avialan status for Oviraptorosauria". Acta Palaeontologica Polonica. 47 (1): 97–116.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Martinelli, Agustín G.; Vera, Ezequiel I. (2007). "Achillesaurus manazzonei, a new alvarezsaurid theropod (Dinosauria) from the Late Cretaceous Bajo de la Carpa Formation, Río Negro Province, Argentina" (PDF). Zootaxa. 1582: 1–17. doi:10.11646/zootaxa.1582.1.1.
- ^ Novas, Fernando E.; Pol, Diego. (2002). "Alvarezsaurid relationships reconsidered". Chiappe shahrida Luis M.; Witmer, Lourens M. (tahr.). Mezozoy qushlari: Dinozavrlarning boshlari ustida. Berkli: Kaliforniya universiteti matbuoti. 121-125 betlar. ISBN 978-0-520-20094-4.
- ^ Sereno, Paul C. (1999). "Dinozavrlarning evolyutsiyasi". Ilm-fan. 284 (5423): 2137–47. doi:10.1126 / science.284.5423.2137. PMID 10381873.
- ^ Perle, Altangerel; Norell, Mark A .; Chiappe, Luis M.; & Clark, James M. (1993). "Mo'g'uliston bo'ridan uchib ketmaydigan qush". Tabiat. 362 (6421): 623–626. Bibcode:1993Natur.362..623A. doi:10.1038 / 362623a0. S2CID 4252852.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Chiappe, Luis M.; Norell, Mark A .; & Clark, James M. (2002). "The Cretaceous, short-armed Alvarezsauridae: Mononykus and its kin". In Chiappe, Luis M.; Witmer, Lawrence M. (eds.). Mezozoy qushlari: Dinozavrlarning boshlari ustida. Berkli: Kaliforniya universiteti matbuoti. 87–119-betlar. ISBN 978-0-520-20094-4.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Forster, Ketrin A.; Sampson, Skott D.; Chiappe, Luis M.; & Krause, David W. (1998). "Qushlarning theropod ajdodi: Madagaskarning so'nggi bo'ridan yangi dalillar". Ilm-fan. 279 (5358): 1915–9. Bibcode:1998 yil ... 279.1915F. doi:10.1126 / science.279.5358.1915. PMID 9506938.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Makovicky, Piter J.; Apestegiya, Sebastyan; & Agnolín, Federico L. (2005). "Janubiy Amerikadan eng qadimgi dromaeosaurid terropodi". Tabiat. 437 (7061): 1007–11. Bibcode:2005 yil 53-iyun, 1997 yil. doi:10.1038 / nature03996. PMID 16222297. S2CID 27078534.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Pol, Gregori S. (2002). Havoning dinozavrlari: evolyutsiyasi va dinozavrlar va qushlarda parvoz yo'qolishi. Baltimor: Jons Xopkins universiteti matbuoti. p.472pp. ISBN 978-0-8018-6763-7.
- ^ a b Mayr, Jerald; Pohl, Burkhard; & Peters, D. Stefan (2005). "Yaxshi saqlanib qolgan Arxeopteriks teropod xususiyatlariga ega namuna ". Ilm-fan. 310 (5753): 1483–6. Bibcode:2005Sci...310.1483M. doi:10.1126/science.1120331. PMID 16322455. S2CID 28611454.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Chatterjee, Immoor; L. Immoor (9 September 2005). "The Dinosaurs of the Jurassic Park Movies". Geolor.com. Arxivlandi asl nusxasi 2008 yil 12 mayda. Olingan 23 iyun, 2007.
- ^ Wellnhofer, P. (1988). "Ein neuer Exemplar von Arxeopteriks". Arxeopteriks. 6: 1–30.
- ^ Xu X, Norell MA, Kuang X, Wang X, Zhao Q, Jia C (October 2004). "Xitoydan kelgan bazal tirannosauroidlar va tirannosauroidlardagi protofeatherlarga dalillar". Tabiat. 431 (7009): 680–4. Bibcode:2004 yil natur.431..680X. doi:10.1038 / tabiat02855. PMID 15470426. S2CID 4381777.
