Katexin - Catechin
 | |
| Ismlar | |
|---|---|
| IUPAC nomi (2R,3S) -2- (3,4-Dihidroksifenil) -3,4-dihidro-2H-xromen-3,5,7-triol | |
| Boshqa ismlar Sianidanol Siyanidanol (+) - katexin D-katexin Katexin kislotasi Katexuik kislota Sianidol Dekssiyanidanol (2R,3S) -Katexin 2,3-trans-Katexin (2R,3S) -Flavan-3,3 ′, 4 ′, 5,7-pentol | |
| Identifikatorlar | |
| |
3D model (JSmol ) | |
| 3DMet | |
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| ECHA ma'lumot kartasi | 100.005.297 |
| EC raqami |
|
| KEGG | |
PubChem CID | |
| UNII | |
CompTox boshqaruv paneli (EPA) | |
| |
| |
| Xususiyatlari | |
| C15H14O6 | |
| Molyar massa | 290.271 g · mol−1 |
| Tashqi ko'rinish | Rangsiz qattiq |
| Erish nuqtasi | 175 dan 177 ° C gacha (347 dan 351 ° F; 448 dan 450 K gacha) |
| UV-vis (λmaksimal) | 276 nm |
Chiralning aylanishi ([a]D.) | +14.0° |
| Xavf | |
| Asosiy xavf | Sutemizuvchilarning somatik hujayralari uchun mutagen, bakteriyalar va xamirturushlar uchun mutagen |
| Xavfsizlik ma'lumotlari varaqasi | ilmiy kitob AppliChem[doimiy o'lik havola ] |
| GHS piktogrammalari |  |
| GHS signal so'zi | Ogohlantirish |
| H315, H319, H335 | |
| P261, P264, P271, P280, P302 + 352, P304 + 340, P305 + 351 + 338, P312, P321, P332 + 313, P337 + 313, P362, P403 + 233, P405, P501 | |
| O'lim dozasi yoki konsentratsiyasi (LD, LC): | |
LD50 (o'rtacha doz ) | (+) - katexin: kalamushda 10000 mg / kg (RTECS) Sichqonchada 10000 mg / kg Sichqoncha bilan 3,890 mg / kg (boshqa manba) |
| Farmakologiya | |
| Og'zaki | |
| Farmakokinetikasi: | |
| Siydik | |
Boshqacha ko'rsatilmagan hollar bundan mustasno, ulardagi materiallar uchun ma'lumotlar keltirilgan standart holat (25 ° C [77 ° F], 100 kPa da). | |
| Infobox ma'lumotnomalari | |
Katexin /ˈkætɪtʃɪn/ a flavan-3-ol, tabiiy fenolning bir turi va antioksidant. Bu o'simlik ikkilamchi metabolit. Bu guruhga kiradi flavan-3-ols (yoki oddiygina flavanollar), kimyoviy oilasining bir qismi flavonoidlar.
Katexin kimyoviy oilasining nomi kelib chiqadi katexu, bu tanin sharbati yoki qaynatilgan ekstrakti Mimoza katexi (Acacia catechu L.f).[1]
Kimyo
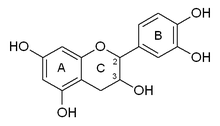
Katexin ikkita narsaga ega benzol uzuklari (A va B halqalari deb ataladi) va a dihidropiran a bilan heterosikl (C halqasi) gidroksil guruhi uglerodda 3. A halqa a ga o'xshaydi rezortsinol qism, B halqasi a ga o'xshash katexol qism. Ikki bor chiral molekulada markazlari 2 va 3 ga teng. Shuning uchun uning to'rttasi bor diastereoizomerlar. Izomerlardan ikkitasi trans konfiguratsiyasi va deyiladi katexin va qolgan ikkitasi cis konfiguratsiyasi va deyiladi epikatexin.
Katexinning eng keng tarqalgan izomeri (+) - katexindir. Boshqa stereoizomer (-) - katexin yoki ent- katexin. Eng keng tarqalgan epikatexin izomeri (-) - epikatechin (shuningdek, L-epikatechin, epikatexol, (-) - epikatexol, l-akatsatechin, l-epikatexol, epi-katexin, 2,3-sis-epikatexin yoki ( 2018-04-02 121 2R,3R) - (-) - epikatexin).
Turli epimerlardan foydalanib ajratish mumkin chiral ustunli kromatografiya.[2]
Hech qanday izomerga murojaat qilmasdan, molekulani faqat katexin deb atash mumkin. Turli enantiomerlarning aralashmalarini (+/-) - katexin yoki DL-katexin va (+/-) - epikatechin yoki DL-epikatexin deb atash mumkin.
Katexin va epikatexin - bu qurilish bloklari proantosiyanidinlar, quyultirilgan tanin turi.
