Odamning bezgakka genetik qarshiligi - Human genetic resistance to malaria - Wikipedia
Ushbu maqolada bir nechta muammolar mavjud. Iltimos yordam bering uni yaxshilang yoki ushbu masalalarni muhokama qiling munozara sahifasi. (Ushbu shablon xabarlarini qanday va qachon olib tashlashni bilib oling) (Ushbu shablon xabarini qanday va qachon olib tashlashni bilib oling)
|
Odamning bezgakka genetik qarshiligi -dagi merosxo'r o'zgarishlarga ishora qiladi DNK qarshilikni kuchaytiradigan odamlarning bezgak va natijada ushbu genetik o'zgarishlarga ega bo'lgan odamlarning omon qolish darajasi oshadi. Bularning mavjudligi genotiplar ehtimol tufayli evolyutsion bosim jinsning parazitlari tomonidan amalga oshiriladi Plazmodium bezgak kasalligini keltirib chiqaradi. Bezgak yuqtirganligi sababli qizil qon hujayralari, bu genetik o'zgarishlar ko'pincha qizil qon hujayralarining ishlashi uchun zarur bo'lgan molekulalarning o'zgarishi (va shuning uchun parazitlarning omon qolishi), masalan gemoglobin yoki boshqa hujayralardagi oqsillar yoki eritrotsitlar fermentlari. Ushbu o'zgarishlar, odatda, qizil qon hujayralarini bosqinlardan himoya qiladi Plazmodium qizil qon tanachasida parazitlar yoki parazitlarning ko'payishi.
Gemoglobin yoki boshqa xarakterli oqsillarning irsiy o'zgarishi, bu sutemizuvchilar biokimyosining juda muhim va o'zgarmas xususiyatlari bo'lib, odatda irsiy kasallikni keltirib chiqaradi. Shuning uchun ular odatda nomlari bilan ataladi qon kasalliklari ular bilan bog'liq, shu jumladan o'roqsimon xastalik, talassemiya, glyukoza-6-fosfat dehidrogenaza etishmovchiligi va boshqalar. Ushbu qon kasalliklari ko'payib boradi kasallanish va bezgak kam tarqalgan dunyodagi o'lim.
Bezgakka genetik qarshilikni rivojlantirish
Mikroskopik parazitlar, viruslar, bezgakni keltirib chiqaradigan protozoanlar va boshqalar singari, o'zlarini ko'paytira olmaydi va hayot tsikllarini davom ettirish uchun mezbonga ishonadi. Ular xostlar xujayralarini bosib olish va o'zlarini takrorlash uchun uyali aloqa vositalarini o'zlashtirish orqali takrorlanadi. Oxir oqibat, tekshirilmagan replikatsiya hujayralarni yorilishiga olib keladi, hujayralarni o'ldiradi va yuqumli organizmlarni boshqa hujayralarga yuqtirishi mumkin bo'lgan qon oqimiga chiqaradi. Hujayralar nobud bo'lganda va invaziv organizm replikatsiyasining toksik mahsulotlari to'planganda kasallik alomatlari paydo bo'ladi. Ushbu jarayon yuqumli organizm tomonidan ishlab chiqariladigan o'ziga xos oqsillarni va mezbon hujayrani o'z ichiga olganligi sababli, juda muhim bo'lgan oqsilning ozgina o'zgarishi ham infektsiyani qiyinlashtirishi yoki imkonsiz qilishi mumkin. Bunday o'zgarishlar oqsilni kodlaydigan genning mutatsion jarayoni natijasida paydo bo'lishi mumkin. Agar o'zgarish jinsiy hujayrada bo'lsa, ya'ni spermatozoid yoki tuxum birlashib, odam bo'lib o'sadigan zigota hosil qilsa, himoya mutatsiya meros qilib olinadi. O'limga olib keladigan kasalliklar himoya mutatsiyasiga ega bo'lmagan ko'plab odamlarni o'ldirganligi sababli, vaqt o'tishi bilan o'limga olib keladigan kasalliklar tarqalgan mintaqalarda ko'plab odamlar himoya mutatsiyalariga ega bo'lishadi.
Qachon P. falciparum parazit xujayra xujayrasini yuqtiradi, u qizil qon hujayralari membranasining xususiyatlarini o'zgartiradi va uni boshqa hujayralarga "yopishqoq" qiladi. Parazitlangan qizil qon hujayralarining klasterlari kapillyar qon aylanish hajmidan oshib ketishi mumkin endoteliy va aylanishni blokirovka qilish. Ushbu to'siqlar miyani o'rab turgan qon tomirlarida paydo bo'lganda, ular sabab bo'ladi miya yarim gipoksiya, ni natijasida nevrologik belgilari ma'lum miya bezgagi. Ushbu holat chalkashlik, yo'nalishni buzish va ko'pincha terminal bilan tavsiflanadi koma. Bu bezgak o'limining 80% ni tashkil qiladi. Shuning uchun bezgak infektsiyasidan va o'limdan himoya qiluvchi mutatsiyalar sezilarli ustunlikka ega.
Bezgak ma'lum bo'lgan eng kuchli odamni joylashtirdi selektiv bosim ustida inson genomi so'nggi 10 ming yil ichida qishloq xo'jaligi paydo bo'lganidan beri.[1][2] Plazmodium falciparum orasida joy olishga qodir emas edi Afrika kattalashguncha populyatsiyalar harakatsiz jamoalar Afrikada mahalliy qishloq xo'jaligi evolyutsiyasi bilan birgalikda paydo bo'lgan ( qishloq xo'jaligi inqilobi ). Bir nechta meros qilib olingan variantlar qizil qon hujayralari Tanlov natijasida bezgak tez-tez uchraydigan dunyoning ba'zi joylarida keng tarqalgan parazit.[3] Ushbu tanlov tarixiy jihatdan birinchi hujjatlashtirilgan misol sifatida muhim edi kasallik agenti sifatida tabiiy selektsiya yilda odamlar. Bu shuningdek, genetik jihatdan boshqariladigan birinchi misol edi tug'ma immunitet yuqumli kasalliklarning dastlabki bosqichida ishlaydi, adaptiv immunitetdan oldin, bir necha kundan keyin ta'sir qiladi. Bezgakda, boshqa kasalliklarda bo'lgani kabi, tug'ma immunitet olib keladi va rag'batlantiradi, adaptiv immunitet.
Mutatsiyalar zararli va foydali ta'sirga ega bo'lishi mumkin va har qanday mutatsiya ikkalasiga ham ta'sir qilishi mumkin. Bezgakning yuqishi hujayralar devorlarida va eritrotsitlarning boshqa joylarida mavjud bo'lgan o'ziga xos oqsillarga bog'liq. Himoya mutatsiyalari ushbu oqsillarni bezgak organizmlari uchun mavjud bo'lmaydigan qilib o'zgartiradi. Ammo, bu o'zgarishlar, shuningdek, ochiq yoki qizil qon tanachalarini mikroskopik tekshiruvi natijasida ko'rinadigan ta'sirga ega bo'lishi mumkin bo'lgan qizil qon hujayralarining faoliyati va shaklini o'zgartiradi. Ushbu o'zgarishlar eritrotsitlar faoliyatini turli xil yo'llar bilan buzishi mumkin, bu esa insonning sog'lig'iga yoki uzoq umr ko'rishiga zararli ta'sir ko'rsatadi. Ammo, agar bezgakdan himoya qilishning aniq ta'siri boshqa zararli ta'sirlardan ustun bo'lsa, himoya mutatsiyasi saqlanib qoladi va avloddan avlodga tarqaladi.
Bezgak infektsiyasidan himoya qiluvchi, ammo qizil qon hujayralarini ishdan chiqaradigan bu o'zgarishlar qonning buzilishi deb hisoblanadi, chunki ular ochiq va zararli ta'sirga ega. Ularning himoya funktsiyasi faqat so'nggi paytlarda aniqlandi va tan olindi. Ushbu buzilishlarning ba'zilari o'roqsimon hujayrali anemiya, talassemiya, glyukoza-6-fosfat dehidrogenaza etishmovchiligi, ovalotsitoz, ellipotsitoz va Gerbich antigeni va Duffy antigenini yo'qotish kabi xayoliy va sirli nomlar bilan ma'lum. Ushbu nomlar turli xil oqsillarni, fermentlarni va qizil qon hujayralarining shakli yoki funktsiyasini anglatadi.
Tug'ma qarshilik
Genetika bilan boshqariladigan tug'ma qarshilikning kuchli ta'siri bezgak kasalligi tarqalgan joylarda yosh bolalarning omon qolish ehtimolida aks etadi. Tug'ma immunitetni sezgir yosh guruhida (to'rt yoshdan kichik) o'rganish kerak, chunki katta yoshdagi bolalar va kattalarda tug'ma immunitetning ta'siri adaptiv immunitetga ta'sir qiladi. Shuningdek, tasodifiy foydalaniladigan populyatsiyalarni o'rganish kerak bezgakka qarshi dorilar sodir bo'lmaydi. Umurtqali hayvonlar infektsiyasiga, shu jumladan odamlarga tug'ma qarshilik ko'rsatish bo'yicha ba'zi dastlabki hissa 1-jadvalda keltirilgan.
| Kashf etilgan yil | Patogen | Qarshilik mexanizmi | Mualliflar |
|---|---|---|---|
| 1954 | P. falciparum | O'roqsimon hujayralardagi heterozigota | Allison[4] |
| 1975 | P. knowlesi | Duffy antigenining qizil hujayralarga ta'sir etmasligi | Miller va boshq. |
| 1976 | P. vivax | Duffy antigenining qizil hujayralarga ta'sir etmasligi | Miller va boshq.[5] |
Kashshof tadqiqotlarning ikkitasi bezgak bilan bog'liqligi diqqatga sazovordir. In Toll retseptorlari bo'yicha klassik tadqiqotlar Drosophila mevali chivin[6] tez kengaytirildi Pullikga o'xshash retseptorlar yilda sutemizuvchilar[7] va keyin boshqasiga naqshni aniqlash retseptorlari, tug'ma immunitetda muhim rol o'ynaydigan. Biroq, bezgakka dastlabki qo'shilishlar vaqt sinovidan o'tgan tug'ma qarshilikning klassik namunalari bo'lib qolmoqda.
