Sitoxrom P450 - Cytochrome P450 - Wikipedia
| Sitoxrom P450 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
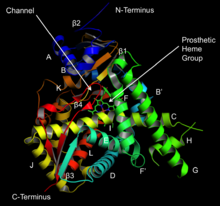 Lanosterol 14a-demetilaza tuzilishi (CYP51 ) | |||||||||
| Identifikatorlar | |||||||||
| Belgilar | p450 | ||||||||
| Pfam | PF00067 | ||||||||
| InterPro | IPR001128 | ||||||||
| PROSITE | PDOC00081 | ||||||||
| SCOP2 | 2cpp / QOIDA / SUPFAM | ||||||||
| OPM superfamily | 39 | ||||||||
| OPM oqsili | 2 mlrd | ||||||||
| Membranom | 265 | ||||||||
| |||||||||
Sitoxromlar P450 (CYP-lar) a superfamily ning fermentlar o'z ichiga olgan heme kabi kofaktor sifatida ishlaydi monooksigenazlar.[1][2][3] Sutemizuvchilarda bu oqsillar oksidlanadi steroidlar, yog 'kislotalari va ksenobiotiklar va uchun muhimdir tozalash turli xil birikmalar, shuningdek gormon sintezi va parchalanishi uchun. O'simliklarda bu oqsillar biosintezi uchun muhimdir himoya aralashmalari yog 'kislotalari va gormonlar.[2]
CYP fermentlari hammasi aniqlangan shohliklar hayot: hayvonlar, o'simliklar, qo'ziqorinlar, protistlar, bakteriyalar va arxey, shuningdek viruslar.[4] Biroq, ular hamma joyda mavjud emas; masalan, ular ichida topilmadi Escherichia coli.[3][5] 2018 yildan boshlab[yangilash], 300000 dan ortiq alohida CYP oqsillari ma'lum.[6][7]
CYP'lar, umuman olganda, terminal oksidazdir fermentlar yilda elektronlar almashinuvi sifatida keng tasniflangan zanjirlar P450 o'z ichiga olgan tizimlar. "P450" atamasi spektrofotometrik cho'qqisi to'lqin uzunligi ning assimilyatsiya maksimal ferment (450nm ) qachon bo'lsa kamaytirilgan holati va murakkabligi uglerod oksidi. Ko'pgina CYP'lar bir yoki bir nechtasini etkazib berish uchun oqsil sherigiga muhtoj elektronlar kamaytirish uchun temir (va oxir-oqibat molekulyar kislorod ).
Nomenklatura
Genlar kodlash CYP fermentlari va fermentlarning o'zi, bilan belgilanadi ildiz belgisi CYP uchun superfamily, keyin raqamni ko'rsatuvchi genlar oilasi, subfamilani ko'rsatadigan katta harf va individual gen uchun boshqa raqam. Konventsiya kursiv genga murojaat qilganda ism. Masalan, CYP2E1 fermentni kodlovchi gendir CYP2E1 - ishtirok etgan fermentlardan biri paratsetamol (asetaminofen) metabolizmi. The CYP nomenklatura - vaqti-vaqti bilan bo'lsa ham rasmiy nomlash konvensiyasi CYP450 yoki CYP450 sinonim sifatida ishlatiladi. Shu bilan birga, CYPlarning ba'zi genlari yoki fermentlari nomlari ushbu nomenklaturadan farq qilishi mumkin, bu katalitik faollikni va substrat sifatida ishlatiladigan birikmaning nomini bildiradi. Bunga misollar kiradi CYP5A1, tromboksan A2 sintaz, qisqartirilgan TBXAS1 (ThromBoXane A2 Synthase 1) va CYP51A1, lanosterol 14-a-demetilaza, ba'zan uning substratiga ko'ra norasmiy ravishda LDM ga qisqartiriladi (Lanosterol) va faollik (D.eMetilatsiya).[8]
Amaldagi nomenklatura bo'yicha ko'rsatmalar shuni ko'rsatadiki, yangi CYP oilalari a'zolari kamida 40% ulushga ega aminokislota subfamila a'zolari kamida 55% aminokislota identifikatoriga ega bo'lishlari kerak. Ikkala asosiy gen nomlarini belgilaydigan va kuzatadigan nomenklatura qo'mitalari mavjud (Sitoxrom P450 bosh sahifasi ) va allel ismlar (CYP Allele nomenklatura qo'mitasi ).[9][10]
Tasnifi
Elektron o'tkazuvchan oqsillarning tabiatiga asoslanib, CYPlarni bir necha guruhlarga bo'lish mumkin:[11]
- Mikrosomal P450 tizimlari
- unda elektronlar o'tkaziladi NADPH orqali sitokrom P450 reduktaza (turli xil CPR, POR yoki CYPOR). Sitoxrom b5 (cyb5) tomonidan kamaytirilgandan so'ng ushbu tizimning quvvatini kamaytirishga yordam berishi mumkin sitoxrom b5 reduktaza (CYB5R).
- Mitoxondrial P450 tizimlari
- ishlaydiganlar adrenodoksin reduktaza va adrenodoksin elektronlarni NADPH dan P450 ga o'tkazish uchun.
- Bakterial P450 tizimlari
- ishlaydigan a ferredoksin reduktaza va a ferredoksin elektronlarni P450 ga o'tkazish uchun.
- CYB5R / cyb5/ P450 tizimlari, unda CYP talab qiladigan ikkala elektron ham sitoxrom b dan kelib chiqadi5.
- FMN / Fd / P450 tizimlari
- dastlab topilgan Rodokok turlari, unda a FMN - domen o'z ichiga olgan reduktaza CYP bilan birlashtirilgan.
- Faqat P450
- tashqi kamaytiradigan quvvatni talab qilmaydigan tizimlar. E'tiborga loyiq bo'lganlar kiradi tromboksan sintezi (CYP5), prostatsiklin sintaz (CYP8) va CYP74A (allen oksidi sintaz ).
P450 sitoxromlari tomonidan katalizlangan eng keng tarqalgan reaktsiya bu a monooksigenaza reaktsiya, masalan, kislorodning bir atomini organik substrat (RH) ning alifatik holatiga kiritish, boshqa kislorod atomi esa kamaytirilgan suvga:
RH + O2 + NADPH + H+ → ROH + H2O + NADP+
Ko'pchilik gidroksillanish reaktsiyalar (qo'shilishi gidroksil guruhlar) CYP fermentlaridan foydalanadi.
Mexanizm
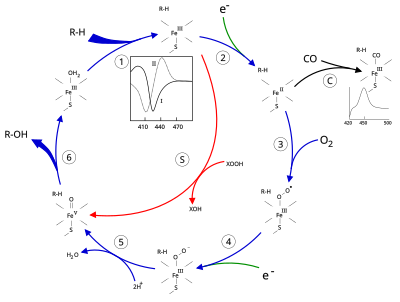
Tuzilishi
Sitoxrom P450 ning faol joyida a mavjud heme - temir markazi. Dazmol a orqali oqsil bilan bog'lanadi sistein tiolat ligand. Ushbu sistein va bir nechta yon qoldiqlar ma'lum CYPlarda yuqori darajada saqlanib qolgan va rasmiydir PROSITE imzoning kelishuv modeli [FW] - [SGNH] - x - [GD] - {F} - [RKHPT] - {P} - C - [LIVMFAP] - [GAD].[12] CYPlar tomonidan katalizlanadigan juda ko'p turli xil reaktsiyalar tufayli, ko'plab CYPlarning faoliyati va xususiyatlari ko'p jihatdan farq qiladi.[13] Umuman olganda, P450 katalitik tsikli quyidagicha davom etadi:
Katalitik tsikl
- Substrat ga yaqin joyda bog'lanadi gem guruhi, eksenel tiolatga qarama-qarshi tomonda. Substratni bog'lash faol joy konformatsiyasining o'zgarishini keltirib chiqaradi, ko'pincha suv molekulasini gem temirining distal eksenel koordinatsion holatidan siljitadi,[14] va gem temirining holatini past spindan yuqori spinga aylantirish.[15]
- Substratni bog'lash NAD (P) H dan elektron uzatishni keltirib chiqaradi sitokrom P450 reduktaza yoki boshqa bog'liq reduktaza.[16]
- Molekulyar kislorod hosil bo'lgan temir gem markaziga distal eksenel koordinatsiya holatida bog'lanib, dastlab a ni beradi dioksigen qo'shimchasi oksi-mioglobindan farqli o'laroq.
