Qushlarning ovozi - Bird vocalization

Qushlarning ovozi ikkalasini ham o'z ichiga oladi qush chaqiradi va qush qo'shiqlari. Texnik bo'lmagan foydalanishda, qush qo'shiqlar - bu inson qulog'iga ohangdor bo'lgan qush tovushlari. Yilda ornitologiya va qush, qo'shiqlar (nisbatan murakkab ovozlar) funktsiyalari bilan qo'ng'iroqlardan ajralib turadi (nisbatan sodda ovozlar).
Ta'rif
Qo'shiqlar va qo'ng'iroqlar o'rtasidagi farq murakkablik, uzunlik va kontekstga asoslangan. Qo'shiqlar uzoqroq va murakkabroq va ular bilan bog'liq hudud[3] va uchrashish va juftlik, qo'ng'iroqlar kabi funktsiyalarni bajarishga moyil signalizatsiya yoki a a'zolarini saqlash suruv aloqada.[4] Xovell va Uebb (1995) kabi boshqa vakolatlar funktsiyaga qarab farqlaydilar, shuning uchun kaptarlar singari qisqa ovozlar, hattoki noo'rin tovushlar, masalan, davul. qarag'aylar va "tanib olish "ning merganlar Ko'rgazmada uchadigan qanotlar, qo'shiqlar deb hisoblanadi.[5] Boshqalar esa qo'shiqni ta'riflaydigan takrorlanadigan va o'zgaruvchan naqshlarga o'xshash hece xilma-xilligi va vaqtinchalik muntazamlikni talab qiladi. musiqa. Odatda qushlar va ornitologiya bo'yicha qaysi tovushlar qo'shiq, qaysi qo'ng'iroqlar ekanligi to'g'risida kelishib olinadi va yaxshi dala qo'llanmasi ikkalasini farq qiladi.
Qushlarning qo'shig'i eng yaxshi rivojlangan buyurtma Passeriformes. Ba'zi guruhlar deyarli ovozsiz, faqat ishlab chiqaradilar zarbli va ritmik kabi tovushlar laylaklar, ularning hisob-kitoblarini chalg'itadigan. Ba'zi manakinlarda (Pipridae ), erkaklar mexanik ovoz ishlab chiqarishning bir qancha mexanizmlarini, shu jumladan mexanizmlarini rivojlantirdilar stridulyatsiya ba'zi hasharotlarda uchraydiganlardan farqli o'laroq emas.[6]
Qo'shiq odatda taniqli perchlardan etkazib beriladi, garchi ba'zi turlar uchayotganda kuylashlari mumkin. Tovushlarni ishlatishdan farqli o'laroq, mexanik vositalar yordamida tovushlarni chiqarish sirinx har xil atama qilingan cholg‘u musiqasi tomonidan Charlz Darvin, mexanik tovushlar[7] va yaqinda sonatsiya.[8] Atama sonat Bill, qanotlar, dum, oyoq va tana patlari kabi shprits bo'lmagan tuzilmalar yordamida ishlab chiqarilgan, ataylab modulyatsiya qilingan kommunikativ signallarga ega bo'lgan ovozli bo'lmagan tovushlarni ishlab chiqarish harakati sifatida belgilangan.[8]
Ekstratropikada Evroosiyo va Amerika qit'asi deyarli barcha qo'shiqlar erkak qushlar tomonidan ishlab chiqarilgan; ammo tropik mintaqalarda va ko'proq darajada cho'l kamarlari Avstraliya va Afrika ayollar uchun erkaklar singari ko'proq qo'shiq aytish odatlidir. Ushbu farqlar qadimdan ma'lum bo'lgan[9][10] va odatda Avstraliya va Afrikaning qurg'oqchil zonalarining odatdagidan va mavsumiy iqlimidan kelib chiqqan holda, qushlar har qanday sharoitda qulay bo'lgan vaqtda ko'paytirishni talab qiladi, garchi ular ko'p yillar davomida nasl berolmasa ham, oziq-ovqat ta'minoti hech qachon minimal darajadan oshmaydi.[9] Vaqti-vaqti bilan muntazam bo'lmagan naslchilik bilan har ikkala jins ham naslga keltirilishi kerak va vokalizatsiya, ayniqsa duet, shu maqsadga xizmat qiladi. Tropik mintaqalarda, Avstraliya va Janubiy Afrikada ayollarning vokalatsiyasining yuqori chastotasi juda past o'lim ko'rsatkichlari bilan bog'liq bo'lib, ular juftlikni bog'lash va hududiylikni kuchaytiradi.[11]
Anatomiya va fiziologiya
Qushlarning vokal a'zosi sirinx;[12] bu pastki qismida suyak tuzilishi traxeya (farqli o'laroq gırtlak ning yuqori qismida sutemizuvchi traxeya). Sirin va ba'zida atrofdagi havo yostig'i parranda havo o'tkazadigan membranalar tomonidan hosil bo'lgan tovush to'lqinlariga ta'sir qiladi. Qush pardani keskinligini o'zgartirib, balandlikni boshqaradi va ekshalatsiya kuchini o'zgartirib, balandlikni ham, hajmini ham boshqaradi. U traxeyaning ikki tomonini mustaqil ravishda boshqarishi mumkin, shuning uchun ba'zi turlar bir vaqtning o'zida ikkita nota hosil qilishi mumkin.
Funktsiya

Qushlar qo'shig'ining ikkita asosiy funktsiyasidan biri bu juftlikni jalb qilishdir.[14] Olimlar qushlar qo'shig'i orqali rivojlangan deb taxmin qilishmoqda jinsiy tanlov va tajribalar shuni ko'rsatadiki, qushlar qo'shig'ining sifati fitnessning yaxshi ko'rsatkichi bo'lishi mumkin.[15] Tajribalar shuni ko'rsatadiki, parazitlar va kasalliklar qo'shiqning tezligi kabi qo'shiq xususiyatlariga bevosita ta'sir ko'rsatishi mumkin, bu esa sog'liqning ishonchli ko'rsatkichlari bo'lib xizmat qiladi.[16][17] Qo'shiq repertuari, shuningdek, ba'zi turlarga mosligini ko'rsatadigan ko'rinadi.[18][19] Erkak qushlarni ushlab turish va reklama qilish qobiliyati hududlar qo'shiqdan foydalanish ularning jismoniy tayyorgarligini ham namoyish etadi. Shuning uchun, urg'ochi qush erkakni qo'shiqlari sifatiga va qo'shiq repertuarining hajmiga qarab tanlashi mumkin.
Qushlar qo'shig'ining ikkinchi asosiy vazifasi hududni himoya qilishdir.[14] Hududiy qushlar bir-biri bilan qo'shiq yordamida o'zaro ta'sir o'tkazib, hudud chegaralarini muhokama qilishadi. Qo'shiq sifatning ishonchli ko'rsatkichi bo'lishi mumkinligi sababli, shaxslar raqiblarining sifatini aniqlay olishlari va baquvvat ravishda qimmat kurashni oldini olishlari mumkin.[15] Qo'shiq repertuariga ega bo'lgan qushlarda jismoniy shaxslar bir xil qo'shiq turini baham ko'rishlari va murakkabroq aloqa qilish uchun ushbu qo'shiq turlaridan foydalanishlari mumkin.[20] Ba'zi qushlar umumiy qo'shiq turiga qo'shiq turidagi moslik bilan javob berishadi (ya'ni bir xil qo'shiq turi bilan).[21] Bu tajovuzkor signal bo'lishi mumkin; ammo, natijalar aralash.[20] Qushlar repertuar-matchlar yordamida ham o'zaro ta'sir qilishi mumkin, bunda qush raqibining repertuaridagi qo'shiq turi bilan javob beradi, lekin u hozir aytayotgan qo'shiq emas.[22] Bu qo'shiq turi bilan taqqoslaganda kamroq tajovuzkor harakat bo'lishi mumkin.[22] Qo'shiqning murakkabligi, shuningdek, erkaklarning hududiy mudofaasi bilan bog'liq bo'lib, yanada murakkab qo'shiqlar katta hududiy tahdid sifatida qabul qilinadi.[23]
Qushlarning qo'ng'iroqlari orqali aloqa bir xil turdagi shaxslar o'rtasida yoki hatto turlar bo'ylab bo'lishi mumkin. Qushlar tahdidga xos bo'lgan vokalizatsiya va harakatlar orqali signal beradi va qushlarning signallari boshqa tahlikani aniqlash va undan himoya qilish uchun boshqa hayvon turlari, shu jumladan boshqa qushlar tomonidan tushunilishi mumkin.[24] Mobbing qo'ng'iroqlar boyo'g'li yoki boshqa yirtqich mavjud bo'lishi mumkin bo'lgan hududga odamlarni jalb qilish uchun ishlatiladi. Ushbu chaqiriqlar keng tarqalgan chastotali spektrlar, keskin boshlanish va tugatish va takrorlanuvchanlik bilan ajralib turadi, ular turlar orasida keng tarqalgan bo'lib, ularni topish oson bo'lganligi sababli boshqa potentsial "mobberlar" ga yordam beradi. Aksariyat turlarning signal qo'ng'iroqlari, aksincha, o'ziga xos balandlikda bo'lib, qo'ng'iroq qiluvchining manzilini aniqlash qiyin.[25]
Shaxsiy qushlar o'zlarining qo'ng'iroqlari orqali bir-birlarini aniqlash uchun etarlicha sezgir bo'lishi mumkin. Koloniyalarda uya qiladigan ko'plab qushlar chaqiruvlari yordamida jo'jalarini topishlari mumkin.[26] Qo'ng'iroqlar ba'zida ekologik tadqiqotlarda ham odam tadqiqotchilari tomonidan individual identifikatsiyalash uchun etarlicha ajralib turadi.[27]
Ko'plab qushlar duet chaqiruvlari bilan shug'ullanadilar. Ba'zi hollarda, duetlar shu qadar mukammal tarzda tuzilganki, ular deyarli bitta qo'ng'iroq kabi ko'rinadi. Bunday qo'ng'iroq antifonal duet deb nomlanadi.[28] Bunday duetlik ko'plab oilalarda, shu jumladan bedanalarda,[29] bushshrikes,[30] babblers kabi scimitar babblers va ba'zi boyqushlar[31] va to'tiqushlar.[32] Hududiy qo'shiq qushlarida qushlar ko'proq uchraydi hisoblash ular o'z hududlariga simulyatsiya qilingan kirib kelish bilan qo'zg'atilganlarida.[33] Bu intraspetsiyalarda tajovuzkor raqobatdagi rolni anglatadi.
Ba'zida naslchilikdan keyingi mavsumda yangragan qo'shiqlar ibrat vazifasini o'taydi o'ziga xos tinglovchilar.[34] Yilda qora tomoqli ko'k jangchilar, muvaffaqiyatli o'stirgan va ko'paygan erkaklar o'zlarining avlodlariga vokal rivojlanishiga ta'sir qilish uchun qo'shiq aytishadi, ko'paytira olmagan erkaklar odatda uyalarni tashlab jim turishadi. Reproduktiv qo'shiq, shuning uchun reproduktiv muvaffaqiyatga erishish ehtimoli yuqori bo'lgan ma'lum yashash joylarining muvaffaqiyatsiz erkaklariga beixtiyor xabar beradi. The ijtimoiy aloqa vokalizatsiya orqali yuqori sifatli yashash joylarini aniqlash uchun yorliq va turli xil o'simlik tuzilmalarini to'g'ridan-to'g'ri baholash muammosidan xalos bo'lish.

