DNKning o'zaro bog'liqligi - Crosslinking of DNA

Yilda genetika, DNKning o'zaro bog'liqligi har xil ekzogen yoki endogen agentlar ikkitasi bilan reaksiyaga kirishganda paydo bo'ladi nukleotidlar ning DNK, ular orasidagi kovalent aloqani hosil qiladi. Ushbu o'zaro bog'liqlik bir xil yo'nalishda (intrastrand) yoki ikki qatorli DNKning (interstrand) qarama-qarshi chiziqlari orasida sodir bo'lishi mumkin. Ushbu qo'shimcha moddalar uyali metabolizmga xalaqit beradi, masalan DNKning replikatsiyasi va transkripsiya, qo'zg'atuvchi hujayralar o'limi. Biroq, bu o'zaro bog'liqliklar bo'lishi mumkin ta'mirlangan eksizyon yoki rekombinatsiya yo'llari orqali.
DNKning o'zaro bog'liqligi, shuningdek, kimyoviy terapiya va saraton hujayralarini maqsadli yo'naltirishda foydali xizmatga ega apoptoz,[1] shuningdek, oqsillarning DNK bilan o'zaro ta'sirini tushunishda.
O'zaro bog'lovchi vositalar
Ko'p xarakterli o'zaro bog'liqlik agentlari bir xil molekula ichida ikkita mustaqil reaktiv guruhga ega, ularning har biri DNKning nukleotid qoldig'i bilan bog'lanishga qodir. Ushbu vositalar kelib chiqish manbalariga qarab ajratiladi va ekzogen yoki endogen deb belgilanadi. Ekzogen o'zaro bog'lovchi moddalar - bu kimyoviy moddalar va tabiiy va sintetik birikmalar, bu atrof-muhit ta'siridan kelib chiqadi, masalan, farmatsevtika va sigaretaning tutuni yoki avtomobil chiqindilari. Endogen o'zaro bog'liqlik agentlari - bu hujayra yoki organizm ichidagi uyali yoki biokimyoviy yo'llardan kiritiladigan birikmalar va metabolitlar.
Ekzogen moddalar
- Azot xantallari ekzogen hisoblanadi alkillovchi moddalar bilan reaksiyaga kirishadigan N7 guaninning pozitsiyasi. Ushbu birikmalar o'zgaruvchan, bis- (2-etilxloro) omin yadroli tuzilishga ega R- guruh, ikkita reaktiv funktsional guruh nukleobazlarni alkilatlash va o'zaro bog'liqlik lezyonini hosil qilish uchun xizmat qiladi. Ushbu agentlar eng maqbul ravishda 1,3 5'-d (GNC) o'zaro bog'lanishni hosil qiladi. Ushbu agentning kiritilishi DNK dupleksini spiral tarkibida agentning mavjudligini ta'minlash uchun ozgina egiladi.[2] Ushbu vositalar ko'pincha farmatsevtika sifatida kiritiladi va ishlatiladi sitotoksik kimyoviy terapiya.[3]
- Sisplatin (cis-diamminedichloroplatinum (II)) va uning hosilalari asosan qo'shni guaninlarga ta'sir qiladi. N7 lavozimlar. Planar birikma nukleobazalar bilan uning xlorid guruhlaridan birining yoki ikkalasining siljishi orqali bog'lanib, sisplatinning DNK yoki RNKga monoeduktlar hosil qilishiga imkon beradi, DNKning intrastrand o'zaro bog'lanishlari, interstrand DNKning o'zaro bog'lanishlari va DNK-oqsil o'zaro bog'lanishlari.[4] Sisplatin DNKning o'zaro bog'lanishini hosil qilganda, u tez-tez 1,2-intrastrand o'zaro bog'lanishini (5'-GG) hosil qiladi, ammo past foizlarda 1,3-intrastrand o'zaro bog'lanishini (5-GNG) hosil qiladi.[5][6] Sisplatin interstrand o'zaro bog'lanishlarni (5'-GC) hosil qilganda, qarama-qarshi iplardagi guaninlar orasidagi masofa qisqarishi va GG o'zaro ta'siri natijasida spiraldan chiqib ketgan sitozin tufayli DNK spiralining qattiq buzilishi mavjud.[7] Azot xantaliga o'xshash sisplatin kimyoviy davolashda tez-tez ishlatiladi - ayniqsa moyak va tuxumdon saratonida.[8]
- Xlor etil nitrozo karbamid (CENU), xususan karustin (BCNU), kimyoviy davolashda, ayniqsa miya shishi uchun keng qo'llaniladigan o'zaro bog'lovchi vositalar. Ushbu vositalar alkilatlashi sababli boshqa o'zaro bog'lovchilardan farq qiladi O6 hosil qilish uchun guanin O6-etanoguanin. Ushbu oraliq birikma keyinchalik GC tayanch punkti orasidagi o'zaro bog'liqlikni keltirib chiqaradi. Ushbu o'zaro bog'lovchi vositalar molekulalarning kichikligi tufayli faqat DNK spiralidagi kichik buzilishlarga olib keladi.