Himoya qilish mexanizmlari
Anormal gemoglobinlarni o'z ichiga olgan yoki G6PD etishmasligi bo'lgan eritrotsitlar qisman himoyalangan mexanizmlar P. falciparum infektsiyalar to'liq tushunilmagan, garchi takliflar etishmasligi bo'lsa. Replikatsiya bezgakning periferik qon bosqichida parazitlar yuqori darajaga ega kislorod iste'mol[8] va ko'p miqdordagi gemoglobinni yutadi.[9] Ehtimol, endotsitik pufakchalardagi HbS oksigensizlanadi, polimerlanadi va kam hazm qilinadi. Anormal gemoglobinlarni o'z ichiga olgan yoki G6PD etishmasligi bo'lgan qizil hujayralarda, kislorod radikallari ishlab chiqariladi va bezgak parazitlari qo'shimcha oksidlanish stresini keltirib chiqaradi.[10] Buning natijasida qizil hujayra membranalari o'zgarishi mumkin, shu jumladan translokatsiya fosfatidilserin ularning yuzasiga[jargon ]so'ngra makrofagni tanib olish va yutish.[11] Mualliflarning ta'kidlashicha, bu mexanizm odatdagi qizil hujayralarga qaraganda g'ayritabiiy tarzda sodir bo'lishi mumkin va shu bilan avvalgi ko'payishni cheklaydi. Bundan tashqari, parazitlangan o'roq hujayralarining endotelial hujayralar bilan birikishi o'zgarganligi sababli sezilarli darajada kamayadi. P. falciparum eritrotsitlar membranasi oqsili-1 (PfMP-1).[12] Ushbu oqsil parazitning hujayra sirtidagi asosiy sitoaderentsiya ligand va virulentlik omilidir. Parazit replikatsiyasining so'nggi bosqichlarida qizil hujayralar venoz endoteliyga yopishadi va bu birikmani inhibe qilish replikatsiyani bostirishi mumkin.
O'roq gemoglobin gem oksigenaz-1 ning ekspressionini keltirib chiqaradi gemopoetik hujayralar. Uglerod oksidi, tomonidan gem katabolizmining yon mahsuloti gem oksigenaz -1 (HO-1), keyin aylanib yuradigan erkin gemning to'planishiga to'sqinlik qiladi Plazmodium eksperimental miya bezgagi patogenezini bostiruvchi infektsiya.[13] Boshqa mexanizmlar, masalan, HO-1 vositachiligidagi kasalliklarga chidamliligi va parazitga mezbon mikro-RNKning translokatsiyasi tufayli parazitar o'sishni pasayishi.[14]
Tug'ma qarshilik turlari
Bezgakdan himoya qilishning birinchi usuli asosan anormal gemoglobinlar va glyukoza-6-fosfat dehidrogenaza etishmovchiligi bilan amalga oshiriladi. Irsiy genetik qarshilikning uchta asosiy turi - o'roqsimon hujayra kasalligi, talassemiya va G6PD etishmovchiligi - mavjud bo'lgan O'rta er dengizi vaqti bilan dunyo Rim imperiyasi.
Gemoglobin anormalliklari
Anormal gemoglobinlarning tarqalishi
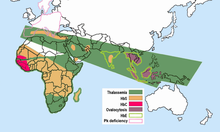
Bezgak dunyoning tropik va subtropik mintaqalaridagi tog'li hududlarning salqinroq, quruqroq iqlim sharoitida ro'y bermaydi, o'n minglab odamlar o'rganilgan va bezgak bo'lmagan biron bir populyatsiyada g'ayritabiiy gemoglobinlarning yuqori chastotalari topilmagan. Turli populyatsiyalardagi g'ayritabiiy gemoglobinlarning chastotalari juda xilma-xil, ammo ba'zilari, shubhasiz, polimorfikdir, chastotalari takroriy mutatsiya kutilganidan yuqori. Shubhasiz, bezgak tanlovi ushbu polimorfizmlarning tarqalishida katta rol o'ynagan. Bularning barchasi bezgakli joylarda,
- O'roq xujayrasi - o'roq xujayrasi bilan bog'liq bo'lgan HbS geni bugungi kunda Afrikaning Sahroi osti qismida, Yaqin Sharqda va Hindiston yarim orolining keng qismida tarqalgan bo'lib, bu erda tashuvchilarning chastotalari aholining 5-40% va undan ko'prog'ini tashkil qiladi. Bezgak sohalarida o'roqsimon hujayralardagi heterozigotlarning chastotalari 20-40% ni tashkil etdi Keniya, Uganda va Tanzaniya. Keyinchalik ko'plab tergovchilar tomonidan olib borilgan tadqiqotlar rasmni to'ldirdi.[15][16] HbS genining yuqori chastotalari keng kamar bilan chegaralanadi Markaziy Afrika, lekin ko'pini hisobga olmaganda Efiopiya va Sharqiy Afrikaning baland tog'lari; bu bezgak yuqtirish sohalariga to'g'ri keladi. 20% gacha bo'lgan o'roq xujayrasi geterozigota chastotalari ham cho'ntaklarda uchraydi Hindiston va Gretsiya ilgari juda bezgakka chalingan.
Talassemiyalar O'rta er dengizi havzasi va Afrikaning ayrim qismlaridan, butun O'rta Sharq, Hindiston yarim oroli, Janubi-Sharqiy Osiyo, Melaneziya va Tinch okean orollariga qadar cho'zilgan keng diapazonga ega.
- a-talassemiya, bu G'arbiy Afrikaning ayrim qismlarida 30% chastotaga etadi;[17]
- b-talassemiya, Italiyaning ayrim qismlarida chastotasi 10% gacha;
- HbE, Tailandda va boshqa Janubi-Sharqiy Osiyo mamlakatlarida chastotalarni 55% gacha oshiradi;[18] HbE Hindiston yarim sharqining sharqiy qismida va butun Janubi-Sharqiy Osiyoda uchraydi, bu erda ba'zi hududlarda tashuvchilar darajasi aholining 60 foizidan oshishi mumkin.
- HbC Shimoliy Gana va Burkina-Fasoda 20% ga yaqin chastotalarga ega. HbC G'arbiy va Shimoliy Afrikaning ayrim qismlarida cheklangan.[iqtibos kerak ]
- bir vaqtning o'zida polimorfizmlar - HbS va b-talassemiya, va HbS va HbC uchun er-xotin heterozigotlar, o'roqsimon hujayrali kasallikning SSga nisbatan engilroq variantlaridan aziyat chekadi, ammo zamonaviy davolanishga qadar fitnesni pasaytiradi. Bashorat qilinganidek, ushbu variant allellari populyatsiyalarda o'zaro eksklyuziv bo'lishga moyil. Yunonistonning turli qismlarida HbS va b-talassemiya chastotalari va G'arbiy Afrikada HbS va HbC chastotalari o'rtasida salbiy bog'liqlik mavjud.[19] Mutatsiyalarning nojo'ya o'zaro ta'siri bo'lmagan joylarda, g'ayritabiiy gemoglobinlar va G6PD etishmovchiligida bo'lgani kabi, populyatsiyalarda ushbu variant allellarning ijobiy o'zaro bog'liqligi kutiladi va topiladi.[19]
O'roqsimon hujayra
Ushbu bo'lim Mendeliyaning genetikasi, o'roqsimon hujayra kasalligi, bezgakdan himoya darajasi, ta'sirlangan plazmodiya turlari, oqsillar kimyosi haqida ma'lumot etishmayapti. (2014 yil aprel) |
O'roq-hujayra kasalligi ma'lum bir oqsilning mutatsiyasiga bog'liq bo'lgan genetik kasallik edi. Poling genetik yo'l bilan yuqadigan molekulyar kasallik sifatida o'zining o'roqsimon hujayra anemiyasi haqidagi muhim kontseptsiyasini taqdim etdi.[20]
O'roqsimon hujayra anemiyasining molekulyar asoslari nihoyat 1959 yilda, Ingram triptik peptid barmoq izlarini olish usullarini takomillashtirganda aniqlandi. 1950-yillarning o'rtalarida, peptidlar va aminokislotalarni ajratishning eng yangi va ishonchli usullaridan biri tripsin fermenti bo'lib, u polipeptid zanjirlarini ikki aminokislotaning lizin va karboksil guruhlari tomonidan hosil bo'lgan kimyoviy bog'lanishlarni maxsus parchalash orqali ajratib yubordi. arginin. Gemoglobin A va S ning kichik farqlari ushbu peptidlarning birida yoki bir nechtasida kichik o'zgarishlarga olib keladi.[21] Ushbu kichik farqlarni aniqlashga urinish uchun Ingram qog'oz elektroforezi va qog'oz xromotografiya usullarini birlashtirdi. Ushbu birikma bilan u ikki o'lchovli usulni yaratdi, bu unga tripsin hazm qilish jarayonida olingan gemoglobin S va A parchalarini qiyosiy ravishda "barmoq izi" olish imkonini berdi. Barmoq izlarida 30 ga yaqin peptid dog'lari aniqlandi, gemoglobin A hazmida aniq ko'rinadigan bitta peptid dog'i bor edi, u barmoq izi gemoglobinida aniq bo'lmagan. HbS gen defekti - bu g-globin genining bitta nukleotidining (A dan T gacha) mutatsiyasidir, aminokislota glutamik kislotani p zanjirning oltinchi pozitsiyasida kam qutbli aminokislota valin bilan almashtiradi.[22]
HbS fiziologik pH-da salbiy zaryad normal kattalar gemoglobiniga qaraganda pastroq. Zaryadlangan aminokislotani hidrofob, neytral aminokislota bilan oddiy almashtirishning natijalari juda kengdir, G'arbiy Afrikada o'tkazilgan so'nggi tadqiqotlar shuni ko'rsatadiki, Hb S ning eng katta ta'siri o'limdan yoki og'ir kasalliklardan himoya qilishga o'xshaydi, ya'ni chuqur anemiya yoki miya bezgagi - infektsiya o'z-o'zidan kamroq ta'sir qiladi. O'roqsimon hujayra geni uchun heterozigotli bolalar, odatdagi gemoglobin geni uchun homozigot bo'lganlar kabi falciparumdan o'lish xavfining atigi o'ndan bir qismiga ega. Parazitlangan o'roq eritrotsitlarining endotelial hujayralar va qon monotsitlari bilan birikishi o'zgargan displey tufayli sezilarli darajada kamayadi. Plazmodium falciparum eritrotsitlar membranasi oqsili 1 (PfEMP-1), eritrotsitlar yuzasida parazitning asosiy sitoaderentsiya ligand va virulentlik omili.[23]