Ba'zi qushlar ajoyib ovozga ega taqlid qilish. Ba'zi tropik turlarda taqlid qilish kabi drongolar shakllanishida rol o'ynashi mumkin aralash ozuqaviy podalar.[35] Vokal mimikriyasi o'ziga xos xususiyatlarni, boshqa turlarni yoki hatto sun'iy tovushlarni ham o'z ichiga olishi mumkin. Vokal mimikriyasi funktsiyalari bo'yicha ko'plab gipotezalar ishlab chiqilgan bo'lib, ular fitnes ko'rsatkichi sifatida harakat qilish, parazitlarga yordam berish yoki yirtqich hayvonlardan himoya qilish orqali jinsiy tanlanishda ishtirok etishi mumkinligi haqidagi takliflar mavjud, ammo har qanday funktsiyani kuchli qo'llab-quvvatlash kam.[36] Ko'plab qushlar, ayniqsa, bo'shliqlarda uya qiladigan qushlar, ilonga o'xshash xirillagan ovoz chiqarishi ma'lum, bu yirtqichlarni yaqin masofada oldini olishga yordam beradi.[37]
G'orlarda yashovchi ba'zi turlar, shu jumladan yog 'qushi[38] va tezyurar (Kollokaliya va Aerodramus turlari),[39] eshitiladigan tovushdan foydalaning (sonik joylashuvining aksariyati 2 va 5 kHz gacha)[40]) ga echolokat qiling g'orlar zulmatida. Faqatgina foydalanadigan qush infratovush (taxminan 20 Hz) bu g'arbiy kaperailli.[41]
Qushlarning eshitish doirasi 50 Gts dan past (infratovush ) 12 kHz atrofida, maksimal sezgirlik 1 dan 5 kHz gacha.[19][42] The qora yakobin taxminan 11,8 kHz chastotada tovushlarni chiqarishda alohida hisoblanadi. Ushbu tovushlarni eshitishlari mumkinmi yoki yo'qmi noma'lum.[43]
Atrof muhitda qushlar chaqiradigan chastotalar diapazoni yashash joyining sifati va atrofdagi tovushlar bilan farq qiladi. Akustik moslashish gipotezasi murakkab o'simlik qatlamlari (tovushlarni yutib yuboradigan va o'chiradigan) yashash joylarida tor tarmoqli kengligi, past chastotalar va uzun elementlar va elementlararo intervallarni topish kerakligini, yuqori chastotalar, keng tarmoqli kengligi, yuqori chastotali modulyatsiyalar ( trillalar), qisqa tutashgan elementlar va inter-elementlar obstruktsion o'simliklarsiz, ochiq joylarda kutilishi mumkin.[44][45][46]
Past chastotali qo'shiqlar to'siq qo'yilgan, zich o'simlik o'sadigan yashash joylari uchun maqbuldir, chunki past chastotali, sekin modulyatsiyalangan qo'shiq elementlari tovushni aks ettiruvchi o'simliklarning reverberatsiyalari orqali signallarning buzilishiga kamroq ta'sir qiladi. Tez modulyatsiyali yuqori chastotali qo'ng'iroqlar ochiq yashash joylari uchun maqbuldir, chunki ular ochiq maydonda kamroq tanazzulga uchraydi.[47][48] Akustik moslashish gipotezasida, shuningdek, qo'shiq xususiyatlari atrof-muhitning foydali akustik xususiyatlaridan foydalanishi mumkinligi aytilgan. Tor chastotali tarmoqli kengligi yozuvlari zich o'sadigan yashash joylarida aks sado berish orqali hajmi va uzunligi bo'yicha oshiriladi.[49]
Mavjud chastota diapazoni bo'linadi va qushlar chaqiradi, shuning uchun chastota va vaqt bo'yicha har xil turlar orasidagi qoplanish kamayadi. Ushbu g'oya "akustik joy" deb nomlangan.[50] Atrofdagi past chastotali shovqin bo'lgan shahar joylarida qushlar balandroq va balandroq balandlikda qo'shiq aytishadi.[51][52] Yo'l harakati shovqinlari reproduktiv muvaffaqiyatni pasaytirishi aniqlandi ajoyib tit (Parus major) akustik chastotadagi ustma-ust tushganligi sababli.[53] Qo'shiq tovushining oshishi sha joylarda qushlarning jismoniy holatini tikladi, shuningdek, yuqori chastotali qo'shiqlar.[54]
Qushlarning qo'shiq murakkabligida kenglik o'zgarishini ko'rsatishi taklif qilingan, ammo kenglik yoki ko'chib yuruvchi xatti-harakatlar bilan qo'shiqning murakkabligi oshib borishi haqida aniq dalillar yo'q.[55]
2019 yilda nashr etilgan tadqiqotga ko'ra oq qo'ng'iroq qushlar uchun qayd etilgan eng baland qo'ng'iroqni 125 ga etkazadidB.[56][57] Rekord ilgari qichqiriq piha 116 dB bilan.[58]
Neyroanatomiya

Qushlar qo'shig'ini sotib olish va o'rganish ikkita birlashtiruvchi yo'lga to'g'ri keladigan miya guruhlari guruhini o'z ichiga oladi:[59]
- Old miya yo'llari (vokal o'rganish ): sutemizuvchilar bazal ganglionlari uchun homolog bo'lgan X maydonidan tashkil topgan; ning lateral qismi magnosellular oldingi nidopalliyning yadrosi (LMAN), shuningdek parranda bazal ganglionlarining bir qismi hisoblangan; va medial talamusning (DLM) dorso-lateral bo'linishi.
- Orqa pastga tushadigan yo'l (vokal ishlab chiqarish): HVC dan tashkil topgan (maxsus nom, ba'zan uni deb ham nomlanadi yuqori vokal markazi ); arkopalliyning mustahkam yadrosi (RA); va trakeosyringeal qismi gipoglossal yadro (nXIIts).[60][61]
Orqa pastga tushadigan yo'l (PDP) qushlarning hayoti davomida normal qo'shiq ishlab chiqarish uchun talab qilinadi, oldingi miya yo'llari (AFP) qo'shiqlarni o'rganish, plastika va parvarish qilish uchun zarur, lekin kattalar uchun qo'shiq ishlab chiqarish uchun emas.[62]
Qo'shiq tizimidagi ikkala asab yo'li ham darajadan boshlanadi HVC, bu RA (premotor yadrosi) ga va oldingi oldingi miyaning X maydoniga ma'lumot beradi. Orqa tushish yo'lidagi ma'lumotlar (shuningdek, vokal ishlab chiqarish yoki vosita yo'li deb ataladi) pastga tushadi HVC RA ga, so'ngra RA dan trakeosyringeal qismiga gipoglossal asab (nXIIts), keyinchalik sirinning mushak qisqarishini boshqaradi.[59][63]
Old oldingi miya yo'lidagi ma'lumotlar prognoz qilinadi HVC X maydonga (bazal ganglionlar), keyin X maydondan DLM ga (talamus) va DLM dan LMAN ga, keyin esa vokal o'rganish va RA-ga ulanish orqali vokal ishlab chiqarish yo'llari. Ba'zi tergovchilar LMAN va RA o'rtasidagi aloqada eshitish geribildirimini baholashga asoslangan (qushning o'z qo'shig'ini yodlangan qo'shiq qolipi bilan taqqoslash) asoslangan ko'rsatma signalini beradigan modelni yaratdilar, bu esa qo'shiq chiqishi uchun motor dasturini o'zgartiradi.[62][64] Ushbu ibratli signalni yaratishda X va LMAN hududidagi eshitish neyronlari yordam berishi mumkin, ular qushning o'ziga xos qo'shig'i (BOS) va uning o'qituvchisi qo'shig'ining vaqtinchalik fazilatlari uchun selektivlikni namoyish etadi, bu BOS va yodlangan repetitor qo'shig'ini taqqoslash uchun maydon yaratadi.[64][65]
Kelajakda AFP va PDP o'rtasidagi xatolarni tuzatishning o'zaro ta'siriga oid modellar ko'rib chiqiladi. Boshqa hozirgi tadqiqotlar asosidagi uyali mexanizmlarni o'rganishga kirishdi HVC qo'shiq tarkibining vaqtinchalik naqshlarini boshqarish va hece hosil bo'lishining RA nazorati.[66]Ikkala yo'lda ishtirok etadigan miya tuzilmalari namoyon bo'ladi jinsiy dimorfizm ko'plab qush turlarida, odatda erkak va urg'ochi ayollarning turlicha qo'shiq aytishiga sabab bo'ladi. Miyada ma'lum bo'lgan dimorfizmlarning ayrim turlariga yadrolarning kattaligi, mavjud bo'lgan neyronlar soni va bitta yadroni boshqasiga bog'laydigan neyronlar kiradi.[67]
Juda dimorfik zebra sichqonlarida (Taeniopygia guttata), odatda faqat erkaklar qo'shiq aytadigan tur, HVC va RA kattaligi erkaklarda ayollarga qaraganda uch-olti baravar katta va X maydon ayollarda taniqli ko'rinmaydi.[68] Tadqiqotlar shuni ko'rsatadiki, erta rivojlanish davrida jinsiy steroidlarga ta'sir qilish miyadagi ushbu farqlar uchun qisman javob beradi. Estradiol bilan davolash qilingan urg'ochi zebra finches, keyin testosteron yoki dihidrotestosteron (DHT) davolash katta yoshdagi erkaklarnikiga o'xshash RA va HVC ni rivojlantiradi va shuningdek, erkaklarga o'xshash qo'shiq xatti-harakatlarini namoyish etadi.[69]
Faqatgina gormonlarni davolash, xuddi erkaklarga o'xshab, miya tuzilishi yoki xulq-atvori bilan urg'ochi ayollarni ishlab chiqarmaydi. Bundan tashqari, boshqa tadqiqotlar sutemizuvchilarning jinsiy farqlanishi haqidagi hozirgi bilimimiz asosida kutilgan narsalarga zid bo'lgan natijalarni ko'rsatdi. Masalan, erkaklar zebra fincheslari kastratsiya qilingan yoki jinsiy steroid inhibitörleri berilgan, chunki ular hali ham odatdagidek erkaklar ashulasini rivojlantiradi.[67] Bu shuni ko'rsatadiki, z xromosomasidagi genlarning faollashishi kabi boshqa omillar ham erkak qo'shig'ining normal rivojlanishida rol o'ynashi mumkin.[70]
Gormonlar, shuningdek, kattalar qushlaridagi qo'shiq va qo'shiq yadrolariga faol ta'sir ko'rsatadi. Kanareykalarda (Serinus kanariya), urg'ochilar odatda erkaklarnikiga qaraganda kamroq va kamroq murakkablikda kuylashadi. Ammo, kattalar urg'ochilariga androgen in'ektsiyalari yuborilganda, ularning qo'shiqlari deyarli erkaklarga o'xshash chastotaga ko'payadi.[71] Bundan tashqari, androgen bilan AOK qilingan kattalar urg'ochi ayollarda HVC va RA mintaqalarida kattalashgan kattalik namoyon bo'ladi.[72] Melatonin kattalardagi qo'shiq xatti-harakatlariga ta'sir qiladi deb hisoblanadigan yana bir gormondir, chunki ko'plab qo'shiq qushlari qo'shiq yadrolari neyronlarida melatonin retseptorlarini ko'rsatadi.[73]