- Psoralenlar o'simliklarda mavjud bo'lgan tabiiy birikmalar (furokumarinlar). Ushbu birikmalar interkalate 5'-AT ketma-ketlik joylarida DNKga kiradi va hosil bo'ladi timidin ishtirokida faollashtirilganda qo'shimchalar Ultra Violet-A (UV-A) nurlari.[9] Ushbu kovalent qo'shimchalar 3, 4 (piron ) yoki 4 ', 5' (furan ) psoralenning 5, 6 juft bog'lanishiga chekkasi timin. Psoralenlar ikki turdagi monoo'tkazgichlar va bitta diaduktuk (interstrand o'zaro bog'liqlik) hosil qilishi mumkin timin.[10] Ushbu qo'shimchalar interkalatsiya joyida DNKning mahalliy buzilishlariga olib keladi. Psoralens teri kasalliklarini tibbiy davolashda ishlatiladi, masalan toshbaqa kasalligi va vitiligo.
- Mitomitsin C (MMC) antibiotiklar sinfidir, ular kimyoviy terapiyada keng qo'llaniladi, ko'pincha oshqozon-ichak trakti bilan bog'liq saraton kasalliklarida. Mitomitsin C faqat DNK nukleotidining kamayishiga ega bo'lganda o'zaro bog'liqlik vazifasini bajarishi mumkin kinon uzuk. Ikki dG qayta tashkil etilib, shu tarzda metillanganida, har bir nukleobazaning ekzo aminlari bilan 5'-GC o'zaro bog'liqlik hosil bo'lishi mumkin. Mitomitsin shuningdek DNK bilan monoduktivlar va intrastrand o'zaro bog'lanishlarni hosil qilish qobiliyatiga ega. Mitomitsin C ning o'zaro bog'lanishlari DNKning kichik chuqurchasida hosil bo'lib, ikkita zanjir ichida molekula mavjudligini ta'minlash uchun o'rtacha kengayish yoki DNK spiraliga cho'zilib ketadi.
Endogen moddalar
- Azot kislotasi oshqozonda yon mahsulot sifatida nitritlarning parhez manbalaridan hosil bo'ladi va DNK tarkibidagi amino guruhlarning karbonillarga aylanishi orqali DNKdagi o'zaro bog'liqlik zararlanishiga olib kelishi mumkin. Ushbu turdagi lezyonlar ko'pincha ikki guanozin o'rtasida uchraydi, 4 ta deaminatsiyalangan guanozinlarning 1 tasi interstrand o'zaro bog'liqlikka olib keladi.[11] Bu interstrand DNKning o'zaro bog'lanishini hosil qiladi aminogrup ekzotsiklik N.2 5'-CG sekanslaridagi guanin. Ushbu lezyon er-xotin spiralni yumshoq tarzda buzadi.