- Ikkinchi elektron ikkalasidan ham o'tkaziladi sitokrom P450 reduktaza, ferredoksinlar, yoki sitoxrom b5, Fe-O ni kamaytirish2 qisqa muddatli perokso holatini berish uchun adduct.
- 4-bosqichda hosil bo'lgan perokso guruhi tezda ikki marta protonlanib, bitta molekula suvini chiqarib, yuqori reaktiv turga aylantiradi. P450 aralashmasi 1 (yoki shunchaki aralashma I). Ushbu yuqori reaktiv qidiruv mahsulot 2010 yilda ajratilgan,[17] P450 Murakkab 1 temir (IV) okso (yoki) feribil ) qo'shimcha oksidlovchi ekvivalenti bo'lgan turlar delokalizatsiya qilingan ustidan porfirin va tiolat ligandlari. Muqobil perferril uchun dalillar temir (V) -oxo [14] etishmayapti.[17]
- Qatnashgan substrat va fermentga qarab P450 fermentlari har xil reaktsiyalarning har qanday katalizatoriga aylanishi mumkin. Gipotetik gidroksillanish ushbu rasmda keltirilgan. Mahsulot faol joydan chiqarilgandan so'ng, ferment yana asl holiga keladi, suv molekulasi temir yadrosining distal koordinatsion holatini egallashga qaytadi.

- Mono-oksigenatsiyaning muqobil yo'li "peroksid shunt" orqali amalga oshiriladi (rasmdagi "S" yo'li). Ushbu yo'l temir-substrat kompleksining peroksid va gipoxlorit kabi kislorod-atom donorlari bilan oksidlanishiga olib keladi.[18] Gipotetik peroksid "XOOH" diagrammada ko'rsatilgan.
Spektroskopiya
Substratning bog'lanishi fermentning spektral xususiyatlarida aks etadi, yutilish qobiliyati 390 nm ga ko'tarilib, 420 nm ga kamayadi. Buni farq spektroskopiyalari bilan o'lchash mumkin va "I tip" farq spektri deb nomlanadi (rasmdagi ichki grafikka qarang). Ba'zi substratlar spektral xususiyatlarning qarama-qarshi o'zgarishini, ya'ni "teskari turdagi I" spektrni hali aniq bo'lmagan jarayonlar bilan keltirib chiqaradi. To'g'ridan-to'g'ri gem temiriga bog'langan ingibitorlar va ayrim substratlar II tipdagi farq spektrini keltirib chiqaradi, maksimal 430 nm, eng kami 390 nm (rasmdagi ichki grafikka qarang). Agar kamaytiradigan ekvivalentlar mavjud bo'lmasa, ushbu kompleks barqaror bo'lib qolishi mumkin, bu esa yutilish o'lchovlari bilan bog'lanish darajasini aniqlashga imkon beradi. in vitro[18]C: Agar uglerod oksidi (CO) kamaytirilgan P450 bilan bog'lansa, katalitik tsikl to'xtatiladi. Ushbu reaktsiya klassik CO farq spektrini maksimal 450 nm ga olib keladi.
Odamlarda P450s
Insonning CYP-lari asosan membranalar bilan bog'liq bo'lgan oqsillardir[19] ning ichki membranasida joylashgan mitoxondriya yoki ichida endoplazmatik to'r hujayralar. CYP'lar minglab metabolizmni amalga oshiradi endogen va ekzogen kimyoviy moddalar. Ba'zi CYPlar faqat bitta (yoki juda oz) substratlarni metabolizm qiladi, masalan CYP19 (aromataza ), boshqalari esa metabolizmga uchrashi mumkin substratlar. Ushbu ikkala xususiyat ham ularning markaziy ahamiyatini hisobga oladi Dori. Sitoxrom P450 fermentlari organizmning ko'pgina to'qimalarida mavjud bo'lib, ular tarkibida muhim rol o'ynaydi gormon sintez va buzilish (shu jumladan estrogen va testosteron sintez va metabolizm), xolesterin sintez va D vitamini metabolizm. Sitoxrom P450 fermentlari potentsial toksik birikmalar, shu jumladan metabolizm uchun ham ishlaydi giyohvand moddalar kabi endogen metabolizm mahsulotlari bilirubin, asosan jigar.
The Inson genomining loyihasi turli xil sitokrom P450 fermentlari uchun kodlovchi 57 inson genini aniqladi.[20]
Dori almashinuvi

CYPlar asosiy fermentlardir dori almashinuvi, umumiy metabolizmning taxminan 75% ni tashkil qiladi.[22] Ko'pgina dorilar to'g'ridan-to'g'ri yoki osonlashtirilgan holda CYP tomonidan deaktivatsiyaga uchraydi ajratish tanadan. Bundan tashqari, ko'plab moddalar mavjud bioaktivlangan kabi faol birikmalar hosil qilish uchun CYP tomonidan trombotsitlarga qarshi dori klopidogrel.
Giyohvand moddalarning o'zaro ta'siri
Ko'pgina dorilar izozim biosintezini keltirib chiqarish orqali turli xil CYP izozimlarining faolligini oshirishi yoki kamaytirishi mumkin (fermentlar induksiyasi ) yoki to'g'ridan-to'g'ri CYP faoliyatini inhibe qilish orqali (fermentlarni inhibatsiyasi ). Klassik misol o'z ichiga oladi epileptik preparatlar, kabi Fenitoin, bu esa undaydi CYP1A2, CYP2C9, CYP2C19 va CYP3A4.
CYP izozim faolligiga ta'siri salbiy ta'sir ko'rsatadigan asosiy manba hisoblanadi dorilarning o'zaro ta'siri, chunki CYP fermentlari faolligining o'zgarishi ta'sir qilishi mumkin metabolizm va tozalash turli xil dorilar. Masalan, agar bitta dori boshqa preparatning CYP vositachiligidagi metabolizmni inhibe qilsa, ikkinchi dori tanada toksik darajagacha to'planishi mumkin. Shunday qilib, ushbu dorilarning o'zaro ta'siri dozani o'zgartirishni yoki CYP tizimiga ta'sir qilmaydigan dorilarni tanlashni talab qilishi mumkin. Dori vositalarining bunday o'zaro ta'sirini bemor uchun hayotiy ahamiyatga ega bo'lgan dori-darmonlarni, ahamiyatli dori-darmonlarni qo'llashda e'tiborga olish ayniqsa muhimdir yon effektlar, yoki tor bilan dorilar terapevtik indeks, ammo har qanday dori o'zgargan dori metabolizmi tufayli plazmadagi kontsentratsiyani o'zgartirishi mumkin.
CYP3A4 uchun ko'plab substratlar, masalan, tor terapevtik indeksli dorilar amiodaron[23] yoki karbamazepin.[24] Ushbu dorilar CYP3A4 tomonidan metabolizmga uchraganligi sababli o'rtacha plazma darajalari Ushbu dorilar ferment inhibisyonu tufayli ortishi yoki ferment indüksiyonu tufayli kamayishi mumkin.