Himoya shuningdek, o'ralgan gemoglobinning beqarorligidan kelib chiqadi, bu esa ustunlik qiluvchi integral hujayra membranasi oqsilini (3-band deb ataladi) to'playdi va fagotsit hujayralari tomonidan tezkor olib tashlanishiga olib keladi. Tabiiy antikorlar bu klasterlarni keksa eritrotsitlarda taniydilar. HbAS bilan himoya qilish nafaqat tug'ma, balki parazitga qarshi immunitetni oshirishni ham o'z ichiga oladi.[24] Vaqtidan oldin denatatsiya qilingan o'roqli gemoglobin bezgakda ham, o'roqsimon hujayra kasalligida ham eritrotsitlar yopishishini boshqaruvchi tabiiy antikorlarning regulyatsiyasini keltirib chiqaradi.[25] Endotelial faollashuviga olib keladigan stimullarga yo'naltirish o'roqning qizil hujayralari yopishqoqligi va vazo-okklyuziyasini inhibe qilishning istiqbolli terapevtik strategiyasini tashkil etadi.[26]
Bu esa, degan farazga olib keldi gomozigotlar chunki o'roqsimon hujayra geni kasallikdan aziyat chekadi, heterozigotlar bezgakdan himoyalangan bo'lishi mumkin.[27] Bezgak o'roqsimon hujayra xususiyati uchun tanlangan omil bo'lib qolmoqda.[28]
Talassemiya
Uzoq vaqt davomida ma'lum bo'lgan anemiya turi ma'lum bo'lgan talassemiya, ba'zi O'rta er dengizi aholisi, shu jumladan yunonlar va janubiy italiyaliklarda yuqori chastotaga ega. Ism yunoncha dengiz so'zlaridan olingan (talassa) ma'nosini anglatadi O'rtayer dengizi va qon (haima). Vernon Ingram talassemiyaning turli shakllarining genetik asoslarini gemoglobinning ikki polipeptid zanjiri sintezidagi muvozanat sifatida tushuntirishga loyiqdir.[29]
Umumiy O'rta er dengizi variantida mutatsiyalar g-zanjiri (b-talassemiya) hosil bo'lishini pasaytiradi. Afrikada va boshqa bir qator mamlakatlarda nisbatan tez-tez uchraydigan a-talassemiyada gemoglobinning a-zanjiri ishlab chiqarilishi buziladi va b-zanjirning nisbatan ortiqcha ishlab chiqarilishi kuzatiladi. B-talassemiya uchun homozigotli odamlar og'ir anemiyaga ega va tirik qolish va ko'payish ehtimoli yo'q, shuning uchun genga qarshi selektsiya kuchli. A-talassemiya uchun homozigota bo'lganlar ham anemiyadan aziyat chekishadi va genga nisbatan ma'lum darajada seleksiya mavjud.
Pastki Himoloy tog 'etaklari va Ichki Teray yoki Doon vodiylari ning Nepal va Hindiston iliq iqlim va quruq davrda er osti suvlari baland tepaliklardan pastga tushib turadigan botqoqliklar tufayli juda bezgaklidir. Bezgak o'rmonlari mudofaa chorasi sifatida Nepal hukmdorlari tomonidan ataylab saqlanib turilgan. Ushbu zonada yashamoqchi bo'lgan odamlar, quritgichdagi balandlikdan yoki pastroqdan o'lim darajasiga qaraganda ancha yuqori bo'lgan Gangetik tekislik. Biroq, Tharu odamlari Ushbu zonada bir necha genlar orqali qarshilikni rivojlantirish uchun etarlicha uzoq vaqt yashagan. Tharu va Tharu bo'lmagan aholisi o'rtasida tibbiy tadqiqotlar Teray Tarus orasida qoldiq bezgak holatlarining tarqalishi qariyb etti baravar past ekanligi haqida dalillar keltirdi. Qarshilik uchun asos a-Talassemiya genining mahalliy aholi tarkibidagi gomozigotliligi sifatida belgilandi.[30] Endogamiya Kasta va etnik yo'nalishlar bo'yicha ushbu genlarning qo'shni populyatsiyalarda keng tarqalishiga to'sqinlik qilgan ko'rinadi.[31]
HbC va HbE eritroidlari
A-talassemiya, HbC va HbE bilan kasallangan odamlarning parazitdan ma'lum darajada himoya qilishiga oid dalillar mavjud.[17][32]Gemoglobin C (HbC) anormal gemoglobin bo'lib, lizin qoldig'ini b-globin zanjirining glutamik kislota qoldig'iga almashtiradi, HbS mutatsiyasi bilan aynan ß-6 pozitsiyasida. HbC uchun "C" belgisi u kashf etilgan shahar nomidan olingan - Yangi Zelandiya, Kristchurch. Ushbu kasallikka chalingan odamlarda, ayniqsa bolalarda, qorin va bo'g'imlarda og'riq, taloq kattalashishi va engil sariqlik bo'lishi mumkin, ammo ular o'roqsimon hujayra kasalliklarida bo'lgani kabi og'ir inqirozlarga duch kelmaydi. Gemoglobin S G'arbiy Afrikaning bezgakli hududlarida, ayniqsa Burkina-Fasoda keng tarqalgan. Burkina-Fasoda 4 348 Mossi sub'ektida o'tkazilgan katta ish nazorati bo'yicha tekshiruvda HbC HbAC heterozigotlarida klinik bezgak xavfining 29% ga va HbCC homozigotlarida 93% ga kamayishi bilan bog'liq. HbC, HbS ning muvozanatli polimorfizmi orqali politsentrik "tez, ammo qimmat" moslashuv bilan taqqoslaganda, vaqtinchalik polimorfizm orqali bezgakka "sekin, ammo bepul" genetik moslashishni anglatadi.[33][34]HbC variant antigenining miqdori va tarqalishini o'zgartiradi P. falciparum infektsiyalangan eritrotsitlar yuzasida eritrotsitlar membranasi oqsili 1 (PfEMP1) va bezgak yuzasi oqsillarining o'zgartirilgan namoyishi parazitlarning yopishqoqligini pasaytiradi (shu bilan taloq tomonidan tozalanishiga yo'l qo'ymaydi) va og'ir kasallik xavfini kamaytirishi mumkin.[35][36]
Gemoglobin E 26-pozitsiyada glutamat-lizin o'rnini bosadigan beta zanjir genidagi bitta nuqta mutatsiyasiga bog'liq. Bu eng keng tarqalgan gemoglobinopatiyalardan biri bo'lib, 30 million odam zarar ko'rgan. Gemoglobin E Janubi-Sharqiy Osiyoda juda keng tarqalgan. HbE eritrotsitlari noma'lum membrana anormalligiga ega, bu RBC populyatsiyasining aksariyat qismini bosqinlarga nisbatan nisbatan chidamli qiladi. P falciparum.[37]
Boshqa eritrotsitlar mutatsiyalari
Gemoglobin anormalliklari bilan bir qatorda qarshilik ko'rsatadigan boshqa genetik mutatsiyalar Plazmodiya infektsiya hujayra yuzasining o'zgarishini o'z ichiga oladi antigenik oqsillar, hujayra membranasi tarkibidagi oqsillar yoki fermentlar glikoliz.
Glyukoza-6-fosfat dehidrogenaza etishmovchiligi

Glyukoza-6-fosfat dehidrogenaza (G6PD) muhim ahamiyatga ega ferment metabolizmga uchragan qizil hujayralarda glyukoza orqali pentoza fosfat yo'li, katabolik oksidlanishning (glikoliz) anabolik alternativasi, kamaytiruvchi muhitni saqlab turganda. G6PD insonning barcha hujayralarida mavjud, ammo qizil qon hujayralari uchun juda muhimdir. Yetuk qizil qon hujayralari etishmasligi sababli yadrolar va sitoplazmatik RNK, ular genetik g'ayritabiiy yoki qarish o'rniga yangi ferment molekulalarini sintez qila olmaydi. Barcha oqsillar, shu jumladan fermentlar, odatda 120 kun bo'lgan qizil qon tanachasining butun hayoti davomida davom etishi kerak.
1956 yilda Alving va uning hamkasblari buni ba'zilarida ko'rsatdilar Afroamerikaliklar bezgakka qarshi preparat primakvin gemolitik anemiya keltirib chiqaradi va bu odamlarda eritrotsitlarda irsiy G6PD etishmovchiligi mavjud.[38] G6PD etishmovchiligi jinsiy aloqada bo'lib, O'rta er dengizi, Afrika va boshqa populyatsiyalarda keng tarqalgan. O'rta er dengizi mamlakatlarida bunday shaxslar gemolitik diatezni rivojlantirishi mumkin (favizm ) iste'mol qilgandan keyin fava loviya. G6PD etishmovchiligi bo'lgan odamlar, shuningdek, primakindan tashqari, bir nechta dorilarga sezgir.
G6PD etishmovchiligi odamlarda eng ko'p uchraydigan ikkinchi ferment tanqisligi (keyin) ALDH2 400 million kishiga ta'sir qilishi taxmin qilinmoqda.[39] Ushbu lokusda ko'plab mutatsiyalar mavjud, ulardan ikkitasi Afrika va O'rta er dengizi aholisining chastotalariga 20% va undan yuqori; bular A va Med mutatsiyalari deb nomlanadi.[40] G6PD-ning mutant navlari tabiiy ravishda uchraydigan fermentga qaraganda beqarorroq bo'lishi mumkin, shuning uchun ularning faolligi qizil hujayralar yoshiga qarab pasayadi.