- Ikki tomonlama aldegidlar orqali endogen ravishda hosil bo'lgan reaktiv kimyoviy moddalardir lipid peroksidatsiyasi va prostoglandin biosintez.[12] Ular tomonidan tashkil etilgan etheno qo'shimchalarini yaratadilar aldegid DNKning qarama-qarshi zanjirlarida o'zaro bog'lanishni hosil qilish uchun qayta tartibga solinadi. Malondialdegid DNKni ikkita ekzosiltsik guanin amino guruhi orqali o'zaro bog'lashi mumkin bo'lgan prototipik misol.[13] Kabi boshqa aldegidlar formaldegid va atsetilaldegid, interstrand o'zaro bog'lanishlarini kiritishi mumkin va ko'pincha ekzogen moddalar rolini bajaradi, chunki ular ko'plab qayta ishlangan ovqatlarda mavjud. Ko'pincha pestitsidlar, tamaki tutuni va avtomobil chiqindilarida topilgan a, b to'yinmagan aldegidlar, masalan akrolein va krotonaldegid, DNKning o'zaro bog'liqligini keltirib chiqaradigan ekzogen moddalardir. Boshqa o'zaro bog'lovchi vositalardan farqli o'laroq, aldegid bilan bog'liq bo'lgan o'zaro bog'liqlik ichki reversibl jarayondir. Ushbu turdagi agentlarning NMR tuzilishi interstrand o'zaro bog'liqlik sifatida 5'-GC qo'shimchasi DNKning ozgina buzilishiga olib kelishini ko'rsatadi, ammo 5'-CG qo'shimchasi spirali beqarorlashtiradi va DNKdagi egilish va burilishni keltirib chiqaradi.[14]
- DNKning o'zaro bog'lanish lezyonlari oksidlanish stresi sharoitida ham hosil bo'lishi mumkin, bu erda erkin kislorod radikallari DNKda reaktiv qidiruv moddalar hosil qiladi va bu lezyonlar qarish va saraton kasalligiga ta'sir qiladi. Tandem DNK lezyonlari sezilarli chastotada ionlashtiruvchi nurlanish va metall katalizli H tomonidan hosil bo'ladi2O2 reaktsiyalar. Anoksik sharoitda, asosan, ikki asosli lezyon guaninni qo'shni 3'-timinning 5-metil guruhiga (G [8,5- Me] T) bog'lab turadigan intrastrand lezyonlarni hosil qiladigan tur hisoblanadi.[15][16]
O'zaro bog'lovchi vositalarning qisqacha jadvali
| O'zaro bog'lovchi agent | Alkilatuvchi agent | O'zaro bog'lanish tuzilishi | Imtiyozli maqsadli ketma-ketlik |
| Azotli xantal |  |  | Intrastrand; 5'-GC |
| Sisplatin |  |  | Intrastrand; 5'-GC Interstrand; 5'-GNG |
| Karmustin (BNCU) |  | 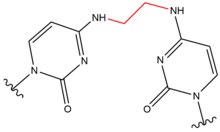 | Tushunmoq; GC tagepair |
| Mitomitsin C |  |  | Interstrand; 5'-GC |
| Psoralen |  |  | Tushunmoq; 5'-TA |
| Malondialdegid |  |  | Interstrand; 5'-GC, 5'-CG |
| Oksidlanish stressi (va nurlanish) |  | Intrastrand; d (GpT) | |
| Azot kislotasi |  | Interstrand; 5'-CG |
DNKning o'zaro bog'lanishini tiklash
O'zaro bog'langan DNK hujayralardagi fermentlar va boshqa omillar birikmasi bilan tiklanadi nukleotid eksizyonini tiklash (NER) yo'l, gomologik rekombinatsiya, va asosiy eksizyonni ta'mirlash (BER) yo'l. Eukaryotlarda interstrand o'zaro bog'lanishlarini tiklash uchun NER dan 3 'qopqoqli endonukleaza, XPF-ERCC1, o'zaro bog'langan DNKga yollangan bo'lib, u o'zaro bog'liqlik uchastkasida 3 'zanjirni ajratib DNKni "ochish" ga yordam beradi. Keyin 5 'torli, yoki tomonidan kesiladi XPF-ERCC1 yoki boshqasi endonukleaza, shakllantirish a ikki qatorli tanaffus (DSB), keyinchalik tomonidan tuzatilishi mumkin gomologik rekombinatsiya yo'l.[17]
DNK o'zaro bog'lanishlari odatda DNKning ikkita zanjiridan ketma-ketlik haqidagi ma'lumotlarning yo'qolishiga olib keladi. Shuning uchun, zararni aniq tiklash, yo'qolgan ma'lumotni buzilmasdan olishiga bog'liq gomologik xromosoma xuddi shu kamerada. Qabul qilish a bilan juftlashish orqali sodir bo'lishi mumkin opa-singil xromosoma replikatsiya oldingi turida ishlab chiqarilgan. A diploid hujayra qidirish opa-singil bo'lmagan ayol bilan juftlashish orqali ham sodir bo'lishi mumkin gomologik xromosoma kabi, ayniqsa paytida sodir bo'ladi mayoz.[iqtibos kerak ] Juftlik sodir bo'lgandan so'ng, o'zaro bog'liqlikni olib tashlash va gomologik rekombinatsiya bilan zararlangan xromosomaga to'g'ri ma'lumotlarni kiritish mumkin.