Ikkalasi ham Evropalik yulduzcha (Sturnus vulgaris) va uy chumchuqi (Passer domesticus) melatonin zulmatiga va sekretsiyasiga ta'sir qilish bilan bog'liq bo'lgan qo'shiq yadrolarining o'zgarishini ko'rsatdi.[74][75] Bu melatonin yil davomida kun yorug'ligi sezilarli darajada o'zgarib turadigan joylarda yashovchi qo'shiq qushlaridagi qo'shiq xatti-harakatlarining mavsumiy o'zgarishlarida rol o'ynashi mumkinligidan dalolat beradi. Boshqa bir qator tadqiqotlar qo'shiq tizimidagi miya tuzilmalari morfologiyasidagi mavsumiy o'zgarishlarni ko'rib chiqdi va ushbu o'zgarishlar (kattalar neyrogenezi, gen ekspressioni) fotoperiod, gormonal o'zgarishlar va xulq-atvor bilan belgilanadi.[76][77]
Gen FOXP2, nuqsonlar nutqni ishlab chiqarishga ham, odamlarda tilni tushunishga ham ta'sir qiladi, X zonasida voyaga etmagan zebra sichqonlari va kattalar kanareykalarida vokal plastika davrida yuqori darajada namoyon bo'ladi.[78]
O'rganish

Turli xil qushlarning qo'shiqlari turlicha va odatda turlarga xosdir. Turlar o'zlarining qo'shiqlarining murakkabligi va ular aytadigan qo'shiq turlarining ko'pligi bilan juda farq qiladi (3000 ga qadar jigarrang trasher ); ayrim turlar ichidagi individlar xuddi shu tarzda farq qiladi. Kabi bir nechta turlarda lira qushlari va masxara qushlar, qo'shiqlar inson hayoti davomida o'rganilgan o'zboshimchalik elementlarini singdirgan, taqlidning bir shakli (garchi "o'zlashtirish" (Ehrlich va boshq.) deb nomlanishi mumkin, chunki qush boshqa turga o'tmaydi). 1773 yildayoq qushlar qo'ng'iroqlarni o'rganishgani va o'zaro faoliyat tajribalar linnet yasashga muvaffaq bo'ldi Acanthis nasha samoviy qo'shiqni o'rganing, Alauda arvensis.[80] Ko'pgina turlarda, asosiy qo'shiq turning barcha a'zolari uchun bir xil bo'lsa-da, yosh qushlar o'zlarining qo'shiqlarining ba'zi tafsilotlarini otalaridan o'rganishadi va bu xilma-xilliklar avlodlar davomida shakllanib boradi. lahjalar.[81]
Voyaga etmagan qushlarda qo'shiqni o'rganish ikki bosqichda sodir bo'ladi: sezgir o'rganish, bu voyaga etmagan bolani otasini yoki boshqa o'ziga xos qushni tinglash va qo'shiqning spektral va vaqtinchalik fazilatlarini yodlashni o'z ichiga oladi (qo'shiq qolipi) va voyaga etmagan qushni ishlab chiqarishni o'z ichiga olgan sensorimotor o'rganish. o'z vokalizatsiyasi va qo'shig'ini yodlangan qo'shiq shabloniga aniq mos kelguncha mashq qilish.[82]
Sensorli motorni o'rganish bosqichida qo'shiq ishlab chiqarish "sub-song" deb nomlanadigan juda o'zgaruvchan sub-vokalizatsiya bilan boshlanadi. gapirish inson go'daklarida. Ko'p o'tmay, balog'atga etmagan bolalar uchun qo'shiq taqlid qilingan kattalar qo'shig'ining ma'lum taniqli xususiyatlarini namoyish etadi, ammo baribir kristallangan qo'shiqning stereotipi yo'q - bu "plastik qo'shiq" deb nomlanadi.[59]
Ikki yoki uch oylik qo'shiqlarni o'rganish va mashq qilishdan so'ng (turlarga qarab) balog'atga etmagan bola spektral va vaqtinchalik stereotipiya (bo'g'inlar hosil bo'lishi va bo'g'in tartibida juda past o'zgaruvchanlik) bilan ajralib turadigan kristallangan qo'shiq hosil qiladi.[83] Kabi ba'zi qushlar zebra finches, qushlar qo'shig'ini tadqiq qilish uchun eng mashhur turlar bo'lgan, sensorli va sensorimotor o'rganish bosqichlari bir-biriga to'g'ri keladi.[79]
Tadqiqotlar shuni ko'rsatdiki, qushlarning qo'shiqni qo'lga kiritish shakli motorli o'rganish bu mintaqalarni o'z ichiga oladi bazal ganglionlar. Keyinchalik, PDP (qarang. Qarang Neyroanatomiya quyida) ko'rib chiqildi gomologik dan kelib chiqqan sutemizuvchilar motorli yo'liga miya yarim korteksi va orqali pastga tushish miya sopi, AFP esa bazal ganglionlar va talamus orqali sutemizuvchilarning kortikal yo'li uchun homolog hisoblanadi.[59] Qushlar qo'shig'ining modellari motorli o'rganish odamlar qanday o'rganishlari uchun modellarni ishlab chiqishda foydali bo'lishi mumkin nutq.[84] Zebra finches kabi ba'zi turlarda qo'shiqni o'rganish birinchi yil bilan cheklangan; ular "yoshi cheklangan" yoki "yaqin" o'quvchilar deb nomlanadi. Kanareykalar kabi boshqa turlar, hatto jinsiy etuk kattalar singari yangi qo'shiqlarni ishlab chiqishi mumkin; bular "ochiq" o'quvchilar deb nomlanadi.[85][86]
Tadqiqotchilar o'rganilgan qo'shiqlar madaniy ta'sir o'tkazish orqali yanada murakkab qo'shiqlarni ishlab chiqishga imkon beradi, shuning uchun qushlarga qarindoshlarni aniqlashda va ularning qo'shiqlarini turli xil akustik muhitga moslashtirishda yordam beradigan ichki dialektlarga imkon beradi deb taxmin qilishdi.[87]
Qushlar qo'shig'ini o'rganishda eshitish teskari aloqasi
1954 yilda Torp tomonidan o'tkazilgan dastlabki tajribalar qushning repetitor qo'shig'ini eshita olishining muhimligini ko'rsatdi. Qushlarni alohida-alohida, o'ziga xos erkaklar ta'siridan uzoqlashtirganda, ular hali ham qo'shiq aytishadi. Ular tomonidan ishlab chiqarilgan "izolyatsiya qilingan qo'shiq" deb nomlangan qo'shiq yovvoyi qushning qo'shig'iga o'xshash bo'lsa-da, u yirtqich qo'shiqdan aniq farqli xususiyatlarni namoyish etadi va uning murakkabligiga ega emas.[88][89] Qushning sensimotor davrda o'z qo'shig'ini eshita olishining ahamiyati keyinchalik Konishi tomonidan aniqlandi. Qo'shiq-kristallanish davri oldidan kar bo'lgan qushlar yovvoyi turdan ajralib turuvchi va alohida bo'lgan qo'shiqlarni yaratishga kirishdilar.[90][91] Ushbu topilmalar paydo bo'lganidan beri tergovchilar sensorli / sensorimotor o'rganishni osonlashtiradigan asab yo'llarini qidirmoqdalar va qushning o'z qo'shig'ini yodlangan qo'shiq shabloniga moslashtirishda vositachilik qildilar.
So'nggi o'n yilliklarda olib borilgan bir qator tadqiqotlar qushlarning qo'shiqlarini ishlab chiqarish yoki parvarish qilish bilan bog'liq bo'lgan tegishli miya tuzilmalariga zarar etkazish yoki qo'shiq kristallanishidan oldin va / yoki undan keyin karlarni quloqsizlantirish yo'li bilan o'rganishning asab mexanizmlarini ko'rib chiqdi. Yaqinda o'tkazilgan yana bir eksperimental yondashuv - qushning qo'shig'ini yozib olish va keyin uni qush kuylayotganda ijro etish, buzilgan eshitish teskari aloqasini keltirib chiqarish (qush o'z qo'shig'ining superpozitsiyasini va oldingi qo'shiq bo'g'inining bo'lak qismini eshitadi).[83] Nordeen va Nordeen'dan keyin[92] Leonardo & Konishi (1999) kattalar qushlaridagi qo'shiqni kristallangan qo'shiq bilan saqlash uchun eshitish teskari aloqasi zarurligini namoyish etishlari bilan muhim kashfiyot qildilar, kattalar qo'shig'ini saqlashda eshitish teskari aloqa rolini o'rganish uchun , buzilgan eshitish teskari ta'siridan keyin kattalar qo'shiqlarining qanday yomonlashishini tekshirish va buzilgan teskari ta'sirdan chiqarilgandan so'ng, kattalar qushlarining vaqt o'tishi bilan kristallangan qo'shiqni tiklab olish darajasini o'rganish. Ushbu tadqiqot kattalar qo'shig'ining barqarorligini saqlashda eshitish teskari aloqa rolini yanada qo'llab-quvvatlashni taklif qildi va kristallangan qushlarning qo'shiqlarini statik emas, balki dinamikligini namoyish etdi.
Brainard & Doupe (2000) qush tomonidan ishlab chiqarilgan qo'shiq va uning yodlangan qo'shiq shabloni o'rtasidagi farqlarni aniqlaganligi sababli xatolarni tuzatishda LMAN (oldingi oldingi miyaning) asosiy rol o'ynaydigan modelni namoyish etadi va qo'shiq ishlab chiqarish uchun motor dasturini tuzatish yoki o'zgartirish uchun vokal ishlab chiqarish yo'lidagi tuzilmalar. Brainard & Doupe (2000) o'zlarining tadqiqotlarida shuni ko'rsatdiki, kattalar qushlarini kar bo'lganida, eshitish teskari aloqasi va motor dasturining moslashuvchan bo'lmagan modifikatsiyasi tufayli qo'shiq stereotipiyasining yo'qolishiga olib keldi, kattalar qushlarining oldingi miya yo'lida LMAN zararlangandi. kar bo'lganligi qo'shiqning barqarorlashishiga olib keldi (kar bo'lgan qushlardagi LMAN shikastlanishi, hece ishlab chiqarish va qo'shiq tarkibining yanada yomonlashishiga yo'l qo'ymadi).
Hozirgi vaqtda LMANning ko'rsatma xato signalini ishlab chiqarishda va uni dvigatel ishlab chiqarish yo'lida aks ettirishdagi rolini yoritib beradigan ikkita raqobatdosh model mavjud:
Qushlarning o'z qo'shig'i (BOS) tomonidan sozlangan xatolarni tuzatish modeli
- Qo'shiq paytida LMAN neyronlarining faollashishi qush tomonidan ishlab chiqarilgan qo'shiq va saqlanadigan qo'shiq shablonlari o'rtasidagi eshitish aloqasi o'rtasidagi bog'liqlikka bog'liq bo'ladi. Agar bu to'g'ri bo'lsa, u holda LMAN neyronlarining otish tezligi eshitish aloqalarining o'zgarishiga sezgir bo'ladi.
Xatolarni tuzatishning effektiv nusxasi
- An efference nusxasi qo'shiq ishlab chiqarish uchun vosita buyrug'i real vaqtda xatolarni tuzatish signalining asosidir. Qo'shiq paytida LMAN neyronlarining faollashishi qo'shiqni yaratish uchun ishlatiladigan vosita signaliga va ushbu vosita buyrug'i asosida kutilgan eshitish teskari aloqa ma'lumotlariga bog'liq bo'ladi. Xatolarni tuzatish ushbu modelda tezroq sodir bo'ladi.