- Diastereoizomerlar galereyasi

(+) - katexin (2R,3S)

(-) - katexin (2S,3R)

(-) - epikatexin (2R,3R)
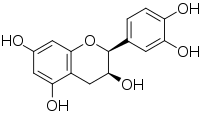
(+) - epikatexin (2S,3S)





Bundan tashqari, C halqasining egiluvchanligi ikkitasini beradi konformatsiya izomerlari, B halqasini psevdoekvatorial holatga (E konformator) yoki psevdoaksial holatga (A konformer) qo'yish. Tadqiqotlar shuni tasdiqladiki, (+) - katexin suvli eritmada A- va E-konformerlar aralashmasini qabul qiladi va ularning konformatsion muvozanati 33:67 ga teng.[3]
Flavonoidlar sifatida katexinlar rol o'ynashi mumkin antioksidantlar yuqori konsentratsiyali bo'lganda in vitro, ammo boshqa flavonoidlar bilan taqqoslaganda ularning antioksidant salohiyati past.[4] Singlet kislorodni so'ndirish qobiliyati katexinning kimyoviy tuzilishi bilan bog'liq bo'lib, B halqasida katekol qismi va S halqasida er-xotin bog'lanishni faollashtiradigan gidroksil guruhi mavjud.[5]
Oksidlanish
Elektrokimyoviy tajribalar shuni ko'rsatadiki (+) - katexin oksidlanish mexanizmi ketma-ket bosqichlarda, katexol va rezortsinol guruhlari va oksidlanish pHga bog'liq. Katexol 3 ', 4'-dihidroksil elektron donorlik guruhlarining oksidlanishi birinchi navbatda, juda past ijobiy potentsiallarda yuzaga keladi va qaytariladigan reaksiya hisoblanadi. Keyinchalik oksidlangan rezortsinol qismining gidroksil guruhlari qaytarilmas oksidlanish reaktsiyasiga kirishganligi ko'rsatilgan.[6]
The laccase /ABTS sistema (+) - katexinni oligomerik mahsulotlarga oksidlaydi[7] ulardan proantosiyanidin A2 dimer.
Spektral ma'lumotlar

| UV-Vis | |
|---|---|
| Lambda-max: | 276 nm |
| Yo'qolib ketish koeffitsienti (log ε) | 4.01 |
| IQ | |
| Asosiy assimilyatsiya bantlari | 1600 sm−1(benzol halqalari) |
| NMR | |
| Proton NMR
| δ : 2.49 (1H, dd, J = 16.0, 8.6 Hz, H-4a), |
| Uglerod-13 NMR | |
| Boshqa NMR ma'lumotlari | |
| XONIM | |
| Massalari asosiy qismlar | ESI-MS [M + H] + m / z: 291.0
|
Tabiiy hodisalar
(+) - katexin va (-) - epikatexin, shuningdek ularnikidan gal kislotasi konjugatlar hamma joyda mavjud bo'lgan tarkibiy qismlardir qon tomir o'simliklar va tez-tez uchraydigan tarkibiy qismlar an'anaviy o'simliklarni davolash vositalari, kabi Uncaria rhynhophylla. Ikki izomerlar asosan sifatida topilgan kakao va choy tarkibiy qismlar, shuningdek Vitis vinifera uzum.[9][10][11]
Oziq-ovqatda
Evropada katexinlarning asosiy parhez manbalari va Qo'shma Shtatlar bor choy va pome mevalar.[12][13]
Katexinlar va epikatexinlar topilgan kakao,[14] ma'lumotlar bazasiga ko'ra, tahlil qilingan ovqatlar orasida eng ko'p katexinlar (108 mg / 100 g) bor, undan keyin o'rik sharbati (25 mg / 100 ml) va keng fasol pod (16 mg / 100 g).[15] Aça neft, açaí xurmo mevasidan olingan (Evterpe oleracea), tarkibida (+) - katexinlar (67 mg / kg).[16]
Katexinlar ovqatlar orasida xilma-xildir,[15] dan shaftoli[17] ga yashil choy va sirka.[15][18] Katexinlar topilgan arpa ular uchun mas'ul bo'lgan asosiy fenolik birikma bo'lgan don xamir rang o'zgarishi.[19] Monomerik (+) - katexin yoki (-) - epikatexin bilan bog'liq bo'lgan ta'm biroz tavsiflanadi biriktiruvchi, lekin achchiq emas.[20]
Metabolizm
Biosintez
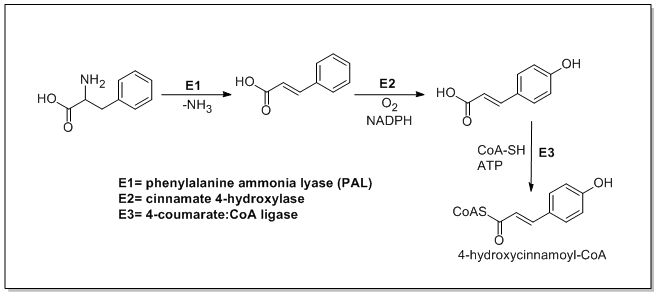
Katexin biosintezi ma bilan boshlanadi 4-gidroksitsinnamoyl CoA uchta qo'shilgan holda zanjir uzaytirilishidan boshlanadigan blok malonil-CoA PKSIII yo'lidan. 4-gidroksitsinnamoyl CoA dan biosintez qilinadi L-fenilalanin Shikimate yo'li orqali. L-fenilalanin dastlab zararsizlantiriladi fenilalanin ammiak liaz (PAL) doljin kislotasini hosil qiladi va keyinchalik u oksidlanadi 4-gidroksitsinnam kislotasi 4-gidroksilaza cinnamati bilan. Keyinchalik xalkon sintaz 4-gidroksitsinnamoil CoA va uchta malonil-KoA molekulalarining kondensatsiyasini katalizlaydi xalkon. Keyinchalik xalkon izomerizatsiya qilinadi naringenin oksidlangan xalkon izomeraza bilan erioditsitol flavonoid 3'- gidroksilaza tomonidan va keyinchalik oksidlanadi taksifolin flavanon 3-gidroksilaza tomonidan. Keyin taksifolin dihidroflavanol 4-reduktaza va tomonidan kamaytiriladi leukoantosiyanidin reduktaza katexin hosil qilish. Katexin biosintezi quyida keltirilgan[21][22][23]