DNKning shakar-fosfat umurtqasidagi deoksiriboz shakar bilan bog'langan nukleobaza orasidagi bog'lanishning parchalanishi er-xotin zanjirli DNKda tubsiz joy qoldiradi. Ushbu tubsiz joylar ko'pincha oraliq shaklda hosil bo'ladi va keyinchalik eksizyonni tiklashda tiklanadi. Ammo, agar ushbu saytlarning davom etishiga ruxsat berilsa, ular DNKning replikatsiyasi va transkripsiyasini inhibe qilishi mumkin.[18] Abasik joylar oqsillar tarkibidagi amin guruhlari bilan reaksiyaga kirishib, DNK-oqsil o'zaro bog'lanishlarini yoki boshqa nukleobazalarning ekzosiklik aminlari bilan o'zaro bog'lanishni hosil qilishi mumkin. Interstrand yoki DNK-oqsil o'zaro bog'lanishining oldini olish uchun BER yo'lidagi fermentlar abasik joyni mahkam bog'lab, uni yaqin atrofdagi reaktiv guruhlardan ajratib olishadi, chunki bu odam alkiladenin DNK glikozilazasida (AAG) va E. coli 3-metiladenin DNK glikozilaza II (AlkA).[19] in vitro dalillar shuni ko'rsatdiki, abasik sayt (DOB-ICL) tomonidan qo'zg'atilgan o'zaro bog'lanishlar replikatsiyani blokirovka qiluvchi va juda noto'g'ri kodlangan lezyon hisoblanadi. Ko'rib chiqilgan bir nechta boshqa TLS qutilari bilan taqqoslaganda pol η DOB-ICL ning TLS vositasida ta'mirlanishiga hissa qo'shishi mumkin. jonli ravishda.[20] O dan foydalanib6-2'-deoksiguanozin-butilen-O6-2'-deoksiguanozin (O6-dG-C4-O6-dG) DNKning zararlanishi, bu kimyoviy jihatdan barqaror tuzilma, bir nechta DNK polimerazalarini chetlab o'tish faolligi tekshirildi va natijada pol η eng yuqori bypass faolligini namoyish etdi; shu bilan birga, bypass mahsulotlarining 70% mutagen tarkibida almashtirish yoki o'chirishni o'z ichiga olgan. Qopqoqsiz qidiruv vositalar hajmining ko'payishi o'chirish mutatsiyasining chastotasini oshiradi. [21]
Davolash E. coli bilan psoralen -plus-ultrabinafsha nur (PUVA ) hujayralar DNKida interstrand o'zaro bog'lanishlarni hosil qiladi. Koul va boshq.[22] va Sinden va Koul[23] dalillarni taqdim etdi a gomologik rekombinatsion genlarning mahsulotlarini talab qiladigan ta'mirlash jarayoni uvrA, uvrB va recA ushbu o'zaro bog'lanishlarni olib tashlashi mumkin E. coli. Ushbu jarayon juda samarali ko'rinadi. Hujayrani inaktiv qilish uchun bitta yoki ikkita ta'mirlanmagan o'zaro bog'liqlik etarli bo'lsa ham, yovvoyi tipdagi bakterial hujayra 53 dan 71 gacha psoralen o'zaro bog'lanishlarini tiklay oladi va tiklaydi. Eukaryotik xamirturush xujayralari qolgan bitta o'zaro bog'liqlik bilan inaktiv qilinadi, ammo yovvoyi turdagi xamirturush xujayralari 120 dan 200 gacha o'zaro bog'lanishda tiklanishi mumkin.[24]
Ilovalar
DNK va oqsilning o'zaro bog'lanishi
Biokimyoviy ta'sir o'tkazish usullari
DNK-oqsilning o'zaro bog'lanishiga turli xil kimyoviy va fizik moddalar, jumladan, o'tish metallari, ionlashtiruvchi nurlanish va endogen aldegidlar sabab bo'lishi mumkin. kimyoviy terapevtik vositalar.[25]DNKning o'zaro bog'lanishiga o'xshab, DNK-oqsil o'zaro bog'liqligi hujayralardagi ultrabinafsha nurlanishidan tez-tez zarar ko'rgan jarohatlardir. UB ta'siri reaktiv o'zaro ta'sirga olib kelishi va DNK va u bilan aloqada bo'lgan oqsillarni o'zaro bog'lanishiga olib kelishi mumkin. Ushbu o'zaro bog'liqliklar juda katta va murakkab jarohatlardir. Ular birinchi navbatda xromosomalarning DNK replikatsiyasiga uchragan va uyali jarayonlarga xalaqit beradigan joylarida uchraydi.