Boshqa moddalarning o'zaro ta'siri
Tabiiy ravishda uchraydigan birikmalar CYP faolligini keltirib chiqarishi yoki inhibe qilishi ham mumkin. Masalan, biofaol tarkibidagi birikmalar greyfurt sharbati va boshqa ba'zi meva sharbatlari, shu jumladan bergamottin, dihidroksibbergamottin va paraditsin-A, ning CYP3A4 vositachiligidagi metabolizmni inhibe qilishi aniqlandi ba'zi dorilar, o'sishiga olib keladi bioavailability va, shunday qilib, kuchli imkoniyat dozani oshirib yuborish.[25] Ushbu xavf tufayli, odatda, giyohvand moddalarni iste'mol qilish paytida greypfrut sharbati va yangi greyfurtlardan saqlanish tavsiya etiladi.[26]
Boshqa misollar:
- Sent-Jonning ziravorlari, umumiy o'simlik vositasi keltirib chiqaradi CYP3A4, shuningdek, inhibe qiladi CYP1A1, CYP1B1.[27][28]
- Tamaki chekish keltirib chiqaradi CYP1A2 (masalan, CYP1A2 substratlari klozapin, olanzapin va fluvoksamin )[29]
- Nisbatan yuqori konsentratsiyalarda, yulduz mevasi sharbatning ham tormozlanishi isbotlangan CYP2A6 va boshqa CYP-lar.[30] Suv sarig'i shuningdek, P450 sitoxromining ma'lum inhibitori hisoblanadi CYP2E1, bu ba'zi bir dori-darmonlarda shaxslar uchun dori almashinuvining o'zgarishiga olib kelishi mumkin (masalan, xlorzoksazon ).[31]
- Tributiltin sitrusli P450 funktsiyasini inhibe qilishi aniqlanib, bu mollyuskalarning erkalashishiga olib keladi.[32]
- Goldenseal, ikkita alkaloid bilan ajralib turadi berberin va gidrastin, P450-marker fermentativ faolligini o'zgartirishi ko'rsatilgan (CYP2C9, CYP2D6 va CYP3A4 bilan bog'liq).[33]
Boshqa o'ziga xos CYP funktsiyalari
Steroid gormonlar
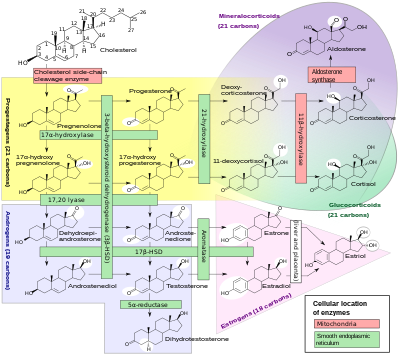
Sitoxrom P450 fermentlarining quyi qismi sintez qilishda muhim rol o'ynaydi steroid gormonlari (steroidogenez ) tomonidan buyrak usti bezlari, jinsiy bezlar va periferik to'qima:
- CYP11A1 (shuningdek, P450scc yoki P450c11a1 deb ham ataladi) buyrak usti bezida mitoxondriya "ilgari 20,22-desmolaza deb nomlangan faollikka" ta'sir qiladi (steroid 20a-gidroksilaza, steroid 22-gidroksilaza, xolesterin yon zanjir scission).
- CYP11B1 (P450c11β oqsilini kodlash) ichki mitoxondriyal membrana ning buyrak usti korteksi steroid 11β-gidroksilaza, steroidga ega 18-gidroksilaza, va steroid 18-metiloksidaza faoliyati.
- CYP11B2 (P450c11AS oqsilini kodlash), faqat buyrak usti mitoxondriyasida uchraydi zona glomerulosa, steroid 11b-gidroksilaza, steroid 18-gidroksilaza va steroid 18-metiloksidaza faolligiga ega.
- CYP17A1, buyrak usti korteksining endoplazmatik to'rida steroid 17a-gidroksilaza va 17,20-liaza faolligiga ega.
- CYP21A2 (P450c21) buyrak usti korteksida 21-gidroksilaza faoliyat.
- CYP19A (P450arom, aromataza ) ichida endoplazmatik to'r ning jinsiy bezlar, miya, yog 'to'qimasi va boshqa joylarda aromatizatsiyani katalizlaydi androgenlar ga estrogenlar.
Ko'p to'yinmagan yog'li kislotalar va eikosanoidlar
Metabolizmada ba'zi bir sitoxrom P450 fermentlari juda muhimdir ko'p to'yinmagan yog 'kislotalari (PUFA) biologik faol, hujayralararo hujayra signalizatsiyasi molekulalar (eikosanoidlar ) va / yoki PUFA biologik faol metabolitlarini kam faol yoki faol bo'lmagan mahsulotlarga aylantirish. Ushbu CYP-larga ega sitoxrom P450 omega gidroksilaza va / yoki epoksigenaza ferment faolligi.
- CYP1A1, CYP1A2 va CYP2E1 signal beruvchi molekulalarga endogen PUFAlarni metabolize qiladi: ular metabolizmga kirishadi arakidon kislotasi (ya'ni AA) dan 19-gidroksyeikosatetraenoik kislotaga (ya'ni 19-HETE; qarang 20-gidroksiekosatetraenoik kislota ); eikosapentaenoik kislota (ya'ni EPA) ga epoksiikosatetraenoik kislotalar (ya'ni EEQ); va dokosaheksaenoik kislota (ya'ni DHA) ga epoksidokosapentaenoik kislotalar (ya'ni EDP).
- CYP2C8, CYP2C9, CYP2C18, CYP2C19 va CYP2J2 signal beruvchi molekulalarga endogen PUFAlarni metabolize qiladi: ular AA ni metabolize qiladi epoksiikosatetraenoik kislotalar (ya'ni EET); EPA dan EEQga; va DHA dan EDPgacha.
- CYP2S1 PUFA-ni signal beruvchi molekulalarga metabolize qiladi: AA-EETs va EPA-ni EEQ-larga almashtiradi.
- CYP3A4 AA dan EET signalizatsiya molekulalarini metabolizadi.
- CYP4A11 signal beruvchi molekulalarga endogen PUFAlarni metabolize qiladi: u AA ni 20-HETE va EETlarga metabolizadi; u shuningdek DHA ni 22-gidroksi-DHA (ya'ni 12-HDHA) ga gidroksillaydi.
- CYP4F2, CYP4F3A va CYP4F3B (qarang CYP4F3 oxirgi ikki CYP uchun) signalizatsiya molekulalari uchun PUFAlarni metabolizmga aylantiradi: ular AA ni 20-HETE ga almashtiradi. Ular, shuningdek, EPK ni 19-gidroksyeikosapentaenoik kislota (19-HEPE) va 20-gidroksyeikosapentaenoik kislota (20-HEPE) ga aylantiradi, shuningdek DHA ni 22-HDA ga aylantiradi. Ular shuningdek signal beruvchi molekulalarning faolligini kamaytiradi yoki kamaytiradi: ular metabolizmga kirishadi leykotrien B4 (LTB4) dan 20-gidroksi-LTB4 gacha, 5-gidroksyeikosatetraenoik kislota (5-HETE) dan 5,20-diHETE gacha, 5-okso-eikosatetraenoik kislota (5-okso-ETE) dan 5-okso, 20-gidroksi-ETEgacha, 12-gidrokseyikosatetraenoik kislota (12-HETE) dan 12,20-diHETE ga, EET dan 20-gidroksi-EETga va lipoksinlar 20-gidroksi mahsulotiga.
- CYP4F8 va CYP4F12 molekulalarni signalizatsiya qilish uchun PUFA-ni metabolize qiladi: ular EPA-ni EEQ-larga va DHA-ni EDP-larga aylantiradi. Ular, shuningdek, AA ni 18-gidroksieikosatetraenoik kislotaga (18-HETE) va 19-HETE ga aylantiradilar.