- ^ Feduccia, A. (2012). Riddle of the Feathered Dragons: Hidden Birds of China. Yale University Press, ISBN 0-300-16435-1, ISBN 978-0-300-16435-0
- ^ Chjan, F.; Kearns, S.L.; Orr, PJ .; Benton, M.J .; Chjou, Z.; Jonson, D.; Xu, X.; and Wang, X. (2010). "Fosilizatsiya qilingan melanosomalar va bo'r dinozavrlari va qushlarning rangi" (PDF). Tabiat. 463 (7284): 1075–1078. Bibcode:2010 yil natur.463.1075Z. doi:10.1038/nature08740. PMID 20107440. S2CID 205219587.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Foth, C (2012). "On the identification of feather structures in stem-line representatives of birds: evidence from fossils and actuopalaeontology". Paläontologische Zeitschrift. 86: 91–102. doi:10.1007/s12542-011-0111-3. S2CID 86362907.
- ^ Currie, PJ .; Chen, P.-j. (2001). "Anatomiya Sinosauropteryx prima from Liaoning, northeastern China". Kanada Yer fanlari jurnali. 38 (1): 705–727. Bibcode:2001CaJES..38.1705C. doi:10.1139/cjes-38-12-1705.
- ^ Bhullar, B. A. S.; Marugan-Lobon, J. S .; Rakimo, F.; Bever, G. S .; Rou, T. B.; Norell, M. A .; Abzhanov, A. (2012). "Birds have paedomorphic dinosaur skulls". Tabiat. 487 (7406): 223–226. Bibcode:2012Natur.487..223B. doi:10.1038/nature11146. PMID 22722850. S2CID 4370675.
- ^ O'Konnor, PM; Claessens, L.P.A.M. (2005). "Qushlarga tegishli bo'lmagan teropod dinozavrlarda asosiy o'pka dizayni va oqimli shamollatish". Tabiat. 436 (7048): 253–6. Bibcode:2005 yil Noyabr.436..253O. doi:10.1038 / nature03716. PMID 16015329. S2CID 4390587.
- ^ Paul C. Sereno, Ricardo N. Martinez, Jeffrey A. Wilson, David J. Varricchio, Oscar A. Alcober, Hans C. E. Larsson (2008). Kemp, Tom (ed.). "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina". PLOS ONE. 3 (9): e3303. Bibcode:2008PLoSO...3.3303S. doi:10.1371/journal.pone.0003303. PMC 2553519. PMID 18825273.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Fisher, P. E.; Rassel, D. A .; Stoskopf, M. K.; Barrick, R. E.; Hammer, M.; Kuzmitz, A. A. (2000). "Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur". Ilm-fan. 288 (5465): 503–5. Bibcode:2000Sci...288..503F. doi:10.1126/science.288.5465.503. PMID 10775107.
- ^ Hillenius, W. J.; Ruben, J. A. (2004). "The evolution of endothermy in terrestrial vertebrates: Who? when? why?". Fiziologik va biokimyoviy zoologiya. 77 (6): 1019–42. doi:10.1086/425185. PMID 15674773.
- ^ Rou, T .; McBride, E. F.; Sereno, P. C .; Rassel, D. A .; Fisher, P. E.; Barrick, R. E.; Stoskopf, M. K. (2001). "Dinosaur with a Heart of Stone". Ilm-fan. 291 (5505): 783. doi:10.1126/science.291.5505.783a. PMID 11157158.
- ^ a b Cleland, Timothy P.; Stoskopf, Michael K.; and Schweitzer, Mary H. (2011). "Histological, chemical, and morphological reexamination of the "heart" of a small Late Cretaceous Felsevra". Naturwissenschaften. 98 (3): 203–211. Bibcode:2011NW.....98..203C. doi:10.1007/s00114-010-0760-1. PMID 21279321. S2CID 2408562.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Chinsamy, Anusuya; and Hillenius, Willem J. (2004). "Physiology of nonavian dinosaurs". Dinozavrlar, 2-chi. 643–659.
- ^ Xu, X.; Norell, MA (2004). "A new troodontid dinosaur from China with avian-like sleeping posture". Tabiat. 431 (7010): 838–41. Bibcode:2004Natur.431..838X. doi:10.1038/nature02898. PMID 15483610. S2CID 4362745. See commentary on the article
- ^ Shveytsar, M.H .; Wittmeyer, J.L .; and Horner, J.R. (2005). "Gender-specific reproductive tissue in ratites and Tyrannosaurus rex". Ilm-fan. 308 (5727): 1456–60. Bibcode:2005Sci...308.1456S. doi:10.1126/science.1112158. PMID 15933198. S2CID 30264554.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Lee, Andrew H.; Werning, Sarah (2008). "Sexual maturity in growing dinosaurs does not fit reptilian growth models". Milliy fanlar akademiyasi materiallari. 105 (2): 582–7. Bibcode:2008PNAS..105..582L. doi:10.1073/pnas.0708903105. PMC 2206579. PMID 18195356.