Strukturani identifikatsiyalash usullarining rivojlanishi rivojlanib bormoqda va DNK va oqsil o'rtasidagi o'zaro ta'sirlarni o'lchash qobiliyatiga qo'shilish biokimyoviy jarayonlarni to'liq anglash uchun talabdir. DNK-oqsil komplekslarining tuzilishini xaritada ko'rish mumkin fotokrossinging Bu ikki makromolekula o'rtasida yoki bitta makromolekulaning ikki xil qismi o'rtasida kovalent bog'lanish hosil bo'lishining fototizimidir. Uslubiyat, maqsadli ketma-ketlikka xos bo'lgan DNKni bog'laydigan oqsilning DNK bilan bog'laydigan motifini ultrabinafsha nurlari ta'sirida DNK nukleotidlari bilan reaksiyaga kirishishga qodir bo'lgan fotoaktivlanuvchi o'zaro bog'liqlik agenti bilan kovalent ravishda bog'lashni o'z ichiga oladi. Ushbu usul o'zaro bog'liqlikdagi DNK va oqsil o'rtasidagi o'zaro bog'liqlik haqida ma'lumot beradi.[26]
Klinik muolajalar
DNKni tiklash yo'llari hosil bo'lishiga olib kelishi mumkin o'simta hujayralari. Saraton davolash DNKning replikatsiyasini blokirovka qilish uchun DNKning azotli asoslari bilan ta'sir o'tkazish uchun DNKni o'zaro bog'laydigan vositalar yordamida ishlab chiqilgan. Ushbu o'zaro bog'liqlik agentlari saraton hujayralarida o'ziga xos nukleotidlarni nishonga olish va yo'q qilish orqali bitta agentli terapiya sifatida harakat qilish qobiliyatiga ega. Ushbu natija saraton hujayralarining aylanishini va o'sishini to'xtatadi; u o'ziga xos DNKni tiklash yo'llarini inhibe qilganligi sababli, ushbu yondashuv kamroq yon ta'sirga ega bo'lishi mumkin bo'lgan afzalliklarga ega.[27]
Odamlarda saraton o'limining asosiy sababi o'pka saratoni, shu jumladan kichik hujayrali bo'lmagan o'pka karsinomasi (NSCLC) bu Qo'shma Shtatlardagi barcha o'pka saratoni holatlarining 85 foizini tashkil qiladi.[28] NSCLC bilan kasallangan shaxslar ko'pincha terapevtik platinaviy birikmalar bilan davolanadi (masalan, sisplatin, karboplatin yoki oksaliplatin) (qarang. O'pka saratoniga qarshi kimyoviy terapiya ) interstrand DNKning o'zaro bog'liqligini keltirib chiqaradi. NSLK bilan kasallanganlar orasida, ko'krak bezi saratoni 1 genining past ifodasi (BRCA1 ) birlamchi o'smada platinani o'z ichiga olgan kimyoviy terapiyadan so'ng omon qolish darajasi yaxshilandi.[29][30] Ushbu o'zaro bog'liqlik shuni anglatadiki, saraton kasalligidagi BRCA1 miqdori pastligi va natijada DNKning tiklanish darajasi pastligi saraton kasalligini DNKning o'zaro bog'lanish agentlari tomonidan davolanishiga olib keladi. Yuqori BRCA1 saraton hujayralarini himoya qilishi mumkin gomologik rekombinatsion platina preparatlari tomonidan kiritilgan DNKdagi zararni bartaraf etadigan yo'lni tiklash. Darajasi BRCA1 ekspression potentsial o'pka saratonini davolashda kimyoviy terapiyani moslashtirish uchun muhim vosita hisoblanadi.[29][30]
Klinik ximoterapiya vositasi fermentativ va fermentativ bo'lmagan DNK-protein bilan o'zaro bog'liqlikni keltirib chiqarishi mumkin. Ushbu induksiyaning misoli platina hosilalari, masalan, sisplatin va oksaliplatin. Ular xromatin bilan o'zaro ta'sir qiluvchi oqsillarni DNK bilan o'ziga xos bo'lmagan o'zaro bog'lanish orqali fermentativ bo'lmagan DNK-oqsil o'zaro bog'lanishlarini hosil qiladi. O'zaro bog'lanish boshqa terapevtik vositalarda kovalent DNK-oqsil reaktsiyasi vositalarini barqarorlashtirish yoki DNKdagi fermentni ushlab turuvchi psevdosubstrat yaratish orqali ham mumkin. Kemptotsin hosilalari, masalan irinotekan va topotekan, o'ziga xos DNKni maqsad qilib oladi va ushlaydi. topoizomeraza 1 (TOP1) ferment-DNK interfeysi ichida interkalatatsiya qilish yo'li bilan. Ushbu dorilarning toksikligi TOP1 tuzoqqa bog'liq bo'lganligi sababli, ushbu birikmalarga uyali sezgirlik to'g'ridan-to'g'ri TOP1 ekspression darajalariga bog'liq. Natijada, ushbu dorilarning vazifasi inhibitorlar emas, balki ferment zahari bo'lib xizmat qilishdir. Buni TOP 2 ferment zaharlari yordamida o'sma hujayralarini davolash uchun qo'llash mumkin.[31]
Adabiyotlar
- ^ Dekanlar, AJ; G'arbiy, SC (24 iyun 2011). "DNK oralig'idagi o'zaro bog'liqlikni tiklash va saraton". Tabiat sharhlari. Saraton. 11 (7): 467–80. doi:10.1038 / nrc3088. PMC 3560328. PMID 21701511.