- CYP4F11 signal beruvchi molekulalarning faolligini faolsizlantiradi yoki kamaytiradi: u LTB4 dan 20-gidroksi-LTB4 gacha, (5-HETE) 5,20-diHETE gacha, (5-okso-ETE) 5-okso, 20-gidroksi-ETE, ( 12-HETE) - 12,20-diHETE, EETs - 20-gidroksi-EET va lipoksinlar 20-gidroksi mahsulotiga.
- CYP4F22 b-gidroksilatlar juda uzun "juda uzun zanjirli yog 'kislotalari ", ya'ni 28 yoki undan ko'p uglerodli yog 'kislotalari. Ushbu maxsus yog' kislotalarining b-gidroksillanishi terining suv to'siqni funktsiyasini yaratish va saqlash uchun juda muhimdir; CYP4F22 ning autosomal retsessiv inaktivatsion mutatsiyalari Lamellar ichtiyozisi pastki turi Tug'ma ichtioiform formali eritrodema odamlarda.[35]
Odamlarda CYP oilalari
Odamlar 57 genga ega va 59 dan ortiq pseudogenlar sitoxrom P450 genlarining 18 ta oilasi va 43 ta subfamiliya o'rtasida bo'lingan.[36] Bu genlar va ular kodlagan oqsillarning qisqacha mazmuni. Batafsil ma'lumot uchun sitoxrom P450 Nomenklatura qo'mitasining bosh sahifasini ko'ring.[20]
| Oila | Funktsiya | A'zolar | Genlar | pseudogenlar |
| CYP1 | dori va steroid (ayniqsa estrogen metabolizm, benzo [a] piren toksifikatsiya (shakllantirish) (+) - benzo [a] piren-7,8-dihidrodiol-9,10-epoksid ) | 3 subfamila, 3 gen, 1 psevdogen | CYP1A1, CYP1A2, CYP1B1 | CYP1D1P |
| CYP2 | dori va steroid metabolizm | 13 subfamila, 16 gen, 16 pseudogenlar | CYP2A6, CYP2A7, CYP2A13, CYP2B6, CYP2C8, CYP2C9, CYP2C18, CYP2C19, CYP2D6, CYP2E1, CYP2F1, CYP2J2, CYP2R1, CYP2S1, CYP2U1, CYP2W1 | Ro'yxat uchun juda ko'p |
| CYP3 | dori va steroid (shu jumladan testosteron ) metabolizm | 1 subfamila, 4 gen, 4 pseudogenlar | CYP3A4, CYP3A5, CYP3A7, CYP3A43 | CYP3A51P, CYP3A52P, CYP3A54P, CYP3A137P |
| CYP4 | arakidon kislotasi yoki yog 'kislotasi metabolizmi | 6 subfamila, 12 gen, 10 pseudogenlar | CYP4A11, CYP4A22, CYP4B1, CYP4F2, CYP4F3, CYP4F8, CYP4F11, CYP4F12, CYP4F22, CYP4V2, CYP4X1, CYP4Z1 | Ro'yxat uchun juda ko'p |
| CYP5 | tromboksan A2 sintaz | 1 subfamila, 1 gen | CYP5A1 | |
| CYP7 | safro kislotasi steroid yadrosining 7-alfa gidroksilaza biosintezi | 2 subfamila, 2 gen | CYP7A1, CYP7B1 | |
| CYP8 | turli xil | 2 subfamila, 2 gen | CYP8A1 (prostatsiklin sintaz), CYP8B1 (safro kislotasi biosintezi) | |
| CYP11 | steroid biosintez | 2 subfamila, 3 gen | CYP11A1, CYP11B1, CYP11B2 | |
| CYP17 | steroid biosintez, 17-alfa gidroksilaza | 1 subfamila, 1 gen | CYP17A1 | |
| CYP19 | steroid biosintez: aromataza sintez qiladi estrogen | 1 subfamila, 1 gen | CYP19A1 | |
| CYP20 | noma'lum funktsiya | 1 subfamila, 1 gen | CYP20A1 | |
| CYP21 | steroid biosintez | 1 subfamila, 1 gen, 1 psevdogen | CYP21A2 | CYP21A1P |
| CYP24 | D vitamini tanazzul | 1 subfamila, 1 gen | CYP24A1 | |
| CYP26 | retinoik kislota gidroksilaza | 3 subfamila, 3 gen | CYP26A1, CYP26B1, CYP26C1 | |
| CYP27 | turli xil | 3 subfamila, 3 gen | CYP27A1 (safro kislotasi biosintez), CYP27B1 (D vitamini3 1-alfa gidroksilaza, D vitaminini faollashtiradi3), CYP27C1 (noma'lum funktsiya) | |
| CYP39 | 24-gidroksixolesterinning 7-alfa gidroksillanishi | 1 subfamila, 1 gen | CYP39A1 | |
| CYP46 | xolesterin 24-gidroksilaza | 1 subfamila, 1 gen, 1 psevdogen | CYP46A1 | CYP46A4P |
| CYP51 | xolesterin biosintez | 1 subfamila, 1 gen, 3 pseudogenes | CYP51A1 (lanosterol 14-alfa demetilaza) | CYP51P1, CYP51P2, CYP51P3 |
Boshqa turlardagi P450s
Hayvonlar
Ko'pgina hayvonlarda CYP genlari odamlarga qaraganda ko'proq yoki ko'pdir. Xabar qilingan raqamlar shimgichdagi 35 gendan iborat Amfimedon queenslandica sefaloxordatdagi 235 gengacha Branchiostoma floridae.[37] Sichqonlar 101 CYP uchun genlarga ega va dengiz kirpi undan ham ko'proq (ehtimol 120 ta gen) bo'lishi mumkin.[38]Ko'pgina CYP fermentlari monooksigenaza faolligiga ega deb taxmin qilinadi, chunki tekshirilgan ko'plab sutemizuvchilarning CYP-larida bo'lgani kabi (masalan, CYP19 va CYP5 ). Gen va genomlar ketma-ketligi juda orqada biokimyoviy fermentlarning funktsiyasini tavsiflash, ammo ko'plab genlar yaqin homologiya funktsiyasi ma'lum bo'lgan CYP-lar topilgan bo'lib, ularning funktsional imkoniyatlari haqida ma'lumot beradi.
Odam bo'lmagan hayvonlarda ko'pincha o'rganiladigan CYP sinflari, ular ishtirok etadiganlardir rivojlanish (masalan, retinoik kislota yoki gormon metabolizm) yoki toksik birikmalar metabolizmida ishtirok etadi (masalan heterosiklik aminlar yoki poliaromatik uglevodorodlar ). Ko'pincha farqlar mavjud genlarni tartibga solish yoki ferment funktsiyasi zaharli birikmalarga nisbatan sezgirlikdagi kuzatilgan farqlarni tushuntiradigan (masalan, kofein kabi ksantinlarni metabolizmga qodir emasligi) qarindosh hayvonlarning CYP-laridan iborat. Ba'zi dorilar har xil turdagi fermentlar orqali metabolizmga uchraydi, natijada turli metabolitlar paydo bo'ladi, boshqa dorilar esa bir turda metabolizmga uchraydi, ammo boshqa turda o'zgarmagan holda chiqariladi. Shu sababli, bir turdagi moddaga reaktsiyasi, bu moddalarning odamlarga ta'sirini ishonchli ko'rsatma emas. Ning sozlangan ifodasini ishlatadigan Sonoran Desert Drosophila turi CYP28A1 kaktuslar rotini zararsizlantirish uchun gen Drosophila mettleri. Ushbu turdagi chivinlar mezbon o'simliklarda yuqori darajadagi alkaloidlar ta'siri tufayli ushbu genning regulyatsiyasini moslashtirgan.