- ^ Norell, M. A., Clark, J. M., Dashzeveg, D., Barsbold, T., Chiappe, L. M., Davidson, A. R., McKenna, M. C. and Novacek, M. J. (November 1994). "A theropod dinosaur embryo and the affinities of the Flaming Cliffs Dinosaur eggs". Ilm-fan. 266 (5186): 779–82. Bibcode:1994Sci ... 266..779N. doi:10.1126/science.266.5186.779. PMID 17730398. S2CID 22333224.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Wings O (2007). "Gastrolit funktsiyasini qazib olingan umurtqali hayvonlar uchun ta'siri va qayta ko'rib chiqilgan tasnifi". Paleontologica Polonica. 52 (1): 1–16.
- ^ Embery, G; Milner, AC; Waddington, RJ; Hall, RC; Langley, MS; Milan, AM (2003). "Identification of proteinaceous material in the bone of the dinosaur Iguanodon". Birlashtiruvchi to'qimalarni tadqiq qilish. 44 (Suppl 1): 41–6. doi:10.1080/03008200390152070. PMID 12952172. S2CID 2249126.
- ^ Schweitzer, MH; Marshall, M; Carron, K; Bohle, DS; Busse, SC; Arnold, EV; Barnard, D; Horner, JR; Starkey, JR (June 1997). "Heme compounds in dinosaur trabecular bone". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 94 (12): 6291–6. Bibcode:1997PNAS...94.6291S. doi:10.1073/pnas.94.12.6291. PMC 21042. PMID 9177210.
- ^ Schweitzer, MH; Wittmeyer, JL; Horner, JR; Toporski, JK (2005). "Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex". Ilm-fan. 307 (5717): 1952–5. Bibcode:2005 yil ... 307.1952 yil. doi:10.1126 / science.1108397. PMID 15790853. S2CID 30456613. Also covers the Reproduction Biology paragraph in the Feathered dinosaurs and the bird connection section.
- ^ Asara, JM; Schweitzer MH, Freimark LM, Phillips M, Cantley LC (2007). "Protein Sequences from Mastodon and Tyrannosaurus Rex Revealed by Mass Spectrometry". Ilm-fan. 316 (5822): 280–5. Bibcode:2007Sci...316..280A. doi:10.1126/science.1137614. PMID 17431180. S2CID 85299309.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Organ, CL; Schweitzer, MH; Chjen, Vt; Freimark, LM; Cantley, LC; Asara, JM (2008). "Molecular phylogenetics of mastodon and Tyrannosaurus rex". Ilm-fan. 320 (5875): 499. doi:10.1126/science.1154284. PMID 18436782. S2CID 24971064.
- ^ Schweitzer, M. H.; Zheng W., Organ C. L., Avci R., Suo Z., Freimark L. M., Lebleu V. S., Duncan M. B., Vander Heiden M. G., Neveu J. M., Lane W. S., Cottrell J. S., Horner J. R., Cantley L. C., Kalluri R. & Asara J. M. (2009). "Biomolecular Characterization and Protein Sequences of the Campanian Hadrosaur B. canadensis". Ilm-fan. 324 (5927): 626–31. Bibcode:2009Sci...324..626S. doi:10.1126/science.1165069. PMID 19407199. S2CID 5358680.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Kaye, TG; Gaugler, G; Sawlowicz, Z (2008). "Dinosaurian soft tissues interpreted as bacterial biofilms". PLOS ONE. 3 (7): e2808. Bibcode:2008PLoSO...3.2808K. doi:10.1371/journal.pone.0002808. PMC 2483347. PMID 18665236.
- ^ Peterson, JE; Lenczewski, ME; Scherer, RP (2010). "Influence of microbial biofilms on the preservation of primary soft tissue in fossil and extant archosaurs". PLOS ONE. 5 (10): e13334. Bibcode:2010PLoSO...513334P. doi:10.1371/journal.pone.0013334. PMC 2953520. PMID 20967227.