- ^ Guainaczi, Angelo; Schärer, Orlando D. (2010-11-01). "Ta'mirlash yo'llarini aniqlash va saraton ximiyaterapiyasining yangi terapevtik maqsadlarini aniqlash uchun sintetik DNK interstrand o'zaro bog'lanishlaridan foydalanish". Uyali va molekulyar hayot haqidagi fanlar. 67 (21): 3683–3697. doi:10.1007 / s00018-010-0492-6. ISSN 1420-682X. PMC 3732395. PMID 20730555.
- ^ Saraton kasalligi, Klivlend klinikasi. "Azotli xantal - kimyoviy terapiya preparatlari - ximokare". chemocare.com. Olingan 2017-10-09.
- ^ Jeymison, E. R .; Lippard, S. J. (1999-09-08). "Sisplatin-DNK qo'shimchalarining tuzilishi, tan olinishi va qayta ishlanishi". Kimyoviy sharhlar. 99 (9): 2467–2498. doi:10.1021 / cr980421n. ISSN 1520-6890. PMID 11749487.
- ^ Poklar N, Pilch DS, Lippard SJ, Redding EA, Dunham SU, Breslauer KJ (iyul 1996). "Sisplatin intrastrand o'zaro bog'lanishining 20 merlikli DNK dupleksining konformatsiyasi, issiqlik barqarorligi va energetikasiga ta'siri". Proc. Natl. Akad. Ilmiy ish. AQSH. 93 (15): 7606–11. doi:10.1073 / pnas.93.15.7606. PMC 38793. PMID 8755522.
- ^ Rudd GN, Xartli JA, Souhami RL (1995). "Keksa va yosh odamlarning periferik qonidagi mononukleer hujayralardagi sisplatin ta'sirida DNKning o'zaro bog'lanishining barqarorligi". Saraton kasalligi. Farmakol. 35 (4): 323–6. doi:10.1007 / BF00689452. PMID 7828275.
- ^ Kost, F .; Malinge, J. M .; Serre, L .; Shepard, V.; Rot, M.; Len, M .; Zelwer, C. (1999-04-15). "1.63 da sisplatin interstrand o'zaro bog'liqligini o'z ichiga olgan ikki zanjirli DNKning kristalli tuzilishi. Ruxsat berish: platinlangan joyda hidratsiya". Nuklein kislotalarni tadqiq qilish. 27 (8): 1837–1846. doi:10.1093 / nar / 27.8.1837. ISSN 0305-1048. PMC 148391. PMID 10101191.
- ^ "Sisplatin". Milliy saraton instituti. 2007-03-02. Olingan 2017-10-09.
- ^ Cimino, G. D .; Gamper, H. B .; Ayzaks, S. T .; Xerst, J. E. (1985). "Psoralenlar nuklein kislota tuzilishi va funktsiyasining fotoaktiv zondlari sifatida: organik kimyo, fotokimyo va biokimyo". Biokimyo fanining yillik sharhi. 54: 1151–1193. doi:10.1146 / annurev.bi.54.070185.005443. ISSN 0066-4154. PMID 2411210.
- ^ Qi Vu, Laura A Kristensen, Rendi J Legerski va Karen M Vaskes, Mismatchni tuzatish inson hujayralarida DNKning interstrand o'zaro bog'liqligini xatosiz qayta ishlashda ishtirok etadi, EMBO 6, 6, 551-557 hisobotlari (2005).
- ^ Kirchner, Jeyms J .; Sigurdsson, Snorri T.; Xopkins, Pol B. (1992-05-01). "Dupleks DNKning azot kislotasi bilan o'zaro bog'liqligi: 5'-CG ketma-ketlikda dG-dG o'zaro bog'liqligining kovalent tuzilishi". Amerika Kimyo Jamiyati jurnali. 114 (11): 4021–4027. doi:10.1021 / ja00037a001. ISSN 0002-7863.