Leykosiyanidin reduktaza (LCR) 2,3-trans-3,4-cis- dan foydalanadileukosiyanidin ishlab chiqarish uchun (+) - katexin va tarkibidagi birinchi ferment hisoblanadi proantosiyanidinlar (PA) - maxsus yo'l. Uning faoliyati barglar, gullar va dukkakli o'simliklarning urug'lari bilan o'lchandi Medicago sativa, Lotus japonicus, Lotus uliginosus, Hedysarum sulfuresens va Robiniya pseudoacacia.[24] Ferment ham mavjud Vitis vinifera (uzum).[25]

Biologik parchalanish
Katexin parchalanishining asosiy fermenti bo'lgan katexin oksigenaz zamburug'lar va bakteriyalarda mavjud.[26]
Bakteriyalar orasida (+) - katexin parchalanishiga erishish mumkin Acinetobacter calcoaceticus. Katexin metabolizmga uchraydi protatexuik kislota (PCA) va floroglyucinol karboksilik kislota (PGCA).[27] Shuningdek, bu degradatsiyaga uchragan Bradyrhizobium japonicum. Phloroglucinol karboksilik kislota bundan keyin dekarboksillangan ga floroglyucinol, bu degidroksillangan ga rezortsinol. Resorsinol gidroksillanadi gidroksikinol. Protokatexuik kislota va gidroksikinol ta'sir qiladi intradiol dekolte orqali 3,4-dioksigenazni protokatlashtiring va gidroksikinol 1,2-dioksigenaza shakllantirmoq b-karboksi sis, sis-mukon kislotasi va maleyl asetat.[28]
Qo'ziqorinlar orasida katexinning parchalanishiga erishish mumkin Chaetomium cupreum.[29]
Odamlarda metabolizm


Katexinlar qabul qilinganidan keyin metabolizmga uchraydi oshqozon-ichak trakti, xususan jejunum,[31] va jigar, natijada strukturaviy bog'liq epikatexin metabolitlari (SREM) paydo bo'ladi.[32] SREMlar uchun asosiy metabolik yo'llar glyukuronidatsiya, sulfatlanish va metilatsiya ning katexol guruh tomonidan katekol-O-metil transferaza, plazmada faqat oz miqdordagi moddalar aniqlangan.[33][30] Ammo parhezli katexinlarning aksariyati metabolizmga uchraydi yo'g'on ichak mikrobiomi ga gamma-valerolaktonlar va gipur kislotalari bundan keyin o'tadigan biotransformatsiya, glyukuronidatsiya, sulfatlanish va metilatsiya ichida jigar.[33]
Katexinlarning stereokimyoviy konfiguratsiyasi ularning qabul qilinishi va metabolizmiga kuchli ta'sir qiladi, chunki qabul qilish (-) - epikatezin uchun eng yuqori va (-) - katexin uchun eng past ko'rsatkichdir.[34]
Tadqiqot

Nanozarralar katexinlarni etkazib berish tizimlari sifatida dastlabki tadqiqotlar olib borilmoqda.[35] Kakao katexinlari xavfga ta'sir ko'rsatishi uchun dastlabki tadqiqotlar olib borilmoqda yurak-qon tomir kasalliklari.[36] Bir cheklangan meta-tahlil shuni ko'rsatdiki, yashil choy va uning katexinlari iste'molini kuniga etti stakangacha oshirish, ozgina pasayishni ta'minladi. prostata saratoni.[37]
Biotransformatsiya
(+) - katexinning biotransformatsiyasi taksifolin ikki bosqichli oksidlanish orqali erishish mumkin Burxolderiya sp.[38]
(+) - Katexin va (-) - epikatexin endofitik filamentli qo'ziqorin bilan o'zgaradi Diaport sp. 3,4-sis-dihidroksiflavan hosilalariga, (+) -(2R, 3S, 4S) -3,4,5,7,3 ', 4'-geksahidroksiflavan (leukosiyanidin) va (-) - (2R, 3R, 4R) -3,4,5,7,3 ', 4'-heksahidroksiflavan navbati bilan, (-) - katexin va (+) - 2S-fenil guruhi bo'lgan epikatexin biooksidlanishga qarshilik ko'rsatdi.[39]
Leykoantosiyanidin reduktaza (LAR) foydalanadi (2R,3S) -cateechin, NADP+ va H22,3-trans-3,4-cis- ishlab chiqarish uchun Oleukosiyanidin, NADPH va H+. Uning gen ekspressioni uzum mevalari va uzum barglarini rivojlantirishda o'rganilgan.[40]
Glikozidlar