- ^ Bern, M; Phinney, BS; Goldberg, D (2009). "Reanalysis of Tyrannosaurus rex mass spectra". Proteom tadqiqotlari jurnali. 8 (9): 4328–4332. doi:10.1021/pr900349r. PMC 2738754. PMID 19603827.
- ^ Cleland, TP; va boshq. (2015). "Mass spectrometry and antibody-based characterization of blood vessels from Brachylophosaurus canadensis". Proteom tadqiqotlari jurnali. 14 (12): 5252–5262. doi:10.1021/acs.jproteome.5b00675. PMC 4768904. PMID 26595531.
- ^ Bakli, Maykl; Warwood, Stacey; van Dongen, Bart; Kitchener, Endryu S.; Manning, Phillip L. (2017). "A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination". Qirollik jamiyati materiallari B. 284 (1855): 20170544. doi:10.1098/rspb.2017.0544. PMC 5454271. PMID 28566488.
- ^ Wang, HL; Yan, ZY; Jin, DY (1997). "Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil". Molekulyar biologiya va evolyutsiya. 14 (5): 589–91. doi:10.1093/oxfordjournals.molbev.a025796. PMID 9159936.
- ^ Veten, Dennis F.A.E.; va boshq. (2018 yil 13 mart). "Qanot suyagi geometriyasi Arxeopteriksda faol parvozni ochib beradi". Tabiat aloqalari. 9 (923): 923. Bibcode:2018NatCo ... 9..923V. doi:10.1038 / s41467-018-03296-8. PMC 5849612. PMID 29535376.
- ^ Guarino, Ben (2018 yil 13 mart). "Bu tukli dinozavr uchib ketgandir, lekin siz bilgan qushlar kabi emas". Washington Post. Olingan 13 mart 2018.
- ^ Terres, Jon K. (1980). Shimoliy Amerika qushlari Audubon Jamiyati Entsiklopediyasi. Nyu-York, NY: Knopf. pp.398–401. ISBN 978-0-394-46651-4.
- ^ Poling, J. (1996). "Feathers, scutes and the origin of birds". dinosauria.com. Arxivlandi asl nusxasi 2008-04-02. Olingan 2008-03-21.
- ^ Prum, R. & Brush, A.H. (2002). "The evolutionary origin and diversification of feathers" (PDF). Biologiyaning choraklik sharhi. 77 (3): 261–95. doi:10.1086/341993. PMID 12365352. Arxivlandi asl nusxasi (PDF) on 2003-10-15. Olingan 2018-09-17.
- ^ Mayr G, Pohl B, Peters D (2005). "Yaxshi saqlanib qolgan Arxeopteriks teropod xususiyatlariga ega namuna ". Ilm-fan. 310 (5753): 1483–6. Bibcode:2005Sci...310.1483M. doi:10.1126/science.1120331. PMID 16322455. S2CID 28611454.
- ^ Feduccia, A. (1999). Qushlarning kelib chiqishi va evolyutsiyasi. Yel universiteti matbuoti. ISBN 978-0-300-07861-9.
- ^ Feduccia, A. (February 1995). "Explosive Evolution in Tertiary Birds and Mammals". Ilm-fan. 267 (5198): 637–8. Bibcode:1995 yil ... 267..637F. doi:10.1126 / science.267.5198.637. PMID 17745839. S2CID 42829066. Arxivlandi asl nusxasi 2008-03-20.
- ^ Feduccia, A. (1993).[belgilang ]
- ^ Bo'r tracks of a bird with a similar lifestyle have been found - Lockley, M.G., Li, R., Harris, J.D., Matsukawa, M., and Liu, M. (August 2007). "Eng qadimgi zigodaktil qushlarning oyoqlari: Bo'rt davridagi yo'l yo'ldoshiga o'xshash yo'llardan olingan dalillar". Naturwissenschaften. 94 (8): 657–65. Bibcode:2007NW ..... 94..657L. doi:10.1007 / s00114-007-0239-x. PMID 17387416. S2CID 15821251.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Burgers, P. & L. M. Chiappe (1999). "The wing of Archaeopteryx as a primary thrust generator". Tabiat. 399 (6731): 60–62. Bibcode:1999Natur.399...60B. doi:10.1038/19967. S2CID 4430686.