- ^ Stoun, Maykl P.; Cho, Young-Jin; Xuang, Xay; Kim, Xay-Yang; Kozekov, Ivan D.; Kozekova, Albena; Vang, Xao; Minko, Irina G.; Lloyd, R. Stiven (2008-07-01). "Lipid peroksidatsiyasi va atrof-muhit manbalaridan olingan a, b-to'yinmagan aldegidlar tomonidan yaratilgan interstrand DNKning o'zaro bog'liqligi". Kimyoviy tadqiqotlar hisoblari. 41 (7): 793–804. doi:10.1021 / ar700246x. ISSN 0001-4842. PMC 2785109. PMID 18500830.
- ^ Niedernhofer, Laura J.; Daniels, J. Scott; Ruzer, Kerol A.; Grin, Reychel E.; Marnett, Lourens J. (2003-08-15). "Malondialdegid, lipid peroksidatsiyasi mahsuloti, inson hujayralarida mutagendir". Biologik kimyo jurnali. 278 (33): 31426–31433. doi:10.1074 / jbc.m212549200. ISSN 0021-9258. PMID 12775726.
- ^ Duli, Patrisiya A.; Chjan, Minchjou; Korbel, Gregori A.; Nechev, Lyubomir V.; Xarris, Konstans M.; Stoun, Maykl P.; Xarris, Tomas M. (2003-01-08). "5'-d (GpC) motifini o'z ichiga olgan oligodeoksinukleotid dupleksidagi trimetilen oralig'idagi o'zaro bog'liqlikning NMR konformatsiyasini aniqlash". Amerika Kimyo Jamiyati jurnali. 125 (1): 62–72. doi:10.1021 / ja0207798. ISSN 0002-7863. PMID 12515507.
- ^ LC Colis; P Raychaudxuri; AK Basu (2008). "Sutemizuvchi hujayralardagi gamma-radiatsiyadan kelib chiqadigan guanin-timin va timin-guanin intrastrand o'zaro bog'lanishlari va insonning DNK polimeraza eta tomonidan guanin-timin shikastlanishidan o'tgan translesion sintezining mutatsion o'ziga xosligi". Biokimyo. 47 (6): 8070–8079. doi:10.1021 / bi800529f. PMC 2646719. PMID 18616294.
- ^ Box, Garold S.; Budzinski, Edvin E.; Davidzik, Jan D.; Uolles, Jon S.; Evans, Marianne S.; Gobey, Jeyson S. (1996). "D (KpGpTpA) ning oksigenlanmagan eritmalarida Deoksiguanozin va Timidin asosiy qismlari o'rtasida o'zaro bog'liqlikning nurlanish natijasida hosil bo'lishi". Radiatsion tadqiqotlar. 145 (5): 641–643. doi:10.2307/3579285. JSTOR 3579285.
- ^ Klayn Douvel, Daisy; Boonen, Rik AC; Uzoq, Devid T .; Sypowska, Anna A.; Rasl, Markus; Valter, Yoxannes S.; Knipscheer, Puck (2014). "XPF-ERCC1 FANCD2 va FANCP / SLX4 bilan hamkorlikda DNKning interstrand o'zaro bog'lanishida ishlaydi". Molekulyar hujayra. 54 (3): 460–471. doi:10.1016 / j.molcel.2014.03.015. PMC 5067070. PMID 24726325.
- ^ DNKning tiklanishi va mutagenezi. Fridberg, Errol C., Fridberg, Errol C. (2-nashr). Vashington, Kolumbiya: ASM Press. 2006 yil. ISBN 9781555813192. OCLC 59360087.CS1 maint: boshqalar (havola)
- ^ Admiraal, Suzanna J.; O'Brayen, Patrik J. (2015-03-10). "Baza eksizyonni tiklash fermentlari dupleks DNKdagi tub joylarni interstrand o'zaro bog'liqliklaridan himoya qiladi". Biokimyo. 54 (9): 1849–1857. doi:10.1021 / bi501491z. ISSN 0006-2960. PMC 4404639. PMID 25679877.
- ^ Chjao, Linlin; Syu, Venyan (2015-12-02). "Inson translesion sintezi DNK polimerazalari bilan oksidlangan abazik lezyon ta'sirida paydo bo'lgan DNKning o'zaro bog'liqlik analogining mutagenik aylanib o'tishi". Biokimyo. 54 (50): 7409–7422. doi:10.1021 / acs.biochem.5b01027. PMC 4700817. PMID 26626537.