Leonardo [93] bu modellarni to'g'ridan-to'g'ri normal va buzilgan eshitish geribildirimi sharoitida qo'shiq paytida kattalar zebra sichqonlarining bitta LMAN neyronlarida boshoq tezligini qayd etish orqali sinovdan o'tkazdi. Uning natijalari BOS-sozlangan xatolarni tuzatish modelini qo'llab-quvvatlamadi, chunki LMAN neyronlarining otish tezligi eshitish teskari ta'sirining ta'siriga ta'sir qilmadi va shuning uchun LMAN tomonidan ishlab chiqarilgan xato signali eshitish aloqasi bilan bog'liq bo'lmagan holda paydo bo'ldi. Bundan tashqari, ushbu tadqiqot natijalari LMAN neyronlari motor signalining efferentsiya nusxasi bilan qo'shiq paytida (va kutilgan eshitish teskari aloqasi haqidagi bashoratlari) qo'shiq paytida faollashtirilgan nusxa ko'chirish modelining bashoratlarini qo'llab-quvvatladi, bu esa neyronlarning vaqtini aniqroq bo'lishiga imkon berdi. - eshitish mulohazalarining o'zgarishi uchun bloklangan.
Ko'zgu neyronlari va vokal o'rganish
A ko'zgu neyroni a neyron bu biron bir harakatni amalga oshirganda ham, xuddi shu harakatni boshqasi tomonidan amalga oshirilishini sezganda ham bo'shatadi.[94] Ushbu neyronlar birinchi marta kashf etilgan makak maymunlar, ammo so'nggi tadqiqotlar shuni ko'rsatadiki, ko'zgu neyron tizimlari boshqa hayvonlarda, shu jumladan odamlarda ham bo'lishi mumkin.[95]

Ko'zgu neyronlari quyidagi xususiyatlarga ega:[94]
- Ular joylashgan prekotor korteks.
- Ular hissiy va motor xususiyatlarini namoyish etadi.
- Ular harakatga xosdir - ko'zgu neyronlari faqatgina shaxs muayyan harakat turini bajarganda yoki kuzatayotganda (masalan, ob'ektni ushlashda) faollashadi.
Chunki ko'zgu neyronlari ikkalasini ham namoyish etadi sezgir va vosita Ba'zi tadqiqotchilar ko'zgu neyronlari vosita tuzilmalariga hissiy tajribani solishtirishga xizmat qilishi mumkin, degan fikrni bildirishdi.[97] Bu qushlarning qo'shiqlarini o'rganishga ta'sir qiladi - ko'plab qushlar o'zlarining qo'shiqlarini olish va saqlab qolish uchun eshitish fikrlariga ishonadilar. Ko'zgu neyronlari qush nimani eshitishi, uning yodlangan qo'shiq shabloniga solishtirishi va u nimalar ishlab chiqarishi bilan taqqoslashda vositachi bo'lishi mumkin.
Jonatan Prater va Dyuk Universitetining boshqa tadqiqotchilari ushbu eshitish-motorli neyronlarni izlashda bitta neyronlarning faolligini qayd etdilar. HVClar ning botqoq chumchuqlar.[96] Ular HVC dan X maydonga (HVC) tushadigan neyronlarni topdilarX neyronlar) qush o'z qo'shig'ini ijro etishni eshitganda juda sezgir. Qush xuddi shu qo'shiqni aytganda, bu neyronlar ham xuddi shunday naqshlarda yonadi. Botqoq chumchuqlarda 3-5 xil qo'shiq turi mavjud bo'lib, asab faoliyati qaysi qo'shiq eshitilishi yoki kuylanishiga qarab farqlanadi. HVCX neyronlar faqat asosiy qo'shiq turi bo'lgan qo'shiqlardan birining taqdimotiga (yoki kuylanishiga) javoban otashadi. Ular, shuningdek, vaqtincha tanlangan bo'lib, qo'shiq bo'g'inining aniq bosqichida otishadi.
Prater va boshq. qush qo'shiq aytishdan oldin va keyin qisqa vaqt ichida uning HVCX neyronlar befarq bo'lib qoladi eshitish kiritish. Boshqacha qilib aytganda, qush o'z qo'shig'iga "kar" bo'lib qoladi. Bu shuni ko'rsatadiki, bu neyronlar a hosil qiladi xulosa chiqarish, bu vosita chiqishi va eshitish qobiliyatini to'g'ridan-to'g'ri taqqoslash imkonini beradi.[98] Bu eshitish teskari aloqasi orqali o'rganish asosidagi mexanizm bo'lishi mumkin. Ushbu topilmalar, shuningdek, Leonardoning (2004) qushlarni o'qish va ishlab chiqarishda xatolarni to'g'rilashning efferents nusxasi modeliga mos keladi.
Umuman olganda, HVCX botqoq chumchuqlaridagi eshitish vositasi neyronlari kashf etilgan vizual vosita oynasi neyronlariga juda o'xshaydi primatlar. Ko'zgu neyronlari singari, HVCX neyronlar:
- A joylashgan oldingi vosita miya maydoni
- Ham sezgir, ham vosita xususiyatlarini namoyish eting
- Amalga xosmi - javob faqat "asosiy qo'shiq turi" orqali amalga oshiriladi
Ko'zgu neyron tizimining vazifasi hali ham aniq emas. Ba'zi olimlar ko'zgu neyronlari boshqalarning harakatlarini tushunishda rol o'ynashi mumkin deb taxmin qilishadi, taqlid, ong nazariyasi va tilni o'rganish ammo hozircha bu etarli emas neyrofiziologik ushbu nazariyalarni tasdiqlovchi dalillar.[97] Xususan, qushlarga nisbatan, ko'zgu neyronlari tizimi umumiy mexanizm bo'lib xizmat qilishi mumkin vokal o'rganish, ammo qo'shimcha tadqiqotlar o'tkazish kerak. Qo'shiqni o'rganishga ta'sir qilishdan tashqari, ko'zgu neyron tizimi ham rol o'ynashi mumkin hududiy xatti-harakatlar qo'shiq turiga mos kelish va hisoblash.[99][100]
Madaniy uzatish orqali o'rganish
Hayvonlarda madaniyat odatda ma'lum bir populyatsiyalarga xos bo'lgan ijtimoiy uzatiladigan xatti-harakatlar shakllaridan ("urf-odatlar") iborat deb belgilanadi.[101] Qushlar qo'shig'ining o'rganilgan tabiati va "dialekt" ga o'xshash mahalliy o'zgarishlarning dalillari ba'zi tadqiqotchilarning mavjudligini faraz qilishga undadi. parrandalar madaniyati.
Yuqorida aytib o'tilganidek yuqorida, qushlar qo'shig'ining o'rganishga bog'liqligi Torp tomonidan o'rganilgan va buni aniqlagan chaffinches hayotining birinchi haftasidan boshlab ajratilgan holda ko'tarilib, boshqa chaffinches bilan taqqoslaganda juda g'ayritabiiy va unchalik murakkab bo'lmagan qo'shiqlar tug'diradi.[102] Bu qo'shiq qushlaridagi qo'shiqni rivojlantirishning ko'p jihatlari bir xil yoshdagi a'zolarning repetitorligiga bog'liqligini ko'rsatdi. Keyinchalik tadqiqotlar ko'tarilgan chaffinch qo'shig'ida kanareyka o'xshash elementlarni kuzatdi kanareykalar,[103] voyaga etmagan qushlar tomonidan qo'shiqni o'rganishda repetitorlarning kuchli rolidan dalolat beradi.
Shunga o'xshash chaffinch qo'shiq turlari (alohida elementlari va tartibiga qarab turkumlanadi) o'xshash geografik hududlarda klaster qilinganligi kuzatildi,[104] va bu kashfiyot qushlar qo'shig'ida "dialektlar" haqidagi farazlarni keltirib chiqardi. O'shandan beri ushbu qo'shiq turlarining farqlari yo'q deb e'lon qilingan lahjalar biz inson tilida topilgan narsalar kabi. Buning sababi shundaki, ma'lum bir geografik hududning barcha a'zolari bir xil qo'shiq turiga mos kelmaydi, shuningdek, uni boshqa barcha turlardan ajratib turadigan qo'shiq turining o'ziga xos xususiyati mavjud emas (ba'zi so'zlar ma'lum dialektlarga xos bo'lgan odam shevalaridan farqli o'laroq) ).[101]
O'rganishning ushbu dalillariga va mahalliylashtirilgan qo'shiq turlariga asoslanib, tadqiqotchilar qushlarning qo'shiqlarini ijtimoiy o'rganishni madaniy uzatish shakli sifatida o'rganishni boshladilar. Ushbu madaniyatni tashkil etuvchi xulq-atvor naqshlari qo'shiqlarning o'zi va qo'shiq turlarini an'analar deb hisoblash mumkin.
Dopamin zanjirlari va madaniy uzatish
Yaqinda o'tkazilgan tadqiqot shuni ko'rsatdiki, zopak finchlaridagi dopamin sxemasi repetitorlardan qushlar qo'shig'ini ijtimoiy o'rganishga yordam berishi mumkin.[105] Ularning ma'lumotlari shuni ko'rsatadiki, balog'atga etmagan zebra sichqonlarida miya sohalari zebra finch qo'shig'ini ijro etadigan karnaylar tomonidan emas, balki o'ziga xos (ya'ni bir xil) repetitorlarning qo'shiqlari bilan hayajonlanadi. Bundan tashqari, ular HVC-ga chiqarilgan dopaminning qo'shiqni kodlashda yordam berishini ko'rsatmoqdalar.
Qushlarning vokal ta'limini evolyutsion tarzda saqlash
Madaniy tuzoq gipotezasi
20-asr davomida qushlar qo'shig'i haqida juda ko'p tadqiqotlar olib borilgan bo'lsa ham, hech kim qushlarning qo'shiqlari evolyutsiyasi "ishlatilishini" aniqlay olmadi, ayniqsa katta vokal repertuarlari bilan bog'liq. Bunga javoban Lachlan va Slater qo'shiqlarning turlarining turg'unligini tushuntirish uchun "madaniy tuzoq" modelini taklif qilishdi.[106] Ushbu model "filtrlar" tushunchasiga asoslangan bo'lib, unda:
- erkak qo'shiqchining (ya'ni qo'shiqchining) filtrida u rivojlanishi mumkin bo'lgan qo'shiqlar doirasi mavjud
- urg'ochi qushning (ya'ni qabul qiluvchining) filtrida turmush o'rtog'i tanlashi uchun maqbul deb topilgan qo'shiqlar oralig'i mavjud
Mumkin bo'lgan bitta vaziyatda populyatsiya asosan keng filtrlarga ega qushlardan iborat. Ushbu populyatsiyada tor filtrli kam sonli ayol kamdan-kam hollarda keng filtrga ega bo'lgan erkak qo'shiqchini tanlaydi (chunki erkakning qo'shig'i torroq filtrga tushishi mumkin emas). Endi bu urg'ochi bilan juftlashadigan erkaklar nisbatan kichik tanlovga ega bo'ladi, shuning uchun ayolning tor filtrining genetik asoslari saqlanib qolmaydi. Mumkin bo'lgan yana bir holat asosan tor filtrlarga ega bo'lgan aholi bilan bog'liq. So'nggi populyatsiyada keng filtrlangan erkaklar yoshi kattaroq, tor filtrli erkaklardan o'rganish orqali turmush o'rtog'ining tanlovidan voz kechishlari mumkin. Shu sababli, keng filtrli qushlarning o'rtacha reproduktiv muvaffaqiyati o'rganish imkoniyati bilan yaxshilanadi va vokal o'rganish va katta qo'shiq repertuarlari (ya'ni keng filtrlar) qo'lma-qo'l yuradi.[106][101]
The cultural trap hypothesis is one example of gene-culture coevolution, in which selective pressures emerge from the interaction between genotypes and their cultural consequences.[106]
Possible correlation with cognitive ability
Various studies have shown that adult birds that underwent stress during critical developmental periods produce less complex songs and have smaller HVC brain regions.[107][108] These has led some researchers to hypothesize that sexual selection for more complex songs indirectly selects for stronger cognitive ability in males.[109] Further investigation showed that male qo'shiq chumchuqlar with larger vocal repertoires required less time to solve detour-reaching cognitive tasks.[110] Some have proposed that bird song (among other sexually selected traits such as flashy coloring, body symmetry, and elaborate courtship) allow female songbirds to quickly assess the cognitive skills and development of multiple males.