CYPlar keng ko'lamli tekshiruvdan o'tkazildi sichqonlar, kalamushlar, itlar va kamroq zebrafish, ulardan foydalanishni osonlashtirish maqsadida model organizmlar yilda giyohvand moddalarni kashf qilish va toksikologiya. Yaqinda parranda turlarida, xususan kurka kurkalarida CYPlar topildi, ular odamlarda saraton kasalligini o'rganish uchun foydali model bo'lishi mumkin.[39] CYP1A5 va CYP3A37 kurkalarda odamga juda o'xshash ekanligi aniqlandi CYP1A2 va CYP3A4 navbati bilan, ularning kinetik xususiyatlari, shuningdek aflatoksin B1 metabolizmida.[40]
CYP'lar ham juda ko'p o'rganilgan hasharotlar, ko'pincha tushunish uchun pestitsidga qarshilik. Masalan, CYP6G1 insektitsidga chidamliligi bilan bog'liq DDT - chidamli Drosophila melanogaster[41] va CYP6M2 chivin ichida bezgak vektor Anopheles gambiae to'g'ridan-to'g'ri metabolizmga qodir piretroidlar.[42]
Mikrobial
Mikrobiyal sitoxromlar P450 ko'pincha eriydigan fermentlar bo'lib, turli xil metabolik jarayonlarda ishtirok etadi. Bakteriyalarda P450 ning tarqalishi juda o'zgaruvchan, chunki P450s aniqlanmagan ko'plab bakteriyalar (masalan, E.coli). Ba'zi bakteriyalar, asosan aktinomitsetlar, ko'plab P450larga ega (masalan,[43][44]). Hozirgacha aniqlanganlar odatda ksenobiotik birikmalarning biotransformatsiyasida ishtirok etadi (masalan, CYP105A1 dan Streptomyces griseolus sulfanilüre gerbitsidlarini kamroq toksik hosilalarga metabolize qiladi,[45]) yoki maxsus metabolitli biosintez yo'llarining bir qismidir (masalan. CYP170B1 ichida sesquiterpenoid albaflavenone ishlab chiqarishni kataliz qiladi Streptomyces albus[46]). Hali ham P450 mikrobda muhim ekanligi isbotlanmagan bo'lsa-da, CYP105 oilasi har birida vakili bilan yuqori darajada saqlanib qolgan streptomitset genom hozirgacha ketma-ketlikda.[47] Bakterial P450 fermentlarining eruvchanligi tufayli ular asosan membrana bilan bog'langan ökaryotik P450larga qaraganda osonroq ishlaydi. Bu ular katalizlaydigan ajoyib kimyo bilan birlashganda, in vitro heterologik ta'sir ko'rsatadigan oqsillardan foydalangan holda ko'plab tadqiqotlarga olib keldi. P450-larning in vivo jonli ravishda nima qilishini, tabiiy substrat (lar) ni va P450-larning tabiiy muhitdagi bakteriyalarning saqlanib qolishida qanday hissa qo'shishini o'rgangan bir nechta tadqiqotlar mavjud. Strukturaviy va mexanik tadqiqotlar uchun katta hissa qo'shgan uchta misol bu erda keltirilgan, ammo juda ko'p turli xil oilalar mavjud.
- Sitoxrom P450 kamerasi (CYP101A1) dastlab Pseudomonas putida ko'plab P450 sitoxromlari uchun namuna sifatida ishlatilgan va rentgen-kristallografiya bilan hal qilingan birinchi o'lchovli oqsil P450 sitokromi bo'lgan. Ushbu ferment kofur-gidroksilatlovchi katalitik tsiklning bir qismidir, bu ikki elektron o'tkazish bosqichidan iborat putidaredoksin, tarkibida 2Fe-2S klasteri bo'lgan protein kofaktori.
- Sitoxrom P450 eryF (CYP107A1) dastlab aktinomitset bakteriyasidan Sachcharopolyspora eritreya ning biosintezi uchun javobgardir antibiotik eritromitsin makrolid 6-deoksieritronolid B ning C6-gidroksillanishi bilan.
- Sitoxrom P450 BM3 (CYP102A1) tuproq bakteriyasidan Bacillus megaterium bir nechtasining NADPHga bog'liq gidroksillanishini katalizlaydi uzoq zanjirli yog 'kislotalari ω – 1 dan ω – 3 gacha bo'lgan holatlarda. Deyarli har qanday ma'lum bo'lgan CYPdan farqli o'laroq (CYP505A1, sitokrom P450 tulkisi bundan mustasno), u CYP domeni va elektron xayr-ehson qiluvchi kofaktor o'rtasidagi tabiiy termoyadroviy oqsilni tashkil qiladi. Shunday qilib, BM3 biotexnologik dasturlarda juda foydali.[48][49]
- Sitoxrom P450 119 (CYP119A1 ) dan ajratilgan termofil arxey Sulfolobus solfatarikus [50] turli mexanistik tadqiqotlarda qo'llanilgan.[17] Termofil fermentlar evolyutsiyasi tufayli yuqori haroratda ishlaydi, ular xona haroratida sekinroq ishlashga moyil (umuman bo'lsa) va shuning uchun ajoyib mexanistik modellardir.
Qo'ziqorinlar
Odatda ishlatiladi azol qo'ziqorinlarga qarshi vositalar qo'ziqorinni inhibe qilish bilan ishlaydi sitoxrom P450 14a-demetilaza. Bu konvertatsiya qilishni to'xtatadi lanosterol ga ergosterol, qo'ziqorin hujayralari membranasining tarkibiy qismi. (Bu faqat odamlarning P450-si boshqa sezgirlikka ega bo'lgani uchun foydalidir; bu sinf shunday qo'ziqorinlarga qarshi vositalar ish.)[51]
P450 qo'ziqorinlari bo'yicha bir qator tadqiqotlar olib borilmoqda, chunki bir qator qo'ziqorinlar mavjud patogen odamlarga (masalan Candida xamirturush va Aspergillus ) va o'simliklarga.
Kanninghamella eleganlari sutemizuvchilar dori almashinuvi uchun namuna sifatida foydalanish uchun nomzoddir.
O'simliklar
O'simliklar sitoxromi P450lar ko'plab biosintez reaktsiyalarida qatnashadilar va turli xil biomolekulalarni maqsad qilib oladilar. Ushbu reaktsiyalar turli xillikka olib keladi yog 'kislotasi konjugatlar, o'simlik gormonlari, ikkilamchi metabolitlar, ligninlar va turli xil himoya aralashmalari.[52] O'simliklar genomining izohlari shuni ko'rsatadiki, sitoxrom P450 genlari o'simlik genlarining 1% ni tashkil qiladi. P450 genlarining soni va xilma-xilligi, qisman, bioaktiv birikmalarning ko'pligi uchun javobgardir.[53]
Sitoxrom P450 aromatik O-demetilaza Ikki xil ravshan qismdan iborat: sitoxrom P450 oqsili (GcoA) va uchta domen reduktaza, o'simlik hujayralari devorlarida keng tarqalgan aromatik biopolimer Ligninni katabolik reaktsiyalar to'plamida qayta tiklanadigan uglerod zanjiriga aylantirish qobiliyati bilan muhimdir. . Muxtasar qilib aytganda, bu Lignin konversiyasidagi muhim bosqichni osonlashtiruvchidir.