Ushbu savol Tanzaniya, Sharqiy Afrikada bezgakka qarshi dorilar ishlatilmaydigan izolyatsiya qilingan populyatsiyalarda o'rganilgan[41] va Gambiya Respublikasi, G'arbiy Afrika, bolalarni eng sezgir bo'lgan davrda kuzatib borish falciparum bezgak.[42] Ikkala holatda ham parazitlar soni G6PD etishmovchiligida normal qizil hujayralar fermentlariga qaraganda ancha past edi. Birlashma, shuningdek, odamlarda o'rganilgan, bu mumkin, chunki ferment etishmovchiligi jinsiy aloqada va ayol heterozigotlari mozaikadir lyonizatsiya, bu erda tasodifiy inaktivatsiya X-xromosoma ba'zi hujayralarda normal qizil qon hujayralari bilan birga bo'lgan G6PD etishmayotgan qizil qon hujayralari populyatsiyasini hosil qiladi. Bezgak parazitlari ferment etishmaydigan hujayralarga qaraganda normal qizil hujayralarda sezilarli darajada tez-tez kuzatilgan.[43] G6PD tanqisligi genlarini bezgak selektsiyasining evolyutsion genetik tahlili Tishkoff va Verelli tomonidan nashr etilgan.[40] Ferment etishmovchiligi bezgakka chalingan yoki ilgari bezgak bilan kasallangan, ammo boshqa joylarda bo'lmagan ko'plab mamlakatlarda keng tarqalgan.
PK etishmovchiligi
Piruvat kinaz (PK) etishmovchiligi, shuningdek, eritrotsitlar piruvat kinaz etishmovchiligi deb ataladi, bu piruvat kinaza fermentining irsiy metabolik buzilishi. Bunday holatda piruvat kinaz etishmasligi glikoliz jarayonini sekinlashtiradi. Ushbu ta'sir ayniqsa mitoxondriyaga ega bo'lmagan hujayralarda halokatli bo'ladi, chunki bu hujayralar anaerob glikolizni yagona energiya manbai sifatida ishlatishi kerak, chunki TCA tsikli mavjud emas. Masalan, qizil qon hujayralari, ular piruvat kinaz etishmovchiligi holatida tezda ATP etishmovchiligiga uchraydi va gemolizga uchrashi mumkin. Shuning uchun piruvat kinaz etishmovchiligi gemolitik anemiyani keltirib chiqarishi mumkin.
Ning zo'ravonligi o'rtasida sezilarli bog'liqlik mavjud PK etishmovchiligi va bezgakdan himoya darajasi.[44]
Elliptitoz
Ellipotsitoz qonning buzilishi bo'lib, unda bemorning g'ayritabiiy eritrotsitlari elliptikdir. Ta'sir qilganlar orasida juda ko'p genetik o'zgaruvchanlik mavjud. Irsiy elliptotsitozning uchta asosiy shakli mavjud: oddiy irsiy elliptotsitoz, sferotsitik elliptotsitoz va janubi-sharqiy Osiyo ovalotsitozi.
Janubi-sharqiy Osiyo ovalotsitozi

Ovalotsitoz bu ellipotsitozning kichik turi bo'lib, eritrotsitlar yumaloq shakl o'rniga ovalga ega bo'lgan irsiy holatdir. Ko'pgina populyatsiyalarda ovalotsitoz kam uchraydi, ammo Janubiy-Sharqiy Osiyo ovalotsitozi (SAO) mahalliy aholining 15 foizida uchraydi. Malayziya va of Papua-Yangi Gvineya. SAO eritrotsitlarining bir nechta anormalliklari, shu jumladan qizil hujayra qattiqligining oshishi va ba'zi qizil hujayra antijenlerinin ekspresyonunun pasayishi haqida xabar berilgan.[46]SAO eritrotsitni kodlovchi gen mutatsiyasidan kelib chiqadi 3-band oqsil. Genda 400-408 kodonlarning yo'q bo'lib ketishi, 3-guruh oqsilining sitoplazmatik va transmembranali domenlari chegarasida 9 ta aminokislotaning yo'q qilinishiga olib keladi.[47] 3-band membranadan iborat skelet uchun asosiy bog'lanish joyi bo'lib xizmat qiladi, bu submembran oqsil tarmog'idir ankirin, spektrin, aktin va 4.1 tasma. Ovalotsitlar tasmasi 3 normal tasma 3 ga nisbatan ankirin bilan qattiqroq bog'lanadi, bu membrana skeletini 3-band anion tashuvchisi bilan bog'laydi. Ushbu sifat nuqsonlari qizil qon hujayralari membranasini hosil qiladi, ular siljish stressiga kamroq bardoshli va doimiy deformatsiyaga ko'proq moyil.
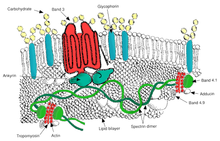
SAO bolalardagi miya bezgagidan himoya bilan bog'liq, chunki u parazitlangan eritrotsitlarning sekvestratsiyasini kamaytiradi. P. falciparum miya mikrovaskulyatsiyasida.[48] Yopish P. falciparumCD36 ga qadar infektsiyalangan qizil qon hujayralari miya bezgagidan himoya qiluvchi SAO xususiyati bilan kuchayadi. SAO odamlarida CD36 orqali sekvestratsiya qilishning yuqori samaradorligi sekvestrlangan infektsiyalangan qizil qon hujayralarining boshqa organ taqsimlanishini aniqlashi mumkin. Bular SAO tomonidan miya bezgagiga qarshi tanlangan afzalligi uchun mumkin bo'lgan tushuntirishni beradi.[49]
Duffy antigen retseptorlari salbiyligi
Plazmodium vivax tropik mamlakatlarda keng tarqalgan, ammo G'arbiy va Markaziy Afrikadagi katta mintaqada yo'q yoki kam uchraydi, bu yaqinda PCR turlarini terish bilan tasdiqlangan.[50] Tarqatishdagi bu bo'shliq ko'plab Sahroi Afrikaliklarning qizil hujayralarida ximokinlar uchun Duffy antigen retseptorlari ekspressionining etishmasligi bilan bog'liq. Duffy salbiy shaxslar DARC alleli uchun homozigot bo'lib, bitta nukleotid mutatsiyasini (DARC 46 T → C) olib yuradilar, bu esa hGATA1 eritroid chizig'i transkripsiyasi omili uchun bog'lanish joyini buzadi.[jargon ][51] Keng tarqalgan in vitro va jonli ravishda tadqiqotlar, Miller va boshq. Daffi qon guruhi retseptoridir P. vivax va qizil hujayralarda Duffy qon guruhining yo'qligi qarshilik omilidir P. vivax afrikadan kelib chiqqan shaxslarda.[5] Maqsad hujayralarida agent uchun retseptor yo'qligi sababli, bu yuqumli kasallikka qarshi tug'ma qarshilikning taniqli namunasi bo'ldi.
Ammo, kuzatuvlar Millerning asl ma'ruzasi malakaga muhtojligini ko'rsatmoqda. Inson tadqiqotlarida P. vivax uzatish, etkazish uchun dalillar mavjud P. vivax G'arbiy Keniyadagi salbiy ta'sirli populyatsiyalar orasida,[52] braziliyalik Amazon mintaqa,[53] va Madagaskar.[54] The Malagasiya xalqi Madagaskarda turli millatlarga mansub Duffy-pozitiv va Duffy-salbiy odamlarning aralashmasi mavjud.[55] Orol aholisining 72% Duffy-salbiy ekanligi aniqlandi. P. vivax ijobiy, 476 asemptomatik Duffy-salbiy odamning 8,8 foizida va klinikada aniqlandi P. vivax bunday 17 kishida bezgak kasalligi aniqlangan. Genotiplash shuni ko'rsatdiki, ko'plik P. vivax shtammlar Duffy-salbiy odamlarning qizil hujayralariga kirib borardi. Mualliflar, Malagas populyatsiyasida chivin yuqishini va jigar infektsiyasini saqlab qolish uchun Duffy-musbat odamlar etarli ekanligini ta'kidlamoqda. Yaqinda Duffy salbiy shaxslari ikki xil shtammni yuqtirishdi P. vivax topilgan Angola va Ekvatorial Gvineya; yanada, P. vivax yuqumli kasalliklar odamlarda ham, chivinlarda ham topilgan, bu esa faol yuqish sodir bo'lishini anglatadi. Bunday uzatish chastotasi hali ham noma'lum.[56] Dunyoning turli burchaklaridan kelgan bir nechta hisobotlar tufayli ba'zi variantlari aniq P. vivax qizil hujayralarida DARC ni ko'rsatmaydigan odamlarga yuqadi. Xuddi shu hodisa Yangi Dunyo maymunlarida ham kuzatilgan. [Izoh 1] Biroq, DARC hali ham odamga yuqadigan asosiy retseptor bo'lib ko'rinadi P. vivax.
Afrikada Duffy negativining tarqalishi aniq bilan o'zaro bog'liq emas P. vivax yuqish.[50] Parfit yuqadigan Sharqiy Afrikada Duffy negativining chastotalari yuqori (80% dan yuqori), G'arbiy Afrikada bo'lgani kabi. Ning kuchi P. vivax tabiiy selektsiya agenti sifatida noma'lum va har bir joyda farq qilishi mumkin. DARC negativligi infektsiyaga tug'ma qarshilik ko'rsatishning yaxshi namunasi bo'lib qolmoqda, ammo u nisbatan qarama-qarshilikni keltirib chiqaradi P. vivax yuqish.