Identification and systematics

The specificity of bird calls has been used extensively for species identification. The calls of birds have been described using words or nonsense syllables or line diagrams.[111] Common terms in English include words such as quack, chirillash va chirrup. These are subject to imagination and vary greatly; a well-known example is the oq tomoqli chumchuq 's song, given in Kanada kabi O sweet Canada Canada Canada va Yangi Angliya kabi Old Sam Peabody Peabody Peabody (shuningdek Where are you Frederick Frederick Frederick?). In addition to nonsense words, grammatically correct phrases have been constructed as likenesses of the vocalizations of birds. Masalan, taqiqlangan boyqush produces a motif which some bird guides describe as Who cooks for you? Who cooks for you all? with the emphasis placed on siz.[112] The term "warblish" has been coined to explain this approach to bird call description.[113]

Kay Electric Company started by former Bell Labs engineers Harry Foster and Elmo Crump made a device that was marketed as the "Sona-Graph" in 1948. This was adopted by early researchers[114] including C.E.G.Bailey who demonstrated its use for studying bird song in 1950.[115] Dan foydalanish spektrogramlar to visualize bird song was then adopted by Donald J. Borror[116] and developed further by others including W. H. Thorpe.[117][118] These visual representations are also called sonograms or sonagrams. Beginning in 1983, some field guides for birds use sonograms to document the calls and songs of birds.[119] The sonogram is objective, unlike descriptive phrases, but proper interpretation requires experience. Sonograms can also be roughly converted back into sound.[120][121]
Bird song is an integral part of bird courtship and is a pre-zygotic isolation mechanism involved in the process of spetsifikatsiya. Ko'pchilik allopatrik sub-species show differences in calls. These differences are sometimes minute, often detectable only in the sonograms. Song differences in addition to other taxonomic attributes have been used in the identification of new species.[122] The use of calls has led to proposals for splitting of species complexes such as those of the Mirafra bushlarks.[123]
Smartphone apps such as Bird Song Id USA[124] enables automatic recognition of birds using sounds.[125] Yoqdi Shazam, those apps work by comparing spektrografik database for matches.
Qush tili
The qushlarning tili has long been a topic for anecdote and speculation. That calls have meanings that are interpreted by their listeners has been well demonstrated. Ichki tovuqlar have distinctive alarm calls for aerial and ground predators, and they respond to these alarm calls appropriately.[126][127]
Biroq, a til has, in addition to words, grammatika (that is, structures and rules). Studies to demonstrate the existence of language have been difficult due to the range of possible interpretations. For instance, some have argued that in order for a communication system to count as a language it must be "combinatorial",[128] having an open ended set of grammar-compliant sentences made from a finite vocabulary.
Bo'yicha tadqiqotlar to'tiqushlar tomonidan Irene Pepperberg is claimed to demonstrate the innate ability for grammatical structures, including the existence of concepts such as nouns, adjectives and verbs.[129] Yovvoyi tabiatda, Black-capped Chickadees innate vocalizations have been rigorously shown[128] to have combinatorial language. Bo'yicha tadqiqotlar starling vocalizations have also suggested that they may have recursive structures.[130]
Atama bird language may also more informally refer to patterns in bird vocalizations that communicate information to other birds or other animals in general.[131]
Some birds have two distinct "languages", one for internal communications and one for use in flocks. All birds have a separate type of communication for "songs" vs. communicating danger and other information. Konrad Lorenz demonstrated that Jackdaws have "names" identifying each individual in the flock and when beginning flight preparations each of them say one other bird's name creating a "chain". In his book King Solomon's Ring, he describes the name he was given by the birds and how he was recognized several years later in a far away location following WWII.
Studies in parakeets have shown a striking similarity between gapiruvchi qush 's verbal areas in the brain and the equivalent human brain areas, suggesting that mimicry has much to do with the construction of language and its structures and order.[132] Recent research showed birds construct sentence like communications with a syntax and grammar.[133]
Madaniyatda
Yozib olish
Ushbu bo'lim kengayishga muhtoj. Siz yordam berishingiz mumkin unga qo'shilish. (2011 yil sentyabr) |
The first known recording of birdsong was made in 1889 by Lyudvig Koch,[134] who went on to become an eminent wildlife recordist and BBC natural history presenter.[134]
Other notable birdsong recordists include Erik Simms, Kris Uotson and, in France, Jean-Claude Roché, François Charron, Fernand Deroussen.
Musiqada
Musiqada, qushlar qo'shig'i has influenced composers and musicians in several ways: they can be inspired by birdsong; they can intentionally imitate bird song in a composition, as Vivaldi va Betxoven did, along with many later composers; kabi qushlarning yozuvlarini o'z asarlariga kiritishi mumkin Ottorino Respighi birinchi qildi; yoki shunga o'xshash Beatrice Harrison va Devid Rothenberg, they can duet with birds.[135][136][137][138] Authors including Rothenberg have claimed that birds sing on traditional scales as used in human music,[139][140][141] but at least one songbird does not choose notes in this way.[142]
Among birds which habitually borrow phrases or sounds from other species, the way they use variations of ritm, munosabatlar musiqiy pitch, and combinations of eslatmalar musiqaga o'xshash bo'lishi mumkin.[143] Hollis Taylor's in-depth analysis of qarag'ay qassob vocalizations provides a detailed rebuttal to objections of birdsong being judged as music.[144] The similar motor constraints on human and avian song may have driven these to have similar song structures, including "arch-shaped and descending melodic contours in musical phrases", long notes at the ends of phrases, and typically small differences in pitch between adjacent notes, at least in birds with a strong song structure like the Eurasian treecreeper Certhia tanish.[145]
She'riyatda
Bird song is a popular subject in she'riyat. Famous examples inspired by bird song include the 1177 Forscha she'r "Qushlar konferentsiyasi ", in which the birds of the world assemble under the wisest bird, the xoop, to decide who is to be their king.[146] In English poetry, Jon Kits 's 1819 "Bulbulga odob "va Persi Byishe Shelli 's 1820 "Skylarkga "mashhur klassiklardir.[147][148] Ted Xyuz 's 1970 collection of poems about a bird character, "Qarg'a ", is considered one of his most important works.[149] Bird poems by Jerar Manli Xopkins include "Sea and Skylark" and "Windhover ".[150]
Shuningdek qarang
Adabiyotlar
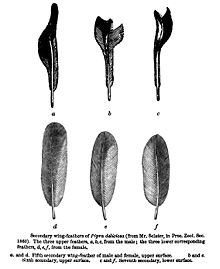
- ^ Sclater, P. L. (1860). "List of Birds collected by Mr. Fraser in Ecuador, at Nanegal, Calacali, Perucho, and Puellaro, with notes and descriptions of new species". Proc. Zool. Soc. London: 83–97.
- ^ Darvin, Charlz (1871). The descent of man and selection in relation to sex. jild 2. London: Jon Myurrey. 65-66 betlar. ISBN 978-1-108-00510-4.
- ^ Boswall, Jeffery. "Why do birds sing?". Britaniya kutubxonasi. Arxivlandi asl nusxasi 2020 yil 29 yanvarda.
- ^ Ehrlich, Pol R.; David S. Dobkin & Darryl Wheye. ""Bird Voices" and "Vocal Development" from Birds of Stanford essays". Olingan 9 sentyabr 2008.
- ^ Howell, Steve N. G. & Sophie Webb (1995). Meksika va Shimoliy Markaziy Amerika qushlari uchun qo'llanma. Oksford universiteti matbuoti. ISBN 978-0-19-854012-0.
- ^ Bostwick, Kimberly S. & Richard O. Prum (2005). "Qo'rqinchli qushlar qotib turadigan qanotli tuklar bilan qo'shiq aytadi". Ilm-fan. 309 (5735): 736. doi:10.1126 / science.1111701. PMID 16051789. S2CID 22278735.
- ^ Manson-Barr, P. and Pye, J. D. (1985). Mechanical sounds. In A Dictionary of Birds (ed. B. Campbell and E. Lack), pp. 342-344. Staffordshire: Poyser.
- ^ a b Bostwick, Kimberly S. & Richard O. Prum (2003). "High-speed video analysis of wing-snapping in two manakin clades (Pipridae: Aves)". Eksperimental biologiya jurnali. 206 (Pt 20): 3693–3706. doi:10.1242/jeb.00598. PMID 12966061. S2CID 8614009.
- ^ a b Robinson, Angus; "The Biological Significance of Bird Song in Australia" yilda Emu; 48(4), pp. 291-315
- ^ Hartshorne, Charles; "Some Biological Principles Applicable to Song Behavior" in Uilson byulleteni Vol. 70, No. 1 (Mar., 1958), pp. 41-56
- ^ Slater Peter J. B., Mann Nigel I. (2004). "Why do the females of many bird species sing in the tropics?". Qushlar biologiyasi jurnali. 35 (4): 289–294. doi:10.1111/j.0908-8857.2004.03392.x.
- ^ Attenboro, D. 1998 yil. Qushlarning hayoti. BBC publication.0563-38792-0
- ^ Avstraliya muzeyi Onlayn. "Crows and Ravens". Arxivlandi asl nusxasidan 2007 yil 1 sentyabrda. Olingan 12 avgust 2007.
- ^ a b Catchpole, C.; Slater, P.J.B (2008). Bird Song: Biological Themes and Variation. Kembrij universiteti matbuoti.
- ^ a b Read, A. W. & D. M. Weary (1990). "Sexual selection and the evolution of bird song: A test of the Hamilton-Zuk hypothesis". Xulq-atvor ekologiyasi va sotsiobiologiyasi. 26 (1): 47–56. doi:10.1007/BF00174024. S2CID 25177326.
- ^ Garamszegi, L. Z.; A. P. Møller; János Török; Gábor Michl; Péter Péczely; Murielle Richard (2004). "Immune challenge mediates vocal communication in a passerine bird: an experiment" (PDF). Xulq-atvor ekologiyasi. 15 (1): 148–157. doi:10.1093/beheco/arg108.
- ^ Redpath, S. M.; Bridget M Appleby; Steve J Petty (2000). "Do male hoots betray parasite loads in Tawny Owls?". Qushlar biologiyasi jurnali. 31 (4): 457–462. doi:10.1034/j.1600-048X.2000.310404.x.
- ^ Reid, J. M.; Peter Arcese; Alice L. E. V. Cassidy; Sara M. Hiebert; James N. M. Smith; Philip K. Stoddard; Amy B. Marr & Lukas F. Keller (2005). "Fitness Correlates of Song Repertoire Size in Free-Living Song Sparrows (Melospiza melodiasi)" (PDF). Amerikalik tabiatshunos. 165 (3): 299–310. doi:10.1086/428299. PMID 15729661. S2CID 12547933.
- ^ a b Møller AP; J. Erritzøe; L. Z. Garamszegi (2005). "Covariation between brain size and immunity in birds: implications for brain size evolution" (PDF). Evolyutsion biologiya jurnali. 18 (1): 223–237. CiteSeerX 10.1.1.585.3938. doi:10.1111/j.1420-9101.2004.00805.x. PMID 15669979. S2CID 21763448.
- ^ a b Searcy, W. A.; Beecher, M.D. (2009). "Song as an aggressive signal in songbirds". Hayvonlar harakati. 78 (6): 1281–1292. doi:10.1016/j.anbehav.2009.08.011. S2CID 30360474.