- (2R, 3S) -Katexin-7-O-b-D-glyukopiranozid dan ajratilishi mumkin arpa (Hordeum vulgare L.) va solod.[41]
- Epigeosid (Katexin-3-O-alfa-L-ramnopiranosil- (1-4) -beta-D-glyukopiranosil- (1-6) -beta-D-glyukopiranozid) ning rizomlaridan ajratilishi mumkin. Epigynum auritum.[42]
Bioaktivlikni o'rganish
Qon tomirlari funktsiyasi

Bir necha asrlar ilgari katexin o'z ichiga olgan ekstraktlar yurak kasalliklarini davolash uchun foydali deb hisoblangan,[44][45] va kapillyarlarning o'tkazuvchanligiga ta'siri 1936 yilda ko'rsatilgan.[46] Parhez tadqiqotlaridagi cheklangan dalillar katexinlarning ta'sir qilishi mumkinligini ko'rsatadi endoteliy - mustaqil vazodilatatsiya normal holatga hissa qo'shishi mumkin qon oqimi odamlarda tartibga solish.[47][48] Yashil choy katexinlari qon bosimini yaxshilashi mumkin, ayniqsa sistolik qon bosimi 130 mmHg dan yuqori bo'lganda.[49] Ovqat hazm qilish jarayonida keng metabolizm tufayli, qon tomirlariga bu ta'sir uchun javobgar bo'lgan katechin metabolitlarining taqdiri va faoliyati, shuningdek, haqiqiy ta'sir usuli noma'lum.[33][50]
The Evropa oziq-ovqat xavfsizligi boshqarmasi kakao flavanollari sog'lom kattalardagi qon tomirlarining ishiga ta'sir qiladi degan xulosaga kelishgan: "kakao flavanollari normal qon oqimiga hissa qo'shadigan endoteliyga bog'liq vazodilatatsiyani saqlashga yordam beradi".[51] Kuzatuv kohort tadqiqotlari ma'lumotlari flavan-3-olni iste'mol qilish va yurak-qon tomir kasalliklari xavfi o'rtasidagi izchil bog'liqlikni ko'rsatmadi.[43]
A meta-tahlil shuningdek, yashil choy katexinlari ijobiy ta'sir ko'rsatishi mumkinligini ko'rsatdi xolesterin.[49]
Immunitet ta'sirlari mumkin
Iste'mol qilinadigan dozaga qarab, katexinlar va ularning metabolitlari qizil qon tanachalari bilan bog'lanib, ularning ajralishini keltirib chiqarishi mumkin otoantikorlar, ni natijasida gemolitik anemiya va buyrak etishmovchiligi.[52] Bu davolash uchun ishlatiladigan katexin o'z ichiga olgan Catergen preparatini olib tashlashga olib keldi virusli gepatit, 1985 yilda Evropa bozoridan.[53][54]
Botanika ta'siri
Ba'zi o'simliklar tomonidan erga tashlanadigan katexinlar qo'shnilarining o'sishiga to'sqinlik qilishi mumkin allelopatiya.[55] Centaurea maculosa, ko'pincha bu xatti-harakatlar uchun o'rganilgan dog'langan knapweed, katexinni chiqaradi izomerlar ildizlari orqali erga, potentsial sifatida ta'sir qilishi mumkin antibiotik yoki gerbitsid. Gipotezalardan biri shundaki, u sabab bo'ladi reaktiv kislorod turlari ildiz hujayralarini o'ldirish uchun maqsadli o'simlik ildizi orqali to'lqin apoptoz.[56] Evropa ekotizimidagi aksariyat o'simliklar katexinga qarshi himoyaga ega, ammo Shimoliy Amerika ekotizimida ozgina o'simliklar unga qarshi himoyalangan. Centaurea maculosa invaziv, nazoratsiz o't hisoblanadi.[55]
Katexin qulupnay barglarida infektsiyani inhibe qiluvchi omil sifatida ishlaydi.[57] Epikatexin va katexin kofe berry kasalligini oldini olish orqali oldini oladi appressorial melanizatsiya Colletotrichum kahawae.[58]
Adabiyotlar
- ^ "Cutch va catechu o'simliklarining kelib chiqishi". Birlashgan Millatlar Tashkilotining Oziq-ovqat va qishloq xo'jaligi tashkiloti. 2011 yil 5-noyabr.
- ^ Rinaldo D, Batista JM, Rodrigues J va boshq. (Avgust 2010). "Kiral HPLC-PAD-CD yordamida Byrsonima turlarining barglaridan katexin diastereomerlarini aniqlash". Chirallik. 22 (8): 726–33. doi:10.1002 / chir.20824. PMID 20143413.
- ^ Kríz Z, Koca J, Imberty A, Charlot A, Auzély-Velty R (iyul 2003). "(+) - katexinni b-siklodekstrin bilan NMR, mikrokalorimetriya va molekulyar modellashtirish texnikasi kombinatsiyasini o'rganish". Org. Biomol. Kimyoviy. 1 (14): 2590–5. doi:10.1039 / B302935M. PMID 12956082.
- ^ Pietta, P. G. (2000). "Flavonoidlar antioksidant sifatida". Tabiiy mahsulotlar jurnali. 63 (7): 1035–42. doi:10.1021 / np9904509. PMID 10924197.