Gerbich antigen retseptorlari salbiyligi
The Gerbich antigen tizimi eritrotsitning ajralmas membrana oqsilidir va eritrotsitlar shaklini saqlashda funktsional jihatdan muhim rol o'ynaydi. Shuningdek, u retseptorlari vazifasini bajaradi P. falciparum eritrotsitlarni bog'laydigan oqsil. To'rtta allellar antigenni kodlaydigan genning Ge-1dan Ge-4gacha. Ge antigenining salbiy uch turi ma'lum: Ge-1, -2, -3, Ge-2, -3 va Ge-2, + 3. nisbatan kam uchraydigan fenotipi bo'lgan Ge-1, -2, -3 bo'lgan shaxslar bosqinga kam ta'sir qiladi (nazorat darajasining ~ 60%). P. falciparum. Bunday shaxslar shartning pastki turiga ega deb nomlanadi irsiy ellipoksitoz, oval yoki elliptik shakldagi eritrotsitlar bilan tavsiflanadi.
Boshqa noyob eritrotsitlar mutatsiyalari
Ning noyob mutatsiyalari glikoforin A va B oqsillar qarshilikka vositachilik qilishlari ham ma'lum P. falciparum.
Odamning leykotsit antijeni polimorfizmlari
G'arbiy Afrikada keng tarqalgan, ammo boshqa irqiy guruhlarda kam uchraydigan inson leykotsit antijeni (HLA) polimorfizmlari og'ir bezgakdan himoya qilish bilan bog'liq. Ushbu genlar guruhi hujayra yuzasida antigen taqdim qiluvchi oqsillarni kodlaydi va boshqa ko'plab funktsiyalarga ega. G'arbiy Afrikada ular kasallik bilan kasallanish darajasining o'roqsimon hujayrali gemoglobin varianti kabi kamaygan. Tadqiqotlar shuni ko'rsatadiki, ning odatiy bo'lmagan polimorfizmi asosiy gistosayish kompleksi genlar asosan yuqumli patogenlar tomonidan tabiiy tanlanish orqali rivojlandi.
Antigen taqdimotida qatnashadigan oqsillarni kodlovchi HLA lokuslaridagi polimorfizmlar bezgak jarayoniga ta'sir qiladi. G'arbiy Afrikada HLA sinf I antigeni (HLA Bw53) va HLA sinf II haplotipi (DRB1 * 13OZ-DQB1 * 0501) og'ir bezgakdan himoya qilish bilan mustaqil ravishda bog'liqdir.[59] Shu bilan birga, HLA korrelyatsiyalari turli xil geografik joylarda farq qiluvchi polimorfik bezgak parazitining genetik konstitutsiyasiga qarab o'zgaradi.[60][61]
Xomilalik gemoglobinning irsiy davomiyligi
Ba'zi tadkikotlar shuni ko'rsatadiki, yuqori darajalar xomilalik gemoglobin (HbF) kattalardagi falciparum bezgakdan himoya qiladi Xomilalik gemoglobinning irsiy davomiyligi.[62]
Bezgak gipotezasini tasdiqlash
Evolyutsion biolog J.B.S. Haldene birinchi bo'lib bezgak va genetik kasallik o'rtasidagi bog'liqlik to'g'risida gipoteza berdi. U birinchi marta 1948 yilda Stokgolmda bo'lib o'tgan "Odam genlarining mutatsion darajasi" mavzusida bo'lib o'tgan Sakkizinchi Xalqaro Genetika Kongressida gipotezasini aytdi.[63] U 1949 yilda nashr etilgan texnik maqolada rasmiylashtirgan va unda bashoratli bayonot bergan: "Anemik heterozigotlarning korpuskalari odatdagidan kichikroq va gipotonik eritmalarga chidamli. Hech bo'lmaganda ularning hujumlariga ham chidamli bo'lishlari mumkin. bezgakni keltirib chiqaradigan sporozoa. "[64] Bu "Haldanening bezgak gipotezasi" yoki qisqacha "bezgak gipotezasi" deb nomlandi.[65]

Yaqin atrofda yashovchi 1022 nafar keniyalik bolalardan iborat kogortani batafsil o'rganish Viktoriya ko'li, 2002 yilda nashr etilgan ushbu bashoratni tasdiqladi.[66] Ko'p SS bolalari hali bir yoshga to'lmasdan vafot etdilar. 2 oydan 16 oygacha AS bolalaridagi o'lim AA bolalariga nisbatan ancha past ekanligi aniqlandi. Ushbu yaxshi tekshirilgan tekshiruv inson populyatsiyasida kasallik orqali tabiiy selektsiyaning doimiy harakatini ko'rsatadi.
Tahlil genomning keng assotsiatsiyasi (GWA) va nozik o'lchamlari assotsiatsiyani xaritalash yuqumli kasalliklar va boshqa kasalliklarga chidamlilik merosini aniqlashning kuchli usuli. GWA-ning afrikaliklarda og'ir falciparum bezgak bilan assotsiatsiyasini ikkita mustaqil dastlabki tahlillari o'tkazildi, ulardan biri Gambiya aholisidagi Malariagen konsortsiumi tomonidan, ikkinchisi Rolf Xorstmann (Gamburgdagi Bernhard Nocht tropik tibbiyot instituti) va uning hamkasblari. . Ikkala holatda ham genomning ahamiyatiga ega bo'lgan assotsiatsiyaning yagona belgisi HBB kodlash joyi β- HbS da anormal bo'lgan gemoglobin zanjiri.[67] Bu HbS falciparum bezgakka tug'ma qarshilik ko'rsatadigan yagona gen ekanligini anglatmaydi; there could be many such genes exerting more modest effects that are challenging to detect by GWA because of the low levels of bog'lanish nomutanosibligi Afrika populyatsiyalarida. However the same GWA association in two populations is powerful evidence that the single gene conferring strongest innate resistance to falciparum malaria is that encoding HbS.
Fitnesses of different genotypes
The fitnesses turli xil genotiplar in an African region where there is intense malarial selection were estimated by Anthony Allison in 1954.[68] In Baamba population living in the Semliki Forest region in Western Uganda the sickle-cell heterozygote (AS) frequency is 40%, which means that the chastota of the sickle-cell gene is 0.255 and 6.5% of children born are SS homozygotes. [Izoh 2]It is a reasonable assumption that until modern treatment was available three quarters of the SS homozygotes failed to reproduce. To balance this loss of sickle-cell genes, a mutatsiya darajasi of 1:10.2 per gene per generation would be necessary. This is about 1000 times greater than mutation rates measured in Drosophila and other organisms and much higher than recorded for the sickle-cell locus in Africans.[69] To balance the polymorphism, Anthony Allison estimated that the fitness of the AS heterozygote would have to be 1.26 times than that of the normal homozygote. Later analyses of survival figures have given similar results, with some differences from site to site. In Gambians, it was estimated that AS heterozygotes have 90% protection against P. falciparum-associated severe anemia and cerebral malaria,[59] whereas in the Luo population of Kenya it was estimated that AS heterozygotes have 60% protection against severe malarial anemia.[66] These differences reflect the intensity of transmission of P. falciparum malaria from locality to locality and season to season, so fitness calculations will also vary. In many African populations the AS frequency is about 20%, and a fitness superiority over those with normal hemoglobin of the order of 10% is sufficient to produce a stable polymorphism.
Shuningdek qarang
Izohlar
- ^ P. vivax can be transmitted in Sincap maymunlari (Saimiri boliviensis va S. sciureus), and Barnwell et al.[57] have obtained evidence that P. vivax kiradi Saimiri monkey red cells independently of the Duffy blood group, showing that P. vivax has an alternative pathway for invading these cells. The Duffy binding protein topilgan Plazmodiya, the one and only invasion ligand for DARC, does not bind to Saimiri erythrocytes although these cells express DARC and obviously become infected with P. vivax.[58]
- ^ If the frequency of the heterozygote is 0.40 the sickle-cell gene frequency (q) can be calculated from the Hardy-Weinberg equation 2q(1-q) = 0,40, whence q = 0.255 and q2, the frequency of sickle-cell homozygotes, is 0.065.
Lug'at
- actin, ankrin, spectrin – proteins that are the major components of the cytoskeleton scaffolding within a cell's cytoplasm
- aerob – uses oxygen for the production of energy (contrast anaerob)
- allel – one of two or more alternative forms of a gene that arise by mutation
- α-chain / β-chain (hemoglobin) – subcomponents of the hemoglobin molecule; two α-chains and two β-chains make up normal hemoglobin (HbA)
- alveolyar – pertaining to the alveoli, the tiny air sacs in the lungs
- aminokislota – any of twenty organic compounds that are subunits of protein in the human body
- anabolik – of or relating to the synthesis of complex molecules in living organisms from simpler ones
- together with the storage of energy; constructive metabolism (contrast katabolik)
- anaerob – refers to a process or reaction which does not require oxygen, but produces energy by other means (contrast aerob)
- anion transporter (organic) – molecules that play an essential role in the distribution and excretion of numerous endogenous metabolic products and exogenous organic anions
- antigen – any substance (as an immunogen or a hapten) foreign to the body that evokes an immune response either alone or after forming a complex with a larger molecule (as a protein) and that is capable of binding with a component (as an antibody or T cell) of the immune system
- ATP – (Adenozin trifosfat ) – an organic molecule containing high energy phosphate bonds used to transport energy within a cell
- katabolik – of or relatig to the breakdown of complex molecules in living organisms to form simpler ones, together with the release of energy; destructive metabolism (contrast anabolik)
- ximokin – are a family of small cytokines, or signaling proteins secreted by cells
- kodon – a sequence of three nucleotides which specify which amino acid will be added next during protein synthesis
- corpuscle – obsolete name for red blood cell
- cytoadherance – infected red blood cells may adhere to blood vellel walls and uninfected red blood cells
- sitoplazma – clear jelly-like substance, mostly water, inside a cell
- diatez – a tendency to suffer from a particular medical condition
- DNK – deoxyribonucleic acid, the hereditary material of the genome
- Drosophila – a kind of fruit fly used for genetic experimentation because of ease of reproduction and manipulation of its genome
- endotsitik – the transport of solid matter or liquid into a cell by means of a coated vacuole or vesicle
- endogamiya – the custom of marrying only within the limits of a local community, clan, or tribe
- endoteliy – of or referring to the thin inner surface of blood vessels
- ferment – a protein that promotes a cellular process, much like a catalyst in an ordinary chemical reaction
- epidemiologiya – the study of the spread of disease within a population
- eritrotsit – red blood cell, which with the leucocytes make up the cellular content of the blood (contrast leykotsit)
- eritroid – of or referring to erythrocytes, red blood cells
- fitness (genetic) – loosely, reproductive success that tends to propagate a trait or traits (see tabiiy selektsiya)
- genom – (abstractly) all the inheritable traits of an organism; represented by its chromosomes
- genotip – the genetic makeup of a cell, an organism, or an individual usually with reference to a specific trait
- glikoliz – the breakdown of glucose by enzymes, releasing energy
- glycophorin – transmembrane proteins of red blood cells
- haplotip – a set of DNA variations, or polymorphisms, that tend to be inherited together.