- ^ Cowen, R. Hayot tarixi. Blackwell Science. ISBN 978-0-7266-0287-0.
- ^ Videler, J.J. 2005: Avian Flight. Oksford universiteti. Press, Oksford.
- ^ Burke, A.C. & Feduccia, A. (1997). "Rivojlanish naqshlari va qushlar qo'lidagi homologiyalarni aniqlash". Ilm-fan. 278 (5338): 666–668. Bibcode:1997 yil ... 278..666B. doi:10.1126/science.278.5338.666. Xulosa qilingan "Embryo Studies Show Dinosaurs Could Not Have Given Rise To Modern Birds". ScienceDaily. 1997 yil oktyabr.
- ^ Chatterjee, S. (April 1998). "Counting the Fingers of Birds and Dinosaurs". Ilm-fan. 280 (5362): 355a–355. Bibcode:1998Sci...280..355C. doi:10.1126 / science.280.5362.355a.
- ^ Vargas, AO, Fallon, JF (2004 yil oktyabr). "Qushlarning dinozavr qanotlari bor: molekulyar dalillar". Eksperimental Zoologiya jurnali B qism: Molekulyar va rivojlanish evolyutsiyasi. 304B (1): 86–90. doi:10.1002 / jez.b.21023. PMID 15515040.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Pennisi, E. (2005 yil yanvar). "Qushlarning qanotlari haqiqatan ham dinozavrlarning qo'llariga o'xshaydi" (PDF). Ilm-fan. 307 (5707): 194b. doi:10.1126 / science.307.5707.194b. PMID 15653478. S2CID 82490156. Arxivlandi asl nusxasi (PDF) 2011-07-27 da.
- ^ Bu erda juda yosh jo'janing videoklipi bor "Qanot parvoz evolyutsiyasi va parvoz evolyutsiyasi".
- ^ Dial, K.P. (2003). "Qanot yordamida nishab yugurish va parvoz evolyutsiyasi". Ilm-fan. 299 (5605): 402–4. Bibcode:2003 yil ... 299..402D. doi:10.1126 / science.1078237. PMID 12532020. S2CID 40712093. Xulosa qilingan Morelle, Rebekka (2008 yil 24-yanvar). "Qushlarning uchish sirlari oshkor qilindi" (Internet). Olimlarning fikriga ko'ra, ular birinchi qushlarning qanday qilib havoga ko'tarilganligi sirini hal qilishda bir qadam yaqinlashishi mumkin. BBC yangiliklari. Olingan 2008-01-25.
- ^ Bundle, MW & Dial, K.P. (2003). "Qanotli nishab mexanikasi (WAIR)" (PDF). Eksperimental biologiya jurnali. 206 (Pt 24): 4553-64. doi:10.1242 / jeb.00673. PMID 14610039. S2CID 6323207.
- ^ a b Senter, P. (2006). "Tropodlarda va bazal qushlarda skapulyar yo'nalish va uchib yurishning kelib chiqishi". Acta Palaeontologica Polonica. 51 (2): 305–313.
- ^ Dececchi, T. Aleksandr; Larsson, Xans C. E. (2011). "Qushlarning o'tmishdagi daraxtlarga moslashishini baholash: parranda parvozining ekologik muhitini sinash". PLOS ONE. 6 (8): e22292. Bibcode:2011PLoSO ... 622292D. doi:10.1371 / journal.pone.0022292. PMC 3153453. PMID 21857918.
- ^ Chatterji, Sankar, Templin, R.J. (2004) "Xitoydan tukli koleurozavrlar: parranda parvozining arboreal kelib chiqishiga yangi yorug'lik" 251-281 betlar. Tukli ajdarlarda: Dinozavrlardan qushlarga o'tishga oid tadqiqotlar (P. J. Currie, E. B. Koppelhus, M. A. Shugar va J. L. Wright (tahr.). Indiana University Press, Bloomington.
- ^ Tarsitano, Samuel F.; Rassel, Entoni P.; Xorn, Frensis; Plummer, Kristofer; Millerchip, Karen (2000). "Aerodinamik va konstruktiv ko'rinish nuqtai nazaridan patlar evolyutsiyasi to'g'risida". Amerika zoologi. 40 (4): 676–686. doi:10.1093 / icb / 40.4.676.