Biotexnologiyada P450s
P450-larning ajoyib reaktivligi va substratning buzilishi uzoq vaqt kimyogarlarning e'tiborini tortgan.[54] So'nggi paytlarda P450-lardan qiyin oksidlanishlar tomon foydalanish potentsialini ro'yobga chiqarishga quyidagilar kiradi: (i) molekulalarni o'z ichiga olgan arzon peroksid bilan almashtirish orqali tabiiy ko-faktorlarga bo'lgan ehtiyojni bartaraf etish;[55] (ii) P450s ning organik erituvchilar bilan mosligini o'rganish,[56] va (iii) P450 oksidlanishini bashoratli ravishda yo'naltirish uchun kichik, chiral bo'lmagan yordamchilardan foydalanish.[iqtibos kerak ]
InterPro subfamilies
Ushbu bo'lim mumkin talab qilish tozalamoq Vikipediya bilan tanishish uchun sifat standartlari. Muayyan muammo: buzilgan havolalar; parchalangan paragraf. (2016 yil sentyabr) (Ushbu shablon xabarini qanday va qachon olib tashlashni bilib oling) |
InterPro subfamilies:
- Sitokrom P450, B-sinf InterPro: IPR002397
- Sitoxrom P450, mitoxondriyal InterPro: IPR002399
- Sitoxrom P450, E-sinf, I guruh InterPro: IPR002401
- Sitoxrom P450, E-sinf, II guruh InterPro: IPR002402
- Sitoxrom P450, E-sinf, IV guruh InterPro: IPR002403
- Aromataza
Klozapin, imipramin, paratsetamol, fenatsetin Geterosiklik aril aminlar Induktiv va CYP1A2 5-10% uroporfirinogenni uroporfiringa (CYP1A2) gem metabolizmasida etishmovchiligiga olib keladi, ammo ular qo'shimcha kashf qilinmagan endogen substratlarga ega bo'lishi mumkin. ba'zi politsiklik uglevodorodlar tomonidan induktsiya qilinadi, ularning ba'zilari sigareta tutunida va charchagan ovqatda uchraydi.
Ushbu fermentlar qiziqish uyg'otadi, chunki tahlillarda ular kanserogenlar tarkibidagi birikmalarni faollashtirishi mumkin. CYP1A2 ning yuqori darajasi yo'g'on ichak saratoni xavfini oshirishi bilan bog'liq. 1A2 fermenti sigaretani chekish orqali paydo bo'lishi mumkinligi sababli, bu chekishni yo'g'on ichak saratoni bilan bog'laydi.[57]
Shuningdek qarang
Adabiyotlar
- ^ Gonsales FJ, Gelboin HV (noyabr 1992). "Inson sitoxromlari P450: evolyutsiyasi va cDNA yo'naltirilgan ifodasi". Atrof muhitni muhofaza qilish istiqbollari. 98: 81–5. doi:10.1289 / ehp.929881. PMC 1519618. PMID 1486867.
- ^ a b "Sitoxrom P450". InterPro.
- ^ a b Danielson PB (2002 yil dekabr). "Sitoxrom P450 superfamilasi: odamlarda biokimyo, evolyutsiya va dori almashinuvi". Hozirgi dori metabolizmi. 3 (6): 561–97. doi:10.2174/1389200023337054. PMID 12369887.
- ^ Lamb DC, Lei L, Warrilow AG, Lepesheva GI, Mullins JG, Waterman MR, Kelly SL (avgust 2009). "Birinchi virusli kodlangan p450 sitoxromi". Virusologiya jurnali. 83 (16): 8266–9. doi:10.1128 / JVI.00289-09. PMC 2715754. PMID 19515774.
- ^ Sigel R, Sigel A, Sigel H (2007). Sitoxrom P450 oqsillarining keng tarqalgan rollari: hayot fanlari bo'yicha metall ionlar. Nyu-York: Vili. ISBN 978-0-470-01672-5.
- ^ Nelson, Devid R. (yanvar 2018). "Hayot daraxtidagi sitoxrom P450 xilma-xilligi". Biochimica et Biofhysica Acta (BBA) - Oqsillar va Proteomikalar. 1866 (1): 141–154. doi:10.1016 / j.bbapap.2017.05.003. PMC 5681887. PMID 28502748.
- ^ Nelson D (2009). "Sitoxrom P450 bosh sahifasi". Inson genomikasi. Tennessi universiteti. 4 (1): 59–65. doi:10.1186/1479-7364-4-1-59. PMC 3500189. PMID 19951895. Olingan 2014-11-13.
- ^ "NCBI ketma-ketligini ko'rish vositasi". Olingan 2007-11-19.
- ^ Nelson, DR (oktyabr 2009). "Sitoxrom p450 bosh sahifasi". Inson genomikasi. 4 (1): 59–65. doi:10.1186/1479-7364-4-1-59. PMC 3500189. PMID 19951895.
- ^ Nelson, Devid R. (2011 yil yanvar). "P450 sitoxromining evolyutsion yo'llarini izlashda taraqqiyot". Biochimica et Biofhysica Acta (BBA) - Oqsillar va Proteomikalar. 1814 (1): 14–18. doi:10.1016 / j.bbapap.2010.08.008. PMID 20736090.
- ^ Hanukoglu I (1996). "P450 sitoxrom tizimlarining elektron o'tkazuvchan oqsillari" (PDF). Adv. Mol. Hujayra biol. Molekulyar va hujayra biologiyasining yutuqlari. 14: 29–55. doi:10.1016 / S1569-2558 (08) 60339-2. ISBN 9780762301133.
- ^ [1]PROSITE P450 uchun konsensus modeli
- ^ Srinivasan, Bxarat (2020-10-08). "Giyohvand moddalarni erta aniqlashda Mixaelis-Menten va atipik kinetikani aniq davolash". dx.doi.org. Olingan 2020-11-09.
- ^ a b Meunier B, de Visser SP, Shaik S (sentyabr 2004). "Sitoxrom p450 fermentlari tomonidan katalizlangan oksidlanish reaktsiyalarining mexanizmi". Kimyoviy sharhlar. 104 (9): 3947–80. doi:10.1021 / cr020443g. PMID 15352783. S2CID 33927145.
- ^ Poulos TL, Finzel BC, Howard AJ (iyun 1987). "P450cam sitoxromining yuqori aniqlikdagi kristalli tuzilishi". Molekulyar biologiya jurnali. 195 (3): 687–700. doi:10.1016/0022-2836(87)90190-2. PMID 3656428.
- ^ Sligar SG, Cinti DL, Gibson GG, Schenkman JB (oktyabr 1979). "P450 jigar sitoxromining oksidlanish-qaytarilish potentsialini spin holatini boshqarish". Biokimyoviy va biofizik tadqiqotlari. 90 (3): 925–32. doi:10.1016 / 0006-291X (79) 91916-8. PMID 228675.
- ^ a b v Rittle J, Green MT (noyabr 2010). "Sitoxrom P450 birikmasi I: ta'qib qilish, tavsiflash va C-H bog'lanishini faollashtirish kinetikasi". Ilm-fan. 330 (6006): 933–7. Bibcode:2010Sci ... 330..933R. doi:10.1126 / science.1193478. PMID 21071661. S2CID 206528205.
- ^ a b Ortiz de Montellano, Pol R.; Pol R. Ortiz de Montellano (2005). Sitoxrom P450: tuzilishi, mexanizmi va biokimyosi (3-nashr). Nyu-York: Kluwer Academic / Plenum nashriyotlari. ISBN 978-0-306-48324-0.
- ^ Berka K, Hendrychová T, Anzenbacher P, Otyepka M (oktyabr 2011). "Ibuprofenning membrana pozitsiyasi sitokrom P450 2C9 faol uchastkasiga kirish yo'li bilan tavsiya etilgan". Jismoniy kimyo jurnali A. 115 (41): 11248–55. Bibcode:2011JPCA..11511248B. doi:10.1021 / jp204488j. PMC 3257864. PMID 21744854.
- ^ a b "P450 jadvali".
- ^ doctorfungus> Antifungal dorilarning o'zaro ta'siri Arxivlandi 2012-08-01 da Arxiv.bugun Tarkib direktori: Rassel E. Lyuis, Pharm.D. 2010 yil 23-yanvarda olingan
- ^ Guengerich FP (2008 yil yanvar). "Sitoxrom p450 va kimyoviy toksikologiya". Toksikologiyada kimyoviy tadqiqotlar. 21 (1): 70–83. doi:10.1021 / tx700079z. PMID 18052394. S2CID 17548932. (Ushbu kontekstdagi metabolizm - bu dorilarning kimyoviy modifikatsiyasi yoki degradatsiyasi.)