- Hb (HbC, HbE, HbS, etc.) hemoglobin (hemoglobin polymorphisms: hemoglobin type C, hemoglobin type E,
- hemoglobin type S)
- gemopoetik (stem cell) – the blood stem cells that give rise to all other blood cells
- heme oxygenase-1 (HO-1) – an enzyme that breaks down heme, the iron-containing non-protein part of hemoglobin
- gemoglobin – iron based organic molecule in red blood cells that transports oxygen and gives blood its red color
- gemoliz – the rupturing of red blood cells and the release of their contents (cytoplasm) into surrounding fluid (e.g., blood plasma)
- heterozigot – possessing only one copy of a gene for a particular trait
- bir jinsli – possessing two identical copies of a gene for a particular trait, one from each parent
- gipotonik – denotes a solution of lower osmotic pressure than another solution with which it is in contact, so that certain molecules will migrate from the region of higher osmotic pressure to the region of lower osmotic pressure, until the pressures are equalized
- in vitro – in a test tube or other laboratory vessel; usually used in regard to a testing protocol
- jonli ravishda – in a live human (or animal); usually used in regard to a testing protocol
- leykotsit – white blood cell, part of the immune system, which together with red blood cells, comprise the cellular component of the blood (contrast eritrotsit)
- ligand – an extracellular signal molecule, which when it binds to a cellular retseptorlari, causes a response by the cell
- lokus (gene or chromosome) – the specific location of a gene or DNA sequence or position on a chromosome
- makrofag – a large white blood cell, part of the immune system that ingests foreign particles and infectious microorganisms
- major histocompatibility complex (MHC) – proteins found on the surfaces of cells that help the immune system recognize foreign substances; also called the human leucocyte antigen (HLA) system
- mikro-RNK – a cellular RNA fragment that prevents the production of a particular protein by binding to and destroying the messenger RNA that would have produced the protein.
- mikrovasulyatsiya – very small blood vessels
- mitoxondriya – energy producing organelles of a cell
- mutatsiya – a spontaneous change to a gene, arising from an error in replication of DNA; usually mutations are referred to in the context of inherited mutations, i.e. changes to the gametes
- tabiiy selektsiya – the gradual process by which biological traits become either more or less common in a population as a function of the effect of inherited traits on the differential reproductive success of organisms interacting with their environment (closely related to fitness)
- nukleotid – organic molecules that are subunits, of nucleic acids like DNA and RNA
- nuklein kislota – a complex organic molecule present in living cells, esp. DNA or RNA, which consist of many nucleotides linked in a long chain.
- oxygen radical – a highly reactive ion containing oxygen, capable of damaging microorganisms and normal tissues.
- patogenez – the manner of development of a disease
- PCR – Polymerase Chain Reaction, an enzymatic reaction by which DNA is replicated in a test tube for subsequent testing or analysis
- fenotip – the composite of an organism's observable characteristics or traits, such as its morphology
- Plazmodium – the general type (genus) of the protozoan microorganisms that cause malaria, though only a few of them do
- polimerizatsiya – to combine replicated subunits into a longer molecule (usually referring to synthetic materials, but also organic molecules)
- polimorfizm – the occurrence of something in several different forms, as for example hemoglobin (HbA, HbC, etc.)
- polipeptid – a chain of amino acids forming part of a protein molecule
- receptor (cellular surface) – specialized integral membrane proteins that take part in communication between the cell and the outside world; receptors are responsive to specific ligands that attach to them.
- reducing environment (cellular) – reducing environment is one where oxidation is prevented by removal of oxygen and other oxidising gases or vapours, and which may contain actively reducing gases such as hydrogen, carbon monoxide and gases that would oxidize in the presence of oxygen, such as hydrogen sulfide.
- RNK – ribonucleic acid, a nucleic acid present in all living cells. Its principal role is to act as a messenger carrying instructions from DNA for controlling the synthesis of proteins
- sekvestratsiya (biology) – process by which an organism accumulates a compound or tissue (as red blood cells) from the environment
- jinsiy aloqada – a trait associated with a gene that is carried only by the male or female parent (contrast with autosomal)
- Sporozoa – a large class of strictly parasitic nonmotile protozoans, including Plazmodiya which cause malaria
- TCA tsikli – TriCarboxylic Acid cycle is a series of enzyme-catalyzed chemical reactions that form a key part of aerobic respiration in cells
- translokatsiya (cellular biology) – movement of molecules from outside to inside (or vice versa) of a cell
- transmembran – existing or occurring across a cell membrane
- venoz – of or referring to the veins
- pufakcha – a small organelle within a cell, consisting of fluid enclosed by a fatty membrane
- virulentlik omillari – enable an infectious agent to replicate and disseminate within a host in part by subverting or eluding host defenses.
Adabiyotlar
- ^ Kwiatkowski DP (August 2005). "Bezgak odam genomiga qanday ta'sir ko'rsatdi va inson genetikasi bezgak haqida bizga nimani o'rgatishi mumkin". Amerika inson genetikasi jurnali. 77 (2): 171–92. doi:10.1086/432519. PMC 1224522. PMID 16001361.
- ^ Hedrick PW (October 2011). "Population genetics of malaria resistance in humans". Irsiyat. 107 (4): 283–304. doi:10.1038/hdy.2011.16. PMC 3182497. PMID 21427751.
- ^ Anstee DJ (June 2010). "The relationship between blood groups and disease". Qon. 115 (23): 4635–43. doi:10.1182/blood-2010-01-261859. PMID 20308598.
- ^ Allison AC (1954). "Protection Afforded by Sickle-cell Trait Against Subtertian Malarial Infection" (PDF). Br Med J. 1 (4857): 290–294. doi:10.1136/bmj.1.4857.290. PMC 2093356. PMID 13115700. Arxivlandi asl nusxasi (PDF) 2011-09-28.
- ^ a b Miller LH, Mason SJ, Clyde DF, McGinniss MH (1976). "The resistance factor to Plasmodium vivax in blacks. The Duffy-blood-group genotype, FyFy". N Engl J Med. 295 (6): 302–4. doi:10.1056/NEJM197608052950602. PMID 778616.
- ^ Lemaitre B, Nicolas E, Michaut L, Reichhart JM, Hoffmann JA (1996). "Dorsoventral regulyativ gen kassetasi spätzle / Toll / kaktus Drosophila kattalaridagi qo'ziqorinlarga qarshi kuchli ta'sirni boshqaradi" (PDF). Hujayra. 86 (6): 973–983. doi:10.1016 / S0092-8674 (00) 80172-5. PMID 8808632. S2CID 10736743.
- ^ Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X, Birdwell D, Alejos E, Silva M, Galanos C, Freudenberg M, Ricciardi-Castagnoli P, Layton B, Beutler B (1998). "C3H / HeJ va C57BL / 10ScCr sichqonlaridagi nuqsonli LPS signalizatsiyasi: Tlr4 genidagi mutatsiyalar". Ilm-fan. 282 (5396): 2085–2088. Bibcode:1998 yil ... 282.2085P. doi:10.1126 / science.282.5396.2085. PMID 9851930.
- ^ Vaidya AB, Mather MW (2009). "Mitochondrial evolution and functions in malaria parasites". Annu Rev Microbiol. 63: 249–267. doi:10.1146/annurev.micro.091208.073424. PMID 19575561.
- ^ Elliott DA, McIntosh MT, Hosgood HD 3rd, Chen S, Zhang G, Baevova P, Joiner KA (2008). "Four distinct pathways of hemoglobin uptake in the malaria parasite Plasmodium falciparum". Proc. Natl. Akad. Ilmiy ish. AQSH. 105 (7): 2463–2468. Bibcode:2008PNAS..105.2463E. doi:10.1073/pnas.0711067105. PMC 2268159. PMID 18263733.
- ^ Kuross SA, Rank BH, Hebbel RP (1988). "Excess heme in sickle erythrocyte inside-out membranes: possible role in thiol oxidation" (PDF). Qon. 71 (4): 876–882. doi:10.1182/blood.V71.4.876.876. PMID 3355895.
- ^ Föller M, Bobbala D, Koka S, Huber SM, Gulbins E, Lang F (2009). "Suicide for survival--death of infected erythrocytes as a host mechanism to survive malaria". Hujayra fiziol biokimyosi. 24 (3–4): 133–140. doi:10.1159/000233238. PMID 19710527.
- ^ Cholera R, Brittain NJ, Gillrie MR, Lopera-Mesa TM, Diakité SA, Arie T, Krause MA, Guindo A, Tubman A, Fujioka H, Diallo DA, Doumbo OK, Ho M, Wellems TE, Fairhurst RM (2008). "Impaired cytoadherence of Plasmodium falciparum-infected erythrocytes containing sickle hemoglobin". Proc. Natl. Akad. Ilmiy ish. AQSH. 105 (3): 991–996. Bibcode:2008PNAS..105..991C. doi:10.1073/pnas.0711401105. PMC 2242681. PMID 18192399.
- ^ Ferreira A, Marguti I, Bechmann I, Jeney V, Chora A, Palha NR, Rebelo S, Henri A, Beuzard Y, Soares MP (2011). "Sickle hemoglobin confers tolerance to Plasmodium infection" (PDF). Hujayra. 145 (3): 398–409. doi:10.1016/j.cell.2011.03.049. PMID 21529713. S2CID 8567718. Arxivlandi asl nusxasi (PDF) 2011-10-03 kunlari.