- ^ Xu, D .; Xou, L .; Zhang, L. & Xu, X. (2009). "OldindanArxeopteriks metatarsda uzun patli Xitoydan troodontid theropod ". Tabiat. 461 (7264): 640–3. Bibcode:2009 yil natur.461..640H. doi:10.1038 / nature08322. PMID 19794491. S2CID 205218015.
- ^ Xopson, Jeyms A. "Qushlar va noavian tropod falanjiy nisbatlar ekomorfologiyasi: Qushlarning parrandaning quruqlikdan kelib chiqishiga qarshi ta'siri" (2001) Qushlarning kelib chiqishi va erta evolyutsiyasi bo'yicha yangi istiqbollardan: Jonning sharafiga bag'ishlangan xalqaro simpozium materiallari. H. Ostrom. J. Gautier va L. F. Gall, nashr. New Haven: Peabody Mus. Nat. Tarix., Yel Univ. ISBN 0-912532-57-2© 2001 Yabel universiteti Peabody tabiiy tarix muzeyi. Barcha huquqlar himoyalangan.
- ^ Glen, KL & Bennett, M.B. (2007 yil noyabr). "Mezozoy qushlari va parranda bo'lmagan teropodlarning yem rejimlari". Hozirgi biologiya. 17 (21): R911-2. doi:10.1016 / j.cub.2007.09.026. PMID 17983564. S2CID 535424. Arxivlandi asl nusxasi 2012-12-08.
- ^ Alonso, PD, Milner, AC, Ketcham, RA, Cokson, M.J va Rowe, TB. (2004 yil avgust). "Arxeopteriksning miyasi va ichki qulog'ining qush tabiati". Tabiat. 430 (7000): 666–9. Bibcode:2004 yil natur.430..666A. doi:10.1038 / tabiat02706. PMID 15295597. S2CID 4391019.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Chiappe, LM (2007-02-02). Shon-sharafli dinozavrlar: Qushlarning kelib chiqishi va erta rivojlanishi. Sidney: UNSW Press. ISBN 978-0-471-24723-4.
- ^ Chjan, F., Chjou, Z., Xu, X. va Vang, X. (2002). "Xitoydan kelgan voyaga etmagan koelurozavr teropodi daraxtlarga xos odatlardan dalolat beradi". Naturwissenschaften. 89 (9): 394–8. Bibcode:2002NW ..... 89..394Z. doi:10.1007 / s00114-002-0353-8. PMID 12435090. S2CID 556221.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ Chatterji, S; Templin, RJ (2007). "Mikroraptor gui patlarli dinozavrning ikki qanotli planformasi va parvoz ko'rsatkichlari" (PDF). Milliy fanlar akademiyasi materiallari. 104 (5): 1576–80. Bibcode:2007PNAS..104.1576C. doi:10.1073 / pnas.0609975104. PMC 1780066. PMID 17242354.
- ^ Beebe, C. W. A. (1915). "Qushlarning ajdodlarida tetrapteryks bosqichi". Zoologica. 2: 38–52.
- ^ Lukas, Frederik A. (Frederik Avgust); Amerika Tabiat tarixi muzeyi (2018 yil 13 iyun). O'tmishdagi hayvonlar: qadimgi dunyodagi ba'zi mavjudotlar haqida ma'lumot. Nyu-York: [Amer. Mus. Press] - Internet Arxivi orqali.
- ^ Pol, G.S. (2002). "Havoning dinozavrlari: evolyutsiyasi va dinozavrlar va qushlarda parvoz yo'qolishi". Baltimor: Jons Xopkins universiteti matbuoti. 257-bet
- ^ Mayr, G. (2006). "Izohga javob Tropod xususiyatlariga ega yaxshi saqlangan Arxeopteriks namunasi". Ilm-fan. 313 (5791): 1238c. Bibcode:2006 yil ... 313.1238 million. doi:10.1126 / science.1130964.
- ^ Korfe, I. J .; Butler, RJ (2006). "Izoh Teropod xususiyatlariga ega yaxshi saqlangan Arxeopteriks namunasi". Ilm-fan. 313 (5791): 1238b. Bibcode:2006 yil ... 313.1238C. doi:10.1126 / science.1130800. PMID 16946054. S2CID 19859320.