- ^ Falls, J. B.; Krebs, J. R.; Makgregor, P.K. (1982). "Song matching in the great tit (Parus major) the effect of similarity and familiarity". Hayvonlar harakati. 30 (4): 997–1009. doi:10.1016/S0003-3472(82)80188-7. S2CID 53189625.
- ^ a b Beecher, MD; Stoddard, P.K .; Cambell, E.S.; Horning, C.L. (1996). "Repertoire matching between neighbouring song sparrows". Hayvonlar harakati. 51 (4): 917–923. doi:10.1006/anbe.1996.0095. S2CID 26372750.
- ^ Hill, S. D.; Brunton, D. H.; Anderson, M. A .; Weihong, J. (2018). "Fighting talk: complex song elicits more aggressive responses in a vocally complex songbird". Ibis. 160 (2): 257–268. doi:10.1111/ibi.12542.
- ^ "A Bird's World: Speaking in a Bird's Language". Boston Ilmiy muzeyi. 2008 yil.
- ^ Marler, P. (1955). "Characteristics of some animal calls". Tabiat. 176 (4470): 6–8. Bibcode:1955Natur.176....6M. doi:10.1038/176006a0. S2CID 4199385.
- ^ Lengagne, T.; J. Lauga & T. Aubin (2001). "Intra-syllabic acoustic signatures used by the King Penguin in parent-chick recognition: an experimental approach" (PDF). Eksperimental biologiya jurnali. 204 (Pt 4): 663–672. PMID 11171348.
- ^ Wayne Delport; Alan C Kemp; J. Willem H Ferguson (2002). "Vocal identification of individual African Wood Owls Strix woodfordii: a technique to monitor long-term adult turnover and residency". Ibis. 144 (1): 30–39. doi:10.1046/j.0019-1019.2001.00019.x.
- ^ Thorpe, W. H. (23 February 1963). "Antiphonal Singing in Birds as Evidence for Avian Auditory Reaction Time". Tabiat. 197 (4869): 774–776. Bibcode:1963Natur.197..774T. doi:10.1038/197774a0. S2CID 30542781.
- ^ Stokes, A. W.; H. W. Williams (1968). "Antiphonal calling in quail" (PDF). Auk. 85 (1): 83–89. doi:10.2307/4083626. JSTOR 4083626.
- ^ Xarris, Toni; Franklin, Kim (2000). Shriklar va Bush-Shrikes. Prinston universiteti matbuoti. 257-260 betlar. ISBN 978-0-691-07036-0.
- ^ Osmaston, B. B. (1941). ""Duetting" in birds". Ibis. 5 (2): 310–311. doi:10.1111/j.1474-919X.1941.tb00620.x.
- ^ Power, D. M. (1966). "Antiphonal duetting and evidence for auditory reaction time in the Orange-chinned Parakeet". Auk. 83 (2): 314–319. doi:10.2307/4083033. JSTOR 4083033.
- ^ Hyman, Jeremy (2003). "Countersinging as a signal of aggression in a territorial songbird" (PDF). Hayvonlar harakati. 65 (6): 1179–1185. doi:10.1006/anbe.2003.2175. S2CID 38239656.
- ^ Betts, M.G.; Hadley, A.S.; Rodenhouse, N.; Nocera, J.J. (2008). "Social Information Trumps Vegetation Structure in Breeding-Site Selection by a Migrant Songbird". Ish yuritish: Biologiya fanlari. 1648. 275 (1648): 2257–2263. doi:10.1098/rspb.2008.0217. PMC 2603235. PMID 18559326.
- ^ Goodale, E. & Kotagama, S. W. (2005). "Testing the roles of species in mixed-species bird flocks of a Sri Lankan rain forest". Tropik ekologiya jurnali. 21 (6): 669–676. doi:10.1017/S0266467405002609.
- ^ Kelley, L. A.; Coe, R. L.; Madden, J. R.; Healy, S. D. (2008). "Vocal mimicry in songbirds". Hayvonlar harakati. 76 (3): 521–528. doi:10.1016/j.anbehav.2008.04.012. S2CID 53192695.
- ^ Marler, Piter; Hans Willem Slabbekoorn (2004). Nature's music: the science of birdsong. Akademik matbuot. p. 145. ISBN 978-0-12-473070-0.
- ^ Suthers RA & Hector DH (1985). "The physiology of vocalization by the echolocating Oilbird, Steatornis caripensis". J. Komp. Fiziol. 156 (2): 243–266. doi:10.1007/BF00610867. S2CID 1279919.
- ^ Suthers RA & Hector DH (1982). "Mechanism for the production of echolocating clicks by the Grey Swiftlet, Kollokaliya spodiopigiyasi". J. Komp. Fiziol. A. 148 (4): 457–470. doi:10.1007/BF00619784. S2CID 39111110.
- ^ Coles RB; Konishi M & Pettigrew JD (1987). "Hearing and echolocation in the Australian Grey Swiftlet, Kollokaliya spodiopigiyasi". J. Exp. Biol. 129: 365–371.
- ^ Lieser M; P. Berthold1 & G. A. Manley (2005). "Infrasound in the capercaillie ( Tetrao urogallus )". Ornitologiya jurnali. 146 (4): 395–398. doi:10.1007/s10336-005-0003-y. S2CID 22412727.
- ^ Dooling, R.J. (1982). Auditory perception in birds. Acoustic Communication in Birds, Vol. 1 (eds D.E. Kroodsma & E.H. Miller). 95-130 betlar.
- ^ Olson, Christopher R.; Fernández-Vargas, Marcela; Portfors, Christine V.; Mello, Claudio V. (2018). "Black Jacobin hummingbirds vocalize above the known hearing range of birds". Hozirgi biologiya. 28 (5): R204–R205. doi:10.1016/j.cub.2018.01.041. PMID 29510104. S2CID 3727714.
- ^ Derryberry, Elizabeth (July 2009). "Ecology Shapes Birdsong Evolution: Variation in Morphology and Habitat Explains Variation in White-Crowned Sparrow Song". Amerikalik tabiatshunos. 174 (1): 24–33. doi:10.1086/599298. PMID 19441960. S2CID 8606774.
- ^ Boncoraglio, G. & Nicola Saino (2007). "Habitat structure and the evolution of bird song: a meta-analysis of the evidence for the acoustic adaptation hypothesis". Funktsional ekologiya. 21: 134–142. doi:10.1111/j.1365-2435.2006.01207.x.
- ^ Morton, E.S. (1975). "Ecological sources of selection on avian sounds". Amerikalik tabiatshunos. 109 (965): 17–34. doi:10.1086/282971. S2CID 55261842.
- ^ Ey, Elodie; Fischer, J. (13 April 2012). "The "acoustic adaptation hypothesis" - a review of the evidence from birds, anurans and mammals". Bioakustika. 19 (1–2): 21–48. doi:10.1080/09524622.2009.9753613. S2CID 84971439.
- ^ Tubaro, Pablo L.; Segura, Enrique T. (November 1994). "Dialect Differences in the Song of Zonotrichia capensis in the Southern Pampas: A Test of the Acoustic Adaptation Hypothesis". Kondor. 96 (4): 1084–1088. doi:10.2307/1369117. JSTOR 1369117.
- ^ Slabbekoorn, Xans; Ellers, Jacintha; Smit, Tomas B. (2002). "Birdsong and sound transmission: the benefits of reverberations" (PDF). Kondor. 104 (3): 564–573. doi:10.1650 / 0010-5422 (2002) 104 [0564: basttb] 2.0.co; 2.
- ^ Krause, Bernard L. (1993). "The Niche Hypothesis" (PDF). The Soundscape Newsletter. 06. Arxivlandi asl nusxasi (PDF) 2008-03-07 da.
- ^ Henrik Brumm (2004). "Hududiy qushda qo'shiq amplitudasiga atrof-muhit shovqinining ta'siri". Hayvonlar ekologiyasi jurnali. 73 (3): 434–440. doi:10.1111 / j.0021-8790.2004.00814.x. S2CID 73714706.
- ^ Slabbekoorn, H. & Peet, M. (2003). "Qushlar shahar shovqinida balandroq balandlikda qo'shiq aytishadi". Tabiat. 424 (6946): 267. Bibcode:2003 yil Noyabr.424..267S. doi:10.1038 / 424267a. PMID 12867967. S2CID 4348883.
- ^ Halfwerk, Wouter; Holleman, L.J.M.; Lessells, C.M.; Slabbekoorn, H. (February 2011). "Negative impact of traffic nosie on avian reproductive success". Amaliy ekologiya jurnali. 48 (1): 210–219. doi:10.1111 / j.1365-2664.2010.01914.x. S2CID 83619284.
- ^ Luther, David A.; Derryberry, E.P. (Aprel 2012). "Birdsongs keep pace with city life: changes in song over time in an urban songbird affects communication". Hayvonlar harakati. 83 (4): 1059–1066. doi:10.1016/j.anbehav.2012.01.034. S2CID 31212627.
- ^ Najar, N.; Benedict, L. (2019). "The relationship between latitude, migration and the evolution of bird song complexity". Ibis. 161 (1): 1–12. doi:10.1111 / ibi.12648.
- ^ Jeffrey Podos; Mario Cohn-Haft (21 October 2019). "Extremely loud mating songs at close range in white bellbirds". Hozirgi biologiya. 29 (20): R1068–R1069. doi:10.1016/j.cub.2019.09.028. PMID 31639347. S2CID 204823663.
- ^ "World's 'loudest bird': Meet the white bellbird". Yangiliklar (video). BBC yangiliklari. 22 oktyabr 2019 yil.
- ^ NEMETH, ERWIN (2004-01-01). "Measuring the Sound Pressure Level of the Song of the Screaming Piha Lipaugus Vociferans: One of the Loudest Birds in the World?". Bioakustika. 14 (3): 225–228. doi:10.1080/09524622.2004.9753527. ISSN 0952-4622. S2CID 84218370.
- ^ a b v d e Nottebohm, F. (2005). "The Neural Basis of Birdsong". PLOS Biol. 3 (5): 163. doi:10.1371/journal.pbio.0030164. PMC 1110917. PMID 15884976.
- ^ Brainard, M. S. & Doupe, A. J. (2000). "Auditory feedback in learning and maintenance of vocal behavior". Neuroscience-ning tabiat sharhlari. 1 (1): 31–40. doi:10.1038/35036205. PMID 11252766. S2CID 5133196.
- ^ Carew, Thomas J. (2000). Xulq-atvor nevrologiyasi: tabiiy xatti-harakatlarning uyali tashkiloti. Sinauer Associates. ISBN 978-0-87893-092-0.
- ^ a b Kao, M.H.; Doupe, A.J.; Brainard, M.S. (2005). "Contributions of an avian basal ganglia-forebrain circuit to real=time modulation of song". Tabiat. 433 (7026): 638–642. Bibcode:2005Natur.433..638K. doi:10.1038/nature03127. PMID 15703748. S2CID 4352436.
- ^ Suthers, R. (2004). "How birds sing and why it matters". In Marler, P.; Slabbekoorn, H. (eds.). Nature's music:The science of birdsong. Akademik matbuot. 272–295 betlar. ISBN 978-0-12-473070-0.
- ^ a b Brainard, M. S. & Doupe, A. J. (2000). "Interruption of a basal ganglia-forebrain circuit prevents plasticity of learned vocalizations". Tabiat. 404 (6779): 762–766. Bibcode:2000Natur.404..762B. doi:10.1038/35008083. PMID 10783889. S2CID 4413588.
- ^ Kojima, S.; Doupe, A. (2008). "Neural encoding of auditory temporal context in a songbird basal ganglia nucleus, and its independence of birds' song experience". Evropa nevrologiya jurnali. 27 (5): 1231–1244. doi:10.1111/j.1460-9568.2008.06083.x. PMC 2408885. PMID 18364039.