- ^ Tournaire C, Croux S, Maurette MT va boshq. (1993 yil avgust). "Flavonoidlarning antioksidant faolligi: singlet kislorodning samaradorligi (1Δgsöndürme ". J. Fotokem. Fotobiol. B, Biol. 19 (3): 205–15. doi:10.1016/1011-1344(93)87086-3. PMID 8229463.
- ^ Janeyro, Patrisiya; Oliveira Bret, Ana Mariya (2004). "Katexinning elektrokimyoviy oksidlanish mexanizmlari". Analytica Chimica Acta. 518 (1–2): 109–115. doi:10.1016 / j.aca.2004.05.038. hdl:10316/5128.
- ^ Usmon, A.M.; Vong, K.K.Y .; Fernyhough, A. (2007). "Lakart / ABTS tizimi (+) - katexinni oligomerik mahsulotlarga oksidlaydi". Ferment va mikroblar texnologiyasi. 40 (5): 1272–1279. doi:10.1016 / j.enzmictec.2006.09.018.
- ^ Lin, Yi-Pei; Chen, Tai-Yuan; Tseng, Tszyan-Ven; Li, Mey-Syen; Chen, Shui-Tein (2009). "Feniks hanceana var. Formosana dan ajratilgan neyron hujayralarni himoya qiluvchi birikmalari". Fitokimyo. 70 (9): 1173–81. doi:10.1016 / j.hytochem.2009.06.006. PMID 19628235.
- ^ Aispurua-Olaizola, Oier; Ormazabal, Markel; Vallexo, Asier; Olivares, Maytan; Patrisiya, Navarro; Etxebarriya, Nestor; Usobiaga, Aresatz (2015-01-01). "Vitis Vinifera uzum chiqindilaridan yog 'kislotalari va polifenollarni ketma-ket ekstraktsiyasini superkritik suyuqligini optimallashtirish". Oziq-ovqat fanlari jurnali. 80 (1): E101-E107. doi:10.1111/1750-3841.12715. ISSN 1750-3841. PMID 25471637.
- ^ Freydenberg, Karl; Koks, Richard F. B.; Braun, Emil (1932). "Kakao loviyasining katexini1". Amerika Kimyo Jamiyati jurnali. 54 (5): 1913–1917. doi:10.1021 / ja01344a026.
- ^ "Michiyo Tsujimura (1888–1969)". Olingan 10-noyabr 2015.
- ^ Chun, O. K .; Chung, S. J .; Song, W. O. (2007). "Flavonoidlarni taxminiy iste'mol qilish va AQSh kattalarining asosiy oziq-ovqat manbalari". Oziqlanish jurnali. 137 (5): 1244–52. doi:10.1093 / jn / 137.5.1244. PMID 17449588.
- ^ Vogiatzoglou, A; Mulligan, A. A .; Lentjes, M. A .; Lyuben, R. N .; Spenser, J. P .; Shroeter, H; Xav, K. T .; Kuhl, G. G. (2015). "Evropa kattalaridagi flavonoidlarni iste'mol qilish (18 yoshdan 64 yoshgacha)". PLOS ONE. 10 (5): e0128132. doi:10.1371 / journal.pone.0128132. PMC 4444122. PMID 26010916.
- ^ Kwik-Uribe C, Bektash RM (2008). "Kakao flavanollari - o'lchov, bioavailability va bioaktivlik" (PDF). Asia Pac J Clin Nutr. 17 (Qo'shimcha 1): 280-3. PMID 18296356.
- ^ a b v "Yashil choy damlamasidagi polifenollar". Fenol-Explorer, v 3.5. 2014 yil. Olingan 1 noyabr 2014.
- ^ Pacheco-Palencia LA, Mertens-Talcott S, Talcott ST (iyun 2008). "Acai (Euterpe oleracea Mart.) Dan olingan fitokimyoviy boyitilgan yog'ning kimyoviy tarkibi, antioksidant xususiyatlari va issiqlik barqarorligi". J. Agric. Oziq-ovqat kimyosi. 56 (12): 4631–6. doi:10.1021 / jf800161u. PMID 18522407.
- ^ Cheng, Givven V.; Krisosto, Karlos H. (1995). "Shaftoli va nektarin teri to'qimalarining tampon ekstraktlarining qorayish potentsiali, fenolik tarkibi va polifenoloksidaza faolligi" (PDF). J. Am. Soc. Hort. Ilmiy ish. 120 (5): 835–838. doi:10.21273 / JASHS.120.5.835. Arxivlandi asl nusxasi (PDF) 2014-05-14. Olingan 2012-02-26.
- ^ Galvez, Migel Karrero; Barroso, Karmelo Garsiya; Peres-Bustamante, Xuan Antonio (1994). "Turli sirka namunalarining polifenolik birikmalarini tahlil qilish". Zeitschrift für Lebensmittel-Untersuchung und -Forschung. 199 (1): 29–31. doi:10.1007 / BF01192948.
- ^ Kvinda-Axtell, Zori; Baik, Byung-Kee (2006). "Arpa donasining fenolik birikmalari va ularning oziq-ovqat mahsulotlarining rangsizlanishiga ta'siri". J. Agric. Oziq-ovqat kimyosi. 54 (26): 9978–9984. doi:10.1021 / jf060974w. PMID 17177530.