- ^ Zahno, A; Brext, K; Morand, R; Maseneni, S; Törok, M; Lindinger, PW; Krähenbühl, S (2011 yil 1-fevral). "HepG2 hujayralarida amiodaron bilan bog'liq toksiklikda CYP3A4 ning roli". Biokimyoviy farmakol. 81 (3): 432–41. doi:10.1016 / j.bcp.2010.11.002. PMID 21070748.
- ^ "Karbamazepin: giyohvand moddalarning ko'plab o'zaro ta'sirlarini kuzatib boring". Pharmacy Times.
- ^ Beyli DG, Dresser GK (2004). "Greypfrut sharbati va yurak-qon tomir dorilarining o'zaro ta'siri". Amerika yurak-qon tomir dori vositalari jurnali. 4 (5): 281–97. doi:10.2165/00129784-200404050-00002. PMID 15449971. S2CID 11525439.
- ^ Zeratskiy K (2008-11-06). "Greypfrut sharbati: Dori vositalarining o'zaro ta'sirini keltirib chiqarishi mumkinmi?". Oziq-ovqat va ovqatlanish bo'yicha mutaxassisdan so'rang. MayoClinic.com. Olingan 2009-02-09.
- ^ Chaudxari A, Uillett KL (2006 yil yanvar). "Avliyo Ioann wortining flavonoidlari tomonidan inson sitoxromining CYP 1 fermentlarini inhibatsiyasi". Toksikologiya. 217 (2–3): 194–205. doi:10.1016 / j.tox.2005.09.010. PMID 16271822.
- ^ Strandell J, Nil A, Karlin G (2004 yil fevral). "O'simliklardan va boshqa tabiiy preparatlardan sitoxrom P450 fermentini inhibatsiyasi potentsialini in vitro baholashga yondashuv". Fitomeditsina. 11 (2–3): 98–104. doi:10.1078/0944-7113-00379. PMID 15070158.
- ^ Kroon LA (sentyabr 2007). "Chekish bilan giyohvandlikning o'zaro ta'siri". Amerika sog'liqni saqlash tizimi farmatsiyasi jurnali. 64 (18): 1917–21. doi:10.2146 / ajhp060414. PMID 17823102. S2CID 5397510.
- ^ Zhang JW, Liu Y, Cheng J, Li V, Ma H, Liu HT, Sun J, Vang LM, He YQ, Vang Y, Vang ZT, Yang L (2007). "Yulduzli meva sharbatlari bilan inson jigarining sitokromi P450 ning inhibatsiyasi". Farmatsiya va farmatsevtika fanlari jurnali. 10 (4): 496–503. doi:10.18433 / j30593. PMID 18261370.
- ^ Leclercq I, Desager JP, Horsmans Y (1998 yil avgust). "Xlorzoksazon metabolizmini inhibe qilish, CYP2E1 uchun klinik tekshiruv, suv terisini bir marta qabul qilish orqali". Klinik farmakologiya va terapiya. 64 (2): 144–9. doi:10.1016 / S0009-9236 (98) 90147-3. PMID 9728894. S2CID 43863786.
- ^ Uolmsli, Simon. "Tributiltinning global miqyosda ifloslanishi. Tegishli va yaqinda o'tkazilgan tadqiqotlarga umumiy nuqtai: ta'siri va muammolari" (PDF). WWF UK.
- ^ Chatterjee P, Franklin MR (2003 yil noyabr). "Inson sitoxromi p450 ning inhibatsiyasi va oltin zaharli ekstrakti va uning metilenedioksifenil komponentlari orqali metabolik-oraliq kompleks hosil bo'lishi". Giyohvand moddalar almashinuvi va joylashishi. 31 (11): 1391–7. doi:10.1124 / dmd.31.11.1391. PMID 14570772. S2CID 2967171.
- ^ Xaggström, Mikael; Richfield, Devid (2014). "Inson steroidogenezi yo'llarining diagrammasi". Tibbiyot bo'yicha WikiJournal. 1 (1). doi:10.15347 / wjm / 2014.005. ISSN 2002-4436.
- ^ Sugiura K, Akiyama M (iyul 2015). "Avtosomal retsessiv konjenital ichtiyoz: soch namunalari yordamida mRNK tahlili genetik diagnostika uchun kuchli vosita". Dermatologiya fanlari jurnali. 79 (1): 4–9. doi:10.1016 / j.jdermsci.2015.04.009. PMID 25982146.
- ^ Nelson D (2003). Odamlarda P450 sitoxromlari. Qabul qilingan 2005 yil 9-may.
- ^ Nelson DR, Goldstone QK, Stegeman JJ (2013 yil fevral). "Sitoxrom P450 genezisining joylashuvi: hayvonlarning P450 sitoxromining kelib chiqishi va evolyutsiyasi". London Qirollik Jamiyatining falsafiy operatsiyalari. B seriyasi, Biologiya fanlari. 368 (1612): 20120474. doi:10.1098 / rstb.2012.0474. PMC 3538424. PMID 23297357.
- ^ Goldstone QK, Hamdoun A, Cole BJ, Howard-Ashby M, Nebert DW, Scally M, Dean M, Epel D, Hahn ME, Stegeman JJ (dekabr 2006). "Kimyoviy defensom: Strongilotsentrotus purpuratus genomidagi atrof-muhitni sezish va javob berish genlari". Rivojlanish biologiyasi. 300 (1): 366–84. doi:10.1016 / j.ydbio.2006.08.066. PMC 3166225. PMID 17097629.
- ^ Rawal S, Kim JE, Coulombe R (dekabr 2010). "Aflatoksin B1 parranda go'shti: toksikologiya, metabolizm va profilaktika". Veterinariya fanidagi tadqiqotlar. 89 (3): 325–31. doi:10.1016 / j.rvsc.2010.04.011. PMID 20462619.
- ^ Rawal S, Coulombe RA (avgust 2011). "Turkiya jigar mikrosomalarida aflatoksin B1 metabolizmi: P450 1A5 va 3A37 sitoxromlarining nisbiy rollari". Toksikologiya va amaliy farmakologiya. 254 (3): 349–54. doi:10.1016 / j.taap.2011.05.010. PMID 21616088.
- ^ McCart C, Ffrench-Constant RH (iyun 2008). "Hasharotlarga chidamliligi bilan bog'liq sitoxrom P450 genini Cyp6g1ni ajratish". Zararkunandalarni boshqarish bo'yicha fan. 64 (6): 639–45. doi:10.1002 / ps.1567. PMID 18338338.
- ^ Ismoil, hanafiy; O'Nil, Pol; Xong, Devid; Fin, Robert; Xenderson, Kolin; Rayt, Aaron; Kravatt, Benjamin; Xeminguey, Janet; Paine, Mark (2013 yil 3-dekabr). "Hasharotlar bilan o'zaro bog'liqligi bilan bog'liq sitokrom P450 faoliyatini profilaktika qilish uchun piretroid faolligiga asoslangan zondlar". PNAS. 110 (49): 19766–19771. Bibcode:2013PNAS..11019766I. doi:10.1073 / pnas.1320185110. PMC 3856776. PMID 24248381.
- ^ McLean KJ, Clift D, Lyuis DG, Sabri M, Balding PR, Satkliffe MJ, Leys D, Munro AW (may 2006). "Mycobacterium tuberculosis genomidagi P450s ning ustunligi". Mikrobiologiya tendentsiyalari. 14 (5): 220–8. doi:10.1016 / j.tim.2006.03.002. PMID 16581251.
- ^ Ikeda H, Ishikava J, Xanamoto A, Shinose M, Kikuchi H, Shiba T, Sakaki Y, Xattori M, Omura S (may 2003). "Streptomyces avermitilis sanoat mikroorganizmlarining to'liq genom ketma-ketligi va qiyosiy tahlili". Tabiat biotexnologiyasi. 21 (5): 526–31. doi:10.1038 / nbt820. PMID 12692562.