- ^ Parsons, Uilyam L.; Parsons, Kristen M. (2015). "Deinonychus antirrhopus (Theropoda, Dromaeosauridae) ontogenezidagi morfologik farqlar". PLOS ONE. 10 (4): e0121476. Bibcode:2015PLoSO..1021476P. doi:10.1371 / journal.pone.0121476. PMC 4398413. PMID 25875499.
- ^ Xutson, Djoel D.; Xutson, Kelda N. (2018). "Tuyaqushda parvozga moslashtirilgan parrandalar qo'shma majmuasini saqlash dinozavrlarda qanotlar qachon rivojlana boshlaganini aniqlashga yordam beradi". Tuyaqush: Afrika ornitologiyasi jurnali. 89 (2): 173–186. doi:10.2989/00306525.2017.1422566. S2CID 91081348.
- ^ Chiappe, Luis (2002-01-01), Patagoniyaning so'nggi bo'ridan (Argentina) parvozsiz Patagopteryx deferrariisi osteologiyasi, 281-316 betlar, ISBN 978-0-520-20094-4, olingan 2020-07-27
Adabiyotlar
- Barsbold, Rinchen (1983): O ptich'ikh chertakh v stroyenii khishchnykh dinozavrov. [Yirtqich dinozavrlar morfologiyasidagi "qush" xususiyatlari]. Qo'shma Sovet Mo'g'uliston Paleontologik ekspeditsiyasining operatsiyalari 24: 96-103. [Original maqola rus tilida.] V. Robert Uels tomonidan tarjima qilingan, nusxasi Kennet Carpenter tomonidan taqdim etilgan va Metyu Karrano tomonidan o'zgartirilgan. PDF to'liq matni
- Borenshteyn, Set (31-iyul, 2014-yil). "Dinozavrlar evolyutsiyasini dastlabki qushlarga o'rganish". AP yangiliklari. Olingan 3 avgust, 2014.
- Bostvik, Kimberli S (2003). "Qushlarning kelib chiqishi va evolyutsiyasi: ma'lumotlar to'planadi, olimlar birlashadi va shu bilan birga" munozaralar "hali ham davom etmoqda". Kladistika. 19 (4): 369–371. doi:10.1016 / S0748-3007 (03) 00069-0.
- Dingus, Louell va Rou, Timo'tiy (1997): Noto'g'ri yo'q bo'lib ketish: dinozavr evolyutsiyasi va qushlarning kelib chiqishi. W. H. Freeman and Company, Nyu-York. ISBN 0-7167-2944-X
- On-layn dinozavralar (1995): Arxeopteriksning zamonaviy qushlar bilan aloqasi. Qabul qilingan 2006-09-30.
- On-layn dinozavralar (1996): Dinozavr sinapomorfiyalari Arxeopteriks. Qabul qilingan 2006-09-30.
- Heilmann, G. (1926): Qushlarning kelib chiqishi. Viterby, London. ISBN 0-486-22784-7 (1972 yilgi Doverning qayta nashr etilishi)
- Mayr, Jerald; Pol, B .; Peters, D. S. (2005). "Yaxshi saqlanib qolgan Arxeopteriks Theropod xususiyatlari bilan namuna ". Ilm-fan. 310 (5753): 1483–1486. Bibcode:2005 yil ... 310.1483M. doi:10.1126 / science.1120331. PMID 16322455. S2CID 28611454.
- Olson, Storrs L. (1985): Qushlarning qoldiqlari. In: Farner, D.S .; King, JR va Parkes, Kennet C. (tahrir): Qushlar biologiyasi 8: 79-238. Academic Press, Nyu-York.
Tashqi havolalar
- "Bizning oramizdagi dinozavrlar" dinozavrlardan qushlarga evolyutsion yo'lni qaytaradi, NY Times, 2016 yil 28 mart
- DinoBuzz Dinozavr-qushlar gipotezasining mashhur darajadagi muhokamasi
- Arxeopteriks - Savol-javoblar Usenet yangiliklar guruhidan talk.origins.
- Bizning oramizdagi dinozavrlar Maqola va video Amerika tabiiy tarixi muzeyi qushlarga olib boruvchi dinozavr evolyutsiyasi ko'rgazmasi
| Vikimedia Commons-ga tegishli ommaviy axborot vositalari mavjud Qoldiqlarni Aves. |
| Fins |  | |
|---|---|---|
| Oyoq-qo'llar | ||
| Qanotlar | ||
| Evolyutsiya | ||
| Bog'liq | ||