- ^ Kielxorn, S; Thorngate Iii, JH (1999). "Flavan-3-ols (+) - katexin va (-) - epikatexin bilan bog'liq og'zaki hislar". Oziq-ovqat sifati va afzalligi. 10 (2): 109–116. doi:10.1016 / S0950-3293 (98) 00049-4.
- ^ Rani, Arti; Singh, Kashmir; Ahuja, Paramvir S.; Kumar, Sanjay (2012). "Choy tarkibidagi katexinlar biosintezining molekulyar regulyatsiyasi [Camellia sinensis (L.) O. Kuntze] "deb nomlangan. Gen. 495 (2): 205–10. doi:10.1016 / j.gene.2011.12.029. PMID 22226811.
- ^ Punyasiri, P.A.N.; Abeysinghe, I. S. B.; Kumar, V .; Treutter, D .; Duy, D .; Gosch, S .; Martens, S .; Forkmann, G.; Fischer, T. C. (2004). "Camellia sinensis choy o'simlikidagi flavonoid biosintezi: taniqli epikatexin va katexin yo'llari fermentlarining xususiyatlari". Biokimyo va biofizika arxivlari. 431 (1): 22–30. doi:10.1016 / j.abb.2004.08.003. PMID 15464723.
- ^ Devik, Pol M. (2009). Dorivor tabiiy mahsulotlar: biosintetik yondashuv (3-nashr). Buyuk Britaniya: John Wiley & Sons. ISBN 978-0-470-74167-2.[sahifa kerak ]
- ^ Skadhauge, Birgitte; Gruber, Margaret Y.; Tomsen, Karl Kristian; Von Vettstein, Diter (1997 yil aprel). "Leykosiyanidin reduktaza faolligi va dukkakli to'qimalarni rivojlanishida proantotsianidinlarning to'planishi". Amerika botanika jurnali. 84 (4): 494–503. doi:10.2307/2446026. JSTOR 2446026.
- ^ Maugé C, Granier T, d'Estaintot BL va boshq. (2010 yil aprel). "Vitis vinifera dan leykoantosiyanidin reduktaza kristalli tuzilishi va katalitik mexanizmi". J. Mol. Biol. 397 (4): 1079–91. doi:10.1016 / j.jmb.2010.02.002. PMID 20138891.
- ^ Katexinning biologik parchalanishi. M Arunachalam, M Mohan Raj, N Mohan va A Mahadevan, Proc. Hind natn Sci Acad. B69 № 4 353-370 betlar (2003) Arxivlandi 2012-03-16 da Orqaga qaytish mashinasi
- ^ Arunachalam, M; Mohan, N; Sugadev, R; Chellappan, P; Mahadevan, A (2003). "(+) - katexinning Acinetobacter calcoaceticus MTC 127 tomonidan parchalanishi". Biochimica et Biofhysica Acta (BBA) - Umumiy mavzular. 1621 (3): 261–265. doi:10.1016 / S0304-4165 (03) 00077-1. PMID 12787923.
- ^ Hopper, Vaxeta; Mahadevan, A. (1997). "Katexinning Bradyrhizobium japonicum tomonidan parchalanishi". Biologik parchalanish. 8 (3): 159–165. doi:10.1023 / A: 1008254812074.
- ^ Sambandam, T .; Mahadevan, A. (1993). "Katexin parchalanishi va katetin oksigenazaning Caetomium cupreum-dan tozalanishi va qisman tavsifi". Butunjahon mikrobiologiya va biotexnologiya jurnali. 9 (1): 37–44. doi:10.1007 / BF00656513. PMID 24419836.
- ^ a b v d Ottaviani, J. I .; Borxes, G; Momma, T. Y .; Spenser, J. P .; Kin, K. L .; Krozye, A; Schroeter, H (2016). "Odamlarda 2- (14) C (-) - epikatexinning metabolomlari: Polifenol bioaktivlarning samaradorligi, xavfsizligi va ta'sir mexanizmlarini baholash uchun ta'siri". Ilmiy ma'ruzalar. 6: 29034. Bibcode:2016 yil NatSR ... 629034O. doi:10.1038 / srep29034. PMC 4929566. PMID 27363516.
- ^ Aktis-Goretta, L; Levek, A; Reyn, M; Teml, A; Schäfer, C; Xofmann, U; Li, H; Shvab, M; Eyxelbaum, M; Uilyamson, G (2013). "Sog'lom odamlarda ichakning emishi, metabolizmi va chiqarilishi (-) - epikatexin ichak perfuziyasi texnikasi yordamida baholanadi". Amerika Klinik Ovqatlanish Jurnali. 98 (4): 924–33. doi:10.3945 / ajcn.113.065789. PMID 23864538.
- ^ Ottaviani, J. I .; Momma, T. Y .; Khnle, G. K .; Kin, K. L .; Schroeter, H (2012). "Tuzilmaviy jihatdan bog'liq bo'lgan (-) - odamlarda epikatexin metabolitlari: de novo kimyoviy sintezlangan haqiqiy standartlardan foydalangan holda baholash". Bepul radikal biologiya va tibbiyot. 52 (8): 1403–12. doi:10.1016 / j.freeradbiomed.2011.12.010. PMID 22240152.
- ^ a b v "Flavonoidlar". Linus Poling instituti, Oregon shtat universiteti, Corvallis. 2016 yil. Olingan 24 iyul 2016.