- ^ Chjao, Linlin; Xu, Venyan (2016-10-21). "Ey6-2′-Deoksiguanosin-butilen-O6-2′-deoksiguanozin DNKning o'zaro bog'liqligi replikatsiya-blokirovka qiluvchi va mutagenli DNK lezyonlari ". Kimyoviy. Res. Toksikol. 29 (11): 1872–1882. doi:10.1021 / acs.chemrestox.6b00278. PMC 5665164. PMID 27768841.
- ^ Cole RS, Levitan D, Sinden RR (1976). "Escherichia coli DNKsidan psoralen interstrand o'zaro bog'liqliklarini olib tashlash: mexanizm va genetik nazorat". J. Mol. Biol. 103 (1): 39–59. doi:10.1016/0022-2836(76)90051-6. PMID 785009.
- ^ Sinden RR, Koul RS (1978). "Psoralen va nur bilan ishlangan o'zaro bog'langan DNKni tiklash va Escherichia coli-ning omon qolishi: genetik rekombinatsiya va DNK metabolizmiga ta'sir qiluvchi mutatsiyalarning ta'siri". J. Bakteriol. 136 (2): 538–47. PMC 218577. PMID 361714.
- ^ Noll DM, Meyson TM, Miller PS (2006). "DNKda interstrand o'zaro bog'liqliklarini shakllantirish va ta'mirlash". Kimyoviy. Vah. 106 (2): 277–301. doi:10.1021 / cr040478b. PMC 2505341. PMID 16464006.
- ^ Tretyakova, Natalya; Groler, Arnold; Ji, Shaofei (2015). "DNK-oqsilning o'zaro bog'liqligi: shakllanishi, tuzilishi va biologik natijalari". Acc Chem Res. 48 (6): 1631–44. doi:10.1021 / hisob raqamlari.5b00056. PMC 4704791. PMID 26032357.
- ^ Pendergrast, P .; Chen, Yan; Ebrayt, Yon; Ebrayt, Richard. "Oqsil-DNK kompleksidagi DNK bilan bog'lanish motifining yo'nalishini fotokrossing orqali aniqlash" (PDF). Proc. Natl. Akad. Ilmiy ish. AQSH. doi:10.1073 / pnas.89.21.10287. PMC 50323. PMID 1332042.
- ^ Smit, Kendrik; Martin, Shetlar. "DNK-PROTEIN CROSSLINKS". Iqtibos jurnali talab qiladi
| jurnal =(Yordam bering) - ^ Molina JR, Yang P, Kassivi SD, Schild SE, Adjei AA (2008). "Kichik hujayrali bo'lmagan o'pka saratoni: epidemiologiya, xavf omillari, davolash va omon qolish". Mayo klinikasi. Proc. 83 (5): 584–94. doi:10.4065/83.5.584. PMC 2718421. PMID 18452692.
- ^ a b Taron M, Rosell R, Felip E, Mendez P, Souglakos J, Ronco MS, Queralt C, Majo J, Sanches JM, Sanches JJ, Maestre J (2004). "BRCA1 mRNA ekspression darajasi o'pka saratonida kimyoviy qarshilik ko'rsatkichi sifatida". Hum. Mol. Genet. 13 (20): 2443–9. doi:10.1093 / hmg / ddh260. PMID 15317748.
- ^ a b Papadaki C, Sfakianaki M, Ioannidis G, Lagoudaki E, Trypaki M, Tryfonidis K, Mavroudis D, Stathopoulos E, Georgoulias V, Souglakos J (2012). "Birlamchi o'smadagi ERCC1 va BRAC1 mRNA ekspression darajasi metastatik kichik hujayrali bo'lmagan o'pka saratoni bilan oldindan davolangan bemorlarda sisplatin asosidagi ikkinchi darajali kimyoviy terapiyaning samaradorligini taxmin qilishi mumkin". J Torak Onkol. 7 (4): 663–71. doi:10.1097 / JTO.0b013e318244bdd4. PMID 22425915.
- ^ Stingele, Julian; Belleli, Roberto; Boulton, Simon (2017 yil sentyabr). "DNK - oqsilning o'zaro bog'liqligini tiklash mexanizmlari". Molekulyar hujayra biologiyasi. 18 (9): 563–573. doi:10.1038 / nrm.2017.56. PMID 28655905.
Tashqi havolalar
- PDB: 1AIO – Sisplatin va DNK qo'shimchasini hosil qilish uchun interaktiv tuzilish
- PDB: 204D – Psoralen va o'zaro bog'langan DNK uchun interaktiv tuzilish
- Psoralen ultrabinafsha nurli davolash