- ^ Leto, O'Kif (1988). "Streptomyces griseolus tarkibidagi konstruktiv va herbitsidni keltirib chiqaradigan P-450 sitoxromlarini aniqlash". Arch Microbiol. 149 (5): 406–12. doi:10.1007 / BF00425579. S2CID 35526991.
- ^ Moody SC, Zhao B, Lei L, Nelson DR, Mullins JG, Waterman MR, Kelly SL, Lamb DC (may 2012). "Albaflavenon biosintezi yo'lining saqlanishini va streptomitsetlarda CYP170 bifunksionalligini o'rganish". FEBS jurnali. 279 (9): 1640–9. doi:10.1111 / j.1742-4658.2011.08447.x. PMID 22151149.
- ^ Moody SC, Loveridge EJ (2014 yil dekabr). "CYP105 turli xil tuzilmalar, funktsiyalar va Streptomitsdagi fermentlarning qiziqarli oilasidagi rollari". Amaliy mikrobiologiya jurnali. 117 (6): 1549–63. doi:10.1111 / jam.12662. PMC 4265290. PMID 25294646.
- ^ Narhi LO, Fulco AJ (iyun 1986). "Bacillus megateriumda barbituratlar tomonidan induktsiya qilingan katalitik jihatdan o'zini o'zi ta'minlaydigan 119000 daltonli sitoxrom P-450 monoxoksigenazaning xarakteristikasi". Biologik kimyo jurnali. 261 (16): 7160–9. PMID 3086309.
- ^ Girvan HM, Waltham TN, Neeli R, Collins HF, McLean KJ, Scrutton NS, Leys D, Munro AW (dekabr 2006). "Flavotsitoxrom P450 BM3 va CYP102 termoyadroviy turlarining kelib chiqishi". Biokimyoviy jamiyat bilan operatsiyalar. 34 (Pt 6): 1173-7. doi:10.1042 / BST0341173. PMID 17073779.
- ^ Rayt RL, Xarris K, Solou B, Oq RH, Kennelli PJ (aprel 1996). "Sulfolobus solfataricus arxeyonidan potentsial P450 sitoxromini klonlash". FEBS xatlari. 384 (3): 235–9. doi:10.1016/0014-5793(96)00322-5. PMID 8617361. S2CID 19579406.
- ^ Vanden Bossche H, Marichal P, Gorrens J, Koen MC (sentyabr 1990). "Og'iz orqali qo'ziqorinlarga qarshi preparatlar faolligi va selektivligining biokimyoviy asoslari". Britaniya Klinik Amaliyot Jurnali. Qo'shimcha. 71: 41–6. PMID 2091733.
- ^ Schuler MA, Werck-Reichhart D (2003-01-01). "P450s funktsional genomikasi". O'simliklar biologiyasining yillik sharhi. 54 (1): 629–67. doi:10.1146 / annurev.arplant.54.031902.134840. PMID 14503006.
- ^ Mizutani M, Sato F (mart 2011). "O'simliklarning ikkilamchi metabolizmasidagi g'ayrioddiy P450 reaktsiyalari". Biokimyo va biofizika arxivlari. P450 kataliz mexanizmlari. 507 (1): 194–203. doi:10.1016 / j.abb.2010.09.026. PMID 20920462.
- ^ Chefson A, Okler K (2006 yil oktyabr). "P450 fermentlaridan osonroq foydalanish yo'lidagi taraqqiyot". Molekulyar biosistemalar. 2 (10): 462–9. doi:10.1039 / b607001a. PMID 17216026.
- ^ Chefson A, Zhao J, Okler K (iyun 2006). "Tabiiy kofaktorlarni tanlangan vodorod peroksid donorlari yoki organik peroksidlar bilan almashtirish CYP3A4 va CYP2D6 faolligini yaxshilaydi". ChemBioChem. 7 (6): 916–9. doi:10.1002 / cbic.200600006. PMID 16671126. S2CID 39329433.
- ^ Chefson A, Okler K (2007 yil iyul). "Organik kosolventlar, ionli suyuqliklar yoki suv bilan aralashmaydigan organik erituvchilar ishtirokidagi CYP3A4 faolligi". ChemBioChem. 8 (10): 1189–97. doi:10.1002 / cbic.200700128. PMID 17526062. S2CID 11845235.
- ^ Petros WP, Younis IR, Ford JN, Weed SA (oktyabr 2012). "Tamaki chekish va nikotinning saraton kasalligini davolashga ta'siri". Farmakoterapiya. 32 (10): 920–31. doi:10.1002 / j.1875-9114.2012.01117. PMC 3499669. PMID 23033231.
Qo'shimcha o'qish
- Gelboin HV, Krausz K (mart 2006). "Monoklonal antikorlar va ko'p funktsiyali sitokrom P450: paradigma sifatida dori almashinuvi". Klinik farmakologiya jurnali. 46 (3): 353–72. doi:10.1177/0091270005285200. PMID 16490812.
- Gelboin HV, Krausz KW, Gonsales FJ, Yang TJ (1999 yil noyabr). "Inson sitoxromi P450 fermentlariga to'sqinlik qiluvchi monoklonal antikorlar: giyohvand moddalarni topish uchun yangi yo'l" (PDF). Farmakologiya fanlari tendentsiyalari. 20 (11): 432–8. doi:10.1016 / S0165-6147 (99) 01382-6. PMID 10542439.
- "Monoklonal antikorlardan foydalangan holda sitoxrom P450 vositachiligidagi dori va kanserogen metabolizmi". home.ccr.cancer.gov. Olingan 2018-04-02.
- Krausz KW, Goldfarb I, Buters JT, Yang TJ, Gonsales FJ, Gelboin HV (noyabr, 2001). "P450 2C8, 2C9 va 2C19 odam sitoxromlariga xos va inhibitori bo'lgan monoklonal antikorlar". Giyohvand moddalar almashinuvi va joylashishi. 29 (11): 1410–23. PMID 11602516.
- Gonsales FJ, Gelboin HV (1994). "Kimyoviy kanserogenlar va toksinlarning metabolik faollashuvida odam sitokromlari P450 ning roli". Giyohvand moddalar almashinuvi bo'yicha sharhlar. 26 (1–2): 165–83. doi:10.3109/03602539409029789. PMID 8082563.
Tashqi havolalar
- Sigaroudi A, Vollbrecht H (2019). "farmakokinetik ta'sir o'tkazish jadvali". Sigaroudi va Vollbrecht.
- Preissner S (2010). "Sitoxrom P450 ma'lumotlar bazasi". Nuklein kislotalarni tadqiq qilish.
- Degtyarenko K (2009-01-09). "P450 o'z ichiga olgan tizimlar ma'lumotnomasi". Xalqaro genetik muhandislik va biotexnologiya markazi. Arxivlandi asl nusxasi 2016-07-16. Olingan 2009-02-10.
- Estabrook RW (2003 yil dekabr). "P450-larga bo'lgan ehtiros (P450 sitoxromi bo'yicha dastlabki tadqiqotlar tarixini esga olish)". Giyohvand moddalar almashinuvi va joylashishi. 31 (12): 1461–73. doi:10.1124 / dmd.31.12.1461. PMID 14625342.
- Flockhart DA (2007). "Sitoxrom P450 dorilar bilan o'zaro ta'sir jadvali". Indiana universiteti-Purdue universiteti Indianapolis. Olingan 2009-02-10.
- Sim SC (2008-09-04). "Inson sitoxromi P450 (CYP) Allele nomenklaturasi qo'mitasi". Karolinska instituti. Olingan 2009-02-10.
- P450 inhibisyonu tadqiqotlarining ishlashi Ning ishlashi in vitro sitokrom P450 inhibisyon tadqiqotlari, shu jumladan ma'lumotlarni tahlil qilish.