- ^ Ottaviani, J. I .; Momma, T. Y .; Xeys, C; Kvik-Uribe, C; Shroeter, H; Keen, L. L. (2011). "Flavanollarning stereokimyoviy konfiguratsiyasi odamlarda flavanollarning darajasi va metabolizmiga va in vivo jonli ravishda ularning biologik faolligiga ta'sir qiladi". Bepul radikal biologiya va tibbiyot. 50 (2): 237–44. doi:10.1016 / j.freeradbiomed.2010.11.005. PMID 21074608.
- ^ Ye, J. H; Augustin, M. A (2018). "Katexinlarni etkazib berish uchun nano- va mikro zarrachalar: fizikaviy va biologik ko'rsatkichlar". Oziq-ovqat fanlari va ovqatlanish sohasidagi tanqidiy sharhlar. 59 (10): 1563–1579. doi:10.1080/10408398.2017.1422110. PMID 29345975.
- ^ Aprotosoaie, A. C; Miron, A; Trifan, A; Luka, V. S; Costache, I. I (2016). "Kakao polifenollarining yurak-qon tomirlariga ta'siri - umumiy nuqtai". Kasalliklar. 4 (4): 39. doi:10.3390 / kasalliklar4040039. PMC 5456324. PMID 28933419.
- ^ Guo, Y; Zhi, F; Chen, P; Chjao, K; Syan, H; Mao, Q; Vang, X; Chjan, X (2017). "Yashil choy va prostata bezi saratoni xavfi: tizimli tahlil va meta-tahlil". Dori. 96 (13): e6426. doi:10.1097 / MD.0000000000006426. PMC 5380255. PMID 28353571.
- ^ Matsuda M, Otsuka Y, Jin S va boshq. (2008 yil fevral). "(+) - katexinni ikki bosqichli oksidlanish orqali taksifolinga biotransformatsiyasi: (+) - katexin metabolizmining yangi bosqichi (+) - katechinni parchalaydigan bakteriyalar, Burkholderia sp. KTC-1, tropik torfdan ajratilgan" . Biokimyo. Biofiz. Res. Kommunal. 366 (2): 414–9. doi:10.1016 / j.bbrc.2007.11.157. PMID 18068670.
- ^ Shibuya H, Agusta A, Ohashi K, Maehara S, Simanjuntak P (iyul 2005). "Choy o'simliklaridan ajratilgan Diaporthe sp. Endofitik qo'ziqorin tomonidan (+) - katexin va (-) - epikatezinning 3,4-dihidroksiflavan hosilalariga biooksidlanishi". Kimyoviy. Farm. Buqa. 53 (7): 866–7. doi:10.1248 / cpb.53.866. PMID 15997157.
- ^ Bogs J, Downey MO, Harvey JS, Eshton AR, Tanner GJ, Robinson SP (oktyabr 2005). "Uzum mevalari va uzumzor barglarini ishlab chiqarishda leukoantotsianidin reduktaza va antosiyanidin reduktaza kodlovchi genlarning proantotsianidin sintezi va ekspresiyasi". O'simliklar fizioli. 139 (2): 652–63. doi:10.1104 / pp.105.064238. JSTOR 4281902. PMC 1255985. PMID 16169968.
- ^ Fridrix, Volfgang; Galensa, Rudolf (2002). "Arpa (Hordeum vulgare L.) va soloddan yangi flavanol glyukozidini aniqlash". Evropa oziq-ovqat tadqiqotlari va texnologiyasi. 214 (5): 388–393. doi:10.1007 / s00217-002-0498-x.
- ^ Jin QD, Mu QZ (1991). "[Epigynum auritum dan glikozidal tarkibiy qismlarni o'rganish]". Yao Xue Xue Bao (xitoy tilida). 26 (11): 841–5. PMID 1823978.
- ^ a b Vogiatzoglou, A; Mulligan, A. A .; Bhaniani, A; Lentjes, M. A .; Maktaggart, A; Lyuben, R. N .; Xeys, C; Kelm, M; Merx, M. V .; Spenser, J. P .; Shroeter, H; Xav, K. T .; Kuhl, G. G. (2015). "Evropaning saraton kasalligini (EPIC-Norfolk) tekshirish bo'yicha Norfolk kogortasida flavan-3-ol va KVH xavfi o'rtasidagi assotsiatsiyalar". Bepul radikal biologiya va tibbiyot. 84: 1–10. doi:10.1016 / j.freeradbiomed.2015.03.005. PMC 4503814. PMID 25795512.
- ^ Shreder, Yoxann (1655). Pharmacopoeia medico-chymica: sive thesaurus pharmacologeus. Ulmae Suevorum: Yoxannis Gerlini.
- ^ Berends, KAW (1829). Pathologie and Therapie qo'l amaliyoti Arzneiwissenschaft oder der speedziellen. Berlin: Enslin.
- ^ Armentano, L; Bentsat, A; Beres, T; Rusznyak, St; Szent-Györgyi, A (1936). "Über den Einfluß von Substanzen der Flavongruppe auf die Permeabilität der Kapillaren. Vitamin P". Deutsche Medizinische Wochenschrift. 62 (33): 1325–1328. doi:10.1055 / s-0028-1141260.