- ^ Gong L, Parikh S, Rosenthal PJ, Greenhouse B (2013). "Biochemical and immunological mechanisms by which sickle cell trait protects against malaria" (PDF). Bezgak jurnali. 12 (1): 317. doi:10.1186/1475-2875-12-317. PMC 3847285. PMID 24025776.
- ^ Allison AC (2009). "Genetic control of resistance to human malaria". Immunologiyaning hozirgi fikri. 21 (5): 499–505. doi:10.1016/j.coi.2009.04.001. PMID 19442502.
- ^ Piel FB, Patil AP, Howes RE, Nyangiri OA, Gething PW, Williams TN, Weatherall DJ, Hay SI (2010). "Global distribution of the sickle cell gene and geographical confirmation of the malaria hypothesis". Nat Commun. 1 (8): 104. Bibcode:2010NatCo...1..104P. doi:10.1038/ncomms1104. PMC 3060623. PMID 21045822.
- ^ a b May J, Evans JA, Timmann C, Ehmen C, Busch W, Thye T, Agbenyega T, Horstmann RD (2007). "Hemoglobin variants and disease manifestations in severe falciparum malaria". JAMA. 297 (20): 2220–2226. doi:10.1001/jama.297.20.2220. PMID 17519411.
- ^ Flatz G (1967). "Hemoglobin E: distribution and population dynamics". Humangenetik. 3 (3): 189–234. doi:10.1007/BF00273124. PMID 6074385. S2CID 22541254.
- ^ a b Allison AC (1955). "Aspects of polymorphism in man". Sovuq bahor harbasi simptomi miqdori biol. 20: 239–251. doi:10.1101/SQB.1955.020.01.023. PMID 13433567.
- ^ Pauling L, Itano H, Singer SJ, Wells I (1949). "Sickle cell anemia, a molecular disease" (PDF). Ilm-fan. 110 (2865): 543–548. Bibcode:1949Sci...110..543P. doi:10.1126/science.110.2865.543. PMID 15395398.
- ^ Ingram VM (1959). "Abnormal human haemoglobins. III. The chemical difference between normal and sickle cell haemoglobins". Biochim Biofhys Acta. 36 (2): 543–548. doi:10.1016/0006-3002(59)90183-0. PMID 13852872.
- ^ Ingram VM (2004). "Sickle-Cell Anemia Hemoglobin: The Molecular Biology of the First "Molecular Disease"—The Crucial Importance of Serendipity". Genetika. 167 (1): 1–7. doi:10.1534 / genetika.167.1.1. PMC 1470873. PMID 15166132.
- ^ Cholera R, Brittain NJ, Gillrie MR, Lopera-Mesa TM, Diakité SA, Arie T, Krause MA, Guindo A, Tubman A, Fujioka H, Diallo DA, Doumbo OK, Ho M, Wellems TE, Fairhurst RM (2008). "Impaired cytoadherence of Plasmodium falciparum-infected erythrocytes containing sickle hemoglobin". Proc Natl Acad Sci AQSh. 105 (3): 991–996. Bibcode:2008PNAS..105..991C. doi:10.1073/pnas.0711401105. PMC 2242681. PMID 18192399.
- ^ Williams TN, Mwangi TW, Roberts DJ, Alexander ND, Weatherall DJ, Wambua S, et al. (2005 yil may). "An immune basis for malaria protection by the sickle cell trait". PLoS tibbiyoti. 2 (5): e128. doi:10.1371/journal.pmed.0020128. PMC 1140945. PMID 15916466.
- ^ Hebbel RP (2003). "Sickle hemoglobin instability: a mechanism for malarial protection". Redoks hisoboti. 8 (5): 238–40. doi:10.1179/135100003225002826. PMID 14962356. S2CID 28951874.
- ^ Kaul DK (2008). "Sickle red cell adhesion: many issues and some answers". Transfusion Clinique Et Biologique. 15 (1–2): 51–5. doi:10.1016/j.tracli.2008.03.012. PMID 18495516.
- ^ Brain P (1952). "Sickle-cell Anaemia in Africa". Br Med J. 2 (4789): 880. doi:10.1136/bmj.2.4789.880. PMC 2021738.
- ^ Elguero E, Délicat-Loembet LM, Rougeron V, Arnathau C, Roche B, Becquart P, et al. (2015). "Malaria continues to select for sickle cell trait in Central Africa". Proc Natl Acad Sci U S A. 112 (22): 7051–4. Bibcode:2015PNAS..112.7051E. doi:10.1073/pnas.1505665112. PMC 4460506. PMID 25941403.
- ^ Ingram VM, Stretton AO (1959). "Genetic basis of the thalassaemia diseases". Tabiat. 184 (4703): 1903–1909. Bibcode:1959Natur.184.1903I. doi:10.1038/1841903a0. PMID 13852871. S2CID 36535895.
- ^ Modiano G, Morpurgo G, Terrenato L, Novelletto A, Di Rienzo A, Colombo B, Purpura M, Mariani M, Santachiara-Benerecetti S, Brega A, Dixit KA, Shrestha SL, Lania A, Wanachiwanawin W, Luzzatto L (1991). "Protection Against Malaria Morbidity: Near Fixation of the α-Thalassemia gene in a Nepalese Population". Am. J. Xum. Genet. 48 (2): 390–397. PMC 1683029. PMID 1990845.
- ^ Terrenato L, Shrestha S, Dixit KA, Luzzatto L, Modiano G, Morpurgo G, Arese P (1988). "Nepalda simpatik populyatsiyalar bilan taqqoslaganda Tharu aholisida bezgak kasalligi kamaygan". Ann Trop Med parazitol. 82 (1): 1–11. doi:10.1080/00034983.1988.11812202. PMID 3041928.
- ^ Hutagalung R, Wilairatana P, Looareesuwan S, Brittenham GM, Aikawa M, Gordeuk VR (1999). "Influence of hemoglobin E trait on the severity of Falciparum malaria". J yuqtirgan disk. 179 (1): 283–286. doi:10.1086/314561. JSTOR 30117260. PMID 9841856.
- ^ Modiano D, Luoni G, Sirima BS, Simporé J, Verra F, Konate A, Rastrelli E, Olivieri A, Calissano C, Paganotti GM, D'Urbano L, Sanou I, Sawadogo A, Modiano G, Coluzzi M (2001). "Gemoglobin S klinik plazmodium falciparum bezgakdan himoya qiladi". Tabiat. 414 (6861): 305–308. Bibcode:2001 yil natur.414..305M. doi:10.1038/35104556. PMID 11713529. S2CID 4360808.
- ^ Modiano D, Bancone G, Ciminelli BM, Pompei F, Blot I, Simporé J, Modiano G (2008). "Haemoglobin S and haemoglobin C: 'quick but costly' versus 'slow but gratis' genetic adaptations to Plasmodium falciparum malaria". Hum Mol Genet. 17 (6): 789–799. doi:10.1093/hmg/ddm350. PMID 18048408.
- ^ Rihet P, Flori L, Tall F, Traore AS, Fumoux F (2004). "Hemoglobin C is associated with reduced Plasmodium falciparum parasitemia and low risk of mild attack" (PDF). Hum Mol Genet. 13 (1): 1–6. doi:10.1093/hmg/ddh002. PMID 14613965.
- ^ Fairhurst RM, Baruch DI, Brittain NJ, Ostera GR, Wallach JS, Hoang HL, Hayton K, Guindo A, Makobongo MO, Schwartz OM, Tounkara A, Doumbo OK, Diallo DA, Fujioka H, Ho M, Wellems TE (2005). "Abnormal display of PfEMP-1 on erythrocytes carrying haemoglobin C may protect against malaria" (PDF). Tabiat. 435 (7045): 1117–1121. Bibcode:2005Natur.435.1117F. doi:10.1038/nature03631. PMID 15973412. S2CID 4412263. Arxivlandi asl nusxasi (PDF) 2014-04-07 da. Olingan 2014-04-01.
- ^ Chotivanich K, Udomsangpetch R, Pattanapanyasat K, Chierakul W, Simpson J, Looareesuwan S, White N (August 2002). "Hemoglobin E: a balanced polymorphism protective against high parasitemias and thus severe P. falciparum malaria". Qon. 100 (4): 1172–6. doi:10.1182/blood.V100.4.1172.h81602001172_1172_1176. PMID 12149194.
- ^ Alving AS, Carson PE, Flanagan CL, Ickes CE (1956). "Enzymatic deficiency in primaquine-sensitive erythrocytes". Ilm-fan. 124 (3220): 484–485. Bibcode:1956Sci...124..484C. doi:10.1126/science.124.3220.484-a. PMID 13360274.
- ^ Cappellini MD, Fiorelli G (January 2008). "Glucose-6-phosphate dehydrogenase deficiency". Lanset. 371 (9606): 64–74. doi:10.1016/S0140-6736(08)60073-2. PMID 18177777. S2CID 29165746.
- ^ a b Tishkoff SA, Verelli BJ (2004). "G6PD deficiency and malarial resistance in humans: insights from evolutionary genetic analysis". In Dronamraju K (ed.). Evolutionary Aspects of Infectious Disease. Kembrij universiteti matbuoti.
- ^ Allison AC, Clyde DF (1961). "Malaria in African Children with Deficient Erythrocyte Glucose-6-phosphate Dehydrogenase". Br Med J. 1 (5236): 1346–1349. doi:10.1136/bmj.1.5236.1346. PMC 1954496. PMID 13682585.
- ^ Ruwende C, Khoo SC, Snow RW, Yates SN, Kwiatkowski D, Gupta S, Warn P, Allsopp CE, Gilbert SC, Peschu N, Newbold CI, Greenwood BM, Marsh K, Hill AV (1995). "Natural selection of hemi- and heterozygotes for G6PD deficiency in Africa by resistance to severe malaria". Tabiat. 376 (6537): 246–249. Bibcode:1995Natur.376..246R. doi:10.1038/376246a0. PMID 7617034. S2CID 4301352.