- ^ Long, M.A .; Jin, D.Z.; Fee, M.S. (2010). "Support for a synaptic chain model of neuronal sequence generation". Tabiat. 468 (7322): 394–399. Bibcode:2010Natur.468..394L. doi:10.1038/nature09514. PMC 2998755. PMID 20972420.
- ^ a b Balthazart, J.; Adkins-Regan (2002). Sexual differentiation of brain and behavior in birds. Hormones, Brain and Behavior. 4. pp. 223–301. doi:10.1016/b978-012532104-4/50068-8. ISBN 9780125321044. PMID 18406680.
- ^ Nottebohm, F. & Arnold, A.P. (1976). "Qo'shiq miyasi vokalni boshqarish sohalarida jinsiy dimorfizm". Ilm-fan. 194 (4261): 211–213. Bibcode:1976Sci...194..211N. doi:10.1126 / science.959852. PMID 959852.
- ^ Gurney, M.E. & Konishi, M. (1980). "Hormone-induced sexual differentiation of brain and behavior in zebra finches". Ilm-fan. 208 (4450): 1380–1383. Bibcode:1980Sci...208.1380G. doi:10.1126/science.208.4450.1380. PMID 17775725. S2CID 11669349.
- ^ Tomaszycki, M.L.; Peabody, C.; Replogle, K.; Clayton, D.F; Tempelman, R.J.; Wade, J. (2009). "Sexual differentiation of the zebra finch song system: potential roles for sex chromosome genes". BMC nevrologiyasi. 10: 24. doi:10.1186/1471-2202-10-24. PMC 2664819. PMID 19309515.
- ^ Leonard, S. L. (1 May 1939). "Induction of Singing in Female Canaries by Injections of Male Hormone". Eksperimental biologiya va tibbiyot. 41 (1): 229–230. doi:10.3181/00379727-41-10631. S2CID 87078020.
- ^ Nottebohm, F. (1980). "Testosterone triggers growth of brain vocal control nuclei in adult female canaries". Miya tadqiqotlari. 189 (2): 429–36. doi:10.1016/0006-8993(80)90102-X. PMID 7370785. S2CID 25845332.
- ^ Ball, G.F. & Balthazart, J. (2002). Neuroendocrine mechanisms regulating reproductive cycles and reproductive behavior in birds. Gormonlar, miya va o'zini tutish. 2. pp. 649–798. doi:10.1016/b978-012532104-4/50034-2. ISBN 9780125321044.
- ^ Bentley, G.E.; Van’t Hof, T.J.; Ball, G.F. (1999). "Seasonal neuroplasticity in the songbird telencephalon: A role for melatonin". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 96 (8): 4674–4679. Bibcode:1999PNAS...96.4674B. doi:10.1073/pnas.96.8.4674. PMC 16391. PMID 10200321.
- ^ Cassone, V.M.; Bartell, P.A.; Earnest D.J. & Kumar, V. (2008). "Duration of melatonin regulates seasonal changes in song control nuclei of the house sparrow, Passer domesticus: Independence from gonads and circadian entrainment". Biologik ritmlar jurnali. 23 (1): 49–58. doi:10.1177/0748730407311110. PMID 18258757. S2CID 206544790.
- ^ Ball, G.F.; Auger, C.J.; Bernard, D.J.; Charlier, T.D.; Sartor, J.J.; Riters, L.V.; Balthazart, J. (2004). "Seasonal plasticity in the song control system: Multiple brain sites of steroid hormone action and the importance of variation in song behavior". Nyu-York Fanlar akademiyasining yilnomalari. 1016 (1): 586–610. Bibcode:2004NYASA1016..586B. doi:10.1196/annals.1298.043. PMID 15313796. S2CID 42818488.
- ^ London, S.E.; Replogle, K.; Clayton, D.F. (2009). "Developmental shifts in gene expression in the auditory forebrain during the sensitive period for song learning". Rivojlanish neyrobiologiyasi. 69 (7): 436–450. doi:10.1002/dneu.20719. PMC 2765821. PMID 19360720.
- ^ Scharff C; Haesler S (2005). "An evolutionary perspective on FoxP2: strictly for the birds?". Neyrobiologiyaning hozirgi fikri. 15 (6): 694–703. doi:10.1016/j.conb.2005.10.004. PMID 16266802. S2CID 11350165.
- ^ a b Brainard, M. S. & Doupe, A. J. (2002). "What songbirds teach us about learning". Tabiat. 417 (6886): 351–358. Bibcode:2002Natur.417..351B. doi:10.1038/417351a. PMID 12015616. S2CID 4329603.
- ^ Barrington, D. (1773). "Experiments and observations on the singing of birds". Qirollik jamiyatining falsafiy operatsiyalari. 63: 249–291. doi:10.1098/rstl.1773.0031. S2CID 186207885.
- ^ Marler, P .; M. Tamura (1962). "Song dialects in three populations of the white-crowned sparrow". Kondor. 64 (5): 368–377. doi:10.2307/1365545. JSTOR 1365545.
- ^ Konishi, M. (2010). "From central pattern generator to sensory template in the evolution of birdsong". Miya va til. 115 (1): 18–20. doi:10.1016/j.bandl.2010.05.001. PMID 20955898. S2CID 205791930.
- ^ a b Leonardo, A.; Konishi, M. (1999). "Decrystallization of adult birdsong by perturbation of auditory feedback". Tabiat. 399 (6735): 466–470. Bibcode:1999Natur.399..466L. doi:10.1038/20933. PMID 10365958. S2CID 4403659.
- ^ Teramitsu I; Kudo LC; London SE; Geschwind DH & White SA (2004). "Parallel FoxP1 and FoxP2 expression in songbird and human brain predicts functional interaction". J. Neurosci. 24 (13): 3152–63. doi:10.1523/JNEUROSCI.5589-03.2004. PMC 6730014. PMID 15056695.
- ^ Nottebohm, F. (2004). "The road we travelled: discovery, choreography, and significance of brain replaceable neurons". Nyu-York Fanlar akademiyasining yilnomalari. 1016 (1): 628–658. Bibcode:2004NYASA1016..628N. doi:10.1196/annals.1298.027. PMID 15313798. S2CID 11828091.
- ^ Brenowitz, Eliot A. & Michael D. Beecher (2005). "Song learning in birds: diversity and plasticity, opportunities and challenges" (PDF). Nörobilimlerin tendentsiyalari. 28 (3): 127–132. doi:10.1016/j.tins.2005.01.004. PMID 15749165. S2CID 14586913.
- ^ Slater, P. J. B. (1989). "Bird song learning: causes and consequences". Etol. Ekol. Evol. 1: 19–46. doi:10.1080/08927014.1989.9525529.
- ^ Thorpe, W. (1954). "The process of song-learning in the chaffinch as studied by means of the sound spectrograph". Tabiat. 173 (4402): 465–469. Bibcode:1954Natur.173..465T. doi:10.1038/173465a0. S2CID 4177465.
- ^ Metzmaxer, M. (2016). "Alauda: Chaffinch song learning : Thorpe conclusions revisited". Alauda. 84: 465–469. hdl:2268/204189.
- ^ Konishi, M. (1965). "The role of auditory feedback on the control of vocalization in the white-crowned sparrow". Zeitschrift für Tierpsychologie. 22 (7): 770–783. doi:10.1111/j.1439-0310.1965.tb01688.x (nofaol 2020-11-09). PMID 5874921.CS1 maint: DOI 2020 yil noyabr holatiga ko'ra faol emas (havola)
- ^ Marler, P. (1970). "A comparative approach to vocal learning: Song development in the white-crowned sparrows". Qiyosiy va fiziologik psixologiya jurnali. 71 (2, Pt.2): 1–25. doi:10.1037/h0029144.
- ^ Nordeen, K.W.; Nordeen, E.J. (1994). "Auditory feedback is necessary for the maintenance of stereotyped song in adult zebra finches". Xulq-atvor va asab biologiyasi. 71 (1): 58–66. doi:10.1016/0163-1047(92)90757-U. PMID 1567334.
- ^ Leonardo, A. (2004). "Experimental test of error-correction birdsong model". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 101 (48): 16935–16940. doi:10.1073/pnas.0407870101. PMC 534752. PMID 15557558.
- ^ a b Rizzolatti G.; Craighero L. (2004). "The mirror-neuron system". Annu. Vahiy Neurosci. 27: 169–192. doi:10.1146 / annurev.neuro.27.070203.144230. PMID 15217330. S2CID 1729870.
- ^ Oberman L. M.; Pineda J. A.; Ramachandran V. S. (2007). "Insonning ko'zgu neyron tizimi: harakatni kuzatish va ijtimoiy ko'nikmalar o'rtasidagi bog'liqlik". Ijtimoiy kognitiv va ta'sirchan nevrologiya. 2 (1): 62–66. doi:10.1093 / scan / nsl022. PMC 2555434. PMID 18985120.
- ^ a b Prather J. F.; Peters S.; Nowicki S.; Mooney R. (2008). "Precise auditory-vocal mirroring in neurons for learned vocal communication". Tabiat. 451 (7176): 305–310. Bibcode:2008Natur.451..305P. doi:10.1038/nature06492. PMID 18202651. S2CID 4344150.
- ^ a b Dinstein I.; Thomas C.; Behrmann M.; Heeger D.J. (2008). "A mirror up to nature". Hozirgi biologiya. 18 (1): R13–18. doi:10.1016/j.cub.2007.11.004. PMC 2517574. PMID 18177704.
- ^ Tchernichovski O.; Wallman J. (2008). "Behavioral neuroscience: Neurons of imitation". Tabiat. 451 (7176): 249–250. Bibcode:2008Natur.451..249T. doi:10.1038/451249a. PMID 18202627. S2CID 205035217.
- ^ Miller G (2008). "Mirror neurons may help songbirds stay in tune". Ilm-fan. 319 (5861): 269. doi:10.1126/science.319.5861.269a. PMID 18202262. S2CID 34367648.
- ^ Richard Mooney (5 June 2014). "Auditory–vocal mirroring in songbirds" (PDF). Philosophical Transactions of the Royal Society B: Biological Sciences Online. Olingan 16 dekabr, 2015.[doimiy o'lik havola ]
- ^ a b v Riebel, Katharina; Lachlan, Robert F.; Slater, Peter J. B. (2015-05-01), Naguib, Marc; Brockmann, H. Jane; Mitani, Jon S.; Simmons, Leigh W. (eds.), "Chapter Six - Learning and Cultural Transmission in Chaffinch Song", Xulq-atvorni o'rganishdagi yutuqlar, Academic Press, 47, pp. 181–227, doi:10.1016/bs.asb.2015.01.001, olingan 2020-01-30
- ^ Thorpe, W. H. (2008-06-28). "The Learning of Song Patterns by Birds, with Especial Reference to the Song of the Chaffinch Fringilla Coelebs". Ibis. 100 (4): 535–570. doi:10.1111/j.1474-919x.1958.tb07960.x. ISSN 0019-1019.
- ^ Slater, P. J. B. (1983-04-01). "Chaffinch Imitates Canary Song Elements and Aspects of Organization". Auk. 100 (2): 493–495. doi:10.1093/auk/100.2.493. ISSN 0004-8038.
- ^ Slater, P. J. B.; Ince, S. A. (1979). "Cultural Evolution in Chaffinch Song". Xulq-atvor. 71 (1/2): 146–166. doi:10.1163/156853979X00142. ISSN 0005-7959. JSTOR 4534000.
- ^ Tanaka, Masashi; Sun, Fangmiao; Li, Yulong; Mooney, Richard (2018). "A mesocortical dopamine circuit enables the cultural transmission of vocal behaviour". Tabiat. 563 (7729): 117–120. Bibcode:2018Natur.563..117T. doi:10.1038/s41586-018-0636-7. ISSN 1476-4687. PMC 6219627. PMID 30333629.