- ^ Hooper, L; Kay, C; Abdelhamid, A; Kroon, P. A .; Kon, J. S .; Rimm, E. B.; Kessidi, A (2012). "Shokolad, kakao va flavan-3-ollarning yurak-qon tomir sog'lig'iga ta'siri: randomizatsiyalangan tekshiruvlarning tizimli tekshiruvi va meta-tahlili". Amerika Klinik Ovqatlanish Jurnali. 95 (3): 740–51. doi:10.3945 / ajcn.111.023457. PMID 22301923.
- ^ Ellinger, S; Reusch, A; Stele, P; Helfrich, H. P. (2012). "Kakao mahsulotlari orqali qabul qilingan epikatexin odamlarda qon bosimini pasaytiradi: Bayes yondashuvi bilan chiziqli bo'lmagan regressiya modeli". Amerika Klinik Ovqatlanish Jurnali. 95 (6): 1365–77. doi:10.3945 / ajcn.111.029330. PMID 22552030.
- ^ a b Xelesi, S; Quyosh, J; Buys, N; Jamshidi, A; Nikbaxt-Nasrabadi, E; Xosravi-Borujeni, H (2014). "Yashil choy katexinlari va qon bosimi: randomizatsiyalangan nazorat ostida o'tkazilgan sinovlarni tizimli ko'rib chiqish va meta-tahlil". Ovqatlanish bo'yicha Evropa jurnali. 53 (6): 1299–311. doi:10.1007 / s00394-014-0720-1. PMID 24861099.
- ^ Shroeter, H; Xeys, C; Balzer, J; Klaynbongard, P; Kin, K. L .; Xollenberg, N. K .; Sies, H; Kvik-Uribe, C; Shmitz, H. H.; Kelm, M (2006). "(-) - Epikatexin flavanolga boy kakao moddalarining odamlarda qon tomir funktsiyasiga foydali ta'sirini anglatadi". Milliy fanlar akademiyasi materiallari. 103 (4): 1024–9. Bibcode:2006 yil PNAS..103.1024S. doi:10.1073 / pnas.0510168103. PMC 1327732. PMID 16418281.
- ^ Diyetik mahsulotlar, ovqatlanish va allergiya bo'yicha EFSA paneli (NDA) (2012). "1924/2006 yildagi (EC) Nizomning 13-moddasi 5-bandiga binoan kakao flavanollari va normal endoteliyga bog'liq vazodilatatsiyani ta'minlash bilan bog'liq sog'liqqa da'voni asoslash bo'yicha ilmiy fikr". EFSA jurnali. 10 (7): 2809. doi:10.2903 / j.efsa.2012.2809. Olingan 25 iyul 2016.
- ^ Martinez SE; Devies NM; Reynolds JK (2013). "Flavonoidlarning toksikologiyasi va xavfsizligi". Tahlil usullari, klinikgacha va klinik farmakokinetikasi, xavfsizligi va toksikologiyasi. John Wiley & Son. p. 257. ISBN 978-0-470-57871-1.
- ^ Bode, JC (1987). Okolicsányi, Lajos; Tsomos, Geza; Krepaldi, Gaetano (tahrir). Gepatobiliyer kasalliklarni baholash va boshqarish. Berlin: Springer-Verlag. p. 371. doi:10.1007/978-3-642-72631-6. ISBN 978-3-642-72631-6.
- ^ "Ruhen der Zulassung für Catergen" (PDF). Deutsches Ärzteblatt. 82 (38): 2706.
- ^ a b Amanda K. Broz va Xorxe M. Vivanko (2006). "O'simliklar invaziyasida ikkilamchi metabolitlar va allelopatiya: Centaurea maculosa holatini o'rganish". Sinauer Associates; O'simliklar fiziologiyasi va rivojlanishi, Ed. 6; Linkoln Taiz, Eduardo Zayger, Yan M. Myuller va Angus Merfi, muharrirlar.
- ^ Bais HP, Vepachedu R, Gilroy S, Callaway RM, Vivanco JM (sentyabr 2003). "Allelopatiya va ekzotik o'simlik bosqini: molekulalar va genlardan turlarning o'zaro ta'siriga qadar". Ilm-fan. 301 (5638): 1377–80. Bibcode:2003 yil ... 301.1377B. doi:10.1126 / science.1083245. PMID 12958360.
- ^ Yamamoto M, Nakatsuka S, Otani H, Kohmoto K, Nishimura S (iyun 2000). "(+) - katexin qulupnay bargida infektsiyani inhibe qiluvchi omil sifatida ishlaydi". Fitopatologiya. 90 (6): 595–600. doi:10.1094 / PHYTO.2000.90.6.595. PMID 18944538.
- ^ Chen Z, Liang J, Zhang C, Rodrigues CJ (2006 yil oktyabr). "Epikatexin va katexin Colletotrichum kahawae ning appressorli melanizatsiyasini inhibe qilish orqali kofe berry kasalligini oldini olishlari mumkin". Biotexnol. Lett. 28 (20): 1637–40. doi:10.1007 / s10529-006-9135-2. PMID 16955359.
Tashqi havolalar
 Bilan bog'liq ommaviy axborot vositalari (+) - katexin Vikimedia Commons-da
Bilan bog'liq ommaviy axborot vositalari (+) - katexin Vikimedia Commons-da