- ^ Luzzatto L (1979). "Genetics of red cells and susceptibility to malaria" (PDF). Qon. 54 (5): 961–976. doi:10.1182/blood.V54.5.961.961. PMID 387115.
- ^ Ayi K, Min-Oo G, Serghides L, Crockett M, Kirby-Allen M, Quirt I, Gros P, Kain KC (2008). "Pyruvate kinase deficiency and malaria". N Engl J Med. 358 (17): 1805–1810. doi:10.1056/NEJMoa072464. PMID 18420493.
- ^ Hempelmann E, Götze O (1984). "Polikromatik kumushni bo'yash orqali membrana oqsillarini xarakteristikasi". Hoppe-Seyler's Z Physiol Chem. 365: 241–242.
- ^ Jarolim P, Palek J, Amato D, Hassan K, Sapak P, Nurse GT, Rubin HL, Zhai S, Sahr KE, Liu SC (1991). "Deletion in erythrocyte band 3 gene in malaria-resistant Southeast Asian ovalocytosis". Proc. Natl. Akad. Ilmiy ish. AQSH. 88 (24): 11022–11026. Bibcode:1991PNAS...8811022J. doi:10.1073/pnas.88.24.11022. PMC 53065. PMID 1722314.
- ^ Liu SC, Zhai S, Palek J, Golan DE, Amato D, Hassan K, Nurse GT, Babona D, Coetzer T, Jarolim P, Zaik M, Borwein S (1990). "Molecular defect of the band 3 protein in southeast Asian ovalocytosis". N Engl J Med. 323 (22): 1530–1538. doi:10.1056/NEJM199011293232205. PMID 2146504.
- ^ Allen SJ, O'Donnell A, Alexander ND, Mgone CS, Peto TE, Clegg JB, Alpers MP, Weatherall DJ (1999). "Prevention of cerebral malaria in children in Papua New Guinea by southeast Asian ovalocytosis band 3". Am J Trop Med Hyg. 60 (6): 1056–1060. doi:10.4269/ajtmh.1999.60.1056. PMID 10403343.
- ^ Cortés A, Mellombo M, Mgone CS, Beck HP, Reeder JC, Cooke BM (2005). "Adhesion of Plasmodium falciparum-infected red blood cells to CD36 under flow is enhanced by the cerebral malaria-protective trait South-East Asian ovalocytosis". Mol biokimyosi parazitol. 142 (2): 252–257. doi:10.1016/j.molbiopara.2005.03.016. PMID 15978955.
- ^ a b Culleton RL, Mita T, Ndounga M, Unger H, Cravo PV, Paganotti GM, Takahashi N, Kaneko A, Eto H, Tinto H, Karema C, D'Alessandro U, do Rosário V, Kobayakawa T, Ntoumi F, Carter R, Tanabe K (2008). "Failure to detect Plasmodium vivax in West and Central Africa by PCR species typing". Malar J. 7 (1): 174–182. doi:10.1186/1475-2875-7-174. PMC 2546428. PMID 18783630.
- ^ Tournamille C, Colin Y, Cartron JP, Le Van Kim C (1995). "Disruption of a GATA motif in the Duffy gene promoter abolishes erythroid gene expression in Duffy-negative individuals". Nat Genet. 10 (2): 224–228. doi:10.1038/ng0695-224. PMID 7663520. S2CID 7125832.
- ^ Ryan JR, Stoute JA, Amon J, Dunton RF, Mtalib R, Koros J, Owour B, Luckhart S, Wirtz RA, Barnwell JW, Rosenberg R (2006). "Evidence for transmission of Plazmodium vivax among a duffy antigen negative population in Western Kenya" (PDF). Am J Trop Med Hyg. 75 (4): 575–581. doi:10.4269/ajtmh.2006.75.575. PMID 17038676.
- ^ Cavasini CE, de Mattos LC, Couto AA, Couto VS, Gollino Y, Moretti LJ, Bonini-Domingos CR, Rossit AR, Castilho L, Machado RL (2007). "Duffy blood group gene polymorphisms among malaria vivax patients in four areas of the Brazilian Amazon region". Malar J. 6 (1): 167. doi:10.1186/1475-2875-6-167. PMC 2244634. PMID 18093292.
- ^ Ménard D, Barnadas C, Bouchier C, Henry-Halldin C, Gray LR, Ratsimbasoa A, Thonier V, Carod JF, Domarle O, Colin Y, Bertrand O, Picot J, King CL, Grimberg BT, Mercereau-Puijalon O, Zimmerman PA (2010). "Plasmodium vivax clinical malaria is commonly observed in Duffy-negative Malagasy people". Proc. Natl. Akad. Ilmiy ish. AQSH. 107 (13): 5967–71. Bibcode:2010PNAS..107.5967M. doi:10.1073/pnas.0912496107. PMC 2851935. PMID 20231434.
- ^ Pierron D, Heiske M, Razafindrazaka H, Pereda-Loth V, Sanchez J, Alva O, et al. (Mart 2018). "Strong selection during the last millennium for African ancestry in the admixed population of Madagascar". Tabiat aloqalari. 9 (1): 932. doi:10.1038/s41467-018-03342-5. PMC 5834599. PMID 29500350.
- ^ Mendes C, Dias F, Figueiredo J, Mora VG, Cano J, de Sousa B, do Rosário VE, Benito A, Berzosa P, Arez AP (2011). Franco-Paredes C (ed.). "Duffy Negative Antigen Is No Longer a Barrier to Plasmodium vivax – Molecular Evidences from the African West Coast (Angola and Equatorial Guinea)". PLOS Negl Trop Dis. 5 (e1192): e1192. doi:10.1371/journal.pntd.0001192. PMC 3119644. PMID 21713024.
- ^ Barnwell JW, Nichols ME, Rubinstein P (1989). "In vitro evaluation of the role of the Duffy blood group in erythrocyte invasion by Plasmodium vivax". J Exp Med. 169 (5): 1795–802. doi:10.1084/jem.169.5.1795. PMC 2189319. PMID 2469769.
- ^ Wertheimer SP, Barnwell JW (1989). "Plazmodium vivax interaction with the human Duffy blood group glycoprotein: identification of a parasite receptor-like protein". Exp Parazitol. 69 (4): 340–350. doi:10.1016/0014-4894(89)90083-0. PMID 2680568.
- ^ a b Hill AV, Allsopp CE, Kwiatkowski D, Anstey NM, Twumasi P, Rowe PA, Bennett S, Brewster D, McMichael AJ, Greenwood BM (1991). "Umumiy g'arbiy Afrika HLA antigenlari og'ir bezgakdan himoya bilan bog'liq". Tabiat. 352 (6336): 595–600. Bibcode:1991 yil Natur.352..595H. doi:10.1038 / 352595a0. PMID 1865923. S2CID 2667496.
- ^ Frodsham AJ, Hill AV (2004). "Genetics of infectious diseases". Hum Mol Genet. 13 Spec No 2: R187–R194. doi:10.1093/hmg/ddh225. PMID 15358724.
- ^ Billig EM, McQueen PG, McKenzie FE (2012). "Foetal haemoglobin and the dynamics of paediatric malaria". Bezgak jurnali. 11: 396. doi:10.1186/1475-2875-11-396. PMC 3538578. PMID 23190739.
- ^ Brenda AkinyiI Webala, "Prevalence of Fetal Hemoglobin and Antibody Responses to Plazmodium falciparum Antigens in Sickle Cell Disease Patients in Western Kenya." Master's thesis, School of Pure and Applied Sciences of Kenyatta University, 2013.
- ^ Bengtsson BO, Tunlid A (July 2010). "The 1948 international congress of genetics in Sweden: people and politics". Genetika. 185 (3): 709–15. doi:10.1534/genetics.110.119305. PMC 2907196. PMID 20660651.
- ^ Haldane, J. B. S. (1949). "The rate of mutation of human genes". Hereditalar. 35 (S1): 267–273. doi:10.1111/j.1601-5223.1949.tb03339.x.
- ^ Lederberg J (September 1999). "J. B. S. Haldane (1949) on infectious disease and evolution". Genetika. 153 (1): 1–3. PMC 1460735. PMID 10471694.
- ^ a b v Aidoo M, Terlouw DJ, Kolczak MS, McElroy PD, ter Kuile FO, Kariuki S, Nahlen BL, Lal AA, Udhayakumar V (2002). "Protective effects of the sickle cell gene against malaria morbidity and mortality" (PDF). Lanset. 359 (9314): 1311–1312. doi:10.1016/S0140-6736(02)08273-9. PMID 11965279. S2CID 37952036. Arxivlandi asl nusxasi (PDF) 2010-06-09 da.
- ^ Jallow M, Teo YY, Small KS, Rockett KA, et al. (2009). "G'arbiy Afrikada bezgakni genom bo'yicha va aniqlik bilan assotsiatsiyasini tahlil qilish". Nat Genet. 41 (6): 657–665. doi:10.1038 / ng.388. PMC 2889040. PMID 19465909.
- ^ Allison AC (1954). "Notes on sickle-cell polymorphism". Inson genetikasi yilnomalari. 19 (1): 39–57. doi:10.1111/j.1469-1809.1954.tb01261.x. PMID 13208024. S2CID 10056569.[o'lik havola ]
- ^ Vandepitte JM, Zuelzer WW, Neel JV, Colaert J (1955). "Evidence concerning the inadequacy of mutation as an explanation of the frequency of the sickle cell gene in the Belgian Congo". Qon. 10 (4): 341–350. doi:10.1182/blood.V10.4.341.341. PMID 14363315.
Qo'shimcha o'qish
- Dronamraju KR, Arese P (2006) Malaria: Genetic and Evolutionary Aspects, Springer; Berlin, ISBN 0-387-28294-7 / ISBN 978-0-387-28294-7
- Faye FBK (2009) Malaria Resistance or Susceptibility in Red Cells Disorders, Nova Science Publishers Inc, New York. ISBN 9781606929438