- ^ a b v Lachlan, Robert F.; Slater, Peter J. B. (1999-04-07). "The maintenance of vocal learning by gene–culture interaction: the cultural trap hypothesis". Qirollik jamiyati materiallari B: Biologiya fanlari. 266 (1420): 701–706. doi:10.1098/rspb.1999.0692. ISSN 0962-8452. PMC 1689831.
- ^ Schmidt, K. L.; MacDougall-Shackleton, E. A.; Kubli, S. P.; MacDougall-Shackleton, S. A. (2014-06-20). "Developmental Stress, Condition, and Birdsong: A Case Study in Song Sparrows". Integrativ va qiyosiy biologiya. 54 (4): 568–577. doi:10.1093/icb/icu090. ISSN 1540-7063. PMID 24951504.
- ^ S., Nowicki; W., Searcy; S., Peters (2002-12-01). "Brain development, song learning and mate choice in birds: a review and experimental test of the "nutritional stress hypothesis"". Qiyosiy fiziologiya jurnali A. 188 (11–12): 1003–1014. doi:10.1007/s00359-002-0361-3. ISSN 0340-7594. PMID 12471497. S2CID 14298372.
- ^ Boogert, N. J .; Fokett, T. V.; Lefebvre, L. (2011-04-18). "Kognitiv xususiyatlar uchun turmush o'rtog'ini tanlash: g'ayriinsoniy umurtqali hayvonlar dalillarini ko'rib chiqish". Xulq-atvor ekologiyasi. 22 (3): 447–459. doi:10.1093 / beheco / arq173. ISSN 1045-2249.
- ^ Boogert, Neeltje J.; Anderson, Rindi S.; Piters, Syuzan; Searcy, William A.; Nowicki, Stephen (2011). "Song repertoire size in male song sparrows correlates with detour reaching, but not with other cognitive measures". Hayvonlar harakati. 81 (6): 1209–1216. doi:10.1016/j.anbehav.2011.03.004. ISSN 0003-3472. S2CID 21724914.
- ^ Saunders, Aretas A (1951) Guide to Bird Songs. Doubleday and Company
- ^ Sibli, Devid (2000). Sibley qushlar uchun qo'llanma. Knopf. ISBN 978-0-679-45122-8.
- ^ Sarvasy, Hannah (2016). "Warblish: Verbal Mimicry of Birdsong". Etnobiologiya jurnali. 36 (4): 765–782. doi:10.2993/0278-0771-36.4.765. hdl:1885/112092. ISSN 0278-0771. S2CID 89990174.
- ^ Baker, Myron C. (2001). "Bird Song Research: The Past 100 years" (PDF). Qushlarning harakati. 14: 3–50.
- ^ Bailey, C. E. G (2008). "Towards an Orthography of Bird Song". Ibis. 92: 115–131. doi:10.1111/j.1474-919X.1950.tb01739.x.
- ^ Borror, Donald J.; Riz, Karl R. (1953). "Vibralayzer vositasida qushlarning qo'shiqlarini tahlil qilish" (PDF). Uilson byulleteni. 65 (4): 271–276.
- ^ Thorpe, W. H. (1958). "The learning of song patterns by birds, with especial reference to the song of the chaffinch Fringilla coelebs". Ibis. 100 (4): 535–570. doi:10.1111 / j.1474-919X.1958.tb07960.x.
- ^ Slater, P. J. B. (2003). "Fifty years of bird song research: a case study in animal behaviour". Hayvonlar harakati. 65 (4): 633–639. doi:10.1006/anbe.2003.2051. S2CID 53157104.
- ^ Robbins, Chandler S.; Bertel Bruun; Herbert S. Zim; Arthur Singer (1983). A Guide To Field Identification: Birds of North America. Golden Field Guides (Second ed.). G'arbiy nashriyot kompaniyasi. p.14. ISBN 978-0-307-33656-9.
- ^ Meijer, P.B.L. (1992). "An Experimental System for Auditory Image Representations". Biomedikal muhandislik bo'yicha IEEE operatsiyalari. 39 (2): 112–121. doi:10.1109/10.121642. PMID 1612614. S2CID 34811735.
- ^ "US Patent. 20030216649. Audible output sonogram analyzer". Freepatentsonline.com. 2003-11-20. Olingan 2014-06-03.
- ^ Alström, P .; Ranft, R. (2003). "The use of sounds in avian systematics, and the importance of bird sound archives". Britaniya ornitologlar klubi byulleteni. 123A: 114–135.
- ^ Alström, P. (1998). "Mirafra assamika kompleksi taksonomiyasi" (PDF). Forktail. 13: 97-107. Arxivlandi asl nusxasi (PDF) 2008-03-07 da.
- ^ "App - Bird Song Id USA: Nature Apps - Sunbird". sunbird.tv. Olingan 2016-06-08.
- ^ Munford, Monty (2015-03-28). "Don't know birdsong? There's a (great) app for that". Telegraph.co.uk. Olingan 2016-06-08.
- ^ Collias, N. E. (1987). "The vocal repertoire of the Red Junglefowl: A spectrographic classification and the code of communication". Kondor. 89 (3): 510–524. doi:10.2307/1368641. JSTOR 1368641. S2CID 87662926.
- ^ Evans, S.S .; Makedoniya, J. M .; Marler, P. (1993). "Effects of apparent size and speed on the response of chickens, Gallus gallus, to computer-generated simulations of aerial predators". Hayvonlar harakati. 46: 1–11. doi:10.1006/anbe.1993.1156. S2CID 53197810.
- ^ a b Hailman, Jack; Ficken, Millicent (1986). "Combinatorial animal communication with computable syntax: Chick-a-dee calling qualifies as "language" by structural linguistics". Hayvonlar harakati. 34 (6): 1899–1901. doi:10.1016/S0003-3472(86)80279-2. S2CID 53172611.
- ^ Pepperberg, I.M. (2000). Aleks tadqiqotlari: kulrang to'tiqushlarning kognitiv va kommunikativ qobiliyatlari. Garvard universiteti matbuoti.
- ^ Marcus, Gary F. (2006-04-27). "Language: Startling starlings". Tabiat. 440 (7088): 1117–1118. Bibcode:2006Natur.440.1117M. doi:10.1038/4401117a. PMID 16641976. S2CID 14191866.
- ^ Young, Jon (2008). "Bird Language: Exploring the Language of Nature with Jon Young". OWLink Media. Arxivlandi asl nusxasi 2010-07-28 da.
- ^ Birds Can Tell Us a Lot about Human Language Adam Fishbein, February 2, 2018 (Ilmiy Amerika )
- ^ Birds use language like humans joining calls together to form sentences, March 8, 2015 The Daily Telegraph (First published in Nature Communications)
- ^ a b "Archive Pioneers - Ludwig Koch and the Music of Nature". BBC Arxivlari. BBC. 2009-04-15. Olingan 2 sentyabr 2011.
- ^ Matthew Head (1997). "Qushlarning qo'shig'i va musiqaning kelib chiqishi". Qirollik musiqiy assotsiatsiyasi jurnali. 122 (1): 1–23. doi:10.1093 / jrma / 122.1.1.
- ^ Klark, Suzanna (2001). Music Theory and Natural Order from the Renaissance to the Early Twentieth Century. Kembrij universiteti matbuoti. ISBN 978-0-521-77191-7.
- ^ Reyx, Ronni (2010 yil 15 oktyabr). "NJIT professori bizning tukli do'stlarimizni tinchlantirishda kuku topolmaydi". Yulduzli kitob. Olingan 19 iyun 2011.
- ^ Taylor, Hollis (2011-03-21). "Bastakor Butcherbird qo'shig'ining bastakorlari tomonidan ajratilishi: Genri Teytning" Avstraliyaning pastki qo'shig'i "yoshi keladi". Onlayn musiqa tadqiqotlari jurnali. 2.
- ^ Rothenberg, David (2005). Nima uchun qushlar qo'shiq aytishadi. Allen Leyn.
- ^ Motion, Andrew (10 December 2005). "To'liq reysda". Guardian. Olingan 24 aprel 2016.
- ^ "Nima uchun qushlar qo'shiq aytishadi". British Broadcasting Corporation (BBC to'rtligi). 2010 yil 1-noyabr. Olingan 24 aprel 2016.
- ^ Andervud, Emili (2016 yil 15-avgust). "Axir, Birds qo'shig'i musiqa emas". Ilm-fan. Olingan 24 aprel 2016.
- ^ Baptista, Luis Felipe; Keister, Robin A. (2005). "Nega Qushlarning qo'shig'i ba'zida musiqaga o'xshaydi". Biologiya va tibbiyotning istiqbollari. 48 (3): 426–443. doi:10.1353 / pbm.2005.0066. PMID 16085998. S2CID 38108417.
- ^ Teylor, Xollis (2017). "Is Birdsong Music? Outback Encounters with an Australian Songbird". Indiana universiteti matbuoti. Olingan 2017-06-25.
- ^ Tierni, Adam T.; Russo, Frank A.; Patel, Aniruddh D. (2011). "Inson va parranda qo'shiqlari tuzilishining motorli kelib chiqishi". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 108 (37): 15510–15515. Bibcode:2011PNAS..10815510T. doi:10.1073 / pnas.1103882108. PMC 3174665. PMID 21876156.
- ^ Attar, Farid al-Din (1984). Darbandi, Afkham; Davis, Dick (eds.). Qushlar konferentsiyasi. Pingvin klassiklari. ISBN 978-0-14-044434-6.
- ^ Brooks, Cleanth; Warren, Robert Penn (1968). Stilinger, Jek (tahrir). The Bulbulga odob. Keats's Odes. Prentice-Hall. 44-47 betlar.
- ^ Sandy, Mark (2002). "To a Skylark". Adabiy entsiklopediya. Olingan 22 aprel 2016.
- ^ "Crow - The Ted Hughes Society Journal". The Ted Hughes Society. 2012. Arxivlangan asl nusxasi 2015 yil 2-iyulda. Olingan 22 aprel 2016.
- ^ Hopkins, Gerard Manley (1985). Poems and Prose. Pingvin kitoblari. ISBN 9780140420159.
Tashqi havolalar
- Avian Vocalizations Center Michigan shtati universiteti
- The Nature Explorers Audio and video of Western North American birds.
- Bird Language: Exploring the Language of Nature with Jon Young A blog with stories and tips for learning the patterns in bird vocalizations.
- Large collection of audio bird calls collected in Arizona dan Biologdan so'rang.
- xeno-canto: Community online database of downloadable bird sounds from around the globe ≈250,000 recordings of ≈9400 species as of Aug 2015. See also kseno-kanto.
- British Library's archive of bird sounds representing more than 8,000 species.
- The Sound Approach A guide to understanding bird sound.
- Listen to Nature includes article "The Language of Birds"
- Bird language articles
- Bird songs in movies: an unnatural history Humor piece on soundtrack errors
- How do Birds Sing? The mechanics and anatomy of birdsong production
- Song Bird Science Shared resource for birdsong scientists
- Bioacoustic Research Program da Kornell ornitologiya laboratoriyasi distributes a number of different free birdsong synthesis & analysis programs.
- Makolay kutubxonasi da Kornell ornitologiya laboratoriyasi is the world's largest collection of animal sounds and associated video.
- Audio Pitch Tracer Accurate transcription of clean recordings of bird vocalizations to midi
| Faoliyat |  | |
|---|---|---|
| Mahsulotlar | ||
| San'atda |
| |
| Turlar | ||
| Odamlar |
| |
| Bog'liq |
| |